
RpS3 Is Required for Spermatogenesis of Drosophila melanogaster




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Fly Strains and Breeding Conditions
2.2. Male Fertility Test
2.3. RNA Isolation and Gene Expression Assay
2.4. Immunofluorescence Staining
2.5. In Situ Hybridization
2.6. Transmission Electron Microscopy (TEM)
2.7. TUNEL Assay
2.8. Data Analysis
2.9. RNA Preparation and RNA-Seq
2.10. Bioinformatic Analysis of RNA-seq Data
3. Results
3.1. The Spatial-Temporal Expression Pattern of RpS3 in Drosophila
3.2. Knockdown of RpS3 Results in a Significant Decline in Drosophila Male Fertility
3.3. RpS3-Knockdown Testes Showed Normal Early Spermatogenesis
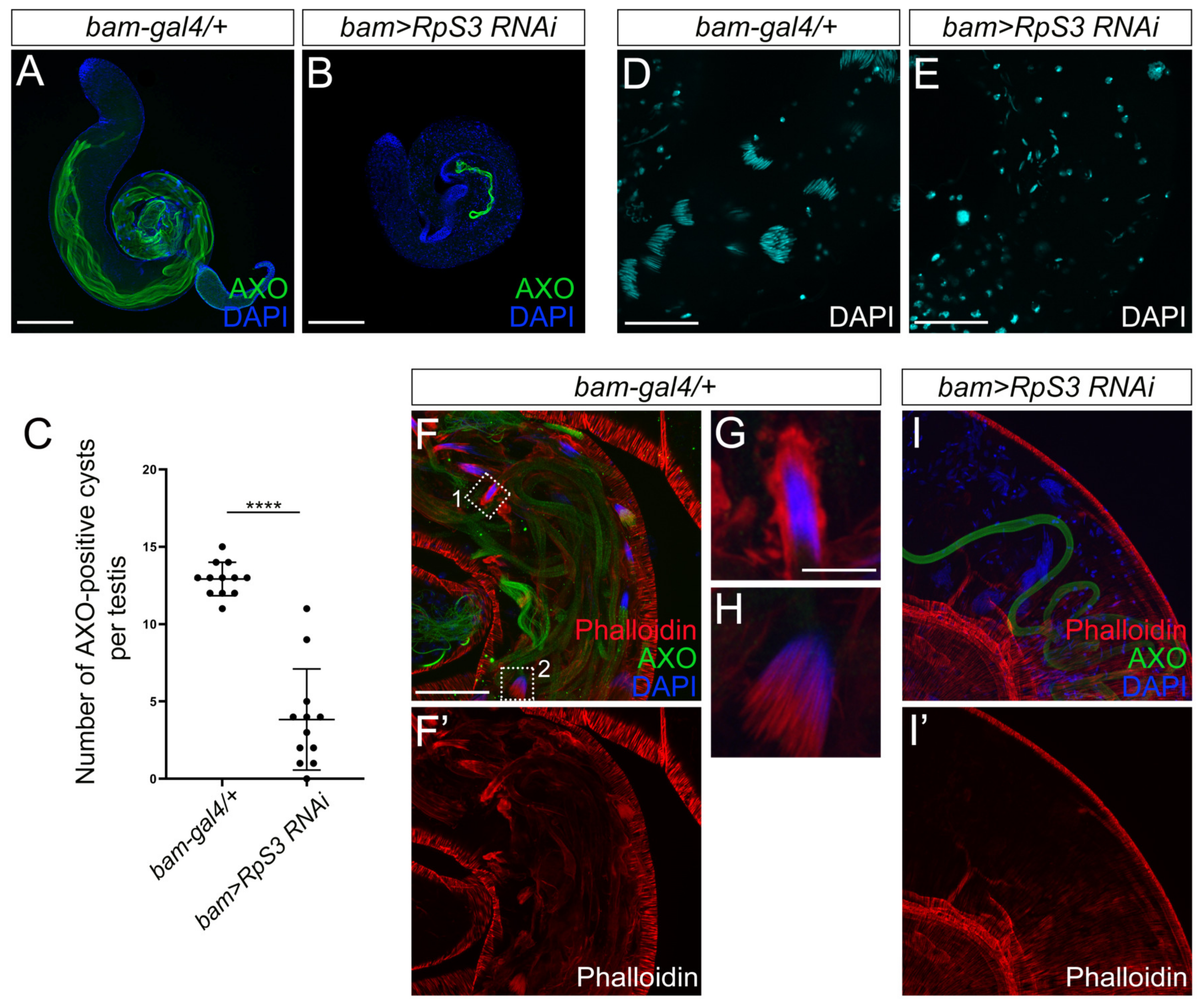
3.4. Spermatid Elongation and Individualization Are Disrupted in RpS3-Knockdown Testes
3.5. RpS3-Knockdown Testes Exhibit Mitochondrial Defects
3.6. Knockdown of RpS3 Induces Apoptosis in Fly Testes
3.7. Knockdown of RpS3 Alters Gene Expression Profiles in Fly Testes
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Norris, K.; Hopes, T.; Aspden, J.L. Ribosome heterogeneity and specialization in development. Wiley Interdiscip Rev. RNA 2021, 12, e1644. [Google Scholar] [CrossRef]
- Shi, Z.; Fujii, K.; Kovary, K.M.; Genuth, N.R.; Rost, H.L.; Teruel, M.N.; Barna, M. Heterogeneous Ribosomes Preferentially Translate Distinct Subpools of mRNAs Genome-wide. Mol. Cell 2017, 67, 71–83. [Google Scholar] [CrossRef] [PubMed]
- Kondrashov, N.; Pusic, A.; Stumpf, C.R.; Shimizu, K.; Hsieh, A.C.; Ishijima, J.; Shiroishi, T.; Barna, M. Ribosome-mediated specificity in Hox mRNA translation and vertebrate tissue patterning. Cell 2011, 145, 383–397. [Google Scholar] [CrossRef] [PubMed]
- Xue, S.; Tian, S.; Fujii, K.; Kladwang, W.; Das, R.; Barna, M. RNA regulons in Hox 5’ UTRs confer ribosome specificity to gene regulation. Nature 2015, 517, 33–38. [Google Scholar] [CrossRef] [PubMed]
- Ludwig, L.S.; Gazda, H.T.; Eng, J.C.; Eichhorn, S.W.; Thiru, P.; Ghazvinian, R.; George, T.I.; Gotlib, J.R.; Beggs, A.H.; Sieff, C.A.; et al. Altered translation of GATA1 in Diamond-Blackfan anemia. Nat. Med. 2014, 20, 748–753. [Google Scholar] [CrossRef]
- Wool, I.G. Extraribosomal functions of ribosomal proteins. Trends Biochem. Sci. 1996, 21, 164–165. [Google Scholar] [CrossRef]
- Kim, S.H.; Lee, J.Y.; Kim, J. Characterization of a wide range base-damage-endonuclease activity of mammalian rpS3. Biochem. Biophys. Res. Commun. 2005, 328, 962–967. [Google Scholar] [CrossRef]
- Kim, Y.; Kim, H.D.; Kim, J. Cytoplasmic ribosomal protein S3 (rpS3) plays a pivotal role in mitochondrial DNA damage surveillance. Biochim. Biophys. Acta 2013, 1833, 2943–2952. [Google Scholar] [CrossRef]
