
Sexually Dimorphic Alterations in the Transcriptome and Behavior with Loss of Histone Demethylase KDM5C
 , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Mice
2.2. mRNA-Seq
2.3. Behavioral Paradigms
2.4. Contextual Fear Conditioning
2.5. Three-Chambered Social Interaction
2.6. Social Dominance Tube Test
2.7. Resident-Intruder Test
3. Results
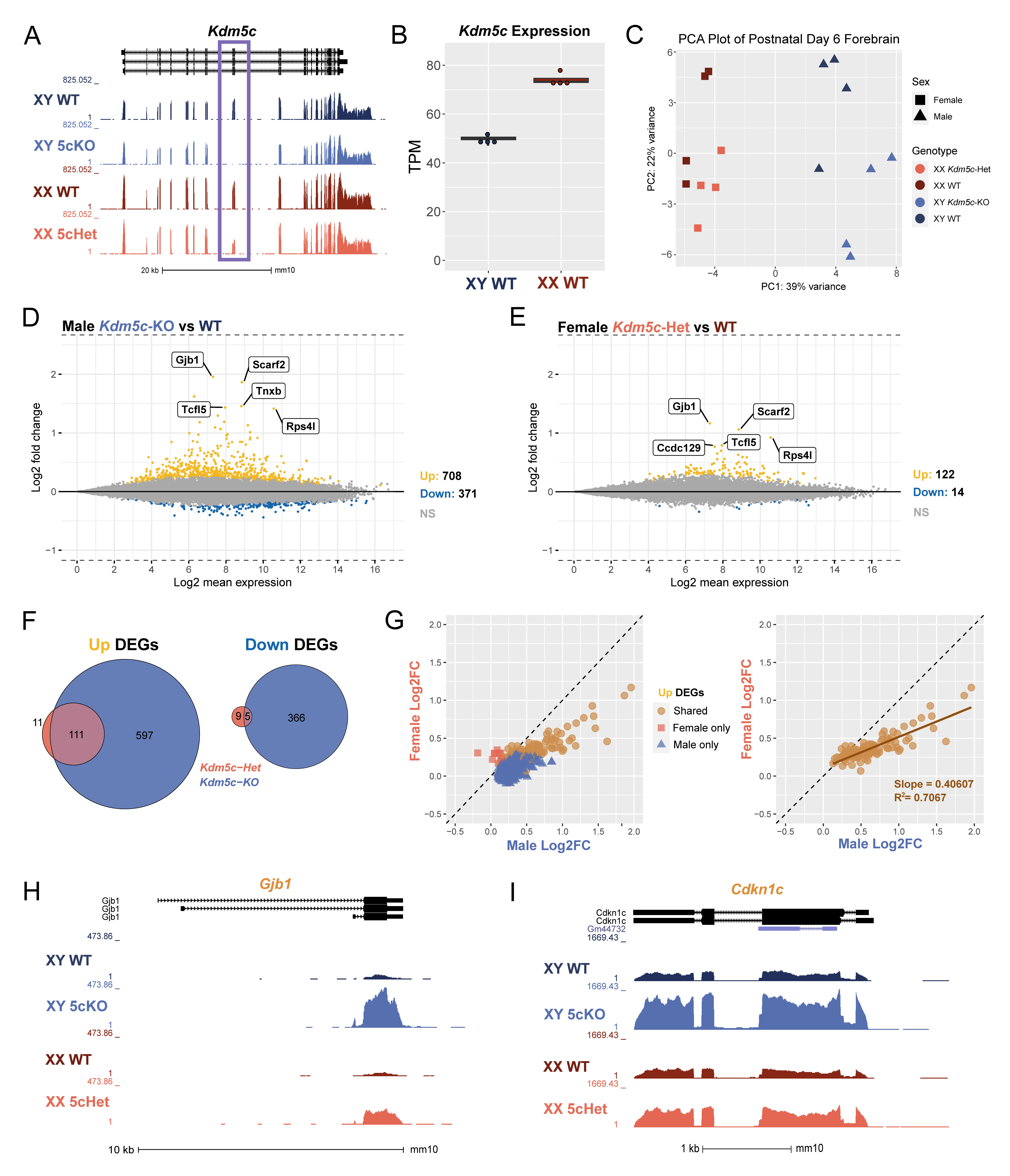
3.1. Kdm5c-Heterozygous Female Brains Show Apparent Gene Expression Changes Similar to Male Mutants
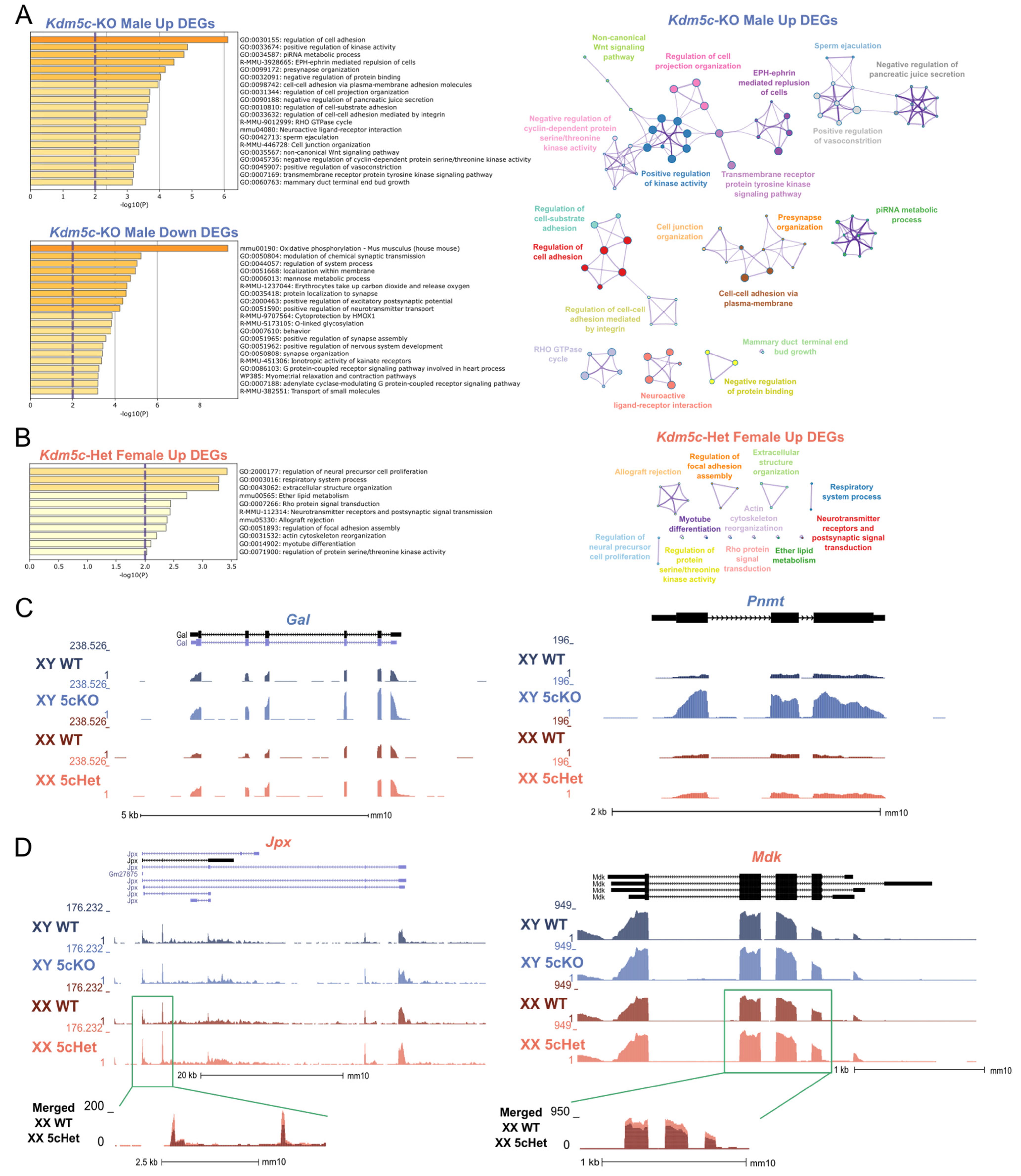
3.2. Kdm5c Deficiency Also Leads to Sex-Specific Gene Expression Changes
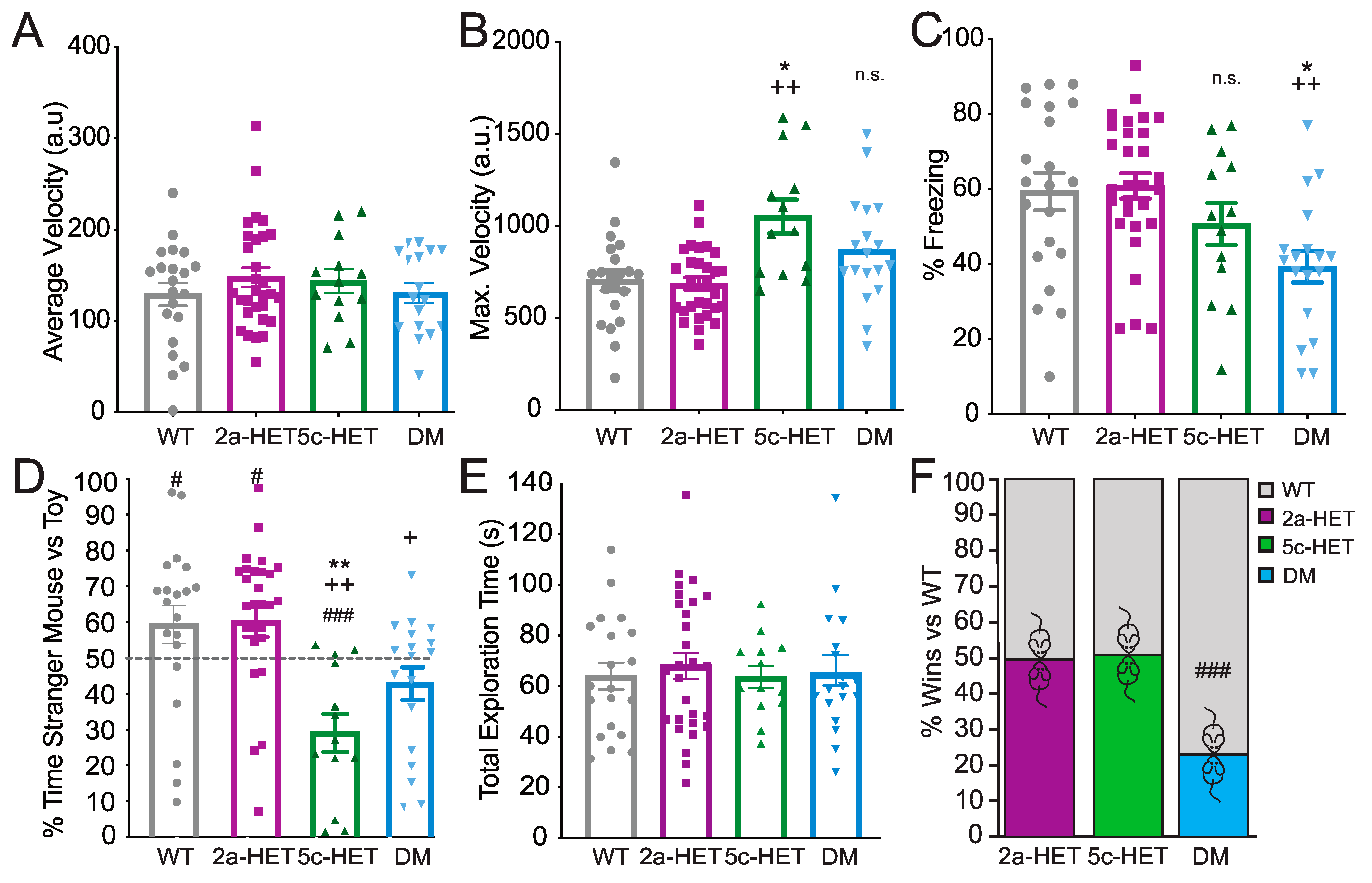
3.3. Kdm5c and Double Kdm5c and Kmt2a Mutations Result in Social Behavior Deficits in Females
3.4. Double Kmt2a and Kdm5c Mutations Mediate Context Fear Memory Impairment in Females
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
Abbreviations
| ATRX | Alpha-thalassemia/mental retardation-X gene |
| DEG | Differentially expressed gene |
| H3K4me | Histone H3K4 methylation |
| KDM5C | Lysine Demethylase 5C |
| Kdm5c-HET | Heterozygous for Kdm5c loss |
| Kdm5c-KO | Kdm5c knockout |
| Log2FC | Log 2 fold change |
| MRXSCJ | Intellectual developmental disorder, X-linked syndromic, Claes-Jensen type |
References
- De Rubeis, S.; He, X.; Goldberg, A.P.; Poultney, C.S.; Samocha, K.; Cicek, A.E.; Kou, Y.; Liu, L.; Fromer, M.; Walker, S.; et al. Synaptic, transcriptional and chromatin genes disrupted in autism. Nature 2014, 515, 209–215. [Google Scholar] [CrossRef] [PubMed]
- Faundes, V.; Newman, W.G.; Bernardini, L.; Canham, N.; Clayton-Smith, J.; Dallapiccola, B.; Davies, S.J.; Demos, M.K.; Goldman, A.; Gill, H.; et al. Histone Lysine Methylases and Demethylases in the Landscape of Human Developmental Disorders. Am. J. Hum. Genet. 2017, 102, 175–187. [Google Scholar] [CrossRef]
- Iossifov, I.; O’Roak, B.J.; Sanders, S.J.; Ronemus, M.; Krumm, N.; Levy, D.; Stessman, H.A.; Witherspoon, K.T.; Vives, L.; Patterson, K.E.; et al. The contribution of de novo coding mutations to autism spectrum disorder. Nature 2014, 515, 216–221. [Google Scholar] [CrossRef]
- Najmabadi, H.; Hu, H.; Garshasbi, M.; Zemojtel, T.; Abedini, S.S.; Chen, W.; Hosseini, M.; Behjati, F.; Haas, S.; Jamali, P.; et al. Deep sequencing reveals 50 novel genes for recessive cognitive disorders. Nature 2011, 478, 57–63. [Google Scholar] [CrossRef] [PubMed]
- The Network and Pathway Analysis Subgroup of the Psychiatric Genomics Consortium Psychiatric genome-wide association study analyses implicate neuronal, immune and histone pathways. Nat. Neurosci. 2015, 18, 199–209. [CrossRef] [PubMed]
- Loke, H.; Harley, V.; Lee, J. Biological factors underlying sex differences in neurological disorders. Int. J. Biochem. Cell Biol. 2015, 65, 139–150. [Google Scholar] [CrossRef]
- van Bokhoven, H. Genetic and Epigenetic Networks in Intellectual Disabilities. Annu. Rev. Genet. 2011, 45, 81–104. [Google Scholar] [CrossRef]
- Hull, L.; Petrides, K.V.; Mandy, W. The Female Autism Phenotype and Camouflaging: A Narrative Review. Rev. J. Autism Dev. Disord. 2020, 7, 306–317. [Google Scholar] [CrossRef]
