
Effects of Garlic on Breast Tumor Cells with a Triple Negative Phenotype: Peculiar Subtype-Dependent Down-Modulation of Akt Signaling

 ,
,  , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Cells and Treatments
2.2. Apoptosis and Cell Cycle Analysis
2.3. Immunochemical Analysis
2.4. Immunocytochemical Analysis
2.5. Real-Time Cell Migration and Invasion Assays
2.6. Statistical Analysis
3. Results
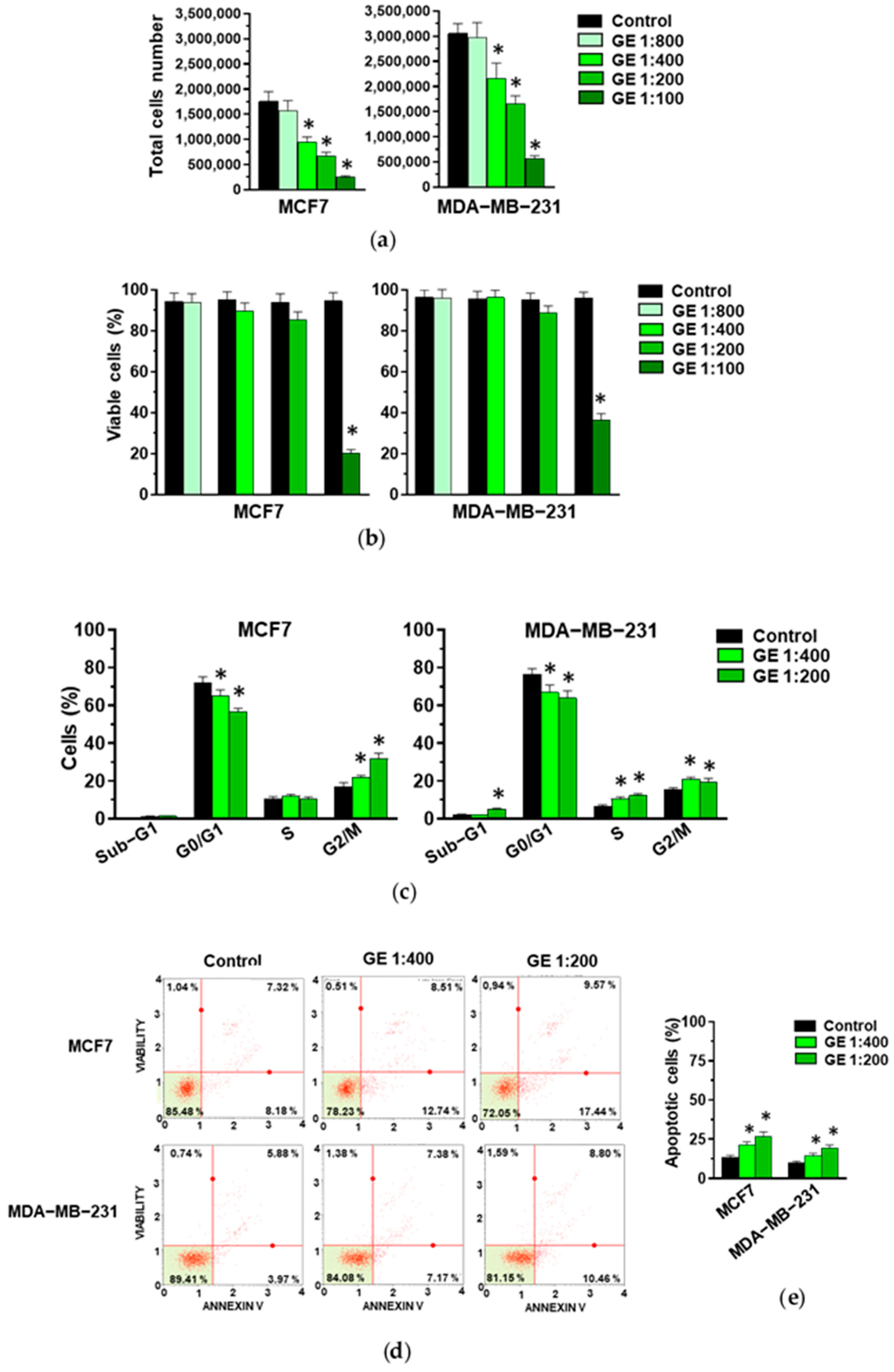
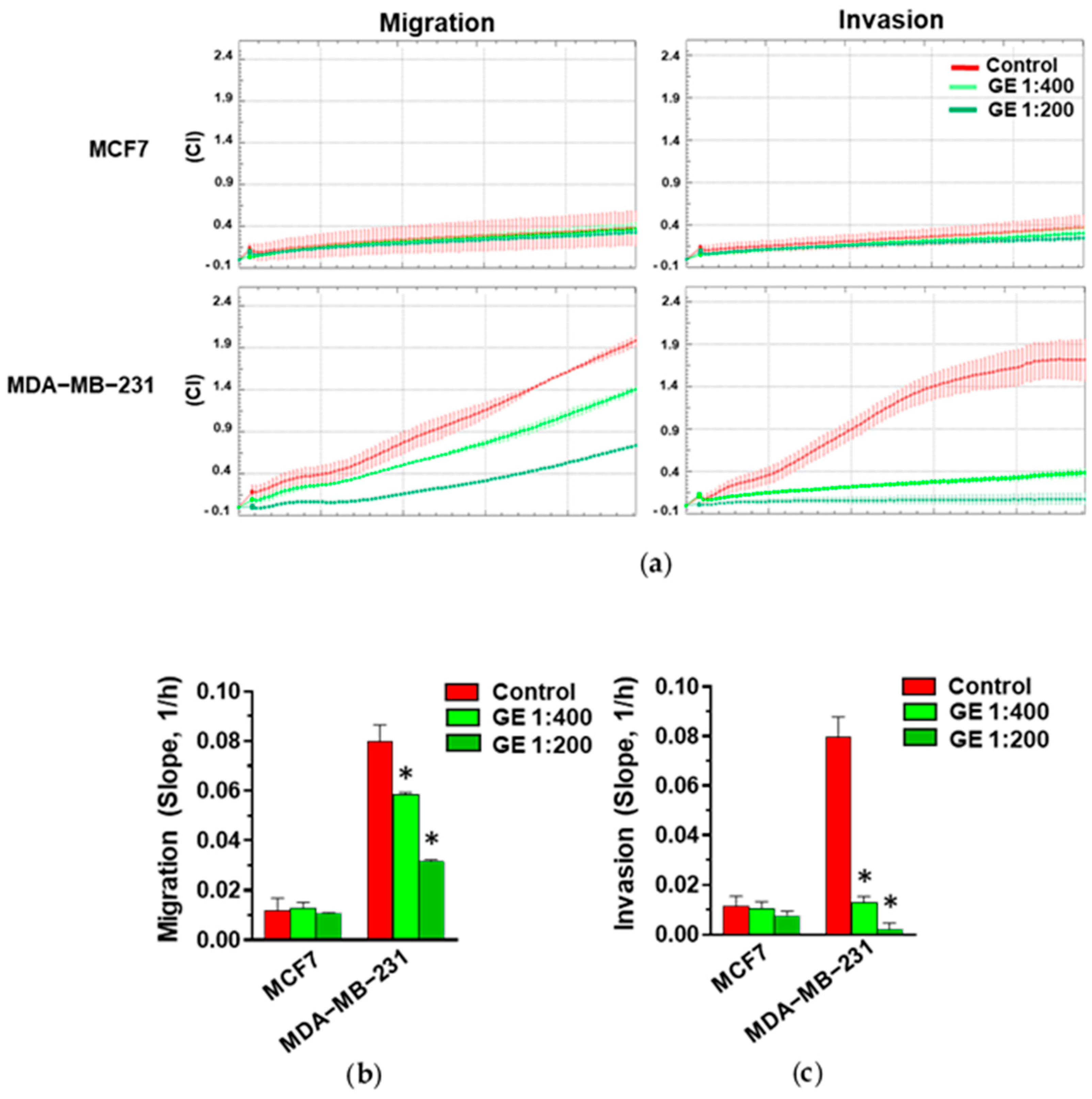
3.1. Effects of Garlic Extract on Growth and Invasive Potential of MCF7 and MDA-MB-231 Breast Tumor Cells
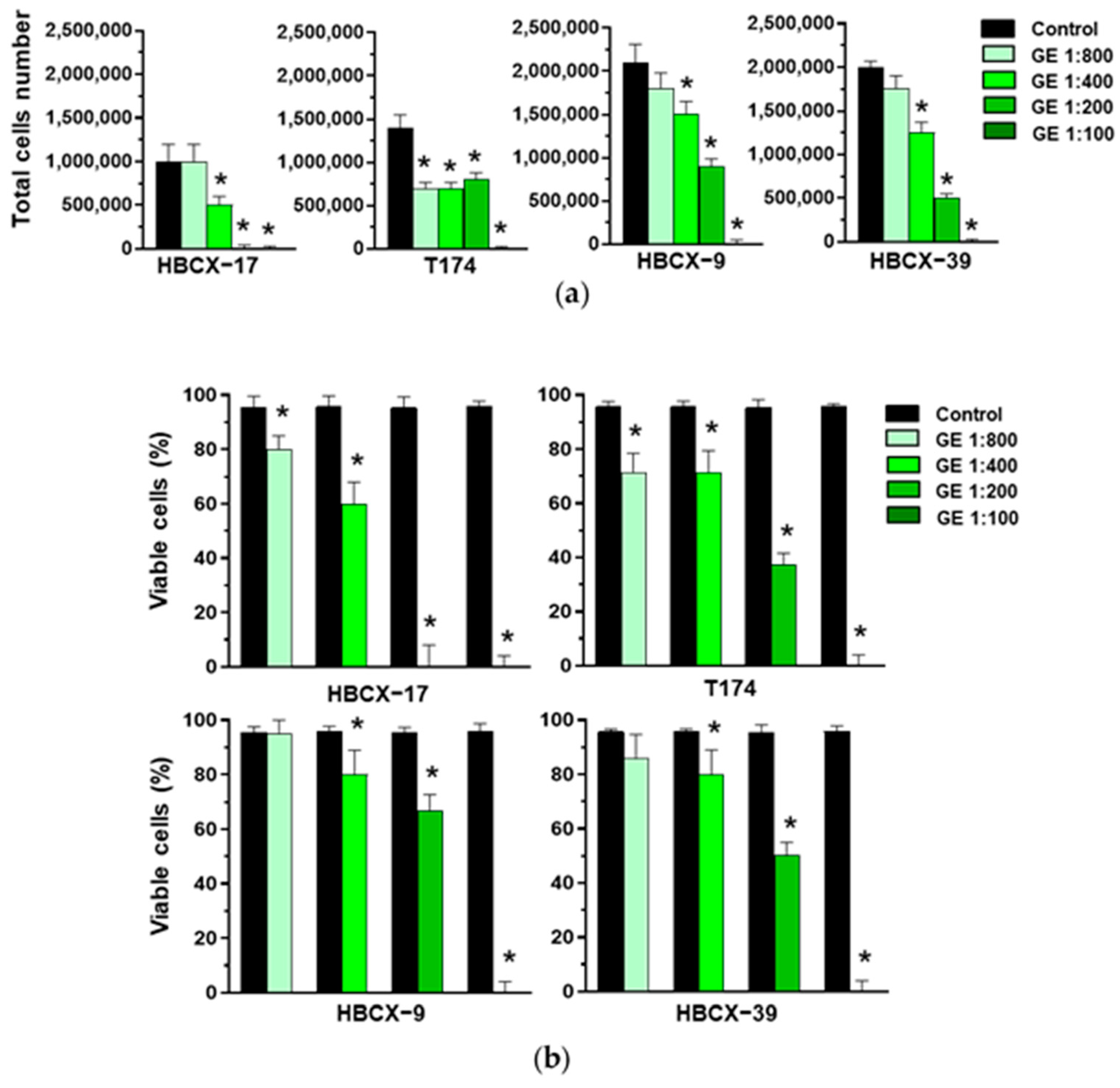
3.2. Effects of Garlic extract on Growth and Invasive Potential of PDX-Derived Breast Tumor Cells with a Triple Negative Phenotype
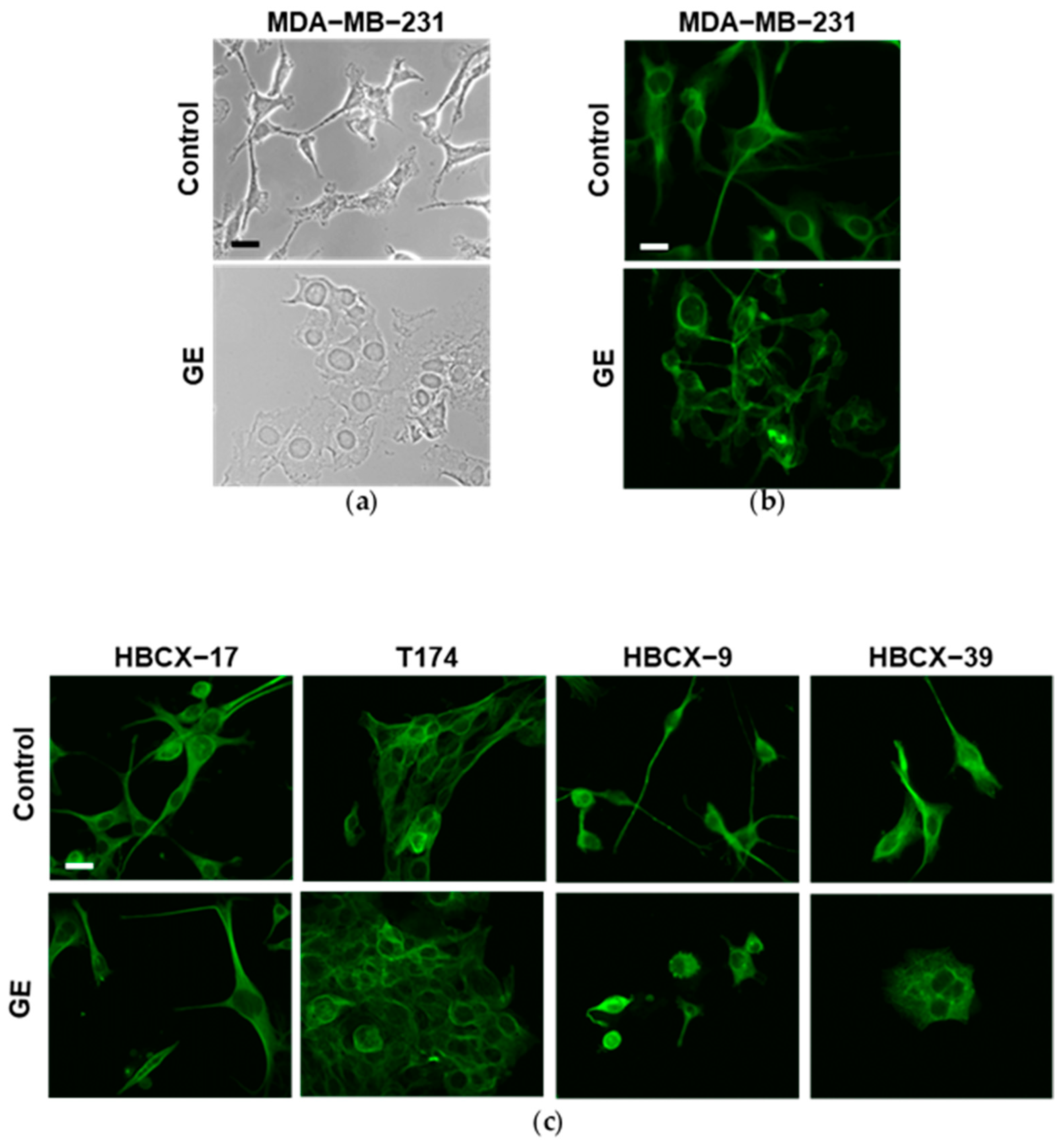
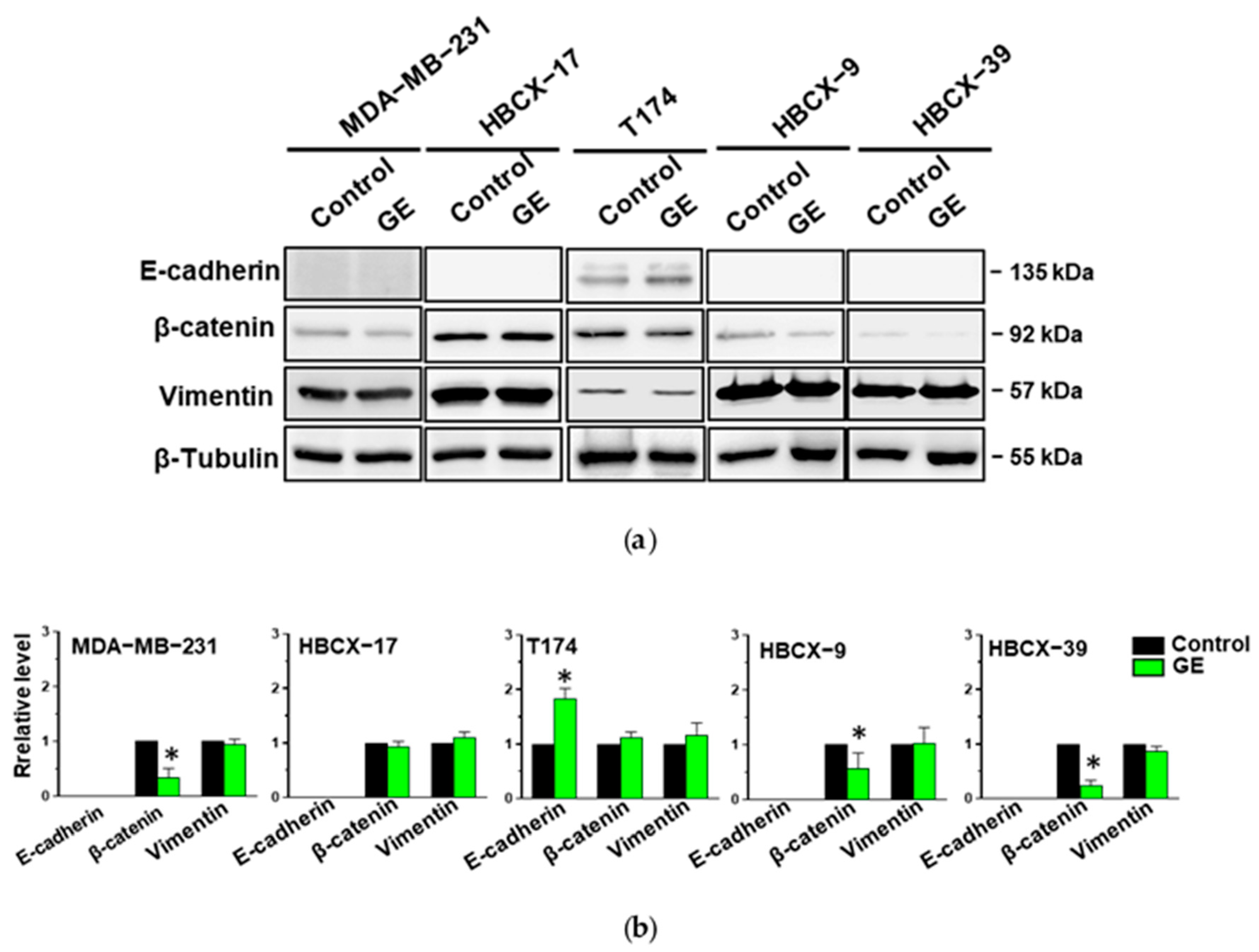
3.3. Effects of Garlic Extract on Epithelial/Mesenchymal Morphology of TNBC Cells
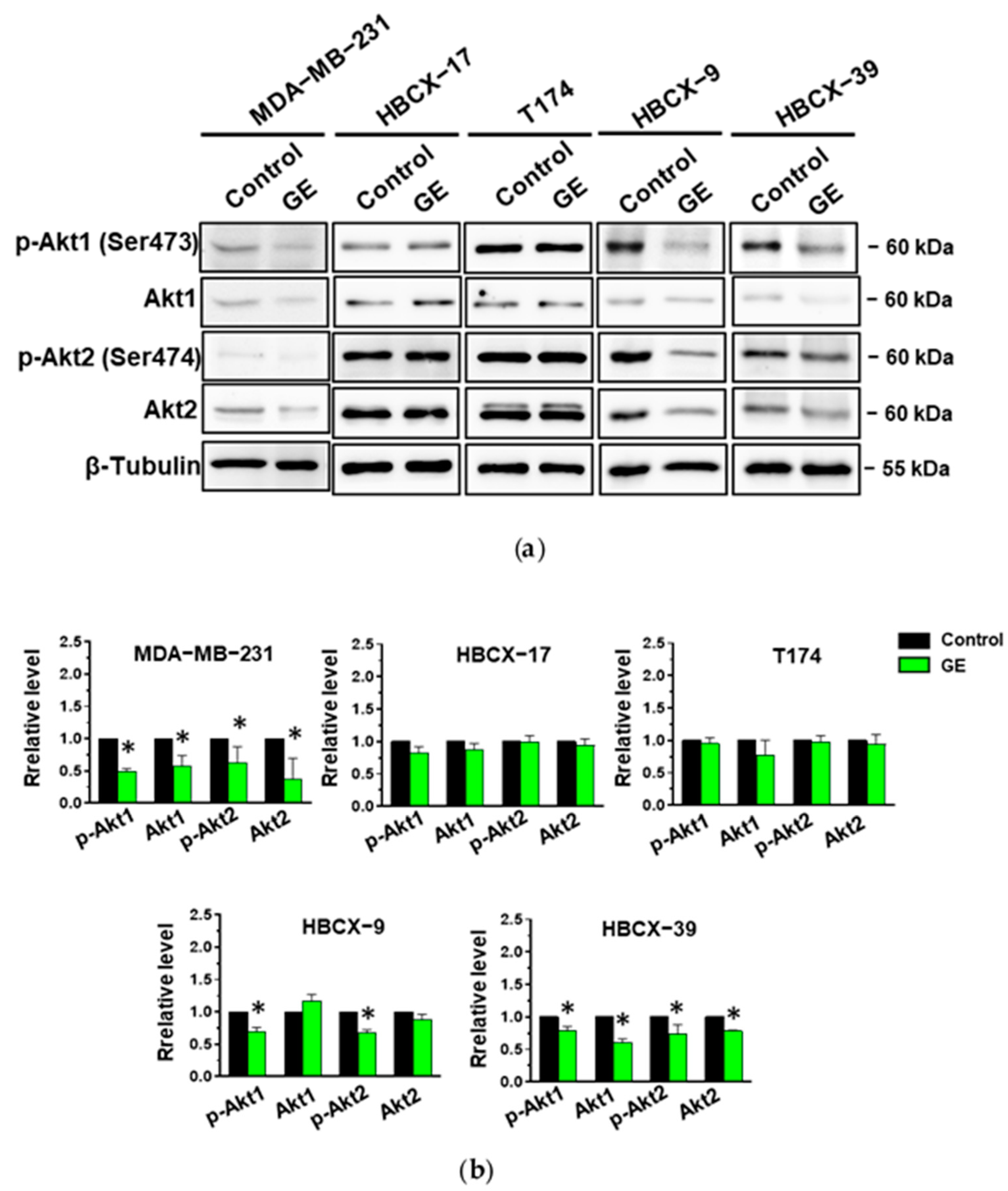
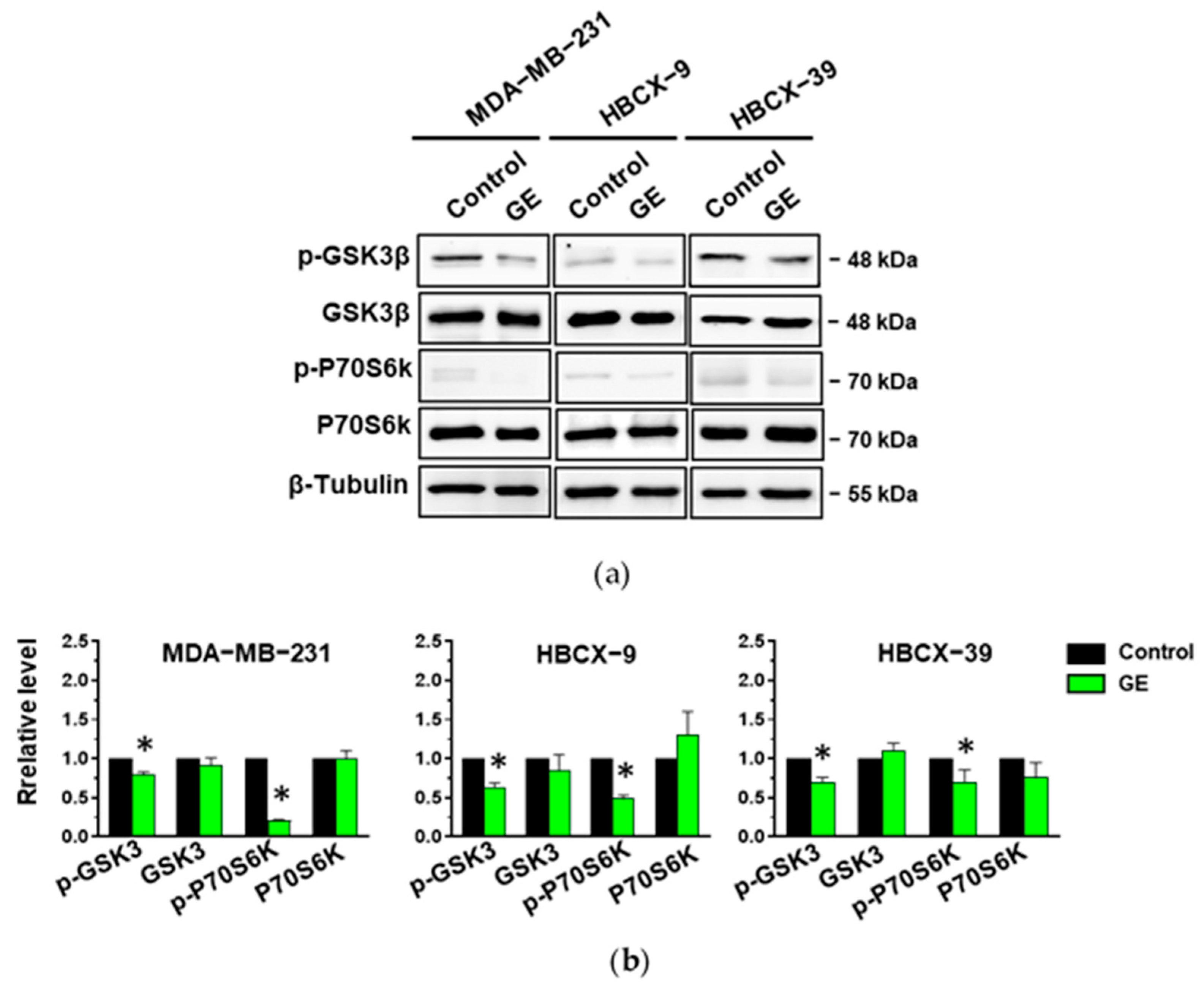
3.4. Effects of GE on Akt in TNBC Cells
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wilkinson, L.; Gathani, T. Understanding Breast Cancer as a Global Health Concern. Br. J. Radiol. 2022, 95, 20211033. [Google Scholar] [CrossRef] [PubMed]
- Iacopetta, D.; Ceramella, J.; Baldino, N.; Sinicropi, M.S.; Catalano, A. Targeting Breast Cancer: An Overlook on Current Strategies. Int. J. Mol. Sci. 2023, 24, 3643. [Google Scholar] [CrossRef] [PubMed]
- Ye, F.; Dewanjee, S.; Li, Y.; Jha, N.K.; Chen, Z.-S.; Kumar, A.; Vishakha; Behl, T.; Jha, S.K.; Tang, H. Advancements in Clinical Aspects of Targeted Therapy and Immunotherapy in Breast Cancer. Mol. Cancer 2023, 22, 105. [Google Scholar] [CrossRef] [PubMed]
- Barrett, S.V. Breast Cancer. J. R. Coll. Physicians Edinb. 2010, 40, 335–338; quiz 339. [Google Scholar] [CrossRef] [PubMed]
- Perou, C.M.; Sørlie, T.; Eisen, M.B.; van de Rijn, M.; Jeffrey, S.S.; Rees, C.A.; Pollack, J.R.; Ross, D.T.; Johnsen, H.; Akslen, L.A.; et al. Molecular Portraits of Human Breast Tumours. Nature 2000, 406, 747–752. [Google Scholar] [CrossRef] [PubMed]
- Bertucci, F.; Finetti, P.; Birnbaum, D. Basal breast cancer: A complex and deadly molecular subtype. Curr. Mol. Med. 2012, 12, 96–110. [Google Scholar] [CrossRef] [PubMed]
