

Senescence and Stress Signaling Pathways in Corneal Cells After Nitrogen Mustard Injury

 , , , , , , , ,
, , , , , , , ,  , and
, and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. NM Preparation
2.3. Cells
2.4. NM Exposure
2.5. Western Blot
2.6. Real-Time PCR
2.7. Immunofluorescence Study
2.8. Proliferative Capacity and Senescence Regulation
2.9. MAPK Signaling Evaluation
2.10. β-Gal Staining Assessment
2.11. Statistical Analysis
3. Results
3.1. Morphological Changes in Human Corneal Cells
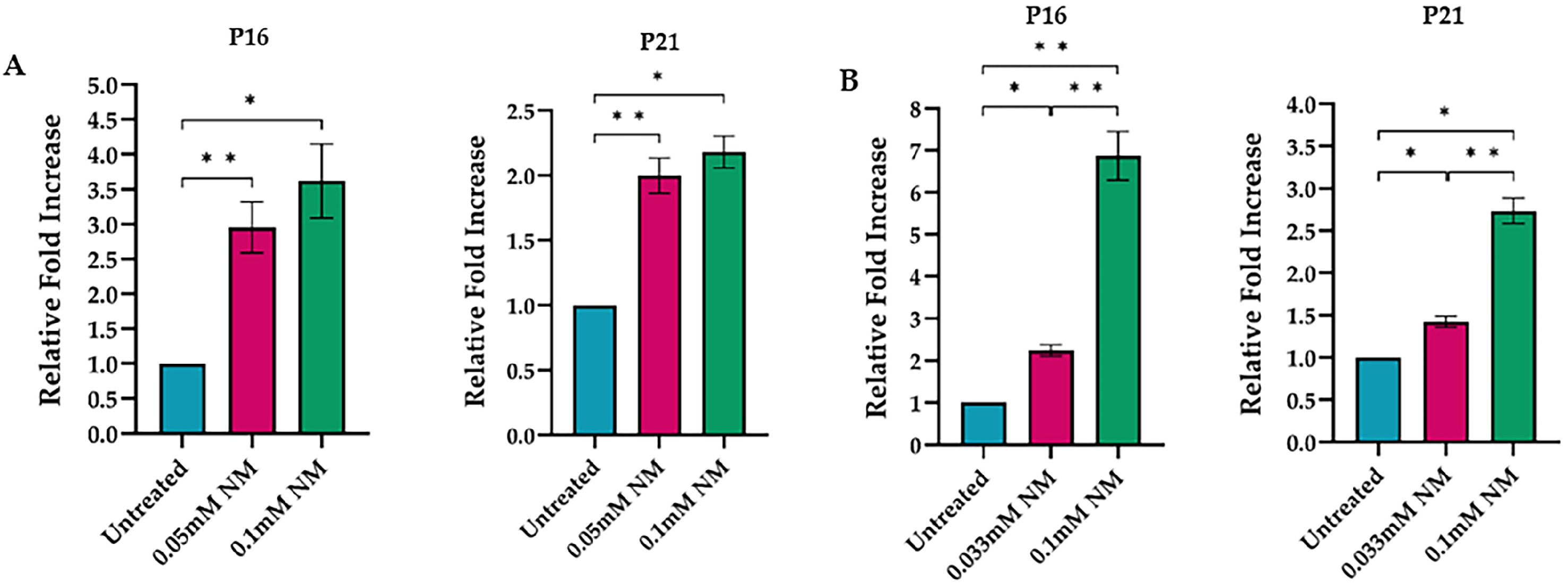
3.2. Proliferation-Related Gene Expression
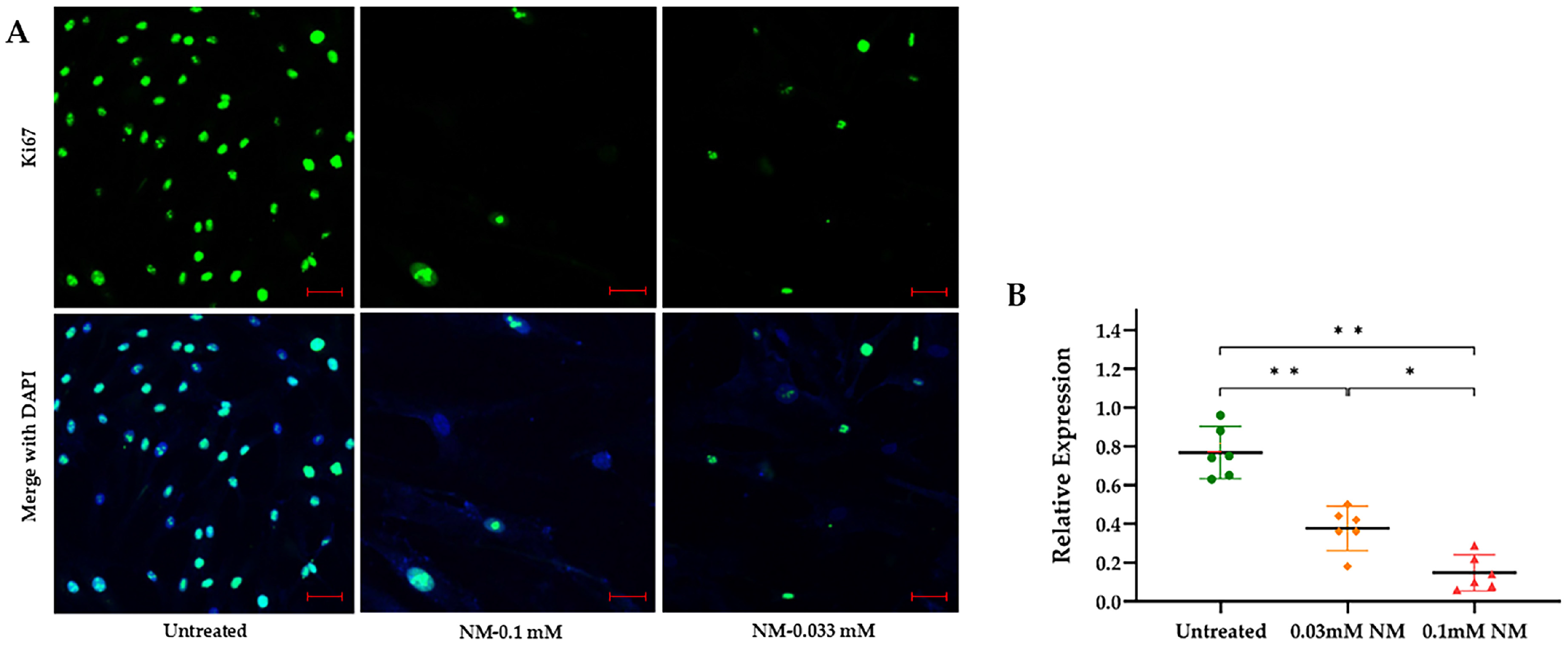
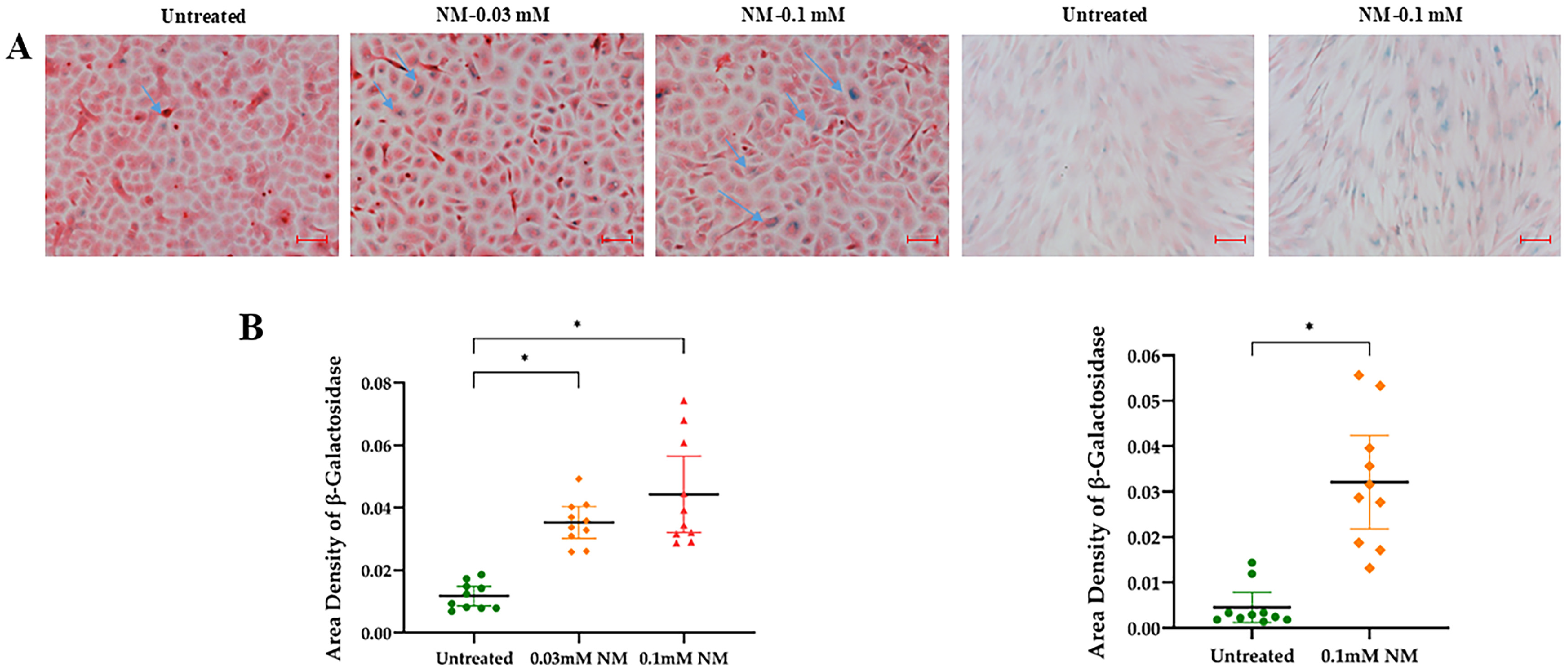
3.3. Activation of Senescence Markers
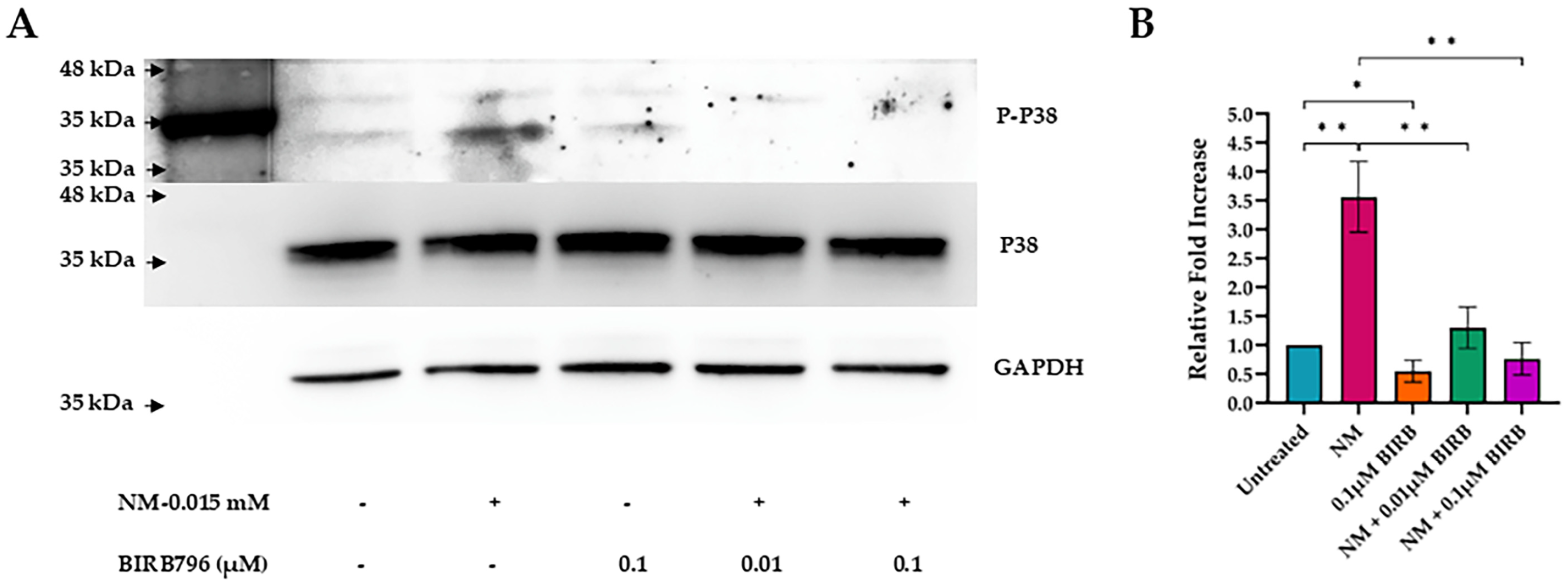
3.4. Activation of MAPK Pathways
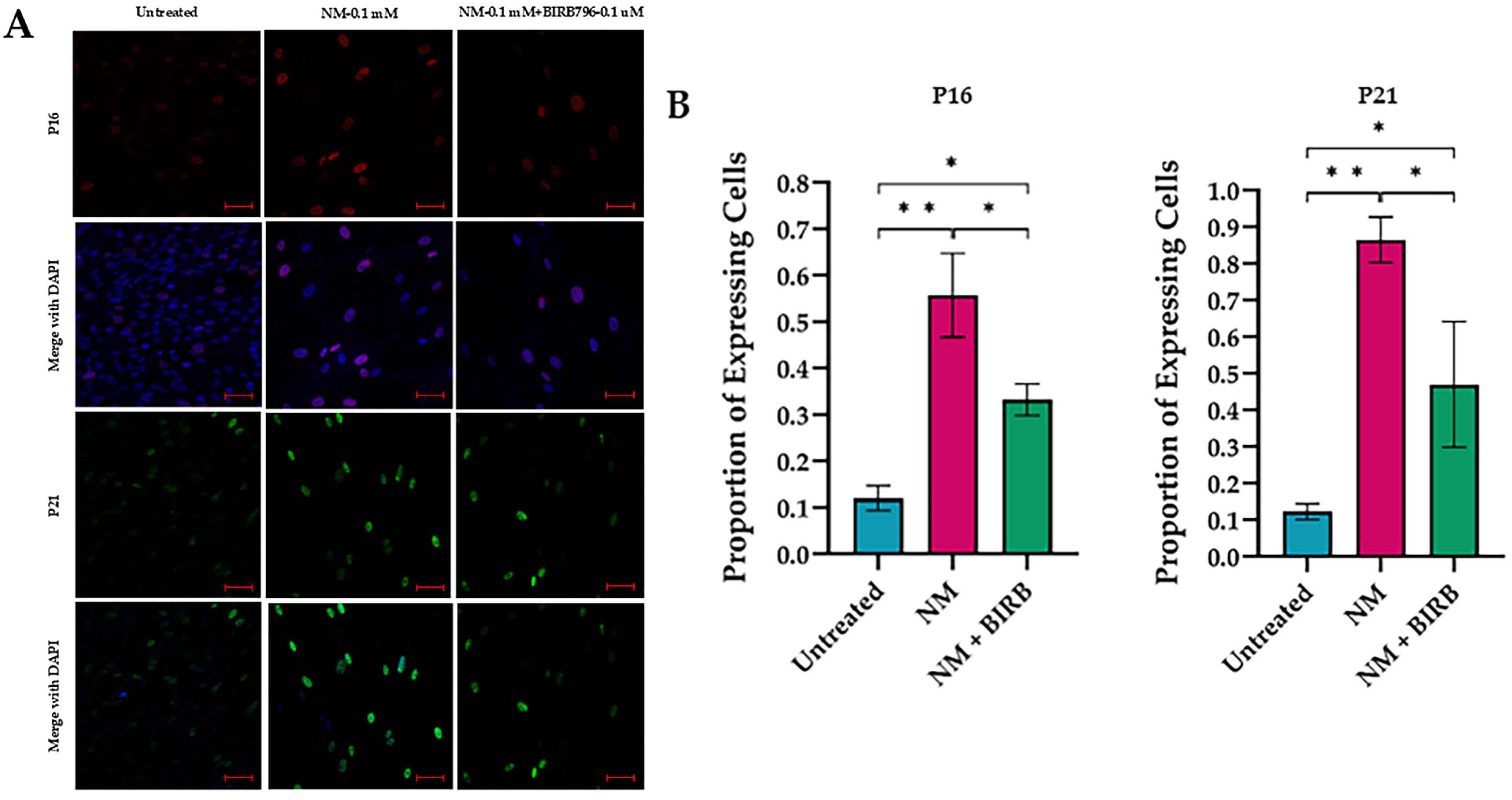
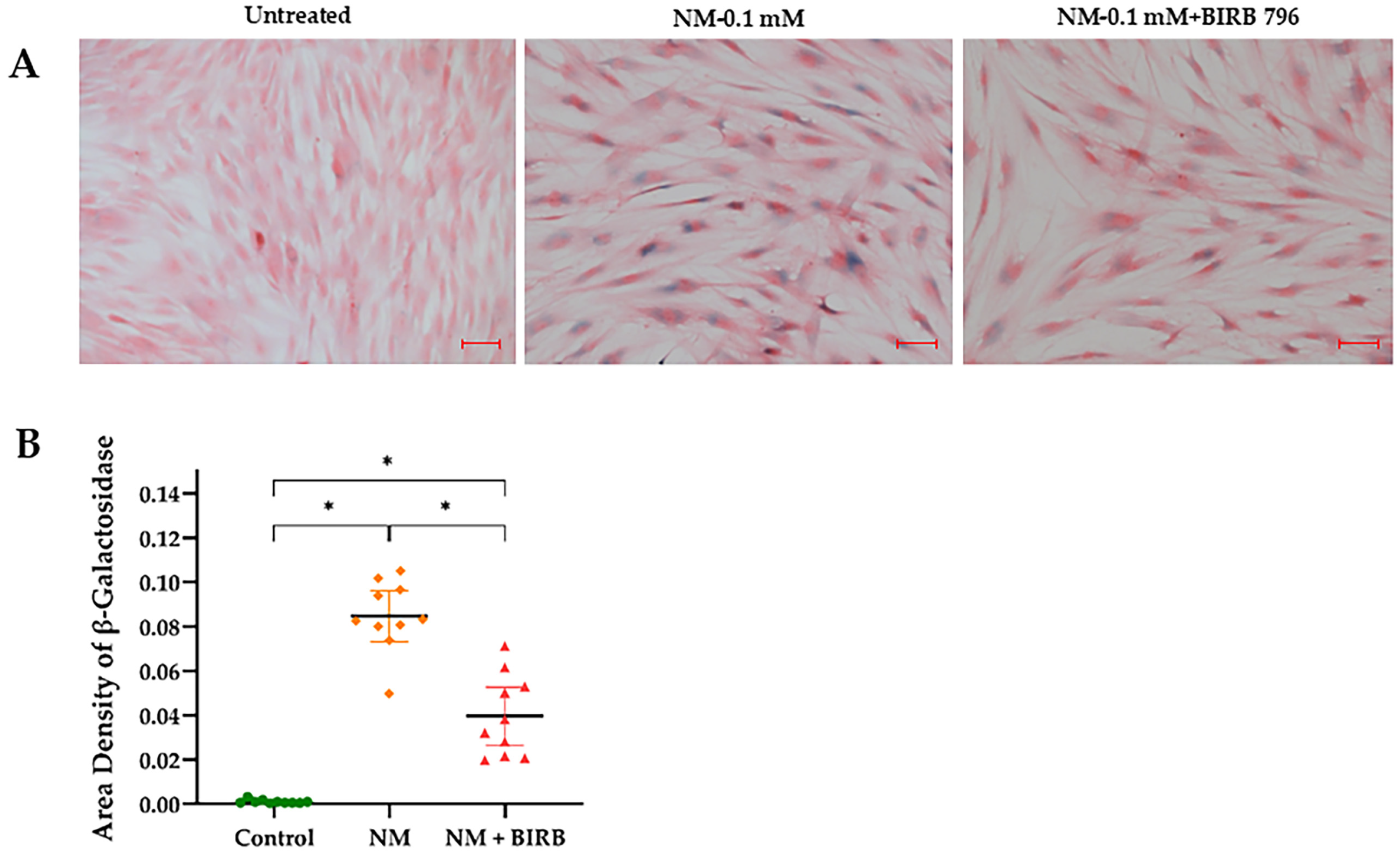
3.5. MAPK Inhibition
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Soleimani, M.; Momenaei, B.; Baradaran-Rafii, A.; Cheraqpour, K.; An, S.; Ashraf, M.J.; Abedi, F.; Javadi, M.A.; Djalilian, A.R. Mustard Gas-Induced Ocular Surface Disorders: An Update on the Pathogenesis, Clinical Manifestations, and Management. Cornea 2023, 42, 776–786. [Google Scholar] [CrossRef] [PubMed]
- Mishra, N.; Kant, R.; Kandhari, K.; Ammar, D.A.; Tewari-Singh, N.; Pantcheva, M.B.; Petrash, J.M.; Agarwal, C.; Agarwal, R. Nitrogen Mustard-Induced Ex Vivo Human Cornea Injury Model and Therapeutic Intervention by Dexamethasone. J. Pharmacol. Exp. Ther. 2024, 388, 484–494. [Google Scholar] [CrossRef] [PubMed]
- McNutt, P. Progress towards a standardized model of ocular sulfur mustard injury for therapeutic testing. Exp. Eye Res. 2023, 228, 109395. [Google Scholar] [CrossRef] [PubMed]