- Bjornsson, H.T.; Benjamin, J.S.; Zhang, L.; Weissman, J.; Gerber, E.E.; Chen, Y.-C.; Vaurio, R.G.; Potter, M.C.; Hansen, K.D.; Dietz, H.C. Histone deacetylase inhibition rescues structural and functional brain deficits in a mouse model of Kabuki syndrome. Sci. Transl. Med. 2014, 6, 256ra135. [Google Scholar] [CrossRef]
- Schaefer, A.; Sampath, S.C.; Intrator, A.; Min, A.; Gertler, T.S.; Surmeier, D.J.; Tarakhovsky, A.; Greengard, P. Control of Cognition and Adaptive Behavior by the GLP/G9a Epigenetic Suppressor Complex. Neuron 2009, 64, 678–691. [Google Scholar] [CrossRef]
- Kerimoglu, C.; Sakib, M.S.; Jain, G.; Benito, E.; Burkhardt, S.; Capece, V.; Kaurani, L.; Halder, R.; Agís-Balboa, R.C.; Stilling, R.; et al. KMT2A and KMT2B Mediate Memory Function by Affecting Distinct Genomic Regions. Cell Rep. 2017, 20, 538–548. [Google Scholar] [CrossRef] [PubMed]
- Jakovcevski, M.; Ruan, H.; Shen, E.Y.; Dincer, A.; Javidfar, B.; Ma, Q.; Peter, C.J.; Cheung, I.; Mitchell, A.C.; Jiang, Y.; et al. Neuronal Kmt2a/Mll1 Histone Methyltransferase Is Essential for Prefrontal Synaptic Plasticity and Working Memory. J. Neurosci. 2015, 35, 5097–5108. [Google Scholar] [CrossRef] [PubMed]
- Vallianatos, C.N.; Raines, B.; Porter, R.S.; Bonefas, K.M.; Wu, M.C.; Garay, P.M.; Collette, K.M.; Seo, Y.A.; Dou, Y.; Keegan, C.E.; et al. Mutually suppressive roles of KMT2A and KDM5C in behaviour, neuronal structure, and histone H3K4 methylation. Commun. Biol. 2020, 3, 1–14. [Google Scholar] [CrossRef]
- Iwase, S.; Brookes, E.; Agarwal, S.; Badeaux, A.I.; Ito, H.; Vallianatos, C.N.; Tomassy, G.S.; Kasza, T.; Lin, G.; Thompson, A.; et al. A Mouse Model of X-linked Intellectual Disability Associated with Impaired Removal of Histone Methylation. Cell Rep. 2016, 14, 1000–1009. [Google Scholar] [CrossRef]
- Iacobucci, S.; Padilla, N.; Gabrielli, M.; Navarro, C.; Lombardi, M.; Vicioso-Mantis, M.; Verderio, C.; de la Cruz, X.; Martínez-Balbás, M.A. The histone demethylase PHF8 regulates astrocyte differentiation and function. Development 2021, 148I, dev194951. [Google Scholar] [CrossRef]
- Balemans, M.C.; Huibers, M.M.; Eikelenboom, N.W.; Kuipers, A.J.; van Summeren, R.C.; Pijpers, M.M.; Tachibana, M.; Shinkai, Y.; van Bokhoven, H.; Van der Zee, C.E. Reduced exploration, increased anxiety, and altered social behavior: Autistic-like features of euchromatin histone methyltransferase 1 heterozygous knockout mice. Behav. Brain Res. 2010, 208, 47–55. [Google Scholar] [CrossRef] [PubMed]
- Martin-Kenny, N.; Bérubé, N.G. Effects of a postnatal Atrx conditional knockout in neurons on autism-like behaviours in male and female mice. J. Neurodev. Disord. 2020, 12, 1–11. [Google Scholar] [CrossRef]
- Tamming, R.J.; Dumeaux, V.; Jiang, Y.; Shafiq, S.; Langlois, L.; Ellegood, J.; Qiu, L.R.; Lerch, J.P.; Bérubé, N.G. Atrx Deletion in Neurons Leads to Sexually Dimorphic Dysregulation of miR-137 and Spatial Learning and Memory Deficits. Cell Rep. 2020, 31, 107838. [Google Scholar] [CrossRef]