- Guido, L.P.; Gomez-Fernandez, C. Advances in the Molecular Taxonomy of Breast Cancer. Arch. Med. Res. 2020, 51, 777–783. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Karakas, C.; Tyburski, H.; Turner, B.M.; Peng, Y.; Wang, X.; Katerji, H.; Schiffhauer, L.; Hicks, D.G. HER2-Low Breast Cancers: Current Insights and Future Directions. Semin. Diagn. Pathol. 2022, 39, 305–312. [Google Scholar] [CrossRef] [PubMed]
- Lehmann, B.D.; Jovanović, B.; Chen, X.; Estrada, M.V.; Johnson, K.N.; Shyr, Y.; Moses, H.L.; Sanders, M.E.; Pietenpol, J.A. Refinement of Triple-Negative Breast Cancer Molecular Subtypes: Implications for Neoadjuvant Chemotherapy Selection. PLoS ONE 2016, 11, e0157368. [Google Scholar] [CrossRef]
- Yin, L.; Duan, J.-J.; Bian, X.-W.; Yu, S. Triple-Negative Breast Cancer Molecular Subtyping and Treatment Progress. Breast Cancer Res. 2020, 22, 61. [Google Scholar] [CrossRef]
- Ensenyat-Mendez, M.; Llinàs-Arias, P.; Orozco, J.I.J.; Íñiguez-Muñoz, S.; Salomon, M.P.; Sesé, B.; DiNome, M.L.; Marzese, D.M. Current Triple-Negative Breast Cancer Subtypes: Dissecting the Most Aggressive Form of Breast Cancer. Front. Oncol. 2021, 11, 681476. [Google Scholar] [CrossRef]
- Keskinkılıc, M.; Gökmen-Polar, Y.; Badve, S.S. Triple Negative Breast Cancers: An Obsolete Entity? Clin. Breast Cancer 2024, 24, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Michaels, E.; Chen, N.; Nanda, R. The Role of Immunotherapy in Triple-Negative Breast Cancer (TNBC). Clin. Breast Cancer 2024, 6, 81526–88209. [Google Scholar] [CrossRef]
- Caputo, R.; Buono, G.; Piezzo, M.; Martinelli, C.; Cianniello, D.; Rizzo, A.; Pantano, F.; Staropoli, N.; Cangiano, R.; Turano, S.; et al. Sacituzumab Govitecan for the treatment of advanced triple negative breast cancer patients: A multi-center real-world analysis. Front. Oncol. 2024, 14, 1362641. [Google Scholar] [CrossRef]
- Wang, D.; Tang, M.; Zhang, P.; Yang, K.; Huang, L.; Wu, M.; Shen, Q.; Yue, J.; Wang, W.; Gong, Y.; et al. Activation of ERβ hijacks the splicing machinery to trigger R-loop formation in triple-negative breast cancer. Proc. Natl. Acad. Sci. USA 2024, 121, e2306814121. [Google Scholar] [CrossRef]