- Alemi, H.; Dehghani, S.; Musayeva, A.; Nadari, A.; Narimatsu, A.; Sharifi, S.; Forouzanfar, K.; Wang, S.; Dohlman, T.H.; Yin, J.; et al. Insights into mustard gas keratopathy: Characterizing corneal layer-specific changes in mice exposed to nitrogen mustard. Exp. Eye Res. 2023, 236, 109495. [Google Scholar] [CrossRef] [PubMed]
- Shahri, S.H.G.; Balali-Mood, M.; Heidarzadeh, H.R.; Abrishami, M. Ophthalmic Complications and Managements of Sulfur Mustard Exposure: A Narrative Review. Arch. Iran. Med. 2022, 25, 647–657. [Google Scholar] [CrossRef]
- Mahaling, B.; Sinha, N.R.; Sokupa, S.; Addi, U.R.; Mohan, R.R.; Chaurasia, S.S. Mustard gas exposure instigates retinal Müller cell gliosis. Exp. Eye Res. 2023, 230, 109461. [Google Scholar] [CrossRef]
- Poudel, S.; Kaffash, E.; Zhao, L.; Pangeni, R.; Chow, W.N.; Xu, Q. Dexamethasone sodium phosphate loaded nanoparticles for prevention of nitrogen mustard induced corneal injury. Exp. Eye Res. 2024, 243, 109902. [Google Scholar] [CrossRef]
- Amini, H.; Solaymani-Dodaran, M.; Mousavi, B.; Alam Beladi, S.N.; Soroush, M.R.; Abolghasemi, J.; Vahedian-Azimi, A.; Salesi, M.; Guest, P.C.; Sahebkar, A.; et al. Long-term Health Outcomes Among Survivors Exposed to Sulfur Mustard in Iran. JAMA Netw. Open 2020, 3, e2028894. [Google Scholar] [CrossRef]
- Darchini-Maragheh, E.; Balali-Mood, M. Delayed Complications and Long-term Management of Sulfur Mustard Poisoning: Recent Advances by Iranian Researchers (Part I of II). Iran. J. Med. Sci. 2018, 43, 103–124. [Google Scholar]
- Fuchs, A.; Giuliano, E.A.; Sinha, N.R.; Mohan, R.R. Ocular toxicity of mustard gas: A concise review. Toxicol. Lett. 2021, 343, 21–27. [Google Scholar] [CrossRef]