- Jeon, S.J.; Gonzales, E.L.; Mabunga, D.F.N.; Valencia, S.T.; Kim, D.G.; Kim, Y.; Adil, K.J.L.; Shin, D.; Park, D.; Shin, C.Y. Sex-specific Behavioral Features of Rodent Models of Autism Spectrum Disorder. Exp. Neurobiol. 2018, 27, 321–343. [Google Scholar] [CrossRef]
- Iwase, S.; Lan, F.; Bayliss, P.; De La Torre-Ubieta, L.; Huarte, M.; Qi, H.; Whetstine, J.R.; Bonni, A.; Roberts, T.M.; Shi, Y. The X-Linked Mental Retardation Gene SMCX/JARID1C Defines a Family of Histone H3 Lysine 4 Demethylases. Cell 2007, 128, 1077–1088. [Google Scholar] [CrossRef]
- Tahiliani, M.; Mei, P.; Fang, R.; Leonor, T.; Rutenberg, M.; Shimizu, F.; Li, J.; Rao, A.; Shi, Y. The histone H3K4 demethylase SMCX links REST target genes to X-linked mental retardation. Nature 2007, 447, 601–605. [Google Scholar] [CrossRef] [PubMed]
- Jensen, L.R.; Amende, M.; Gurok, U.; Moser, B.; Gimmel, V.; Tzschach, A.; Janecke, A.R.; Tariverdian, G.; Chelly, J.; Fryns, J.-P.; et al. Mutations in the JARID1C Gene, Which Is Involved in Transcriptional Regulation and Chromatin Remodeling, Cause X-Linked Mental Retardation. Am. J. Hum. Genet. 2005, 76, 227–236. [Google Scholar] [CrossRef] [PubMed]
- Simensen, R.J.; Rogers, R.C.; Collins, J.S.; Abidi, F.; Schwartz, C.E.; Stevenson, R.E. Short-term memory deficits in carrier females with KDM5C mutations. Genet. Couns. 2012, 23, 31. [Google Scholar] [PubMed]
- Santos-Rebouças, C.B.; Fintelman-Rodrigues, N.; Jensen, L.R.; Kuss, A.W.; Ribeiro, M.G.; Campos, M.; Santos, J.M.; Pimentel, M.M. A novel nonsense mutation in KDM5C/JARID1C gene causing intellectual disability, short stature and speech delay. Neurosci. Lett. 2011, 498, 67–71. [Google Scholar] [CrossRef] [PubMed]
- Õunap, K.; Puusepp-Benazzouz, H.; Peters, M.; Vaher, U.; Rein, R.; Proos, A.; Field, M.; Reimand, T. A novel c.2T > C mutation of the KDM5C/JARID1C gene in one large family with X-linked intellectual disability. Eur. J. Med. Genet. 2012, 55, 178–184. [Google Scholar] [CrossRef]
- Carmignac, V.; Nambot, S.; Lehalle, D.; Callier, P.; Moortgat, S.; Benoit, V.; Ghoumid, J.; Delobel, B.; Smol, T.; Thuillier, C.; et al. Further delineation of the female phenotype with KDM5C disease causing variants: 19 new individuals and review of the literature. Clin. Genet. 2020, 98, 43–55. [Google Scholar] [CrossRef]
- Lippa, N.C.; Barua, S.; Aggarwal, V.; Pereira, E.; Bain, J.M. A novel de novo KDM5C variant in a female with global developmental delay and ataxia: A case report. BMC Neurol. 2021, 21, 358. [Google Scholar] [CrossRef]
- Hatch, H.A.M.; O’Neil, M.H.; Marion, R.W.; Secombe, J.; Shulman, L.H. Caregiver-reported characteristics of children diagnosed with pathogenic variants in KDM5C. Am. J. Med. Genet. Part A 2021, 185, 2951–2958. [Google Scholar] [CrossRef]
- Berletch, J.B.; Yang, F.; Disteche, C.M. Escape from X inactivation in mice and humans. Genome Biol. 2010, 11, 213. [Google Scholar] [CrossRef]
- Samanta, M.K.; Gayen, S.; Harris, C.; Maclary, E.; Murata-Nakamura, Y.; Malcore, R.M.; Porter, R.S.; Garay, P.M.; Vallianatos, C.N.; Samollow, P.B.; et al. Activation of Xist by an evolutionarily conserved function of KDM5C demethylase. Nat. Commun. 2022, 13, 2602. [Google Scholar] [CrossRef]
