

Brain Noradrenergic Innervation Supports the Development of Parkinson’s Tremor: A Study in a Reserpinized Rat Model

 ,
, 
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Drug Treatment Procedures and Dose Selection
2.3. Behavioral Evaluations and Kinematic Analysis
2.4. Tissue Preparation, Immunolabeling and Quantitative Analyses
3. Statistical Analysis
4. Results
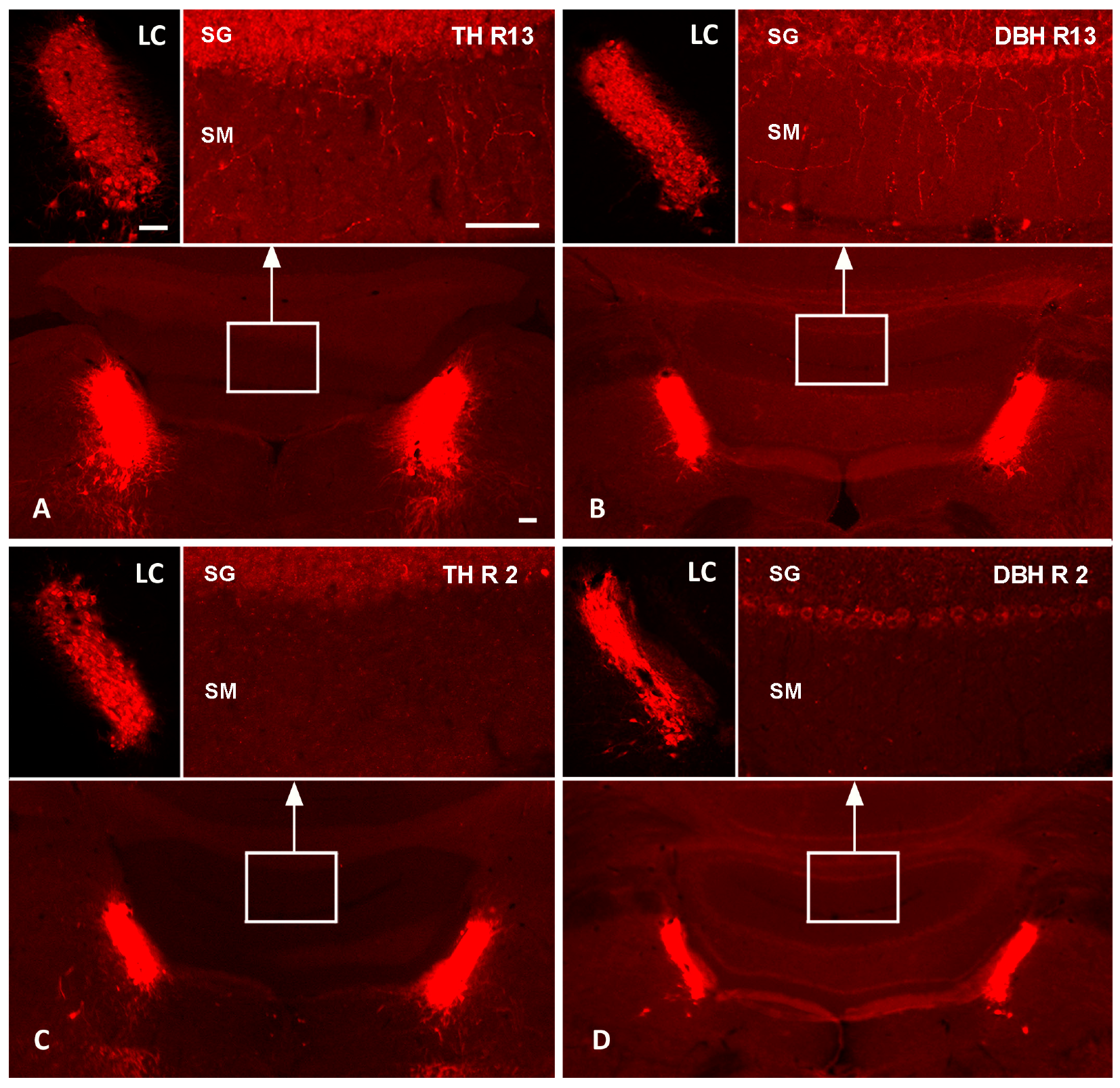
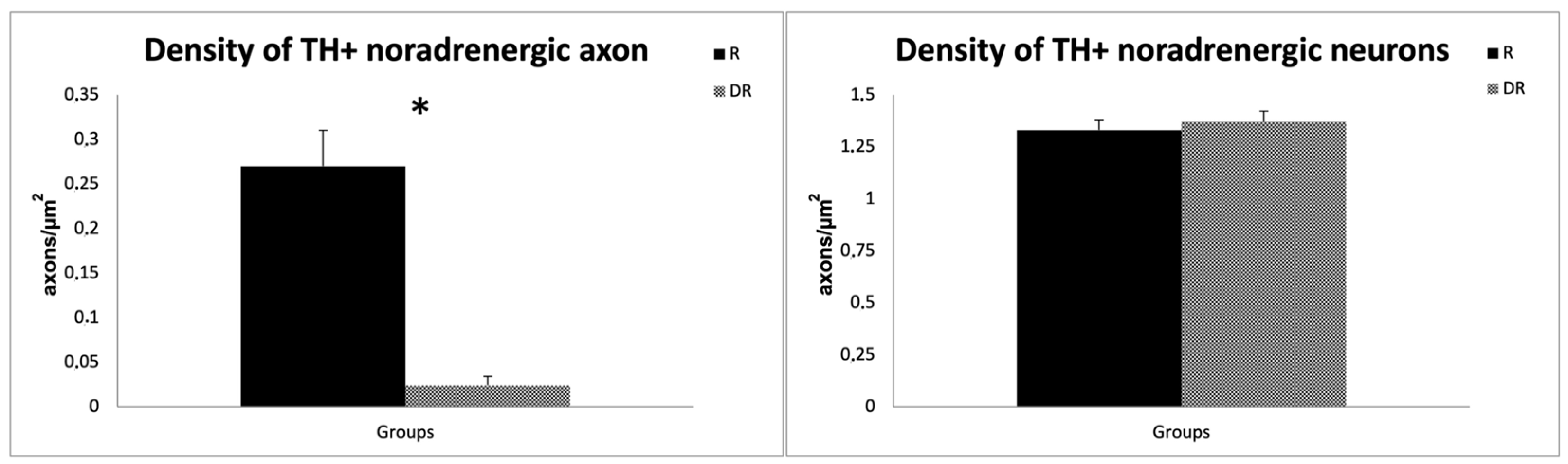
4.1. Assessment of DSP-4-Induced Noradrenergic Denervation
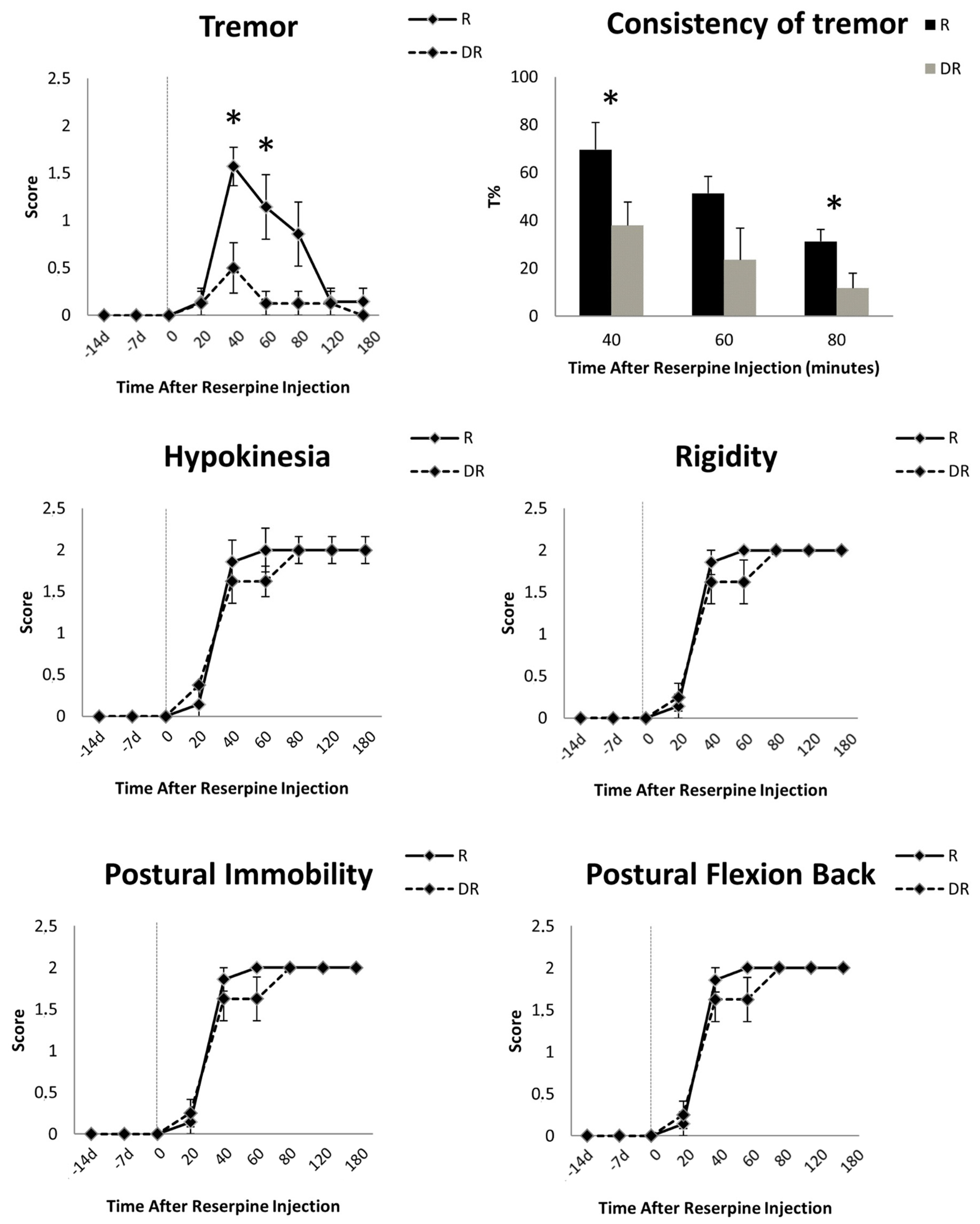
4.2. Visual Scores
4.3. Accelerometer Tremor Measurements
5. Discussion
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Chen, W.; Hopfner, F.; Becktepe, J.S.; Deuschl, G. Rest tremor revisited: Parkinson’s disease and other disorders. Transl. Neurodegener. 2017, 6, 16. [Google Scholar] [CrossRef] [PubMed]
- Hou, M.; Mao, X.; Hou, X.; Li, K. Stigma and Associated Correlates of Elderly Patients with Parkinson’s Disease. Front. Psychiatry 2021, 12, 708960. [Google Scholar] [CrossRef] [PubMed]
- Dirkx, M.F.; Bologna, M. The pathophysiology of Parkinson’s disease tremor. J. Neurol. Sci. 2022, 435, 120196. [Google Scholar] [CrossRef] [PubMed]
- Isaias, I.U.; Marzegan, A.; Pezzoli, G.; Marotta, G.; Canesi, M.; Biella, G.E.M.; Volkmann, J.; Cavallari, P. A role for locus coeruleus in Parkinson tremor. Front. Hum. Neurosci. 2012, 5, 179. [Google Scholar] [CrossRef]
- Isaias, I.U.; Benti, R.; Cilia, R.; Canesi, M.; Marotta, G.; Gerundini, P.; Pezzoli, G.; Antonini, A. [123I]FP-CIT striatal binding in early Parkinson’s disease patients with tremor vs. akinetic-rigid onset. NeuroReport 2007, 18, 1499–1502. [Google Scholar] [CrossRef]
- Benamer, T.S.; Patterson, J.; Grosset, D.G.; Booij, J.; De Bruin, K.; Van Royen, E.; Speelman, J.D.; Horstink, M.H.; Sips, H.J.; A Dierckx, R.; et al. Accurate differentiation of parkinsonism and essential tremor using visual assessment of [123I]-FP-CIT SPECT imaging: The [123I]-FP-CIT study group. Mov. Disord. 2000, 15, 503–510. [Google Scholar]
