
Disulfiram Inhibits Opsonin-Independent Phagocytosis and Migration of Human Long-Lived In Vitro Cultured Phagocytes from Multiple Inflammatory Diseases


Abstract
1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. Cell Preparation
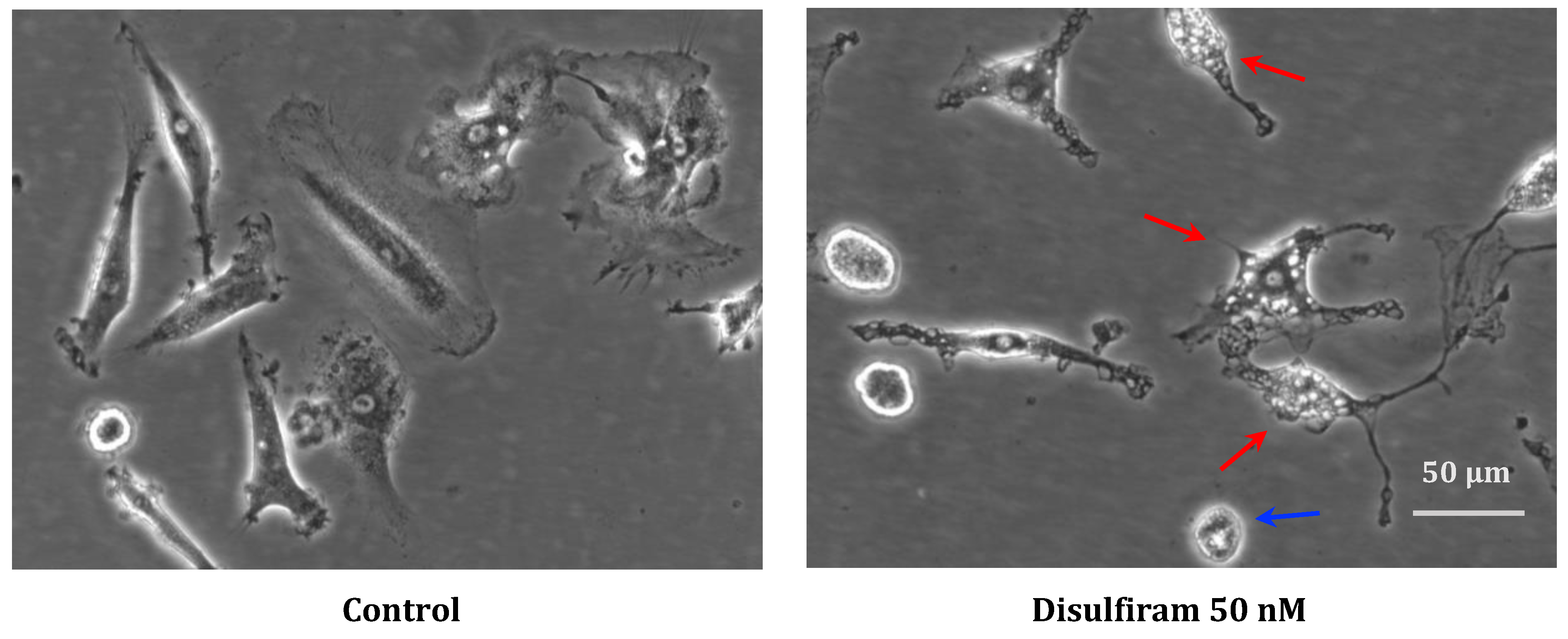
2.3. Macrophage Images Based on Phase–Contrast Microscopy
2.4. Cytospin Preparation and Wright–Giemsa Stain
2.5. Monitoring Phagocytosis by Video Microscopy
2.6. Metabolic Characterization Cultured Macrophages and Dendritic Cells Activated In Vivo
2.7. Flow Cytometry
2.8. Statistics
3. Results
3.1. Quantification of Functional Phenotypes Expressed by Cultured Macrophages
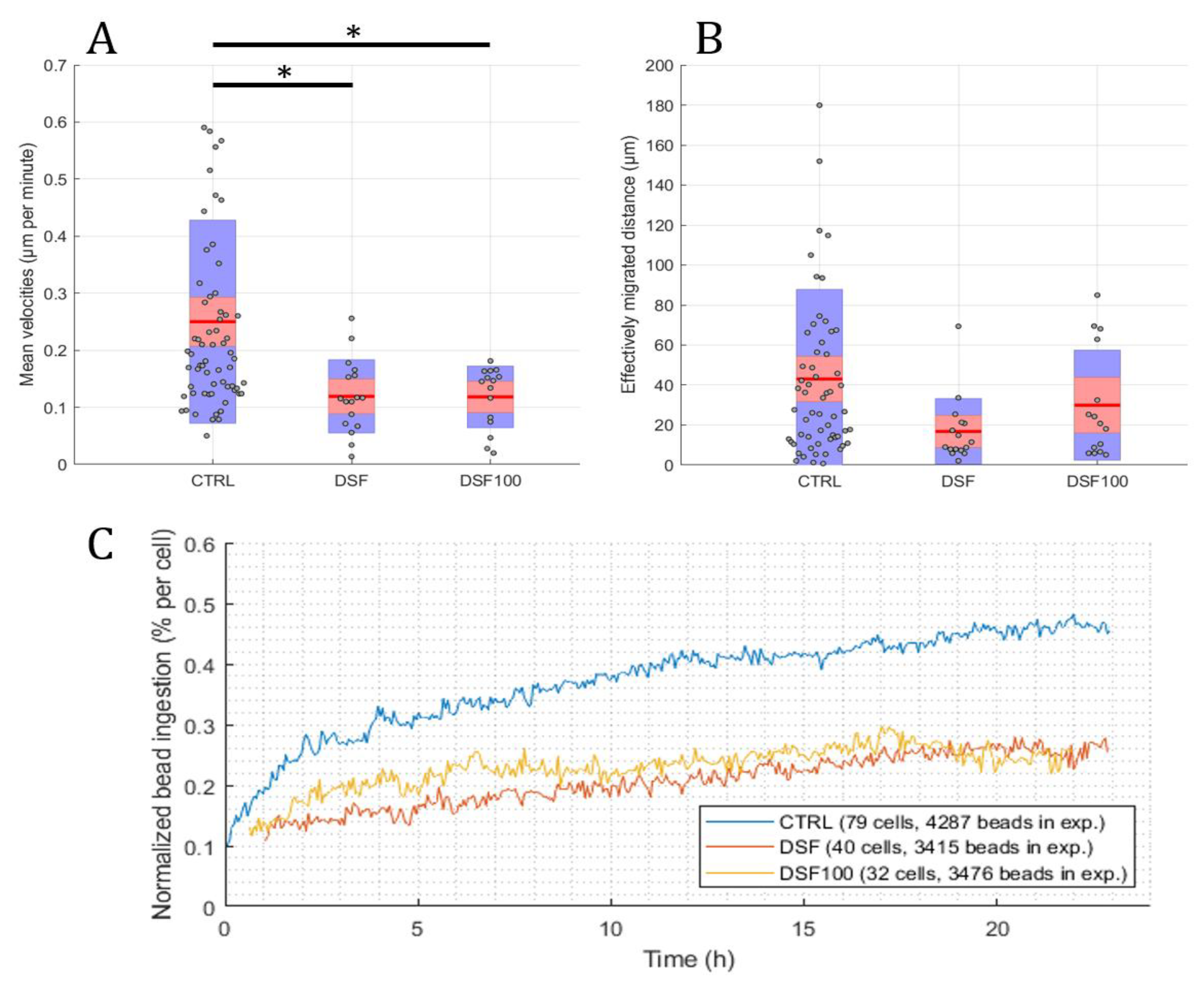
3.1.1. Phagocytosis
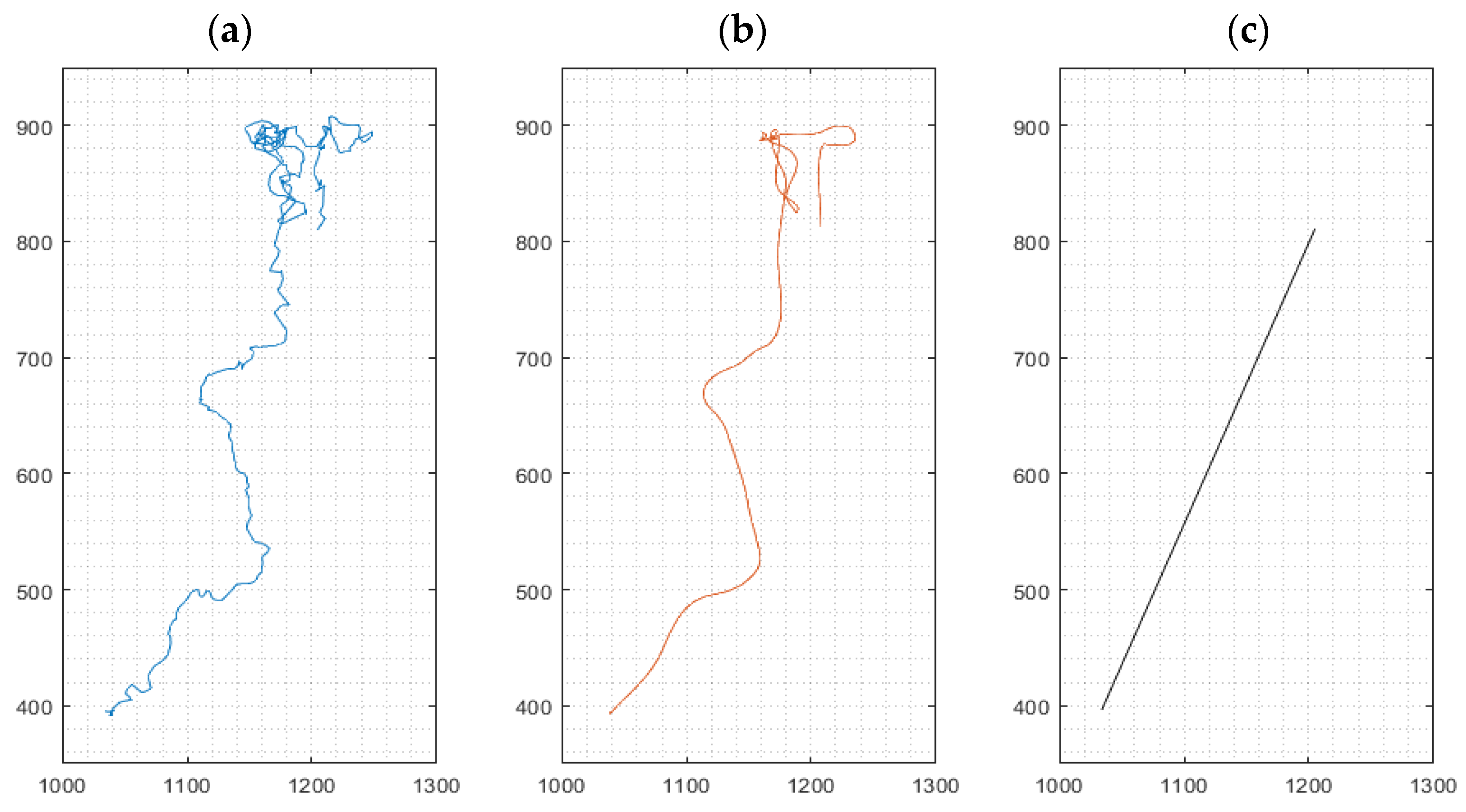
3.1.2. Cell Movement Analysis for Migration Distance Velocity
3.2. Identification of Macrophage Phenotypes
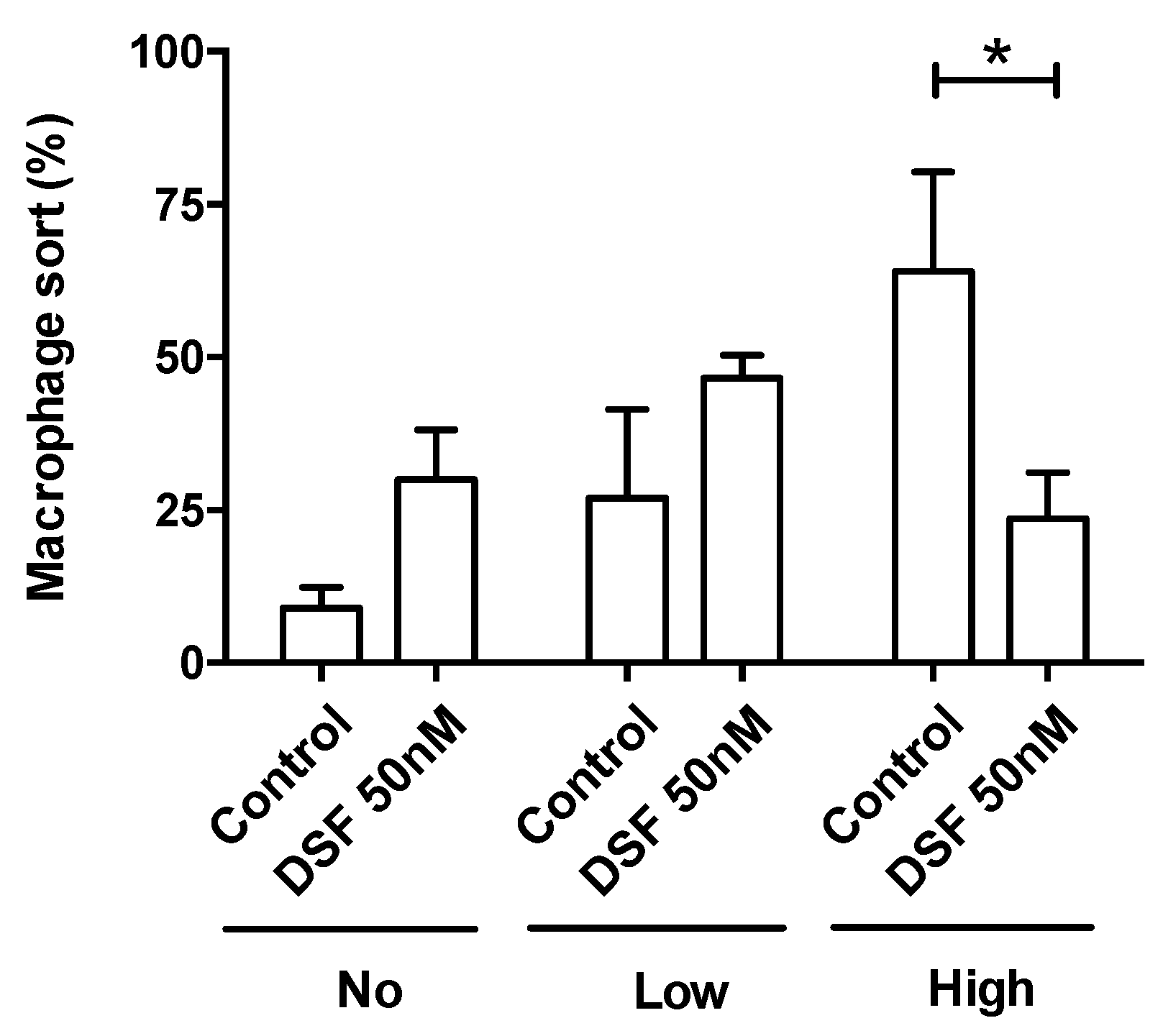
3.3. Modulation of Macrophage and Dendritic Cell Function and Viability by DSF
3.3.1. Correlation of Reduced Phagocytosis and Cell Death Induced by DSF
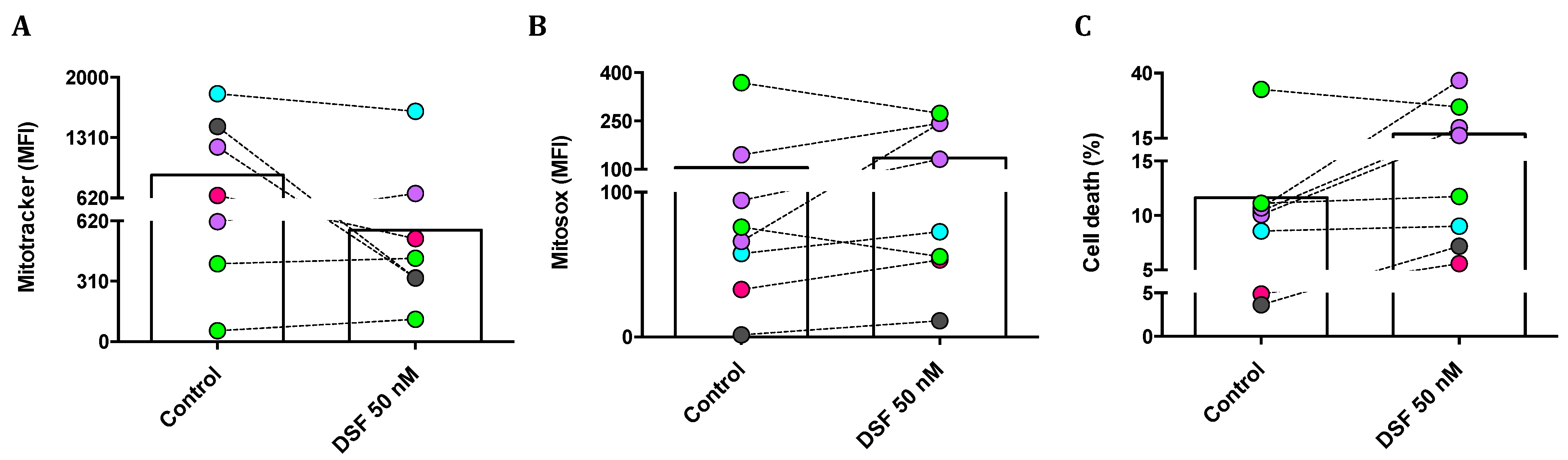
3.3.2. Correlation of DSF-Induced Mitochondrial ROS and Cell Death
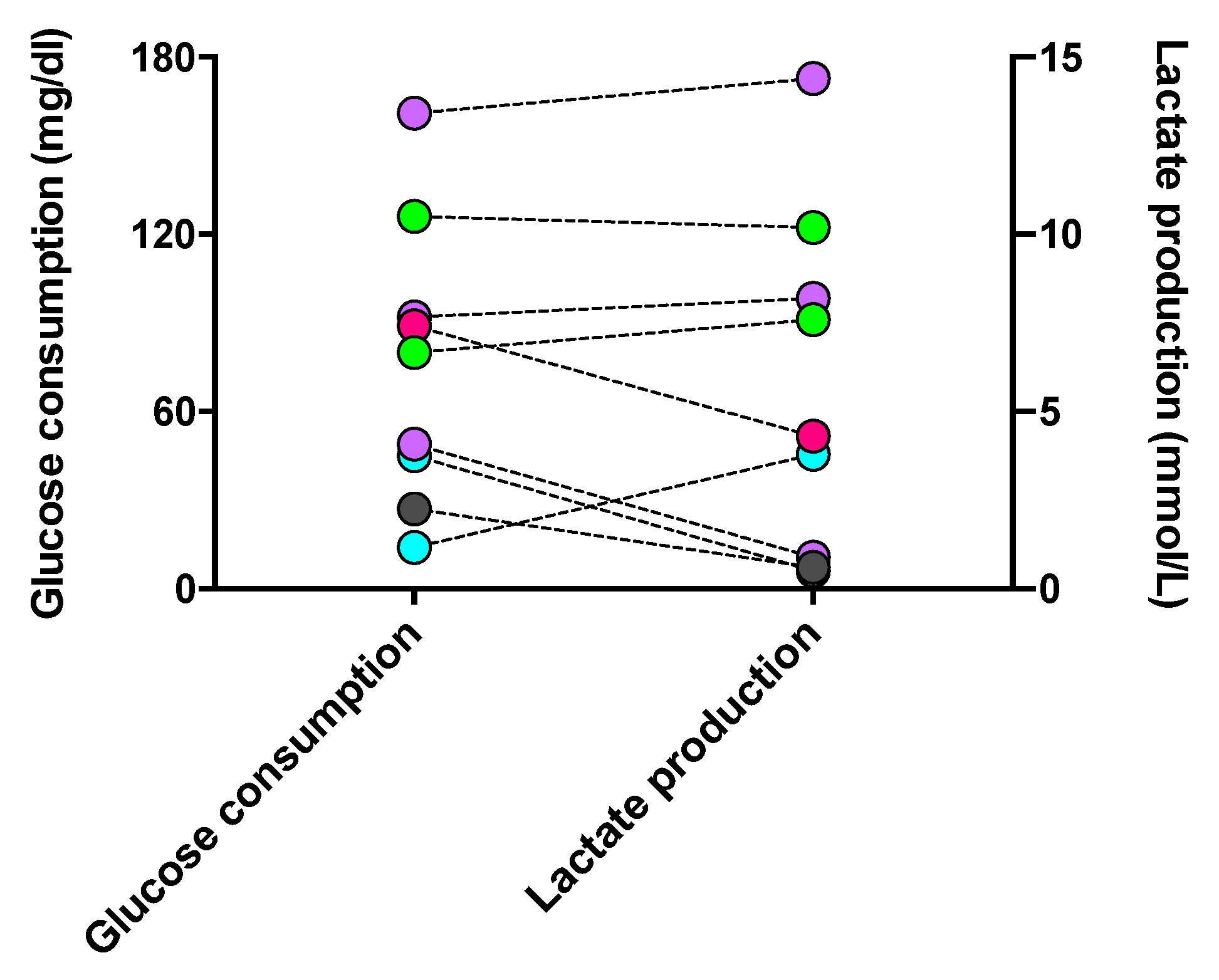
3.3.3. Correlation between Glycolytic Metabolism and Phagocyte Phenotype Regulated by DSF
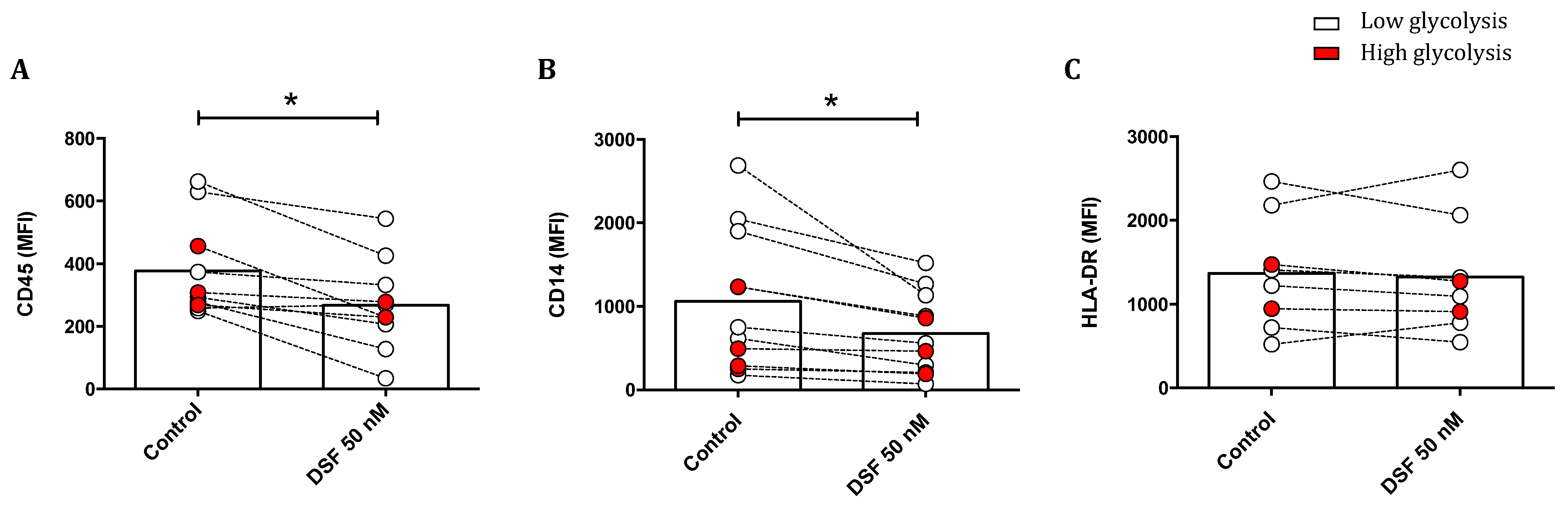
3.3.4. Modulation of Surface Antigen Expression by DSF
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hein, T.M.; Sander, P.; Giryes, A.; Reinhardt, J.O.; Hoegel, J.; Schneider, E.M. Cytokine Expression Patterns and Single Nucleotide Polymorphisms (SNPs) in Patients with Chronic Borreliosis. Antibiotics 2019, 8, 107. [Google Scholar] [CrossRef]
- Del Prete, A.; Salvi, V.; Soriani, A.; Laffranchi, M.; Sozio, F.; Bosisio, D.; Sozzani, S. Dendritic cell subsets in cancer immunity and tumor antigen sensing. Cell. Mol. Immunol. 2023, 20, 432–447. [Google Scholar] [CrossRef]
- Malyshev, I.; Malyshev, Y. Current Concept and Update of the Macrophage Plasticity Concept: Intracellular Mechanisms of Reprogramming and M3 Macrophage “Switch” Phenotype. Biomed. Res. Int. 2015, 2015, 341308. [Google Scholar] [CrossRef]
- Sica, A.; Mantovani, A. Macrophage plasticity and polarization: In vivo veritas. J. Clin. Investig. 2012, 122, 787–795. [Google Scholar] [CrossRef]
- Jablonski, K.A.; Amici, S.A.; Webb, L.M.; Ruiz-Rosado, J.d.D.; Popovich, P.G.; Partida-Sanchez, S.; Guerau-de-Arellano, M. Novel Markers to Delineate Murine M1 and M2 Macrophages. PLoS ONE 2015, 10, e0145342. [Google Scholar] [CrossRef]
