


Single-Cell Transcriptomics of Mtb/HIV Co-Infection
Abstract
1. Introduction
Limitations and Challenges of LTBI Screening in HIV Patients
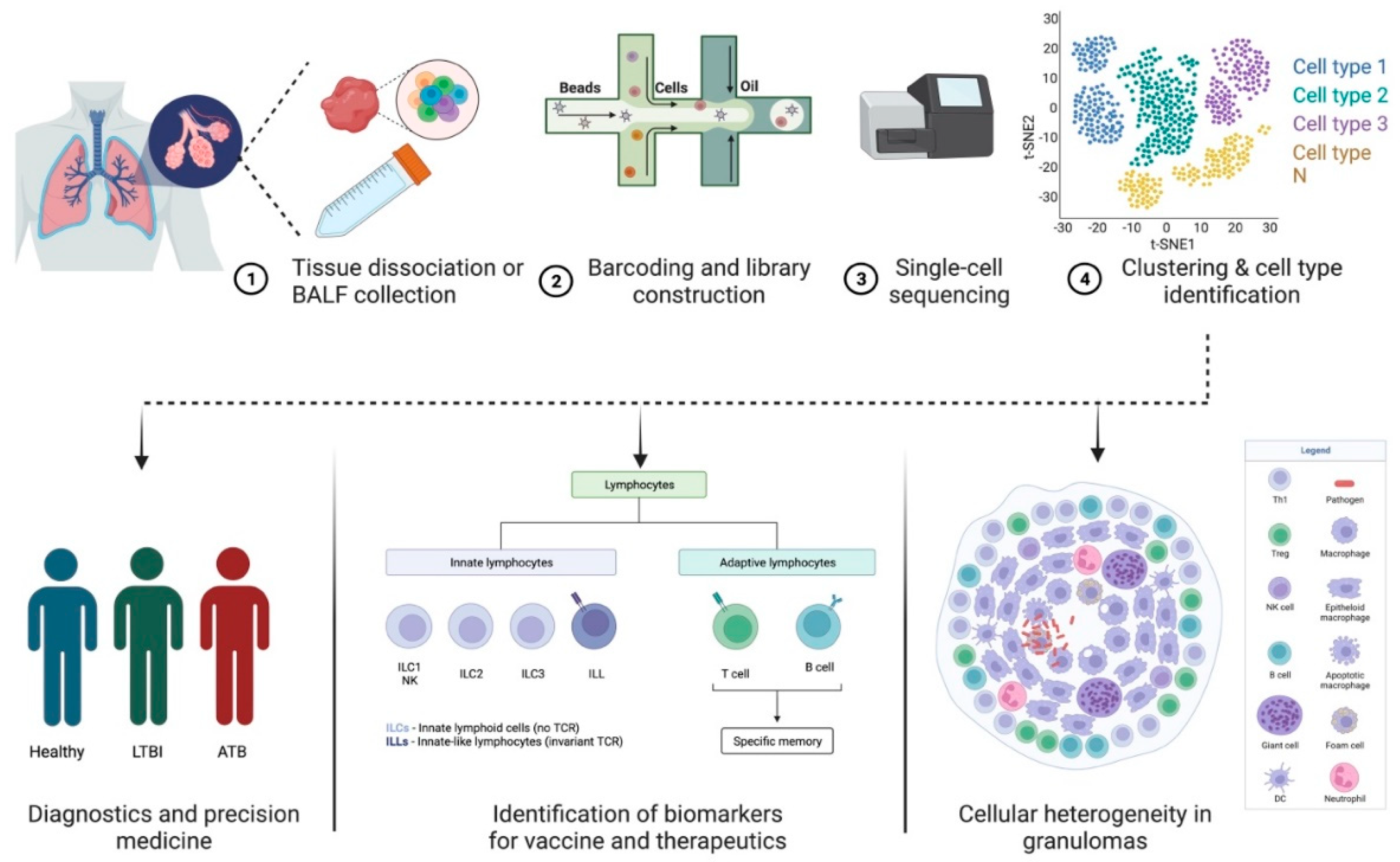
2. scRNA-Seq—A Tool to Study Host–Pathogen Interactions and Biosignatures in TB
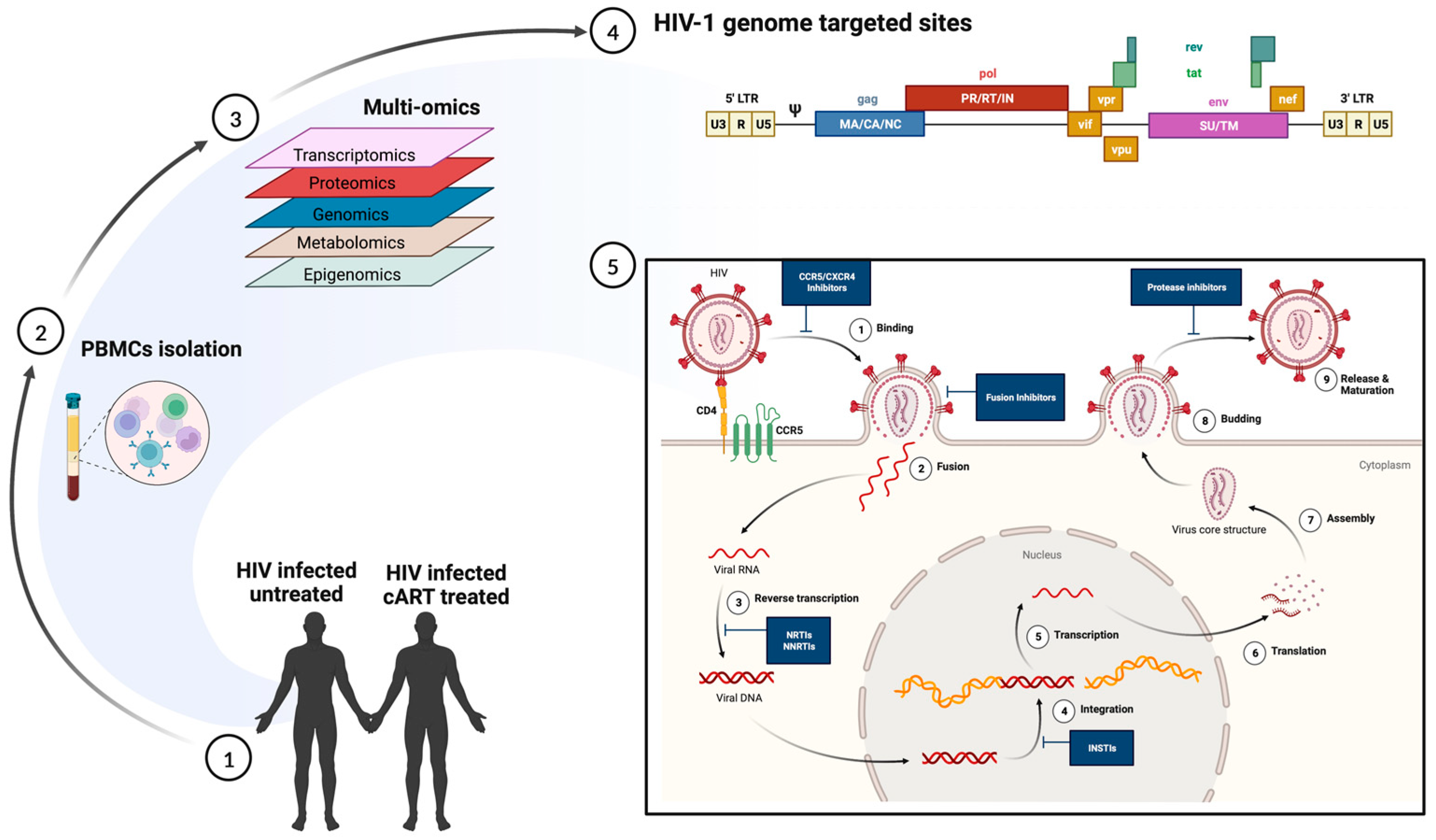
An Insight into the Mechanisms of HIV Latency Using scRNA-Seq and the Effect of Co-Infections on HIV-Reservoir
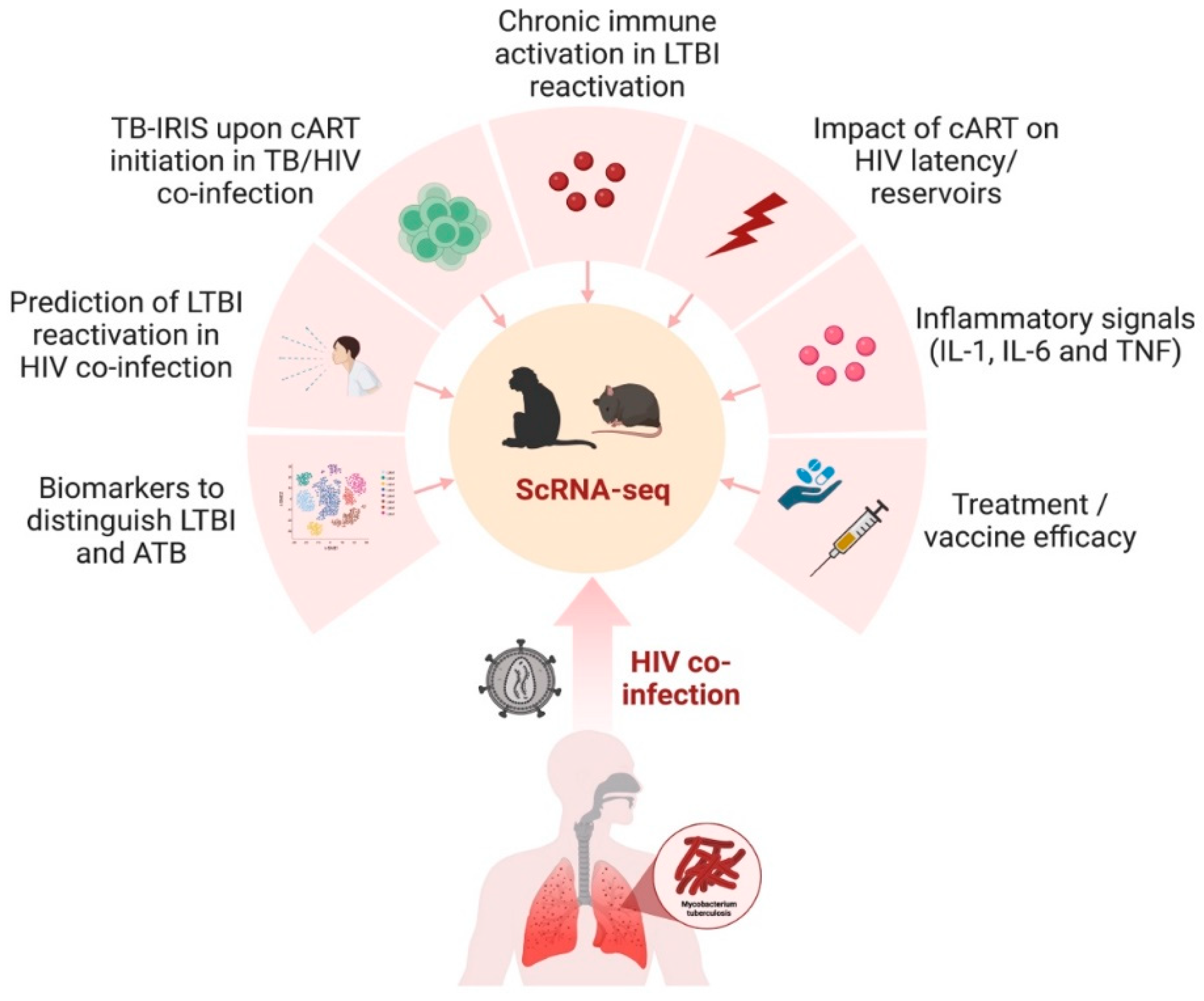
3. Predicting Disease Progression in TB and HIV Using scRNA-Seq

{kind=link}
{kind=link}
{kind=link}
| Study | Technique | Observation | Ref. |
|---|---|---|---|
| Defining HIV-permissive cells | scRNA-seq | TCR stimulation mainly contributes to transcriptional heterogeneity in CD4+ T cells, which regulates HIV transcription. | [120] |
| Immune responses during hyperacute HIV infection | scRNA-seq on peripheral blood mononuclear cells from four untreated individuals before and longitudinally during acute infection | Immune cell responses to HIV infection within the first weeks of infection, such as proliferating natural killer cells, which potentially may be associated with viral control | [30] |
| Restoration of T cell function after ART | scRNA-seq on peripheral T cells on chronic HIV-infected treatment-naïve or ART-treated patients | Significant loss of naïve T cells, prolonged inflammation, and increased response to interferon-α in treatment-naïve patients, partially restored by ART. Granulosyn expressing CD4+ and CD8+ Effector cell clusters correlated with poor immune restoration | [123] |