- Sinha, N.R.; Tripathi, R.; Balne, P.K.; Suleiman, L.; Simkins, K.; Chaurasia, S.S.; Mohan, R.R. Mustard Gas Exposure Actuates SMAD2/3 Signaling to Promote Myofibroblast Generation in the Cornea. Cells 2023, 12, 1533. [Google Scholar] [CrossRef]
- Rajavi, Z.; Safi, S.; Javadi, M.; Jafarinasab, M.; Feizi, S.; Moghadam, M.; Jadidi, K.; Babaei, M.; Shirvani, A.; Baradaran-Rafii, A.; et al. Clinical Practice Guidelines for Prevention, Diagnosis and Management of Early and Delayed-onset Ocular Injuries Due to Mustard Gas Exposure. J. Ophthalmic Vis. Res. 2017, 12, 65. [Google Scholar] [CrossRef] [PubMed]
- Zhylkibayev, A.; Ung, T.T.; Mobley, J.; Athar, M.; Gorbatyuk, M. The Involvement of Unfolded Protein Response in the Mechanism of Nitrogen Mustard-Induced Ocular Toxicity. J. Pharmacol. Exp. Ther. 2024, 388, 518–525. [Google Scholar] [CrossRef] [PubMed]
- Shalwitz, R.; Day, T.; Ruehlmann, A.K.; Julio, L.; Gordon, S.; Vandeuren, A.; Nelson, M.; Lyman, M.; Kelly, K.; Altvater, A.; et al. Treatment of Sulfur Mustard Corneal Injury by Augmenting the DNA Damage Response (DDR): A Novel Approach. J. Pharmacol. Exp. Ther. 2024, 388, 526–535. [Google Scholar] [CrossRef] [PubMed]
- Sprogyte, L.; Park, M.; Di Girolamo, N. Pathogenesis of Alkali Injury-Induced Limbal Stem Cell Deficiency: A Literature Survey of Animal Models. Cells 2023, 12, 1294. [Google Scholar] [CrossRef]
- Cheng, X.; Liu, C.; Yang, Y.; Liang, L.; Chen, B.; Yu, H.; Xia, J.; Liu, S.; Li, Y. Advances in sulfur mustard-induced DNA adducts: Characterization and detection. Toxicol. Lett. 2021, 344, 46–57. [Google Scholar] [CrossRef]
- Ramos, E.; Gil-Martín, E.; De Los Ríos, C.; Egea, J.; López-Muñoz, F.; Pita, R.; Juberías, A.; Torrado, J.J.; Serrano, D.R.; Reiter, R.J.; et al. Melatonin as Modulator for Sulfur and Nitrogen Mustard-Induced Inflammation, Oxidative Stress and DNA Damage: Molecular Therapeutics. Antioxidants 2023, 12, 397. [Google Scholar] [CrossRef]