- Bhattacharya, T.; Dutta, S.; Akter, R.; Rahman, M.H.; Karthika, C.; Nagaswarupa, H.P.; Murthy, H.C.A.; Fratila, O.; Brata, R.; Bungau, S. Role of Phytonutrients in Nutrigenetics and Nutrigenomics Perspective in Curing Breast Cancer. Biomolecules 2021, 11, 1176. [Google Scholar] [CrossRef] [PubMed]
- Ansary, J.; Forbes-Hernández, T.Y.; Gil, E.; Cianciosi, D.; Zhang, J.; Elexpuru-Zabaleta, M.; Simal-Gandara, J.; Giampieri, F.; Battino, M. Potential Health Benefit of Garlic Based on Human Intervention Studies: A Brief Overview. Antioxidants 2020, 9, 619. [Google Scholar] [CrossRef]
- Mondal, A.; Banerjee, S.; Bose, S.; Mazumder, S.; Haber, R.A.; Farzaei, M.H.; Bishayee, A. Garlic Constituents for Cancer Prevention and Therapy: From Phytochemistry to Novel Formulations. Pharmacol. Res. 2022, 175, 105837. [Google Scholar] [CrossRef] [PubMed]
- Okoro, B.C.; Dokunmu, T.M.; Okafor, E.; Sokoya, I.A.; Israel, E.N.; Olusegun, D.O.; Bella-Omunagbe, M.; Ebubechi, U.M.; Ugbogu, E.A.; Iweala, E.E.J. The Ethnobotanical, Bioactive Compounds, Pharmacological Activities and Toxicological Evaluation of Garlic (Allium sativum): A Review. Pharmacol. Res.—Mod. Chin. Med. 2023, 8, 100273. [Google Scholar] [CrossRef]
- Zhang, H.; Wang, H.; Qin, L.; Lin, S. Garlic-Derived Compounds: Epigenetic Modulators and Their Antitumor Effects. Phytother. Res. 2024, 38, 1329–1344. [Google Scholar] [CrossRef]
- Pandey, P.; Khan, F.; Alshammari, N.; Saeed, A.; Aqil, F.; Saeed, M. Updates on the Anticancer Potential of Garlic Organosulfur Compounds and Their Nanoformulations: Plant Therapeutics in Cancer Management. Front. Pharmacol. 2023, 14, 1154034. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Zhao, Y.; Wei, Z.; Tao, L.; Sheng, X.; Wang, S.; Chen, J.; Ruan, J.; Liu, Z.; Cao, Y.; et al. Targeting Thioredoxin System with an Organosulfur Compound, Diallyl Trisulfide (DATS), Attenuates Progression and Metastasis of Triple-Negative Breast Cancer (TNBC). Cell. Physiol. Biochem. 2018, 50, 1945–1963. [Google Scholar] [CrossRef] [PubMed]
- Mitra, S.; Das, R.; Emran, T.B.; Labib, R.K.; Tabassum, N.E.; Islam, F.; Sharma, R.; Ahmad, I.; Nainu, F.; Chidambaram, K.; et al. Diallyl Disulfide: A Bioactive Garlic Compound with Anticancer Potential. Front. Pharmacol. 2022, 13, 943967. [Google Scholar] [CrossRef] [PubMed]