| Immune Exhaustion in Chronic HIV Infection | scRNA-seq on peripheral blood mononuclear cells from HIV patients and healthy subjects | An inhibitory receptor KLRG1 was identified in an HIV-1 specific exhausted CD8+ T cell population expressing KLRG1, TIGIT, and Tbetdim Eomeshi markers | [124] |
| Immune Reconstitution failure in (HAART)-treated HIV patients | scRNA-seq and scATAC-seq analysis of peripheral blood mononuclear cells (PBMCs) derived from immune non-responder [127] and responder (IR) | Low expression of mucosal-associated invariant T (MAIT) cells in INRs, which exhibited transcriptional profiles associated with impaired mitochondrial function and apoptosis signaling | [125] |
| Cellular transcriptome changes induced by active HIV replication in macrophages | An in vitro system to model HIV-1 infection of macrophages and single-cell RNA sequencing (scRNA-seq) to compare the transcriptomes of uninfected cells, cells harboring pre-integration complexes (PIC), and those containing integrated provirus and making late HIV proteins | NFkB- and AP-1-promoted transcription characterize PIC cell transcriptomes, while E2F family transcription products distinguish transcriptomes of cells transcribing from provirus | [121] |
| Differential virus reactivation potential | Primary CD4+ T-cell model expression HIV green fluorescent protein (GFP); scRNA seq | Global transcriptomic profiles of cells with reactivated HIV showed higher cellular activation and metabolic activity | [28] |
| Characterize latent cells reactivated by a single round of stimulation | Latent cell capture; LURE. Purified CD4+ T cells from peripheral blood of ART-treated patients activated by LRS and sorted based on the expression of HIV-env protein and HIV-gag