- Zhao, Z.; Yan, X.; Li, L.; Shu, Y.; He, J.; Wang, L.; Huang, Q.; Xie, J.; Zhao, J.; Peng, S. Proliferating Stem Cells are Acutely Affected by DNA Damage Induced by Sulfur Mustard. DNA Cell Biol. 2022, 41, 716–726. [Google Scholar] [CrossRef]
- Jan, Y.H.; Heck, D.E.; Laskin, D.L.; Laskin, J.D. DNA damage signaling in the cellular responses to mustard vesicants. Toxicol. Lett. 2020, 326, 78–82. [Google Scholar] [CrossRef]
- Mishra, N.; Agarwal, R. Research models of sulfur mustard- and nitrogen mustard-induced ocular injuries and potential therapeutics. Exp. Eye Res. 2022, 223, 109209. [Google Scholar] [CrossRef]
- Horn, G.; Schäfers, C.; Thiermann, H.; Völkl, S.; Schmidt, A.; Rothmiller, S. Sulfur mustard single-dose exposure triggers senescence in primary human dermal fibroblasts. Arch. Toxicol. 2022, 96, 3053–3066. [Google Scholar] [CrossRef] [PubMed]
- Soleimani, M.; Cheraqpour, K.; Koganti, R.; Djalilian, A.R. Cellular senescence and ophthalmic diseases: Narrative review. Graefe’s Arch. Clin. Exp. Ophthalmol. 2023, 261, 3067–3082. [Google Scholar] [CrossRef]
- Rothmiller, S.; Jäger, N.; Meier, N.; Meyer, T.; Neu, A.; Steinritz, D.; Thiermann, H.; Scherer, M.; Rummel, C.; Mangerich, A.; et al. Chronic senescent human mesenchymal stem cells as possible contributor to the wound healing disorder after exposure to the alkylating agent sulfur mustard. Arch. Toxicol. 2021, 95, 727–747. [Google Scholar] [CrossRef] [PubMed]
- Soleimani, M.; Baharnoori, S.M.; Cheraqpour, K.; Momenaei, B.; Mirshahi, R.; Chow, C.; Shahjahan, S.; Nguyen, T.; Ashraf, M.J.; Huang, X.; et al. Cellular senescence implication in mustard keratopathy. Exp. Eye Res. 2023, 233, 109565. [Google Scholar] [CrossRef] [PubMed]