- Zach, H.; Dirkx, M.F.; Roth, D.; Pasman, J.W.; Bloem, B.R.; Helmich, R.C. Dopamine-responsive and dopamine-resistant resting tremor in Parkinson disease. Neurology 2020, 95, e1461–e1470. [Google Scholar] [CrossRef]
- Dirkx, M.F.; Zach, H.; Van Nuland, A.; Bloem, B.R.; Toni, I.; Helmich, R.C. Cerebral differences between dopamine-resistant and dopamine-responsive Parkinson’s tremor. Brain 2019, 142, 3144–3157. [Google Scholar] [CrossRef]
- Deuschl, G.; Bain, P.; Brin, M. Consensus Statement of the Movement Disorder Society on Tremor. Mov. Disord. 2008, 13, 2–23. [Google Scholar] [CrossRef]
- Dirkx, M.F.; Zach, H.; Van Nuland, A.J.; Bloem, B.R.; Toni, I.; Helmich, R.C. Cognitive load amplifies Parkinson’s tremor through excitatory network influences onto the thalamus. Brain 2020, 143, 1498–1511. [Google Scholar] [CrossRef]
- Pasquini, J.; Ceravolo, R. The Molecular Neuroimaging of Tremor. Curr. Neurol. Neurosci. Rep. 2021, 21, 74. [Google Scholar] [CrossRef] [PubMed]
- Prange, S.; Theis, H.; Banwinkler, M.; van Eimeren, T. Molecular Imaging in Parkinsonian Disorders—What’s New and Hot? Brain Sci. 2022, 12, 1146. [Google Scholar] [CrossRef] [PubMed]
- Paulus, W.; Jellinger, K. The Neuropathologic Basis of Different Clinical Subgroups of Parkinson’s Disease. J. Neuropathol. Exp. Neurol. 1991, 50, 743–755. [Google Scholar] [CrossRef]
- Benarroch, E.E. Locus coeruleus. Cell Tissue Res. 2018, 373, 221–232. [Google Scholar] [CrossRef] [PubMed]
- Mure, H.; Hirano, S.; Tang, C.C.; Isaias, I.U.; Antonini, A.; Ma, Y.; Dhawan, V.; Eidelberg, D. Parkinson’s disease tremor-related metabolic network: Characterization, progression, and treatment effects. NeuroImage 2011, 54, 1244–1253. [Google Scholar] [CrossRef]
- Kinnerup, M.B.; Sommerauer, M.; Damholdt, M.F.; Schaldemose, J.L.; Ismail, R.; Terkelsen, A.J.; Stær, K.; Hansen, A.; Fedorova, T.D.; Knudsen, K.; et al. Preserved noradrenergic function in Parkinson’s disease patients with rest tremor. Neurobiol. Dis. 2021, 152, 105295. [Google Scholar] [CrossRef]