- Dwivedi, S.; Rendón-Huerta, E.P.; Ortiz-Navarrete, V.; Montaño, L.F. CD38 and Regulation of the Immune Response Cells in Cancer. J. Oncol. 2021, 2021, 6630295. [Google Scholar] [CrossRef]
- Hu, J.M.; Liu, K.; Liu, J.H.; Jiang, X.L.; Wang, X.L.; Chen, Y.Z.; Li, S.G.; Zou, H.; Pang, L.J.; Liu, C.X.; et al. CD163 as a marker of M2 macrophage, contribute to predicte aggressiveness and prognosis of Kazakh esophageal squamous cell carcinoma. Oncotarget 2017, 8, 21526–21538. [Google Scholar] [CrossRef]
- Viola, A.; Munari, F.; Sánchez-Rodríguez, R.; Scolaro, T.; Castegna, A. The Metabolic Signature of Macrophage Responses. Front. Immunol. 2019, 10, 1462. [Google Scholar] [CrossRef]
- Silva, A.L.; Peres, C.; Conniot, J.; Matos, A.I.; Moura, L.; Carreira, B.; Sainz, V.; Scomparin, A.; Satchi-Fainaro, R.; Préat, V.; et al. Nanoparticle impact on innate immune cell pattern-recognition receptors and inflammasomes activation. Semin. Immunol. 2017, 34, 3–24. [Google Scholar] [CrossRef]
- Ehrchen, J.M.; Roth, J.; Barczyk-Kahlert, K. More Than Suppression: Glucocorticoid Action on Monocytes and Macrophages. Front. Immunol. 2019, 10, 2028. [Google Scholar] [CrossRef]
- Lu, C.; Li, X.; Ren, Y.; Zhang, X. Disulfiram: A novel repurposed drug for cancer therapy. Cancer Chemother. Pharmacol. 2021, 87, 159–172. [Google Scholar] [CrossRef]
- Najlah, M.; Suliman, A.S.; Tolaymat, I.; Kurusamy, S.; Kannappan, V.; Elhissi, A.M.A.; Wang, W. Development of Injectable PEGylated Liposome Encapsulating Disulfiram for Colorectal Cancer Treatment. Pharmaceutics 2019, 11, 610. [Google Scholar] [CrossRef]
- Parny, M.; Bernad, J.; Prat, M.; Salon, M.; Aubouy, A.; Bonnafé, E.; Coste, A.; Pipy, B.; Treilhou, M. Comparative study of the effects of ziram and disulfiram on human monocyte-derived macrophage functions and polarization: Involvement of zinc. Cell Biol. Toxicol. 2021, 37, 379–400. [Google Scholar] [CrossRef]
- Balakirev, M.Y.; Zimmer, G. Mitochondrial injury by disulfiram: Two different mechanisms of the mitochondrial permeability transition. Chem. Biol. Interact. 2001, 138, 299–311. [Google Scholar] [CrossRef]
- Lu, Y.; Cao, L.; Egami, Y.; Kawai, K.; Araki, N. Cofilin contributes to phagocytosis of IgG-opsonized particles but not non-opsonized particles in RAW264 macrophages. Microscopy 2016, 65, 233–242. [Google Scholar] [CrossRef]
- Aderem, A.; Underhill, D.M. Mechanisms of phagocytosis in macrophages. Annu. Rev. Immunol. 1999, 17, 593–623. [Google Scholar] [CrossRef]