- Lopes-Paciencia, S.; Saint-Germain, E.; Rowell, M.C.; Ruiz, A.F.; Kalegari, P.; Ferbeyre, G. The senescence-associated secretory phenotype and its regulation. Cytokine 2019, 117, 15–22. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Han, J.; Elisseeff, J.H.; Demaria, M. The senescence-associated secretory phenotype and its physiological and pathological implications. Nat. Rev. Mol. Cell Biol. 2024, 25, 958–978. [Google Scholar] [CrossRef]
- Wilkinson, H.N.; Hardman, M.J. Cellular Senescence in Acute and Chronic Wound Repair. Cold Spring Harb. Perspect. Biol. 2022, 14, a041221. [Google Scholar] [CrossRef]
- Anerillas, C.; Abdelmohsen, K.; Gorospe, M. Regulation of senescence traits by MAPKs. GeroScience 2020, 42, 397–408. [Google Scholar] [CrossRef]
- Yue, J.; López, J.M. Understanding MAPK Signaling Pathways in Apoptosis. Int. J. Mol. Sci. 2020, 21, 2346. [Google Scholar] [CrossRef]
- Anerillas, C.; Altés, G.; Gorospe, M. MAPKs in the early steps of senescence implemEMTation. Front. Cell Dev. Biol. 2023, 11, 1083401. [Google Scholar] [CrossRef]
- Jabbehdari, S.; Yazdanpanah, G.; Kanu, L.N.; Chen, E.; Kang, K.; Anwar, K.N.; Ghassemi, M.; Hematti, P.; Rosenblatt, M.I.; Djalilian, A.R. Therapeutic Effects of Lyophilized Conditioned-Medium Derived from Corneal Mesenchymal Stromal Cells on Corneal Epithelial Wound Healing. Curr. Eye Res. 2020, 45, 1490–1496. [Google Scholar] [CrossRef] [PubMed]
- Eslani, M.; Putra, I.; Shen, X.; Hamouie, J.; Tadepalli, A.; Anwar, K.N.; Kink, J.A.; Ghassemi, S.; Agnihotri, G.; Reshetylo, S.; et al. Cornea-Derived Mesenchymal Stromal Cells Therapeutically Modulate Macrophage Immunophenotype and Angiogenic Function. Stem Cells 2018, 36, 775–784. [Google Scholar] [CrossRef] [PubMed]