- Park, Y.J.; Kim, S.H.; Kim, T.S.; Lee, S.M.; Cho, B.S.; Seo, C.I.; Kim, H.D.; Kim, J. Ribosomal protein S3 associates with the TFIIH complex and positively regulates nucleotide excision repair. Cell Mol. Life Sci. 2021, 78, 3591–3606. [Google Scholar] [CrossRef]
- Yadavilli, S.; Mayo, L.D.; Higgins, M.; Lain, S.; Hegde, V.; Deutsch, W.A. Ribosomal protein S3: A multi-functional protein that interacts with both p53 and MDM2 through its KH domain. DNA Repair 2009, 8, 1215–1224. [Google Scholar] [CrossRef]
- Jang, C.-Y.; Lee, J.Y.; Kim, J. RpS3, a DNA repair endonuclease and ribosomal protein, is involved in apoptosis. FEBS Lett. 2004, 560, 81–85. [Google Scholar] [CrossRef]
- Wan, F.; Anderson, D.E.; Barnitz, R.A.; Snow, A.; Bidere, N.; Zheng, L.; Hegde, V.; Lam, L.T.; Staudt, L.M.; Levens, D.; et al. Ribosomal protein S3: A KH domain subunit in NF-kappaB complexes that mediates selective gene regulation. Cell 2007, 131, 927–939. [Google Scholar] [CrossRef]
- Wier, E.M.; Neighoff, J.; Sun, X.; Fu, K.; Wan, F. Identification of an N-terminal truncation of the NF-kappaB p65 subunit that specifically modulates ribosomal protein S3-dependent NF-kappaB gene expression. J. Biol. Chem. 2012, 287, 43019–43029. [Google Scholar] [CrossRef]
- Gao, X.; Wan, F.; Mateo, K.; Callegari, E.; Wang, D.; Deng, W.; Puente, J.; Li, F.; Chaussee, M.S.; Finlay, B.B.; et al. Bacterial effector binding to ribosomal protein s3 subverts NF-kappaB function. PLoS Pathog. 2009, 5, e1000708. [Google Scholar] [CrossRef]
- Saeboe-Larssen, S.; Lyamouri, M.; Merriam, J.; Oksvold, M.P.; Lambertsson, A. Ribosomal protein insufficiency and the minute syndrome in Drosophila: A dose-response relationship. Genetics 1998, 148, 1215–1224. [Google Scholar] [CrossRef]
- Fabian, L.; Brill, J.A. Drosophila spermiogenesis: Big things come from little packages. Spermatogenesis 2012, 2, 197–212. [Google Scholar] [CrossRef]
- Hopes, T.; Norris, K.; Agapiou, M.; McCarthy, C.G.P.; Lewis, P.A.; O’Connell, M.J.; Fontana, J.; Aspden, J.L. Ribosome heterogeneity in Drosophila melanogaster gonads through paralog-switching. Nucleic Acids Res. 2022, 50, 2240–2257. [Google Scholar] [CrossRef]
- Yu, J.; Lan, X.; Chen, X.; Yu, C.; Xu, Y.; Liu, Y.; Xu, L.; Fan, H.Y.; Tong, C. Protein synthesis and degradation are essential to regulate germline stem cell homeostasis in Drosophila testes. Development 2016, 143, 2930–2945. [Google Scholar] [CrossRef]