- Marni, R.; Kundrapu, D.B.; Chakraborti, A.; Malla, R. Insight into Drug Sensitizing Effect of Diallyl Disulfide and Diallyl Trisulfide from Allium sativum L. on Paclitaxel-Resistant Triple-Negative Breast Cancer Cells. J. Ethnopharmacol. 2022, 296, 115452. [Google Scholar] [CrossRef] [PubMed]
- Kusza, D.A.; Hunter, R.; Schäfer, G.; Smith, M.; Katz, A.A.; Kaschula, C.H. Activity-Based Proteomic Identification of the S-Thiolation Targets of Ajoene in MDA-MB-231 Breast Cancer Cells. J. Agric. Food Chem. 2022, 70, 14679–14692. [Google Scholar] [CrossRef] [PubMed]
- Hahm, E.-R.; Singh, S.V. Gene Expression Changes by Diallyl Trisulfide Administration in Chemically-Induced Mammary Tumors in Rats. J. Cancer Prev. 2022, 27, 22–30. [Google Scholar] [CrossRef] [PubMed]
- Brugnoli, F.; Tedeschi, P.; Grassilli, S.; Maietti, A.; Brandolini, V.; Bertagnolo, V. Ethanol-Based Garlic Extract Prevents Malignant Evolution of Non-Invasive Breast Tumor Cells Induced by Moderate Hypoxia. Biomed. Pharmacother. 2021, 142, 112052. [Google Scholar] [CrossRef]
- Tedeschi, P.; Brugnoli, F.; Merighi, S.; Grassilli, S.; Nigro, M.; Catani, M.; Gessi, S.; Bertagnolo, V.; Travagli, A.; Caboni, M.F.; et al. The Effect of Different Storage Conditions on Phytochemical Composition, Shelf-Life, and Bioactive Compounds of Voghiera Garlic PDO. Antioxidants 2023, 12, 499. [Google Scholar] [CrossRef] [PubMed]
- Grassilli, S.; Vezzali, F.; Cairo, S.; Brugnoli, F.; Volinia, S.; De Mattei, M.; Judde, J.-G.; Bertagnolo, V. Targeting the Vav1/miR-29b Axis as a Potential Approach for Treating Selected Molecular Subtypes of Triple-negative Breast Cancer. Oncol. Rep. 2021, 45, 83. [Google Scholar] [CrossRef]
- Petrovic, V.; Nepal, A.; Olaisen, C.; Bachke, S.; Hira, J.; Søgaard, C.K.; Røst, L.M.; Misund, K.; Andreassen, T.; Melø, T.M.; et al. Anti-Cancer Potential of Homemade Fresh Garlic Extract Is Related to Increased Endoplasmic Reticulum Stress. Nutrients 2018, 10, 450. [Google Scholar] [CrossRef]
- Brugnoli, F.; Grassilli, S.; Al-Qassab, Y.; Capitani, S.; Bertagnolo, V. PLC-Β2 Is Modulated by Low Oxygen Availability in Breast Tumor Cells and Plays a Phenotype Dependent Role in Their Hypoxia-Related Malignant Potential. Mol. Carcinog. 2016, 55, 2210–2221. [Google Scholar] [CrossRef] [PubMed]