- Jabbehdari, S.; Yazdanpanah, G.; Kanu, L.N.; Anwar, K.N.; Shen, X.; Rabiee, B.; Putra, I.; Eslani, M.; Rosenblatt, M.I.; Hematti, P.; et al. Reproducible Derivation and Expansion of Corneal Mesenchymal Stromal Cells for Therapeutic Applications. Transl. Vis. Sci. Technol. 2020, 9, 26. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Lee, H.J.; Shrivastava, A.; Wang, R.; McQuiston, T.J.; Challberg, S.S.; Pollok, B.A.; Wang, T. Long-Term In Vitro Expansion of Epithelial Stem Cells Enabled by Pharmacological Inhibition of PAK1-ROCK-Myosin II and TGF-β Signaling. Cell Rep. 2018, 25, 598–610.e595. [Google Scholar] [CrossRef]
- Gipson, I.K.; Spurr-Michaud, S.; Argüeso, P.; Tisdale, A.; Ng, T.F.; Russo, C.L. Mucin gene expression in immortalized human corneal-limbal and conjunctival epithelial cell lines. Investig. Ophthalmol. Vis. Sci. 2003, 44, 2496–2506. [Google Scholar] [CrossRef]
- Soleimani, M.; Mirzaei, A.; Cheraqpour, K.; Baharnoori, S.M.; Arabpour, Z.; Ashraf, M.J.; Ghassemi, M.; Djalilian, A.R. The Potential of Mesenchymal Stem/Stromal Cell Therapy in Mustard Keratopathy: Discovering New Roads to Combat Cellular Senescence. Cells 2023, 12, 2744. [Google Scholar] [CrossRef]
- Rabiee, B.; Anwar, K.N.; Shen, X.; Putra, I.; Liu, M.; Jung, R.; Afsharkhamseh, N.; Rosenblatt, M.I.; Fishman, G.A.; Liu, X.; et al. Gene dosage manipulation alleviates manifestations of hereditary PAX6 haploinsufficiency in mice. Sci. Transl. Med. 2020, 12, eaaz4894. [Google Scholar] [CrossRef]
- Putra, I.; Rabiee, B.; Anwar, K.N.; Gidfar, S.; Shen, X.; Babalooee, M.; Ghassemi, M.; Afsharkhamseh, N.; Bakhsh, S.; Missiakas, D.; et al. Staphylococcus aureus alpha-hemolysin impairs corneal epithelial wound healing and promotes intracellular bacterial invasion. Experiment. Eye Res. 2019, 181, 263–270. [Google Scholar] [CrossRef]