mRNA | CD4+ T cells harboring proviruses with identical Env sequences also showed identical TCRs. Reservoir cells arise by clonal expansion, and the reservoir is maintained by balanced cell division and cell death. Reactivated latent cells express a distinct transcriptional program that suppresses HIV-1 transcription, which helps them survive | [78] |
| Characterize reactivated cells within 24 h of latency reversal | Sortseq; stimulated peripheral blood CD4+ T cells from ART-treated, virally suppressed patients | HIV+ cells enriched in TH1 phenotype upregulate cellular factors that support HIV transcription and promote cellular survival. HIV promoter drives high aberrant host gene transcription downstream of the integration site | [79] |
| Transcriptome of quiescent reservoir memory CD4+ T cells harboring intact HIV provirus | Quiescent reservoir memory CD4+ T cells enriched using their unique TCR as a molecule, identified using scRNA-seq | HIV–proviral integration and latency did not induce a specific transcriptional program | [85] |
| Characterize HIV reservoir cells after suppressive ART-therapy | ECCITE-seq, which captures surface protein expression, cellular transcriptome, HIV-1 RNA, and TCR sequence within the same single cells in longitudinally archived paired samples during actual viremia and one-year post-ART | HIV revised in heterogenous granzyme B+ Th1 effector memory CD4+ T cells with robust antigen response, proliferation potential, and long-term clonal stability | [80] |