- Palecanda, A.; Kobzik, L. Receptors for unopsonized particles: The role of alveolar macrophage scavenger receptors. Curr. Mol. Med. 2001, 1, 589–595. [Google Scholar] [CrossRef]
- Kobzik, L. Lung macrophage uptake of unopsonized environmental particulates. Role of scavenger-type receptors. J. Immunol. 1995, 155, 367–376. [Google Scholar] [CrossRef]
- Palecanda, A.; Paulauskis, J.; Al-Mutairi, E.; Imrich, A.; Qin, G.; Suzuki, H.; Kodama, T.; Tryggvason, K.; Koziel, H.; Kobzik, L. Role of the scavenger receptor MARCO in alveolar macrophage binding of unopsonized environmental particles. J. Exp. Med. 1999, 189, 1497–1506. [Google Scholar] [CrossRef]
- Huth, J.; Buchholz, M.; Kraus, J.M.; Schmucker, M.; von Wichert, G.; Krndija, D.; Seufferlein, T.; Gress, T.M.; A Kestler, H. Significantly improved precision of cell migration analysis in time-lapse video microscopy through use of a fully automated tracking system. BMC Cell Biol. 2010, 11, 24. [Google Scholar] [CrossRef]
- Norling, M.; Karlsson-Lindsjö, O.E.; Gourlé, H.; Bongcam-Rudloff, E.; Hayer, J. MetLab: An In Silico Experimental Design, Simulation and Analysis Tool for Viral Metagenomics Studies. PLoS ONE 2016, 11, e0160334. [Google Scholar] [CrossRef]
- Grout, M.M.; Mitchell, K.B. Disulfiram-Mitigating Unintended Effects. Antibiotics 2023, 12, 262. [Google Scholar] [CrossRef]
- Gordon, P.; Okai, B.; Hoare, J.I.; Erwig, L.P.; Wilson, H.M. SOCS3 is a modulator of human macrophage phagocytosis. J. Leukoc. Biol. 2016, 100, 771–780. [Google Scholar] [CrossRef] [PubMed]
- Patente, T.A.; Pinho, M.P.; Oliveira, A.A.; Evangelista, G.C.M.; Bergami-Santos, P.C.; Barbuto, J.A.M. Human Dendritic Cells: Their Heterogeneity and Clinical Application Potential in Cancer Immunotherapy. Front. Immunol. 2018, 9, 3176. [Google Scholar] [CrossRef]
- Kis-Toth, K.; Hajdu, P.; Bacskai, I.; Szilagyi, O.; Papp, F.; Szanto, A.; Posta, E.; Gogolak, P.; Panyi, G.; Rajnavolgyi, E. Voltage-gated sodium channel Nav1.7 maintains the membrane potential and regulates the activation and chemokine-induced migration of a monocyte-derived dendritic cell subset. J. Immunol. 2011, 187, 1273–1280. [Google Scholar] [CrossRef]
- Comi, M.; Avancini, D.; de Sio, F.S.; Villa, M.; Uyeda, M.J.; Floris, M.; Tomasoni, D.; Bulfone, A.; Roncarolo, M.G.; Gregori, S. Coexpression of CD163 and CD141 identifies human circulating IL-10-producing dendritic cells (DC-10). Cell. Mol. Immunol. 2020, 17, 95–107. [Google Scholar] [CrossRef]