- Lambert, W.S.; Pasini, S.; Collyer, J.W.; Formichella, C.R.; Ghose, P.; Carlson, B.J.; Calkins, D.J. Of Mice and Monkeys: Neuroprotective Efficacy of the p38 Inhibitor BIRB 796 Depends on Model Duration in Experimental Glaucoma. Sci. Rep. 2020, 10, 1–20. [Google Scholar] [CrossRef]
- González-Gualda, E.; Baker, A.G.; Fruk, L.; Muñoz-Espín, D. A guide to assessing cellular senescence in vitro and in vivo. FEBS J. 2021, 288, 56–80. [Google Scholar] [CrossRef]
- Hou, X.; Shi, J.; Sun, L.; Song, L.; Zhao, W.; Xiong, X.; Lu, Y. The involvement of ERK1/2 and p38 MAPK in the premature senescence of melanocytes induced by H2O2 through a p53-independent p21 pathway. J. Dermatol. Sci. 2022, 105, 88–97. [Google Scholar] [CrossRef] [PubMed]
- An, S.; Shen, X.; Anwar, K.; Ashraf, M.; Lee, H.; Koganti, R.; Djalilian, A.R. Therapeutic Potential of Mesenchymal Stem Cell-Secreted Factors on Delay in Corneal Wound Healing by Nitrogen Mustard. Int. J. Mol. Sci. 2022, 23, 11510. [Google Scholar] [CrossRef] [PubMed]
- Behboudi, H.; Noureini, S.K.; Ghazanfari, T.; Ardestani, S.K. DNA damage and telomere length shortening in the peripheral blood leukocytes of 20 years SM-exposed veterans. Int. Immunopharmacol. 2018, 61, 37–44. [Google Scholar] [CrossRef] [PubMed]
- Goswami, D.G.; Tewari-Singh, N.; Dhar, D.; Kumar, D.; Agarwal, C.; Ammar, D.A.; Agarwal, R. Nitrogen Mustard-Induced Corneal Injury Involves DNA Damage and Pathways Related to Inflammation, Epithelial-Stromal Separation, and Neovascularization. Cornea 2016, 35, 257–266. [Google Scholar] [CrossRef]
- Kaluzhny, Y.; Kinuthia, M.W.; Lapointe, A.M.; Truong, T.; Klausner, M.; Hayden, P. Oxidative stress in corneal injuries of different origin: Utilization of 3D human corneal epithelial tissue model. Experiment. Eye Res. 2020, 190, 107867. [Google Scholar] [CrossRef]