- Scandaglia, M.; Lopez-Atalaya, J.P.; Medrano-Fernandez, A.; Lopez-Cascales, M.T.; del Blanco, B.; Lipinski, M.; Benito, E.; Olivares, R.; Iwase, S.; Shi, Y.; et al. Loss of Kdm5c Causes Spurious Transcription and Prevents the Fine-Tuning of Activity-Regulated Enhancers in Neurons. Cell Rep. 2017, 21, 47–59. [Google Scholar] [CrossRef]
- Strom, S.P.; Lozano, R.; Lee, H.; Dorrani, N.; Mann, J.; O’Lague, P.F.; Mans, N.; Deignan, J.L.; Vilain, E.; Nelson, S.F.; et al. De Novo variants in the KMT2A (MLL) gene causing atypical Wiedemann-Steiner syndrome in two unrelated individuals identified by clinical exome sequencing. BMC Med. Genet. 2014, 15, 49. [Google Scholar] [CrossRef] [PubMed]
- Jones, W.D.; Dafou, D.; McEntagart, M.; Woollard, W.J.; Elmslie, F.V.; Holder-Espinasse, M.; Irving, M.; Saggar, A.K.; Smithson, S.; Trembath, R.C.; et al. De Novo Mutations in MLL Cause Wiedemann-Steiner Syndrome. Am. J. Hum. Genet. 2012, 91, 358–364. [Google Scholar] [CrossRef] [PubMed]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed]
- Zhu, A.; Ibrahim, J.G.; Love, M.I. Heavy-tailed prior distributions for sequence count data: Removing the noise and preserving large differences. Bioinformatics 2018, 35, 2084–2092. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Zhou, B.; Pache, L.; Chang, M.; Khodabakhshi, A.H.; Tanaseichuk, O.; Benner, C.; Chanda, S.K. Metascape provides a biologist-oriented resource for the analysis of systems-level datasets. Nat. Commun. 2019, 10, 1523. [Google Scholar] [CrossRef] [PubMed]
- Hong, G.; Zhang, W.; Li, H.; Shen, X.; Guo, Z. Separate enrichment analysis of pathways for up- and downregulated genes. J. R. Soc. Interface 2014, 11, 20130950. [Google Scholar] [CrossRef]
- Keiser, A.A.; Turnbull, L.M.; Darian, M.A.; Feldman, D.E.; Song, I.; Tronson, N.C. Sex Differences in Context Fear Generalization and Recruitment of Hippocampus and Amygdala during Retrieval. Neuropsychopharmacology 2016, 42, 397–407. [Google Scholar] [CrossRef]
- Sheardown, S.; Norris, D.; Fisher, A.; Brockdorff, N. The mouse Smcx gene exhibits developmental and tissue specific variation in degree of escape from X inactivation. Hum. Mol. Genet. 1996, 5, 1355–1360. [Google Scholar] [CrossRef]
- Agulnik, A.I.; Avner, P.A.; Lerner, J.L.; Mitchell, M.J.; Mattei, M.-G.; Borsani, G.; Bishop, C.E. A novel X gene with a widely transcribed Y-linked homologue escapes X-inactivation in mouse and human. Hum. Mol. Genet. 1994, 3, 879–884. [Google Scholar] [CrossRef]
- Carrel, L.; Hunt, P.A.; Willard, H.F. Tissue and lineage-specific variation in inactive X chromosome expression of the murine Smcx gene. Hum. Mol. Genet. 1996, 5, 1361–1366. [Google Scholar] [CrossRef] [PubMed]
- Nadarajah, B.; Thomaidou, D.; Evans, W.; Parnavelas, J. Gap junctions in the adult cerebral cortex: Regional differences in their distribution and cellular expression of connexins. J. Comp. Neurol. 1996, 376, 326–342. [Google Scholar] [CrossRef]