- Mantovani, A.; Sica, A.; Sozzani, S.; Allavena, P.; Vecchi, A.; Locati, M. The chemokine system in diverse forms of macrophage activation and polarization. Trends Immunol. 2004, 25, 677–686. [Google Scholar] [CrossRef]
- Zizzo, G.; Hilliard, B.A.; Monestier, M.; Cohen, P.L. Efficient clearance of early apoptotic cells by human macrophages requires M2c polarization and MerTK induction. J. Immunol. 2012, 189, 3508–3520. [Google Scholar] [CrossRef]
- Jiao, Y.; Hannafon, B.N.; Ding, W.Q. Disulfiram’s Anticancer Activity: Evidence and Mechanisms. Anticancer Agents Med. Chem. 2016, 16, 1378–1384. [Google Scholar] [CrossRef]
- Lin, H.; Sun, F.; Li, T.; Zhang, Y.; Guo, X.; Li, M.; Liang, M.; Zhou, X.; Fang, Z. Disulfiram enhances chemotherapeutic effects of doxorubicin liposomes against human hepatocellular carcinoma via activating ROS-induced cell stress response pathways. Cancer Chemother. Pharmacol. 2022, 90, 455–465. [Google Scholar] [CrossRef]
- Moerke, C.; Mueller, P.; Nebe, B. Attempted caveolae-mediated phagocytosis of surface-fixed micro-pillars by human osteoblasts. Biomaterials 2016, 76, 102–114. [Google Scholar] [CrossRef]
- Mateo, C.; Grazu, V.; Guisan, J.M. Immobilization of enzymes on monofunctional and heterofunctional epoxy-activated supports. Methods Mol. Biol. 2013, 1051, 43–57. [Google Scholar] [PubMed]
- Gu, B.J.; Duce, J.A.; Valova, V.A.; Wong, B.; Bush, A.I.; Petrou, S.; Wiley, J.S. P2X7 receptor-mediated scavenger activity of mononuclear phagocytes toward non-opsonized particles and apoptotic cells is inhibited by serum glycoproteins but remains active in cerebrospinal fluid. J. Biol. Chem. 2012, 287, 17318–17330. [Google Scholar] [CrossRef] [PubMed]
- Gauthier, J.D.; Feig, B.; Vasta, G.R. Effect of fetal bovine serum glycoproteins on the in vitro proliferation of the oyster parasite Perkinsus marinus: Development of a fully defined medium. J. Eukaryot. Microbiol. 1995, 42, 307–313. [Google Scholar] [CrossRef] [PubMed]
- Kholmukhamedov, A.; Schwartz, J.M.; Lemasters, J.J. Isolated mitochondria infusion mitigates ischemia-reperfusion injury of the liver in rats: Mitotracker probes and mitochondrial membrane potential. Shock 2013, 39, 543. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Zhang, B.; Tao, Y.; Wang, Y.; Wei, H.; Zhao, J.; Huang, R.; Pei, Z. DL-3-n-butylphthalide protects endothelial cells against oxidative/nitrosative stress, mitochondrial damage and subsequent cell death after oxygen glucose deprivation in vitro. Brain Res. 2009, 1290, 91–101. [Google Scholar] [CrossRef]