- Eveleth, D.D.; Eveleth, J.J.; Subramaniam, A.; Hahn, R.; Zhou, P.; Gordon, M.K.; Bradshaw, R.A. An Engineered Human Fibroblast Growth Factor-1 Derivative, TTHX1114, Ameliorates Short-term Corneal Nitrogen Mustard Injury in Rabbit Organ Cultures. Investig. Ophthalmol. Vis. Sci. 2018, 59, 4720–4730. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Anwar, K.N.; Soleimani, M.; Ashraf, M.J.; Moghtader, A.; Koganti, R.; Ghalibafan, S.; Baharnoori, M.; Arabpour, Z.; Cheraqpour, K.; Sebhat, A.M.; et al. Senescence and Stress Signaling Pathways in Corneal Cells After Nitrogen Mustard Injury. Cells 2024, 13, 2021. https://doi.org/10.3390/cells13232021
Anwar KN, Soleimani M, Ashraf MJ, Moghtader A, Koganti R, Ghalibafan S, Baharnoori M, Arabpour Z, Cheraqpour K, Sebhat AM, et al. Senescence and Stress Signaling Pathways in Corneal Cells After Nitrogen Mustard Injury. Cells. 2024; 13(23):2021. https://doi.org/10.3390/cells13232021
Chicago/Turabian StyleAnwar, Khandaker N., Mohammad Soleimani, Mohammad Javad Ashraf, Amirhossein Moghtader, Raghuram Koganti, Seyyedehfatemeh Ghalibafan, Mahbod Baharnoori, Zohreh Arabpour, Kasra Cheraqpour, Aron M. Sebhat, and et al. 2024. "Senescence and Stress Signaling Pathways in Corneal Cells After Nitrogen Mustard Injury" Cells 13, no. 23: 2021. https://doi.org/10.3390/cells13232021
APA StyleAnwar, K. N., Soleimani, M., Ashraf, M. J., Moghtader, A., Koganti, R., Ghalibafan, S., Baharnoori, M., Arabpour, Z., Cheraqpour, K., Sebhat, A. M., Abtahi, M., Yao, X., Ghassemi, M., & Djalilian, A. R. (2024). Senescence and Stress Signaling Pathways in Corneal Cells After Nitrogen Mustard Injury. Cells, 13(23), 2021. https://doi.org/10.3390/cells13232021