- Nadarajah, B.; Jones, A.M.; Evans, W.H.; Parnavelas, J.G. Differential Expression of Connexins during Neocortical Development and Neuronal Circuit Formation. J. Neurosci. 1997, 17, 3096–3111. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Shen, H.; Naus, C.; Zhang, L.; Carlen, P. Upregulation of gap junction connexin 32 with epileptiform activity in the isolated mouse hippocampus. Neuroscience 2001, 105, 589–598. [Google Scholar] [CrossRef] [PubMed]
- Imaizumi, Y.; Furutachi, S.; Watanabe, T.; Miya, H.; Kawaguchi, D.; Gotoh, Y. Role of the imprinted allele of the Cdkn1c gene in mouse neocortical development. Sci. Rep. 2020, 10, 1884. [Google Scholar] [CrossRef]
- McNamara, G.I.; Davis, B.A.; Browne, M.; Humby, T.; Dalley, J.W.; Xia, J.; John, R.M.; Isles, A.R. Dopaminergic and behavioural changes in a loss-of-imprinting model of Cdkn1c. Genes Brain Behav. 2017, 17, 149–157. [Google Scholar] [CrossRef]
- Goldstein, M.; Fuxe, K.; Hökfelt, T. Characterization and tissue localization of catecholamine synthesizing enzymes. Pharmacol. Rev. 1972, 24, 293–309. [Google Scholar]
- Sørensen, D.B.; Johnsen, P.F.; Bibby, B.M.; Böttner, A.; Bornstein, S.R.; Eisenhofer, G.; Pacak, K.; Hansen, A.K. PNMT Transgenic Mice Have an Aggressive Phenotype. Horm. Metab. Res. 2005, 37, 159–163. [Google Scholar] [CrossRef]
- Mechenthaler, I. Galanin–25 years with a multitalented neuropeptide. Cell. Mol. Life Sci. 2008, 65, 1826–1835. [Google Scholar] [CrossRef]
- Mills, E.G.; Izzi-Engbeaya, C.; Abbara, A.; Comninos, A.N.; Dhillo, W.S. Functions of galanin, spexin and kisspeptin in metabolism, mood and behaviour. Nat. Rev. Endocrinol. 2020, 17, 97–113. [Google Scholar] [CrossRef]
- Winkler, C.; Yao, S. The midkine family of growth factors: Diverse roles in nervous system formation and maintenance. Br. J. Pharmacol. 2014, 171, 905–912. [Google Scholar] [CrossRef] [PubMed]
- Muramatsu, H.; Shirahama, H.; Yonezawa, S.; Maruta, H.; Muramatsu, T. Midkine, A Retinoic Acid-Inducible Growth/Differentiation Factor: Immunochemical Evidence for the Function and Distribution. Dev. Biol. 1993, 159, 392–402. [Google Scholar] [CrossRef] [PubMed]
- Tian, D.; Sun, S.; Lee, J.T. The Long Noncoding RNA, Jpx, Is a Molecular Switch for X Chromosome Inactivation. Cell 2010, 143, 390–403. [Google Scholar] [CrossRef] [PubMed]
- Sun, S.; Del Rosario, B.C.; Szanto, A.; Ogawa, Y.; Jeon, Y.; Lee, J.T. Jpx RNA Activates Xist by Evicting CTCF. Cell 2013, 153, 1537–1551. [Google Scholar] [CrossRef] [PubMed]
- Nudel, R.; Simpson, N.H.; Baird, G.; O’Hare, A.; Conti-Ramsden, G.; Bolton, P.F.; Hennessy, E.R.; SLIConsortium Ring, S.M.; Davey Smith, G.; Francks, C. Genome-wide association analyses of child genotype effects and parent-of-origin effects in specific language impairment. Genes Brain Behav. 2014, 13, 418–429. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, K.; Wanaka, A.; Takatsuji, K.; Muramatsu, H.; Tohyama, M. A novel family of heparin-binding growth factors, pleiotrophin and midkine, is expressed in the developing rat cerebral cortex. Dev. Brain Res. 1994, 79, 229–241. [Google Scholar] [CrossRef]