- Lo, H.M.; Chen, C.L.; Yang, C.M.; Wu, P.H.; Tsou, C.J.; Chiang, K.W.; Wu, W.B. The carotenoid lutein enhances matrix metalloproteinase-9 production and phagocytosis through intracellular ROS generation and ERK1/2, p38 MAPK, and RARβ activation in murine macrophages. J. Leukoc. Biol. 2013, 93, 723–735. [Google Scholar] [CrossRef]
- Lai, J.C.Y.; Wlodarska, M.; Liu, D.J.; Abraham, N.; Johnson, P. CD45 regulates migration, proliferation, and progression of double negative 1 thymocytes. J. Immunol. 2010, 185, 2059–2070. [Google Scholar] [CrossRef]
- Saunders, A.E.; Johnson, P. Modulation of immune cell signalling by the leukocyte common tyrosine phosphatase, CD45. Cell. Signal. 2010, 22, 339–348. [Google Scholar] [CrossRef]
- Zwingmann, C.; Leibfritz, D.; Hazell, A.S. Energy metabolism in astrocytes and neurons treated with manganese: Relation among cell-specific energy failure, glucose metabolism, and intercellular trafficking using multinuclear NMR-spectroscopic analysis. J. Cereb. Blood Flow Metab. 2003, 23, 756–771. [Google Scholar] [CrossRef]
- Kummer, L.; Zaradzki, M.; Vijayan, V.; Arif, R.; Weigand, M.A.; Immenschuh, S.; Wagner, A.H.; Larmann, J. Vascular Signaling in Allogenic Solid Organ Transplantation—The Role of Endothelial Cells. Front. Physiol. 2020, 11, 443. [Google Scholar] [CrossRef] [PubMed]
- Thibodeau, J.; Moulefera, M.A.; Balthazard, R. On the structure-function of MHC class II molecules and how single amino acid polymorphisms could alter intracellular trafficking. Hum. Immunol. 2019, 80, 15–31. [Google Scholar] [CrossRef] [PubMed]
- Crux, N.B.; Elahi, S. Human Leukocyte Antigen (HLA) and Immune Regulation: How Do Classical and Non-Classical HLA Alleles Modulate Immune Response to Human Immunodeficiency Virus and Hepatitis C Virus Infections? Front. Immunol. 2017, 8, 832. [Google Scholar] [CrossRef]
- Vaglini, F.; Viaggi, C.; Piro, V.; Pardini, C.; Gerace, C.; Scarselli, M.; Corsini, G.U. Acetaldehyde and parkinsonism: Role of CYP450 2E1. Front. Behav. Neurosci. 2013, 7, 71. [Google Scholar] [CrossRef] [PubMed]
- Boscá, L.; González-Ramos, S.; Prieto, P.; Fernández-Velasco, M.; Mojena, M.; Martín-Sanz, P.; Alemany, S. Metabolic signatures linked to macrophage polarization: From glucose metabolism to oxidative phosphorylation. Biochem. Soc. Trans. 2015, 43, 740–744. [Google Scholar] [CrossRef] [PubMed]