- Marsden, C.D.; Owen, D.A.L. Mechanisms underlying emotional variation in parkinsonian tremor. Neurology 1967, 17, 711. [Google Scholar] [CrossRef]
- Jiménez-Jiménez, F.J.; Alonso-Navarro, H.; García-Martín, E.; Agúndez, J.A.G. Neurochemical Features of Rem Sleep Behaviour Disorder. J. Pers. Med. 2021, 11, 880. [Google Scholar] [CrossRef]
- Colpaert, F. Pharmacological characteristics of tremor, rigidity and hypokinesia induced by reserpine in rat. Neuropharmacology 1987, 26, 1431–1440. [Google Scholar] [CrossRef]
- Carlsson, A.; Lindqvist, M.; Magnusson, T. 3,4-Dihydroxyphenylalanine and 5-hydroxytryptophan as reserpine antagonists. Nature 1957, 180, 1200. [Google Scholar] [CrossRef]
- Carlsson, A. The occurrence, distribution and physiological role of catecholamines in the nervous system. Pharmacol. Rev. 1959, 11, 490. [Google Scholar] [PubMed]
- Jonsson, G.; Hallman, H.; Ponzio, F.; Ross, S. DSP4 (N-(2-chloroethyl)-N-ethyl-2-bromobenzylamine)—A useful denervation tool for central and peripheral noradrenaline neurons. Eur. J. Pharmacol. 1981, 72, 173–188. [Google Scholar] [CrossRef] [PubMed]
- Flach, F.F. Clinical effectiveness of reserpine. Ann. N. Y. Acad. Sci. 1955, 61, 161–166. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, V.S.; Santos, J.R.; Leão, A.H.; Medeiros, A.M.; Melo, T.G.; Izídio, G.S.; Cabral, A.; Ribeiro, R.A.; Abílio, V.C.; Ribeiro, A.M.; et al. Repeated treatment with a low dose of reserpine as a progressive model of Parkinson’s disease. Behav. Brain Res. 2012, 231, 154–163. [Google Scholar] [CrossRef]
- Gerlach, M.; Riederer, P. Animal models of Parkinson’s disease: An empirical comparison with the phenomenology of the disease in man. J. Neural Transm. 1996, 103, 987–1041. [Google Scholar] [CrossRef]
- Tieu, K. A guide to neurotoxic animal models of Parkinson’s disease. Cold Spring Harb. Perspect. Med. 2011, 1, a009316. [Google Scholar] [CrossRef]
- Caudle, W.M.; Colebrooke, R.E.; Emson, P.C.; Miller, G.W. Altered vesicular dopamine storage in Parkinson’s disease: A premature demise. Trends Neurosci. 2008, 31, 303–308. [Google Scholar] [CrossRef]