- Zheng, Y.; Bi, J.; Hou, M.Y.; Shen, W.; Zhang, W.; Ai, H.; Yu, X.Q.; Wang, Y.F. Ocnus is essential for male germ cell development in Drosophila melanogaster. Insect Mol. Biol. 2018, 27, 545–555. [Google Scholar] [CrossRef]
- Zheng, Y.; Ren, P.P.; Wang, J.L.; Wang, Y.F. Wolbachia-induced cytoplasmic incompatibility is associated with decreased Hira expression in male Drosophila. PLoS ONE 2011, 6, e19512. [Google Scholar] [CrossRef]
- Mao, X.; Cai, T.; Olyarchuk, J.G.; Wei, L. Automated genome annotation and pathway identification using the KEGG Orthology (KO) as a controlled vocabulary. Bioinformatics 2005, 21, 3787–3793. [Google Scholar] [CrossRef] [PubMed]
- Young, M.D.; Wakefield, M.J.; Smyth, G.K.; Oshlack, A. Gene ontology analysis for RNA-seq: Accounting for selection bias. Genome Biol. 2010, 11, R14. [Google Scholar] [CrossRef] [PubMed]
- Carbonell, A.; Perez-Montero, S.; Climent-Canto, P.; Reina, O.; Azorin, F. The Germline Linker Histone dBigH1 and the Translational Regulator Bam Form a Repressor Loop Essential for Male Germ Stem Cell Differentiation. Cell Rep. 2017, 21, 3178–3189. [Google Scholar] [CrossRef] [PubMed]
- Kibanov, M.V.; Kotov, A.A.; Olenina, L.V. Multicolor fluorescence imaging of whole-mount Drosophila testes for studying spermatogenesis. Anal. Biochem. 2013, 436, 55–64. [Google Scholar] [CrossRef]
- Yu, J.; Zheng, Q.; Li, Z.; Wu, Y.; Fu, Y.; Wu, X.; Lin, D.; Shen, C.; Zheng, B.; Sun, F. CG6015 controls spermatogonia transit-amplifying divisions by epidermal growth factor receptor signaling in Drosophila testes. Cell Death Dis. 2021, 12, 491. [Google Scholar] [CrossRef]
- Bre, M.H.; Redeker, V.; Quibell, M.; Darmanaden-Delorme, J.; Bressac, C.; Cosson, J.; Huitorel, P.; Schmitter, J.M.; Rossler, J.; Johnson, T.; et al. Axonemal tubulin polyglycylation probed with two monoclonal antibodies: Widespread evolutionary distribution, appearance during spermatozoan maturation and possible function in motility. J. Cell Sci. 1996, 109 Pt 4, 727–738. [Google Scholar] [CrossRef]
- Noguchi, T.; Miller, K.G. A role for actin dynamics in individualization during spermatogenesis in Drosophila melanogaster. Development 2003, 130, 1805–1816. [Google Scholar] [CrossRef]
- Desai, B.S.; Shirolikar, S.; Ray, K. F-actin-based extensions of the head cyst cell adhere to the maturing spermatids to maintain them in a tight bundle and prevent their premature release in Drosophila testis. BMC Biol. 2009, 7, 19. [Google Scholar] [CrossRef]
- Vedelek, V.; Laurinyecz, B.; Kovacs, A.L.; Juhasz, G.; Sinka, R. Testis-Specific Bb8 Is Essential in the Development of Spermatid Mitochondria. PLoS ONE 2016, 11, e0161289. [Google Scholar] [CrossRef]