- Orihuela, R.; McPherson, C.A.; Harry, G.J. Microglial M1/M2 polarization and metabolic states. Br. J. Pharmacol. 2016, 173, 649–665. [Google Scholar] [CrossRef] [PubMed]
- Mills, E.L.; O’Neill, L.A. Reprogramming mitochondrial metabolism in macrophages as an anti-inflammatory signal. Eur. J. Immunol. 2016, 46, 13–21. [Google Scholar] [CrossRef]
- Liu, Y.; Xu, R.; Gu, H.; Zhang, E.; Qu, J.; Cao, W.; Huang, X.; Yan, H.; He, J.; Cai, Z. Metabolic reprogramming in macrophage responses. Biomark. Res. 2021, 9, 1. [Google Scholar] [CrossRef]
- Lin, M.-H.; Moses, D.C.; Hsieh, C.-H.; Cheng, S.-C.; Chen, Y.-H.; Sun, C.-Y.; Chou, C.-Y. Disulfiram can inhibit MERS and SARS coronavirus papain-like proteases via different modes. Antivir. Res. 2018, 150, 155–163. [Google Scholar] [CrossRef]
- Fu, L.; Ye, F.; Feng, Y.; Yu, F.; Wang, Q.; Wu, Y.; Zhao, C.; Sun, H.; Huang, B.; Niu, P.; et al. Both Boceprevir and GC376 efficaciously inhibit SARS-CoV-2 by targeting its main protease. Nat. Commun. 2020, 11, 4417. [Google Scholar] [CrossRef]
- Froggatt, H.M.; Heaton, B.E.; Heaton, N.S. Development of a Fluorescence-Based, High-Throughput SARS-CoV-2 3CL(pro) Reporter Assay. J. Virol. 2020, 94, e01265-20. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.J.; Liu, X.; Xia, S.; Zhang, Z.; Zhang, Y.; Zhao, J.; Ruan, J.; Luo, X.; Lou, X.; Bai, Y.; et al. FDA-approved disulfiram inhibits pyroptosis by blocking gasdermin D pore formation. Nat. Immunol. 2020, 21, 736–745. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.; Fei, C.Y.; Chen, Y.P.; Sargsyan, K.; Chang, C.P.; Yuan, H.S.; Lim, C. Synergistic Inhibition of SARS-CoV-2 Replication Using Disulfiram/Ebselen and Remdesivir. ACS Pharmacol. Transl. Sci. 2021, 4, 898–907. [Google Scholar] [CrossRef] [PubMed]
- Potula, H.S.K.; Shahryari, J.; Inayathullah, M.; Malkovskiy, A.V.; Kim, K.-M.; Rajadas, J. Repurposing Disulfiram (Tetraethylthiuram Disulfide) as a Potential Drug Candidate against Borrelia burgdorferi In Vitro and In Vivo. Antibiotics 2020, 9, 633. [Google Scholar] [CrossRef] [PubMed]
- Troxell, B.; Ye, M.; Yang, Y.; Carrasco, S.E.; Lou, Y.; Yang, X.F. Manganese and zinc regulate virulence determinants in Borrelia burgdorferi. Infect. Immun. 2013, 81, 2743–2752. [Google Scholar] [CrossRef] [PubMed]
- Lanz, J.; Biniaz-Harris, N.; Kuvaldina, M.; Jain, S.; Lewis, K.; Fallon, B.A. Disulfiram: Mechanisms, Applications, and Challenges. Antibiotics 2023, 12, 524. [Google Scholar] [CrossRef]