| Isolate reservoir cells from HIV patients treated with long-term ART | FIND-seq, simultaneous capture of polyadenylated RNA and HIV DNA from single cells, and scRNA-seq without latency reversal | HIV-DNA+ memory CD4+ T cells inhibited death receptor, necroptosis, and antiproliferative signaling but showed higher expression of known negative regulators of HIV transcription, thus favoring long-term survival of the cells and HIV silencing | [88] |
| The transcriptome of HIV-infected and HIV-exposed peripheral blood monocytes | scRNA seq | Monocytes provide varying degrees of permissive cellular environment for HIV infection, and ART has a detrimental effect on monocyte function | [94] |
| scRNA profiling of HIV-infected cells in CNS | Single nucleus RNA-seq of archived brain tissue | HIV-induced interferon response modulates host chromatin conformation and HIV integration sites in microglia | [95] |
4. Implications of scRNA-Seq in Animal Models of TB and HIV
5. Gaps in Knowledge and Future Directions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sharan, R.; Bucşan, A.N.; Ganatra, S.; Paiardini, M.; Mohan, M.; Mehra, S.; Khader, S.A.; Kaushal, D. Chronic Immune Activation in TB/HIV Co-infection. Trends Microbiol. 2020, 28, 619–632. [Google Scholar] [CrossRef]
- Jones-López, E.C.; Namugga, O.; Mumbowa, F.; Ssebidandi, M.; Mbabazi, O.; Moine, S.; Mboowa, G.; Fox, M.P.; Reilly, N.; Ayakaka, I.; et al. Cough aerosols of Mycobacterium tuberculosis predict new infection: A household contact study. Am. J. Respir. Crit. Care Med. 2013, 187, 1007–1015. [Google Scholar] [CrossRef]
- World Health Organization. Global Tuberculosis Report 2022. 2022. Available online: https://www.who.int/teams/global-tuberculosis-programme/tb-reports/global-tuberculosis-report-2022 (accessed on 29 May 2023).