- Metzendorf, C.; Lind, M.I. Drosophila mitoferrin is essential for male fertility: Evidence for a role of mitochondrial iron metabolism during spermatogenesis. BMC Dev. Biol. 2010, 10, 68. [Google Scholar] [CrossRef]
- Tokuyasu, K.T.; Peacock, W.J.; Hardy, R.W. Dynamics of spermiogenesis in Drosophila melanogaster. I. Individualization process. Z. Zellforsch. Mikrosk. Anat. 1972, 124, 479–506. [Google Scholar] [CrossRef]
- Noguchi, T.; Koizumi, M.; Hayashi, S. Sustained elongation of sperm tail promoted by local remodeling of giant mitochondria in Drosophila. Curr. Biol. 2011, 21, 805–814. [Google Scholar] [CrossRef]
- Senos Demarco, R.; Uyemura, B.S.; D’Alterio, C.; Jones, D.L. Mitochondrial fusion regulates lipid homeostasis and stem cell maintenance in the Drosophila testis. Nat. Cell Biol. 2019, 21, 710–720. [Google Scholar] [CrossRef]
- Arama, E.; Bader, M.; Rieckhof, G.E.; Steller, H. A ubiquitin ligase complex regulates caspase activation during sperm differentiation in Drosophila. PLoS Biol. 2007, 5, e251. [Google Scholar] [CrossRef]
- Goldstein, L.S.; Hardy, R.W.; Lindsley, D.L. Structural genes on the Y chromosome of Drosophila melanogaster. Proc. Natl. Acad. Sci. USA 1982, 79, 7405–7409. [Google Scholar] [CrossRef]
- Witt, E.; Benjamin, S.; Svetec, N.; Zhao, L. Testis single-cell RNA-seq reveals the dynamics of de novo gene transcription and germline mutational bias in Drosophila. eLife 2019, 8. [Google Scholar] [CrossRef]
- Martin, E.T.; Blatt, P.; Nguyen, E.; Lahr, R.; Selvam, S.; Yoon, H.A.M.; Pocchiari, T.; Emtenani, S.; Siekhaus, D.E.; Berman, A.; et al. A translation control module coordinates germline stem cell differentiation with ribosome biogenesis during Drosophila oogenesis. Dev. Cell 2022, 57, 883–900 e810. [Google Scholar] [CrossRef]
- Fang, Y.; Zong, Q.; He, Z.; Liu, C.; Wang, Y.F. Knockdown of RpL36 in testes impairs spermatogenesis in Drosophila melanogaster. J. Exp. Zool. B Mol. Dev. Evol 2021, 336, 417–430. [Google Scholar] [CrossRef]
- Yu, J.; Yan, Y.; Luan, X.; Qiao, C.; Liu, Y.; Zhao, D.; Xie, B.; Zheng, Q.; Wang, M.; Chen, W.; et al. Srlp is crucial for the self-renewal and differentiation of germline stem cells via RpL6 signals in Drosophila testes. Cell Death Dis. 2019, 10, 294. [Google Scholar] [CrossRef]
- Yu, J.; Luan, X.; Yan, Y.; Qiao, C.; Liu, Y.; Zhao, D.; Xie, B.; Zheng, Q.; Wang, M.; Chen, W.; et al. Small ribonucleoprotein particle protein SmD3 governs the homeostasis of germline stem cells and the crosstalk between the spliceosome and ribosome signals in Drosophila. FASEB J. 2019, 33, 8125–8137. [Google Scholar] [CrossRef]