- Grage-Griebenow, E.; Flad, H.-D.; Ernst, M. Heterogeneity of human peripheral blood monocyte subsets. J. Leukoc. Biol. 2001, 69, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Coillard, A.; Segura, E. In vivo Differentiation of Human Monocytes. Front. Immunol. 2019, 10, 1907. [Google Scholar] [CrossRef]
- del Hoyo, G.M.; Martín, P.; Vargas, H.H.; Ruiz, S.; Arias, C.F.; Ardavín, C. Characterization of a common precursor population for dendritic cells. Nature 2002, 415, 1043–1047. [Google Scholar] [CrossRef]
- Gabrusiewicz, K.; Rodriguez, B.; Wei, J.; Hashimoto, Y.; Healy, L.M.; Maiti, S.N.; Thomas, G.; Zhou, S.; Wang, Q.; Elakkad, A.; et al. Glioblastoma-infiltrated innate immune cells resemble M0 macrophage phenotype. JCI Insight 2016, 1, e85841. [Google Scholar] [CrossRef]
- Tremble, L.F.; Forde, P.F.; Soden, D.M. Clinical evaluation of macrophages in cancer: Role in treatment, modulation and challenges. Cancer Immunol. Immunother. 2017, 66, 1509–1527. [Google Scholar] [CrossRef]
- Li, W.; Li, Y.; Jin, X.; Liao, Q.; Chen, Z.; Peng, H.; Zhou, Y. CD38: A Significant Regulator of Macrophage Function. Front. Oncol. 2022, 12, 775649. [Google Scholar] [CrossRef]
- Malavasi, F.; Deaglio, S.; Funaro, A.; Ferrero, E.; Horenstein, A.L.; Ortolan, E.; Vaisitti, T.; Aydin, S. Evolution and function of the ADP ribosyl cyclase/CD38 gene family in physiology and pathology. Physiol. Rev. 2008, 88, 841–886. [Google Scholar] [CrossRef] [PubMed]
- Zilber, M.-T.; Gregory, S.; Mallone, R.; Deaglio, S.; Malavasi, F.; Charron, D.; Gelin, C. CD38 expressed on human monocytes: A coaccessory molecule in the superantigen-induced proliferation. Proc. Natl. Acad. Sci. USA 2000, 97, 2840–2845. [Google Scholar] [CrossRef] [PubMed]
- Buechler, C.; Ritter, M.; Orsó, E.; Langmann, T.; Klucken, J.; Schmitz, G. Regulation of scavenger receptor CD163 expression in human monocytes and macrophages by pro- and antiinflammatory stimuli. J. Leukoc. Biol. 2000, 67, 97–103. [Google Scholar] [CrossRef] [PubMed]
- Tippett, E.; Cheng, W.-J.; Westhorpe, C.; Cameron, P.U.; Brew, B.J.; Lewin, S.R.; Jaworowski, A.; Crowe, S.M. Differential expression of CD163 on monocyte subsets in healthy and HIV-1 infected individuals. PLoS ONE 2011, 6, e19968. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Diagnosis | Age | Gender | Phenotype | Exosome * | |
|---|---|---|---|---|---|
| 20231 | Xanthogranuloma | 43 | M | 80% cDC, 20% M1 | 21% |
| 20315 | Glioblastoma °IV | 73 | M | iDC | 21% |
| 20331 | Glioblastoma °IV | 66 | F | cDC | 16% |
| 20365 | Pancreatic cancer | 55 | M | M2a | 26% |