- Adhikari, N.; Bhattarai, R.B.; Basnet, R.; Joshi, L.R.; Tinkari, B.S.; Thapa, A.; Joshi, B. Prevalence and associated risk factors for tuberculosis among people living with HIV in Nepal. PLoS ONE 2022, 17, e0262720. [Google Scholar] [CrossRef] [PubMed]
- Runels, T.; Ragan, E.J.; Ventura, A.S.; Winter, M.R.; White, L.F.; Horsburgh, C.R.; Samet, J.H.; Saitz, R.; Jacobson, K.R. Testing and treatment for latent tuberculosis infection in people living with HIV and substance dependence: A prospective cohort study. BMJ Open 2022, 12, e058751. [Google Scholar] [CrossRef]
- Shea, K.M.; Kammerer, J.S.; Winston, C.A.; Navin, T.R.; Horsburgh, C.R., Jr. Estimated rate of reactivation of latent tuberculosis infection in the United States, overall and by population subgroup. Am. J. Epidemiol. 2014, 179, 216–225. [Google Scholar] [CrossRef] [PubMed]
- Bruchfeld, J.; Correia-Neves, M.; Källenius, G. Tuberculosis and HIV Coinfection. Cold Spring Harb. Perspect. Med. 2015, 5, a017871. [Google Scholar] [CrossRef] [PubMed]
- Lavalett, L.; Rodriguez, H.; Ortega, H.; Sadee, W.; Schlesinger, L.S.; Barrera, L.F. Alveolar macrophages from tuberculosis patients display an altered inflammatory gene expression profile. Tuberculosis 2017, 107, 156–167. [Google Scholar] [CrossRef] [PubMed]
- Guo, Q.; Zhong, Y.; Wang, Z.; Cao, T.; Zhang, M.; Zhang, P.; Huang, W.; Bi, J.; Yuan, Y.; Ou, M.; et al. Single-cell transcriptomic landscape identifies the expansion of peripheral blood monocytes as an indicator of HIV-1-TB co-infection. Cell Insight 2022, 1, 100005. [Google Scholar] [CrossRef] [PubMed]
- McCaffrey, E.F.; Donato, M.; Keren, L.; Chen, Z.; Delmastro, A.; Fitzpatrick, M.B.; Gupta, S.; Greenwald, N.F.; Baranski, A.; Graf, W.; et al. The immunoregulatory landscape of human tuberculosis granulomas. Nat. Immunol. 2022, 23, 318–329. [Google Scholar] [CrossRef]
- Tilahun, M.; Shibabaw, A.; Kiflie, A.; Bewket, G.; Abate, E.; Gelaw, B. Latent tuberculosis infection and associated risk factors among people living with HIV and apparently healthy blood donors at the University of Gondar referral hospital, Northwest Ethiopia. BMC Res. Notes 2019, 12, 515. [Google Scholar] [CrossRef]
- Hoerter, A.; Arnett, E.; Schlesinger, L.S.; Pienaar, E. Systems biology approaches to investigate the role of granulomas in TB-HIV coinfection. Front. Immunol. 2022, 13, 1014515. [Google Scholar] [CrossRef]
- Bucşan, A.N.; Chatterjee, A.; Singh, D.K.; Foreman, T.W.; Lee, T.H.; Threeton, B.; Kirkpatrick, M.G.; Ahmed, M.; Golden, N.; Alvarez, X.; et al. Mechanisms of reactivation of latent tuberculosis infection due to SIV coinfection. J. Clin. Investig. 2019, 129, 5254–5260. [Google Scholar] [CrossRef] [PubMed]
- Bell, L.C.K.; Noursadeghi, M. Pathogenesis of HIV-1 and Mycobacterium tuberculosis co-infection. Nat. Rev. Microbiol. 2018, 16, 80–90. [Google Scholar] [CrossRef] [PubMed]