- Varuzhanyan, G.; Chen, H.; Rojansky, R.; Ladinsky, M.S.; McCaffery, J.M.; Chan, D.C. Mitochondrial fission factor (Mff) is required for organization of the mitochondrial sheath in spermatids. Biochim. Biophys. Acta Gen. Subj. 2021, 1865, 129845. [Google Scholar] [CrossRef] [PubMed]
- Bazinet, C.; Rollins, J.E. Rickettsia-like mitochondrial motility in Drosophila spermiogenesis. Evol. Dev. 2003, 5, 379–385. [Google Scholar] [CrossRef] [PubMed]
- Lei, Y.; Zhang, X.; Xu, Q.; Liu, S.; Li, C.; Jiang, H.; Lin, H.; Kong, E.; Liu, J.; Qi, S.; et al. Autophagic elimination of ribosomes during spermiogenesis provides energy for flagellar motility. Dev. Cell 2021, 56, 2313–2328. [Google Scholar] [CrossRef] [PubMed]
- Aravindan, R.G.; Kirn-Safran, C.B.; Smith, M.A.; Martin-DeLeon, P.A. Ultrastructural changes and asthenozoospermia in murine spermatozoa lacking the ribosomal protein L29/HIP gene. Asian J. 2014, 16, 925–926. [Google Scholar] [CrossRef]
- Jiang, L.; Li, T.; Zhang, X.; Zhang, B.; Yu, C.; Li, Y.; Fan, S.; Jiang, X.; Khan, T.; Hao, Q.; et al. RPL10L Is required for male meiotic division by compensating for RPL10 during meiotic sex chromosome inactivation in mice. Curr. Biol. 2017, 27, 1498–1505. [Google Scholar] [CrossRef]
- Zou, Q.; Yang, L.; Shi, R.; Qi, Y.; Zhang, X.; Qi, H. Proteostasis regulated by testis-specific ribosomal protein RPL39L maintains mouse spermatogenesis. iScience 2021, 24, 103396. [Google Scholar] [CrossRef]
- Bansal, S.K.; Gupta, N.; Sankhwar, S.N.; Rajender, S. Differential genes expression between fertile and infertile spermatozoa revealed by transcriptome analysis. PLoS ONE 2015, 10, e0127007. [Google Scholar] [CrossRef]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fang, Y.; Zhang, F.; Zhan, Y.; Lu, M.; Xu, D.; Wang, J.; Li, Q.; Zhao, L.; Su, Y. RpS3 Is Required for Spermatogenesis of Drosophila melanogaster. Cells 2023, 12, 573. https://doi.org/10.3390/cells12040573
Fang Y, Zhang F, Zhan Y, Lu M, Xu D, Wang J, Li Q, Zhao L, Su Y. RpS3 Is Required for Spermatogenesis of Drosophila melanogaster. Cells. 2023; 12(4):573. https://doi.org/10.3390/cells12040573
Chicago/Turabian StyleFang, Yang, Fengchao Zhang, Yunhui Zhan, Meina Lu, Di Xu, Jiajia Wang, Qiujun Li, Long Zhao, and Ying Su. 2023. "RpS3 Is Required for Spermatogenesis of Drosophila melanogaster" Cells 12, no. 4: 573. https://doi.org/10.3390/cells12040573
APA StyleFang, Y., Zhang, F., Zhan, Y., Lu, M., Xu, D., Wang, J., Li, Q., Zhao, L., & Su, Y. (2023). RpS3 Is Required for Spermatogenesis of Drosophila melanogaster. Cells, 12(4), 573. https://doi.org/10.3390/cells12040573