- Brugnoli, F.; Grassilli, S.; Piazzi, M.; Palomba, M.; Nika, E.; Bavelloni, A.; Capitani, S.; Bertagnolo, V. In Triple Negative Breast Tumor Cells, PLC-Β2 Promotes the Conversion of CD133high to CD133low Phenotype and Reduces the CD133-Related Invasiveness. Mol. Cancer 2013, 12, 165. [Google Scholar] [CrossRef] [PubMed]
- Souto, E.P.; Dobrolecki, L.E.; Villanueva, H.; Sikora, A.G.; Lewis, M.T. In Vivo Modeling of Human Breast Cancer Using Cell Line and Patient-Derived Xenografts. J. Mammary Gland Biol. Neoplasia 2022, 27, 211–230. [Google Scholar] [CrossRef] [PubMed]
- Cao, C.; Lu, X.; Guo, X.; Zhao, H.; Gao, Y. Patient-Derived Models: Promising Tools for Accelerating the Clinical Translation of Breast Cancer Research Findings. Exp. Cell Res. 2023, 425, 113538. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Wu, W.; Cai, C.; Zhang, H.; Shen, H.; Han, Y. Patient-Derived Xenograft Models in Cancer Therapy: Technologies and Applications. Signal Transduct. Target. Ther. 2023, 8, 160. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Peng, Y.; Tao, X.; Ding, X.; Li, R.; Jiang, Y.; Zuo, W. Microtubule Organization Is Essential for Maintaining Cellular Morphology and Function. Oxid. Med. Cell. Longev. 2022, 2022, e1623181. [Google Scholar] [CrossRef] [PubMed]
- Cerma, K.; Piacentini, F.; Moscetti, L.; Barbolini, M.; Canino, F.; Tornincasa, A.; Caggia, F.; Cerri, S.; Molinaro, A.; Dominici, M.; et al. Targeting PI3K/AKT/mTOR Pathway in Breast Cancer: From Biology to Clinical Challenges. Biomedicines 2023, 11, 109. [Google Scholar] [CrossRef] [PubMed]
- Ito, K.; Ogata, H.; Honma, N.; Shibuya, K.; Mikami, T. Expression of mTOR Signaling Pathway Molecules in Triple-Negative Breast Cancer. Pathobiology 2019, 86, 315–321. [Google Scholar] [CrossRef] [PubMed]
- Vijay, G.V.; Zhao, N.; Den Hollander, P.; Toneff, M.J.; Joseph, R.; Pietila, M.; Taube, J.H.; Sarkar, T.R.; Ramirez-Pena, E.; Werden, S.J.; et al. GSK3β Regulates Epithelial-Mesenchymal Transition and Cancer Stem Cell Properties in Triple-Negative Breast Cancer. Breast Cancer Res. 2019, 21, 37. [Google Scholar] [CrossRef]
- He, R.; Du, S.; Lei, T.; Xie, X.; Wang, Y. Glycogen Synthase Kinase 3β in Tumorigenesis and Oncotherapy. Oncol. Rep. 2020, 44, 2373–2385. [Google Scholar] [CrossRef]