- Souriant, S.; Balboa, L.; Dupont, M.; Pingris, K.; Kviatcovsky, D.; Cougoule, C.; Lastrucci, C.; Bah, A.; Gasser, R.; Poincloux, R.; et al. Tuberculosis Exacerbates HIV-1 Infection through IL-10/STAT3-Dependent Tunneling Nanotube Formation in Macrophages. Cell Rep. 2019, 26, 3586–3599. [Google Scholar] [CrossRef] [PubMed]
- Wyndham-Thomas, C.; Corbière, V.; Selis, E.; Payen, M.C.; Goffard, J.C.; Van Vooren, J.P.; Mascart, F.; Dirix, V. Immune Activation by Mycobacterium tuberculosis in HIV-Infected and -Uninfected Subjects. J. Acquir. Immune Defic. Syndr. 2017, 74, 103–111. [Google Scholar] [CrossRef]
- LaVergne, S.; Umlauf, A.; McCutchan, A.; Heaton, R.; Benson, C.; Kumarasamy, N.; Bharti, A.R. Impact of Latent Tuberculosis Infection on Neurocognitive Functioning and Inflammation in HIV-Infected and Uninfected South Indians. J. Acquir. Immune Defic. Syndr. 2020, 84, 430–436. [Google Scholar] [CrossRef]
- Sharan, R.; Ganatra, S.R.; Bucsan, A.N.; Cole, J.; Singh, D.K.; Alvarez, X.; Gough, M.; Alvarez, C.; Blakley, A.; Ferdin, J.; et al. Antiretroviral therapy timing impacts latent tuberculosis infection reactivation in a Mycobacterium tuberculosis/SIV coinfection model. J. Clin. Investig. 2022, 132. [Google Scholar] [CrossRef]
- Ganatra, S.R.; Bucşan, A.N.; Alvarez, X.; Kumar, S.; Chatterjee, A.; Quezada, M.; Fish, A.; Singh, D.K.; Singh, B.; Sharan, R.; et al. Antiretroviral therapy does not reduce tuberculosis reactivation in a tuberculosis-HIV coinfection model. J. Clin. Investig. 2020, 130, 5171–5179. [Google Scholar] [CrossRef]
- Teklu, A.M.; Nega, A.; Mamuye, A.T.; Sitotaw, Y.; Kassa, D.; Mesfin, G.; Belayihun, B.; Medhin, G.; Yirdaw, K. Factors Associated with Mortality of TB/HIV Co-infected Patients in Ethiopia. Ethiop. J. Health Sci. 2017, 27, 29–38. [Google Scholar] [CrossRef]
- Zicari, S.; Sessa, L.; Cotugno, N.; Ruggiero, A.; Morrocchi, E.; Concato, C.; Rocca, S.; Zangari, P.; Manno, E.C.; Palma, P. Immune Activation, Inflammation, and Non-AIDS Co-Morbidities in HIV-Infected Patients under Long-Term ART. Viruses 2019, 11, 200. [Google Scholar] [CrossRef]
- Paiardini, M.; Müller-Trutwin, M. HIV-associated chronic immune activation. Immunol. Rev. 2013, 254, 78–101. [Google Scholar] [CrossRef]
- Wilkinson, K.A.; Meintjes, G.; Seldon, R.; Goliath, R.; Wilkinson, R.J. Immunological characterisation of an unmasking TB-IRIS case. S. Afr. Med. J. 2012, 102, 512–517. [Google Scholar] [CrossRef]
- Luo, G.; Gao, Q.; Zhang, S.; Yan, B. Probing infectious disease by single-cell RNA sequencing: Progresses and perspectives. Comput. Struct. Biotechnol. J. 2020, 18, 2962–2971. [Google Scholar] [CrossRef]
- Penaranda, C.; Hung, D.T. Single-Cell RNA Sequencing to Understand Host-Pathogen Interactions. ACS Infect. Dis. 2019, 5, 336–344. [Google Scholar] [CrossRef]
- Xu, Y.; Tan, Y.; Zhang, X.; Cheng, M.; Hu, J.; Liu, J.; Chen, X.; Zhu, J. Comprehensive identification of immuno-related transcriptional signature for active pulmonary tuberculosis by integrated analysis of array and single cell RNA-seq. J. Infect. 2022, 85, 534–544. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Hu, C.; Lu, W.; Hang, T.; Shao, Y.; Chen, C.; Wang, Y.; Li, N.; Jin, L.; Wu, W.; et al. Characteristics of alveolar macrophages in bronchioalveolar lavage fluids from active tuberculosis patients identified by single-cell RNA sequencing. J. Biomed. Res. 2022, 36, 167–180. [Google Scholar] [CrossRef] [PubMed]