- Kojima, S.; Inui, T.; Muramatsu, H.; Kimura, T.; Sakakibara, S.; Muramatsu, T. Midkine Is a Heat and Acid Stable Polypeptide Capable of Enhancing Plasminogen Activator Activity and Neurite Outgrowth Extension. Biochem. Biophys. Res. Commun. 1995, 216, 574–581. [Google Scholar] [CrossRef]
- Yamamoto, R.; Song, K.; Yanagisawa, H.-A.; Fox, L.; Yagi, T.; Wirschell, M.; Hirono, M.; Kamiya, R.; Nicastro, D.; Sale, W.S. The MIA complex is a conserved and novel dynein regulator essential for normal ciliary motility. J. Cell Biol. 2013, 201, 263–278. [Google Scholar] [CrossRef]
- Sakoda, Y.; Hashimoto, D.; Asakura, S.; Takeuchi, K.; Harada, M.; Tanimoto, M.; Teshima, T. Donor-derived thymic-dependent T cells cause chronic graft-versus-host disease. Blood 2006, 109, 1756–1764. [Google Scholar] [CrossRef]
- Tanaka, T.; de Silanes, I.L.; Sharova, L.V.; Akutsu, H.; Yoshikawa, T.; Amano, H.; Yamanaka, S.; Gorospe, M.; Ko, M.S.H. Esg1, expressed exclusively in preimplantation embryos, germline, and embryonic stem cells, is a putative RNA-binding protein with broad RNA targets. Dev. Growth Differ. 2006, 48, 381–390. [Google Scholar] [CrossRef]
- Guo, L.; Lin, L.; Wang, X.; Gao, M.; Cao, S.; Mai, Y.; Wu, F.; Kuang, J.; Liu, H.; Yang, J.; et al. Resolving Cell Fate Decisions during Somatic Cell Reprogramming by Single-Cell RNA-Seq. Mol. Cell 2019, 73, 815–829.e7. [Google Scholar] [CrossRef] [PubMed]
- Hodge, R.G.; Ridley, A.J. Regulation and functions of RhoU and RhoV. Small GTPases 2020, 11, 8–15. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Lai, F.; Niswander, L. The ubiquitin ligase mLin41 temporally promotes neural progenitor cell maintenance through FGF signaling. Genes Dev. 2012, 26, 803–815. [Google Scholar] [CrossRef] [PubMed]
- Cuevas, E.; Rybak-Wolf, A.; Rohde, A.M.; Nguyen, D.T.T.; Wulczyn, F.G. Lin41/Trim71 is essential for mouse development and spe-cifically expressed in postnatal ependymal cells of the brain. Front. Cell Dev. Biol. 2015, 3, 20. [Google Scholar] [CrossRef] [PubMed]
- Ozawa, M.; Muramatsu, T. Reticulocalbin, a novel endoplasmic reticulum resident Ca(2+)-binding protein with multiple EF-hand motifs and a carboxyl-terminal HDEL sequence. J. Biol. Chem. 1993, 268, 699–705. [Google Scholar] [CrossRef] [PubMed]
- Sterky, F.H.; Trotter, J.H.; Lee, S.; Recktenwald, C.V.; Du, X.; Zhou, B.; Zhou, P.; Schwenk, J.; Fakler, B.; Südhof, T.C. Carbonic anhydrase-related protein CA10 is an evolutionarily conserved pan-neurexin ligand. Proc. Natl. Acad. Sci. USA 2017, 114, E1253–E1262. [Google Scholar] [CrossRef]
- Kleopa, K.A.; Abrams, C.K.; Scherer, S.S. How do mutations in GJB1 cause X-linked Charcot–Marie–Tooth disease? Brain Res. 2012, 1487, 198–205. [Google Scholar] [CrossRef]
- Russell, A. A syndrome of intra-uterine dwarfism recognizable at birth with cranio-facial dysostosis, disproportionately short arms, and other anomalies (5 examples). Proc. R. Soc. Med. 1954, 47, 1040–1044. [Google Scholar]