- Berven, L.A.; Crouch, M.F. Cellular function of p70S6K: A role in regulating cell motility. Immunol. Cell Biol. 2000, 78, 447–451. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.-L.; Weng, H.-C.; Hsu, J.-L.; Lin, S.-V.; Guh, J.-H.; Hsu, L.-C. The Combination of MK-2206 and WZB117 Exerts a Synergistic Cytotoxic Effect Against Breast Cancer Cells. Front. Pharmacol. 2019, 10, 1311. [Google Scholar] [CrossRef] [PubMed]
- Miskin, R.P.; Warren, J.S.A.; Ndoye, A.; Wu, L.; Lamar, J.M.; DiPersio, C.M. Integrin α3β1 Promotes Invasive and Metastatic Properties of Breast Cancer Cells through Induction of the Brn-2 Transcription Factor. Cancers 2021, 13, 480. [Google Scholar] [CrossRef] [PubMed]
- Coussy, F.; de Koning, L.; Lavigne, M.; Bernard, V.; Ouine, B.; Boulai, A.; El Botty, R.; Dahmani, A.; Montaudon, E.; Assayag, F.; et al. A large collection of integrated genomically characterized patient-derived xenografts highlighting the heterogeneity of triple-negative breast cancer. Int. J. Cancer. 2019, 145, 1902–1912. [Google Scholar] [CrossRef]
- Sharma, M.P.; Shukla, S.; Misra, G. Recent advances in breast cancer cell line research. Int. J. Cancer 2024, 154, 1683–1693. [Google Scholar] [CrossRef] [PubMed]
- Hiremath, I.S.; Goel, A.; Warrier, S.; Kumar, A.P.; Sethi, G.; Garg, M. The multidimensional role of the Wnt/beta-catenin signaling pathway in human malignancies. J. Cell. Physiol. 2022, 237, 199–238. [Google Scholar] [CrossRef]
- Huang, J.; Yang, B.; Xiang, T.; Peng, W.; Qiu, Z.; Wan, J.; Zhang, L.; Li, H.; Li, H.; Ren, G. Diallyl Disulfide Inhibits Growth and Metastatic Potential of Human Triple-Negative Breast Cancer Cells through Inactivation of the β-Catenin Signaling Pathway. Mol. Nutr. Food Res. 2015, 59, 1063–1075. [Google Scholar] [CrossRef]
- Tsai, P.-J.; Lai, Y.-H.; Manne, R.K.; Tsai, Y.-S.; Sarbassov, D.; Lin, H.-K. Akt: A Key Transducer in Cancer. J. Biomed. Sci. 2022, 29, 76. [Google Scholar] [CrossRef] [PubMed]
- Shin, D.Y.; Kim, G.-Y.; Hwang, H.J.; Kim, W.-J.; Choi, Y.H. Diallyl Trisulfide-Induced Apoptosis of Bladder Cancer Cells Is Caspase-Dependent and Regulated by PI3K/Akt and JNK Pathways. Environ. Toxicol. Pharmacol. 2014, 37, 74–83. [Google Scholar] [CrossRef]