- Dröge, W. Free Radicals in the Physiological Control of Cell Function. Physiol. Rev. 2002, 82, 47–95. [Google Scholar] [CrossRef]
- Tsang, A.H.; Chung, K.K. Oxidative and nitrosative stress in Parkinson’s disease. Biochim. Biophys. Acta (BBA)-Mol. Basis Dis. 2009, 1792, 643–650. [Google Scholar] [CrossRef]
- Fritschy, J.-M.; Grzanna, R. Immunohistochemical analysis of the neurotoxic effects of DSP-4 identifies two populations of noradrenergic axon terminals. Neuroscience 1989, 30, 181–197. [Google Scholar] [CrossRef]
- Grzanna, R.; Berger, U.; Fritschy, J.M.; Geffard, M. Acute action of DSP-4 on central norepinephrine axons: Biochemical and immunohistochemical evidence for differential effects. J. Histochem. Cytochem. 1989, 37, 1435–1442. [Google Scholar] [CrossRef] [PubMed]
- Zaczek, R.; Fritschy, J.-M.; Culp, S.; De Souza, E.B.; Grzanna, R. Differential effects of DSP-4 on noradrenaline axons in cerebral cortex and hypothalamus may reflect heterogeneity of noradrenaline uptake sites. Brain Res. 1990, 522, 308–314. [Google Scholar] [CrossRef] [PubMed]
- Alachkar, A.; Brotchie, J.M.; Jones, O.T. Locomotor response to l-DOPA in reserpine-treated rats following central inhibition of aromatic l-amino acid decarboxylase: Further evidence for non-dopaminergic actions of l-DOPA and its metabolites. Neurosci. Res. 2010, 68, 44–50. [Google Scholar] [CrossRef] [PubMed]
- Delaville, C.; Navailles, S.; Benazzouz, A. Effects of noradrenaline and serotonin depletions on the neuronal activity of globus pallidus and substantia nigra pars reticulata in experimental parkinsonism. Neuroscience 2011, 202, 424–433. [Google Scholar] [CrossRef]
- Asan, E.; Langenhan, T.; Holtmann, B.; Bock, H.; Sendtner, M.; Carroll, P. Ciliary neurotrophic factor in the olfactory bulb of rats and mice. Neuroscience 2003, 120, 99–112. [Google Scholar] [CrossRef]