- Silver, H.K.; Kiyasu, W.; George, J.; Deamer, W.C. Syndrome of Congenital Hemihypertrophy, Shortness of Stature, and Elevated Urinary Gonadotropins. Pediatrics 1953, 12, 368–376. [Google Scholar] [CrossRef]
- Andrews, S.C.; Wood, M.D.; Tunster, S.J.; Barton, S.C.; Surani, M.A.; John, R.M. Cdkn1c (p57 Kip2) is the major regulator of embryonic growth within its imprinted domain on mouse distal chromosome. BMC Dev. Biol. 2007, 7, 53. [Google Scholar] [CrossRef]
- Wrenn, C.; Marriott, L.; Kinney, J.; Holmes, A.; Wenk, G.; Crawley, J. Galanin peptide levels in hippocampus and cortex of galanin-overexpressing transgenic mice evaluated for cognitive performance. Neuropeptides 2002, 36, 413–426. [Google Scholar] [CrossRef] [PubMed]
- Steiner, R.A.; Hohmann, J.G.; Holmes, A.; Wrenn, C.C.; Cadd, G.; Juréus, A.; Clifton, D.K.; Luo, M.; Gutshall, M.; Ma, S.Y.; et al. Galanin transgenic mice display cognitive and neurochemical deficits characteristic of Alzheimer’s disease. Proc. Natl. Acad. Sci. USA 2001, 98, 4184–4189. [Google Scholar] [CrossRef] [PubMed]
- Rustay, N.R.; Wrenn, C.C.; Kinney, J.W.; Holmes, A.; Bailey, K.R.; Sullivan, T.L.; Harris, A.P.; Long, K.C.; Saavedra, M.C.; Starosta, G.; et al. Galanin impairs performance on learning and memory tasks: Findings from galanin transgenic and GAL-R1 knockout mice. Neuropeptides 2005, 39, 239–243. [Google Scholar] [CrossRef] [PubMed]
- Phillips, R.G.; LeDoux, J.E. A confocal microscopic analysis of galaninergic hyperinnervation of cholinergic basal forebrain neurons in Alzheimer’s disease. Brain Pathol. 1997, 7, 723–730. [Google Scholar] [CrossRef]
- Phillips, R.G.; LeDoux, J.E. Lesions of the dorsal hippocampal formation interfere with background but not foreground contextual fear conditioning. Learn. Mem. 1994, 1, 34–44. [Google Scholar] [CrossRef] [PubMed]
- Fanselow, M.S. From contextual fear to a dynamic view of memory systems. Trends Cogn. Sci. 2010, 14, 7–15. [Google Scholar] [CrossRef]
- Tronson, N.C.; Keiser, A.A. A Dynamic Memory Systems Framework for Sex Differences in Fear Memory. Trends Neurosci. 2019, 42, 680–692. [Google Scholar] [CrossRef]
- Bellott, D.W.; Hughes, J.F.; Skaletsky, H.; Brown, L.G.; Pyntikova, T.; Cho, T.-J.; Koutseva, N.; Zaghlul, S.; Graves, T.; Rock, S.; et al. Mammalian Y chromosomes retain widely expressed dosage-sensitive regulators. Nature 2014, 508, 494–499. [Google Scholar] [CrossRef]
- Posynick, B.J.; Brown, C.J. Escape From X-Chromosome Inactivation: An Evolutionary Perspective. Front. Cell Dev. Biol. 2019, 7, 241. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| MGI Symbol | Full Name | Ensembl | Base Mean | XX log2FC | XX padj | XY log2FC | XY padj | Feature Type | Known Functions |
|---|---|---|---|---|---|---|---|---|---|
| Arhgef19 | Rho guanine nucleotide exchange factor (GEF) 19 | ENSMUSG00000028919 | 98.585 | 0.346 | 0.021 | 0.088 | 0.739 | protein coding gene | Not much known in the brain, but SNPs are implicated in language impairments [55]. |
| Gm46620 | predicted gene, 46620 | ENSMUSG00000118012 | 165.056 | 0.305 | 0.061 | −0.185 | 0.253 | lncRNA gene | Gene module with no known function |