- Jiang, X.; Zhu, X.; Xu, H.; Zhao, Z.; Li, S.; Li, S.; Cai, J.; Cao, J. Diallyl Trisulfide Suppresses Tumor Growth through the Attenuation of Nrf2/Akt and Activation of P38/JNK and Potentiates Cisplatin Efficacy in Gastric Cancer Treatment. Acta Pharmacol. Sin. 2017, 38, 1048–1058. [Google Scholar] [CrossRef]
- Liu, X.; Wang, N.; He, Z.; Chen, C.; Ma, J.; Liu, X.; Deng, S.; Xie, L. Diallyl Trisulfide Inhibits Osteosarcoma 143B Cell Migration, Invasion and EMT by Inducing Autophagy. Heliyon 2024, 10, e26681. [Google Scholar] [CrossRef]
- Shin, D.Y.; Kim, G.-Y.; Lee, J.H.; Choi, B.T.; Yoo, Y.H.; Choi, Y.H. Apoptosis Induction of Human Prostate Carcinoma DU145 Cells by Diallyl Disulfide via Modulation of JNK and PI3K/AKT Signaling Pathways. Int. J. Mol. Sci. 2012, 13, 14158–14171. [Google Scholar] [CrossRef] [PubMed]
- Yin, X.; Feng, C.; Han, L.; Ma, Y.; Jiao, Y.; Wang, J.; Jia, L.; Jing, F.; Gao, X.; Zhang, Y.; et al. Diallyl Disulfide Inhibits the Metastasis of Type II Esophageal-gastric Junction Adenocarcinoma Cells via NF-κB and PI3K/AKT Signaling Pathways in Vitro. Oncol. Rep. 2018, 39, 784–794. [Google Scholar] [CrossRef] [PubMed]
- Malki, A.; El-Saadani, M.; Sultan, A.S. Garlic Constituent Diallyl Trisulfide Induced Apoptosis in MCF7 Human Breast Cancer Cells. Cancer Biol. Ther. 2009, 8, 2174–2184. [Google Scholar] [CrossRef] [PubMed]
- Shi, G.; Li, X.; Wang, W.; Hou, L.; Yin, L.; Wang, L. Allicin Overcomes Doxorubicin Resistance of Breast Cancer Cells by Targeting the Nrf2 Pathway. Cell Biochem. Biophys. 2024. [Google Scholar] [CrossRef] [PubMed]
- Basu, A.; Lambring, C.B. Akt Isoforms: A Family Affair in Breast Cancer. Cancers 2021, 13, 3445. [Google Scholar] [CrossRef] [PubMed]
- Johnson, J.; Chow, Z.; Lee, E.; Weiss, H.L.; Evers, B.M.; Rychahou, P. Role of AMPK and Akt in Triple Negative Breast Cancer Lung Colonization. Neoplasia 2021, 23, 429–438. [Google Scholar] [CrossRef] [PubMed]
- Sridharan, S.; Basu, A. Distinct Roles of mTOR Targets S6K1 and S6K2 in Breast Cancer. Int. J. Mol. Sci. 2020, 21, 1199. [Google Scholar] [CrossRef] [PubMed]
- Sokolosky, M.; Chappell, W.H.; Stadelman, K.; Abrams, S.L.; Davis, N.M.; Steelman, L.S.; McCubrey, J.A. Inhibition of GSK3beta activity can result in drug and hormonal resistance and alter sensitivity to targeted therapy in MCF-7 breast cancer cells. Cell Cycle 2014, 13, 820–833. [Google Scholar] [CrossRef]