- Asan, E.; Yilmazer-Hanke, D.; Eliava, M.; Hantsch, M.; Lesch, K.-P.; Schmitt, A. The Corticotropin-Releasing Factor (CRF)-system and monoaminergic afferents in the central amygdala: Investigations in different mouse strains and comparison with the rat. Neuroscience 2005, 131, 953–967. [Google Scholar] [CrossRef]
- Helmich, R.C.; Janssen, M.J.R.; Oyen, W.J.G.; Bloem, B.R.; Toni, I. Pallidal dysfunction drives a cerebellothalamic circuit into Parkinson tremor. Ann. Neurol. 2011, 69, 269–281. [Google Scholar] [CrossRef]
- Hirano, S.; Sugiyama, A.; Arai, K. Noradrenergic Pathway to the Cerebellum: The Study Must Go on. Cerebellum 2022. Online ahead of print. [Google Scholar] [CrossRef]
- Wu, T.; Hallett, M. The cerebellum in Parkinson’s disease. Brain 2013, 136, 696–709. [Google Scholar] [CrossRef]
- Lauro, P.M.; Lee, S.; Akbar, U.; Asaad, W.F. Subthalamic–Cortical Network Reorganization during Parkinson’s Tremor. J. Neurosci. 2021, 41, 9844–9858. [Google Scholar] [CrossRef]
- Helmich, R.C.; Lehéricy, S. Dying-back of ascending noradrenergic projections in Parkinson’s disease. Brain 2021, 144, 2562–2564. [Google Scholar] [CrossRef] [PubMed]
- Samuels, E.R.; Szabadi, E. Functional Neuroanatomy of the Noradrenergic Locus Coeruleus: Its Roles in the Regulation of Arousal and Autonomic Function Part I: Principles of Functional Organisation. Curr. Neuropharmacol. 2008, 6, 235–253. [Google Scholar] [CrossRef] [PubMed]
- Isaias, I.U.; Marotta, G.; Pezzoli, G.; Sabri, O.; Schwarz, J.; Crenna, P.; Classen, J.; Cavallari, P. Enhanced catecholamine transporter binding in the locus coeruleus of patients with early Parkinson disease. BMC Neurol. 2011, 11, 88. [Google Scholar] [CrossRef]
- Doppler, C.E.J.; Kinnerup, M.B.; Brune, C.; Farrher, E.; Betts, M.; Fedorova, T.D.; Schaldemose, J.L.; Knudsen, K.; Ismail, R.; Seger, A.D.; et al. Regional locus coeruleus degeneration is uncoupled from noradrenergic terminal loss in Parkinson’s disease. Brain 2021, 144, 2732–2744. [Google Scholar] [CrossRef] [PubMed]
- Foffani, G.; Obeso, J.A. A Cortical Pathogenic Theory of Parkinson’s Disease. Neuron 2018, 99, 1116–1128. [Google Scholar] [CrossRef] [PubMed]