| Mdk | midkine | ENSMUSG00000027239 | 331.224 | 0.305 | 0.028 | 0.116 | 0.546 | protein coding gene | Growth factor highly expressed during early brain development [56]. Promotes neurite outgrowth and survival [57]. |
| Cfap100 | cilia and flagella associated protein 100 | ENSMUSG00000048794 | 122.031 | 0.305 | 0.098 | 0.140 | 0.493 | protein coding gene | Unknown function, but structurally similar to proteins involved in cilia and flagella motility [58]. |
| H2-Ab1 | histocompatibility 2, class II antigen A, beta 1 | ENSMUSG00000073421 | 15.660 | 0.302 | 0.011 | 0.050 | 0.804 | protein coding gene | Influences antigen processing and presentation via MHC class II [59]. No known brain function. |
| Jpx | Jpx transcript, Xist activator (non-protein coding) | ENSMUSG00000097571 | 154.681 | 0.297 | 0.088 | 0.048 | 0.866 | lncRNA gene | Long non-coding RNA that induces Xist expression for X-inactivation [53,54]. |
| Dppa5a | developmental pluripotency associated 5A | ENSMUSG00000060461 | 18.336 | 0.234 | 0.041 | 0.139 | 0.392 | protein coding gene | Highly expressed in embryonic stem cells and germ cells and thought to play a role in pluripotency [60,61]. |
| Rhov | ras homolog family member V | ENSMUSG00000034226 | 498.992 | 0.220 | 0.098 | 0.021 | 0.936 | protein coding gene | Atypical, constitutively active GTPase implicated in neural crest development [62]. |
| Trim71 | tripartite motif-containing 71 | ENSMUSG00000079259 | 18.205 | 0.196 | 0.068 | 0.151 | 0.166 | protein coding gene | Plays important roles in embryonic neurogenesis [63] and postnatal ependymal cells [64]. |
| Rcn1 | reticulocalbin 1 | ENSMUSG00000005973 | 1336.228 | 0.189 | 0.083 | 0.143 | 0.127 | protein coding gene | Calcium binding protein located in the endoplasmic reticulum [65]. |
| Car11 | carbonic anhydrase 11 | ENSMUSG00000003273 | 2481.408 | 0.169 | 0.039 | 0.103 | 0.227 | protein coding gene | Homologous to Car10, potential extracellular binding partner of neurexin that influence synaptogenesis [66] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bonefas, K.M.; Vallianatos, C.N.; Raines, B.; Tronson, N.C.; Iwase, S. Sexually Dimorphic Alterations in the Transcriptome and Behavior with Loss of Histone Demethylase KDM5C. Cells 2023, 12, 637. https://doi.org/10.3390/cells12040637
Bonefas KM, Vallianatos CN, Raines B, Tronson NC, Iwase S. Sexually Dimorphic Alterations in the Transcriptome and Behavior with Loss of Histone Demethylase KDM5C. Cells. 2023; 12(4):637. https://doi.org/10.3390/cells12040637
Chicago/Turabian StyleBonefas, Katherine M., Christina N. Vallianatos, Brynne Raines, Natalie C. Tronson, and Shigeki Iwase. 2023. "Sexually Dimorphic Alterations in the Transcriptome and Behavior with Loss of Histone Demethylase KDM5C" Cells 12, no. 4: 637. https://doi.org/10.3390/cells12040637
APA StyleBonefas, K. M., Vallianatos, C. N., Raines, B., Tronson, N. C., & Iwase, S. (2023). Sexually Dimorphic Alterations in the Transcriptome and Behavior with Loss of Histone Demethylase KDM5C. Cells, 12(4), 637. https://doi.org/10.3390/cells12040637