- Gerosa, R.; De Sanctis, R.; Jacobs, F.; Benvenuti, C.; Gaudio, M.; Saltalamacchia, G.; Torrisi, R.; Masci, G.; Miggiano, C.; Agustoni, F.; et al. Cyclin-Dependent Kinase 2 (CDK2) Inhibitors and Others Novel CDK Inhibitors (CDKi) in Breast Cancer: Clinical Trials, Current Impact, and Future Directions. Crit. Rev. Oncol. Hematol. 2024, 196, 104324. [Google Scholar] [CrossRef]
- Sanchez-Vega, F.; Mina, M.; Armenia, J.; Chatila, W.K.; Luna, A.; La, K.C.; Dimitriadoy, S.; Liu, D.L.; Kantheti, H.S.; Saghafinia, S.; et al. Oncogenic Signaling Pathways in The Cancer Genome Atlas. Cell 2018, 173, 321–337. [Google Scholar] [CrossRef] [PubMed]
- Mai, N.; Abuhadra, N.; Jhaveri, K. Molecularly Targeted Therapies for Triple Negative Breast Cancer: History, Advances, and Future Directions. Clin. Breast Cancer 2023, 23, 784–799. [Google Scholar] [CrossRef] [PubMed]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Brugnoli, F.; Dell’Aira, M.; Tedeschi, P.; Grassilli, S.; Pierantoni, M.; Foschi, R.; Bertagnolo, V. Effects of Garlic on Breast Tumor Cells with a Triple Negative Phenotype: Peculiar Subtype-Dependent Down-Modulation of Akt Signaling. Cells 2024, 13, 822. https://doi.org/10.3390/cells13100822
Brugnoli F, Dell’Aira M, Tedeschi P, Grassilli S, Pierantoni M, Foschi R, Bertagnolo V. Effects of Garlic on Breast Tumor Cells with a Triple Negative Phenotype: Peculiar Subtype-Dependent Down-Modulation of Akt Signaling. Cells. 2024; 13(10):822. https://doi.org/10.3390/cells13100822
Chicago/Turabian StyleBrugnoli, Federica, Marcello Dell’Aira, Paola Tedeschi, Silvia Grassilli, Marina Pierantoni, Rebecca Foschi, and Valeria Bertagnolo. 2024. "Effects of Garlic on Breast Tumor Cells with a Triple Negative Phenotype: Peculiar Subtype-Dependent Down-Modulation of Akt Signaling" Cells 13, no. 10: 822. https://doi.org/10.3390/cells13100822
APA StyleBrugnoli, F., Dell’Aira, M., Tedeschi, P., Grassilli, S., Pierantoni, M., Foschi, R., & Bertagnolo, V. (2024). Effects of Garlic on Breast Tumor Cells with a Triple Negative Phenotype: Peculiar Subtype-Dependent Down-Modulation of Akt Signaling. Cells, 13(10), 822. https://doi.org/10.3390/cells13100822