- Pasquini, J.; Ceravolo, R.; Qamhawi, Z.; Lee, J.-Y.; Deuschl, G.; Brooks, D.J.; Bonuccelli, U.; Pavese, N. Progression of tremor in early stages of Parkinson’s disease: A clinical and neuroimaging study. Brain 2018, 141, 811–821. [Google Scholar] [CrossRef]
- Ross, S.B.; Stenfors, C. DSP4, a Selective Neurotoxin for the Locus Coeruleus Noradrenergic System. A Review of Its Mode of Action. Neurotox. Res. 2014, 27, 15–30. [Google Scholar] [CrossRef]
- Price, C.J.; Warburton, E.A.; Moore, C.J.; Frackowiak, R.S.; Friston, K.J. Dynamic diaschisis: Anatomically remote and context-sensitive human brain lesions. J. Cogn. Neurosci. 2001, 13, 419–429. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pozzi, N.G.; Bolzoni, F.; Biella, G.E.M.; Pezzoli, G.; Ip, C.W.; Volkmann, J.; Cavallari, P.; Asan, E.; Isaias, I.U. Brain Noradrenergic Innervation Supports the Development of Parkinson’s Tremor: A Study in a Reserpinized Rat Model. Cells 2023, 12, 2529. https://doi.org/10.3390/cells12212529
Pozzi NG, Bolzoni F, Biella GEM, Pezzoli G, Ip CW, Volkmann J, Cavallari P, Asan E, Isaias IU. Brain Noradrenergic Innervation Supports the Development of Parkinson’s Tremor: A Study in a Reserpinized Rat Model. Cells. 2023; 12(21):2529. https://doi.org/10.3390/cells12212529
Chicago/Turabian StylePozzi, Nicoló Gabriele, Francesco Bolzoni, Gabriele Eliseo Mario Biella, Gianni Pezzoli, Chi Wang Ip, Jens Volkmann, Paolo Cavallari, Esther Asan, and Ioannis Ugo Isaias. 2023. "Brain Noradrenergic Innervation Supports the Development of Parkinson’s Tremor: A Study in a Reserpinized Rat Model" Cells 12, no. 21: 2529. https://doi.org/10.3390/cells12212529
APA StylePozzi, N. G., Bolzoni, F., Biella, G. E. M., Pezzoli, G., Ip, C. W., Volkmann, J., Cavallari, P., Asan, E., & Isaias, I. U. (2023). Brain Noradrenergic Innervation Supports the Development of Parkinson’s Tremor: A Study in a Reserpinized Rat Model. Cells, 12(21), 2529. https://doi.org/10.3390/cells12212529