| 20373 | COVID-19 infection after stem cell transplantation | 18 | M | M2a | 10% |
| 20643 | Healthy donor | 33 | M | M1 | 5% |
| 20650 | Motor neuron disease | 58 | M | 25% M1 + 75% iDC | 1% |
| 20667 | Glioblastoma °IV | 40 | M | 50% M1 + 50% iDC | 2% |
| 20705 | Long-COVID | 20 | F | M2a | 3% |
| 20725 | Borreliosis after DSF treatment | 48 | M | M2a | 2% |
| 21161 | Major depressive disorder | 30 | F | cDC | 10% |
| 21162 | Long-COVID | 32 | M | 50% M1 + 50% cDC | 4% |
| 21163 | Chronic fatigue syndrome | 44 | M | cDC | 17% |
| 21164 | Post VAC syndrome | 43 | F | M2a | 22% |
| 21428 | Prostate cancer | 63 | M | DC-10 | 7% |
| PatNo. |
CD14 [%postives] |
CD206 [%postives] |
CD163 [%postives] |
CD38 [%postives] |
CD141 [%postives] |
Kv1.3 [%postives] |
|---|---|---|---|---|---|---|
| 20231 * | 20 | 2 | 3 | 6 | 6 | 7 |
| 20315 * | 30 | 4 | 0.4 | 1 | 2 | 5 |
| 20331 | 83 | 5 | 1 | 1 | 2 | 27 |
| 20365 | 89 | 1 | 1 | 4 | nt | 7 |
| 20373 | 93 | 32 | 65 | 6 | nt | 76 |
| 20643 | 89 | 3 | 2 | 85 | nt | nt |
| 20650 | 88 | 0.3 | 0.2 | 25 | nt | 10 |
| 20667 | 98 | 3 | 2 | 52 | nt | 17 |
| 20705 | 99 | nt | 61 | 48 | nt | 78 |
| 20725 | 97 | 4 | 2 | 20 | nt | 53 |
| 21161 | 86 | 10 | 1 | 18 | nt | nt |
| 21162 | 98 | nt | 4 | 50 | nt | nt |
| 21163 | 63 | nt | 1 | 4 | nt | nt |
| 21164 | 78 | nt | 1 | 2 | nt | nt |
| 21428 | 90 | 54 | 70 | nt | 90 | 23 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, C.; Schneider, J.M.; Schneider, E.M. Disulfiram Inhibits Opsonin-Independent Phagocytosis and Migration of Human Long-Lived In Vitro Cultured Phagocytes from Multiple Inflammatory Diseases. Cells 2024, 13, 535. https://doi.org/10.3390/cells13060535
Li C, Schneider JM, Schneider EM. Disulfiram Inhibits Opsonin-Independent Phagocytosis and Migration of Human Long-Lived In Vitro Cultured Phagocytes from Multiple Inflammatory Diseases. Cells. 2024; 13(6):535. https://doi.org/10.3390/cells13060535
Chicago/Turabian StyleLi, Chen, Julian M. Schneider, and E. Marion Schneider. 2024. "Disulfiram Inhibits Opsonin-Independent Phagocytosis and Migration of Human Long-Lived In Vitro Cultured Phagocytes from Multiple Inflammatory Diseases" Cells 13, no. 6: 535. https://doi.org/10.3390/cells13060535
APA StyleLi, C., Schneider, J. M., & Schneider, E. M. (2024). Disulfiram Inhibits Opsonin-Independent Phagocytosis and Migration of Human Long-Lived In Vitro Cultured Phagocytes from Multiple Inflammatory Diseases. Cells, 13(6), 535. https://doi.org/10.3390/cells13060535