- Golumbeanu, M.; Cristinelli, S.; Rato, S.; Munoz, M.; Cavassini, M.; Beerenwinkel, N.; Ciuffi, A. Single-Cell RNA-Seq Reveals Transcriptional Heterogeneity in Latent and Reactivated HIV-Infected Cells. Cell Rep. 2018, 23, 942–950. [Google Scholar] [CrossRef] [PubMed]
- Pollara, J.; Khanal, S.; Edwards, R.W.; Hora, B.; Ferrari, G.; Haynes, B.F.; Bradley, T. Single-cell analysis of immune cell transcriptome during HIV-1 infection and therapy. BMC Immunol. 2022, 23, 48. [Google Scholar] [CrossRef]
- Kazer, S.W.; Aicher, T.P.; Muema, D.M.; Carroll, S.L.; Ordovas-Montanes, J.; Miao, V.N.; Tu, A.A.; Ziegler, C.G.K.; Nyquist, S.K.; Wong, E.B.; et al. Integrated single-cell analysis of multicellular immune dynamics during hyperacute HIV-1 infection. Nat. Med. 2020, 26, 511–518. [Google Scholar] [CrossRef]
- Hamada, Y.; Sidibe, A.; Matteelli, A.; Dadu, A.; Aziz, M.A.; Del Granado, M.; Nishikiori, N.; Floyd, K.; Getahun, H. Policies and practices on the programmatic management of latent tuberculous infection: Global survey. Int. J. Tuberc. Lung Dis. 2016, 20, 1566–1571. [Google Scholar] [CrossRef]
- White, H.A.; Miller, R.F.; Pozniak, A.L.; Lipman, M.C.; Stephenson, I.; Wiselka, M.J.; Pareek, M. Latent tuberculosis infection screening and treatment in HIV: Insights from evaluation of UK practice. Thorax 2017, 72, 180–182. [Google Scholar] [CrossRef]
- Reaves, E.J.; Shah, N.S.; France, A.M.; Morris, S.B.; Kammerer, S.; Skarbinski, J.; Bradley, H. Latent tuberculous infection testing among HIV-infected persons in clinical care, United States, 2010–2012. Int. J. Tuberc. Lung Dis. 2017, 21, 1118–1126. [Google Scholar] [CrossRef] [PubMed]
- Adams, J.W.; Howe, C.J.; Andrews, A.C.; Allen, S.L.; Vinnard, C. Tuberculosis screening among HIV-infected patients: Tuberculin skin test vs. interferon-gamma release assay. AIDS Care 2017, 29, 1504–1509. [Google Scholar] [CrossRef] [PubMed]
- National Institutes of Health; Centers for Disease Control and Prevention; HIV Medicine Association of the Infectious Diseases Society of America; Panel on Opportunistic Infections in Adults and Adolescents with HIV. Guidelines for the prevention and treatment of opportunistic infections in adults and adolescents with HIV: Recommendations from the Centers for Disease Control and Prevention, the National Institutes of Health, and the HIV Medicine Association of the Infectious Diseases Society of America. Mycobact. Tuberc. Infect. Dis. 2022. Available online: https://clinicalinfo.hiv.gov/en/guidelines/hiv-clinical-guidelines-adult-and-adolescent-opportunistic-infections/whats-new (accessed on 29 May 2023).
- Sterling, T.R.; Njie, G.; Zenner, D.; Cohn, D.L.; Reves, R.; Ahmed, A.; Menzies, D.; Horsburgh, C.R., Jr.; Crane, C.M.; Burgos, M.; et al. Guidelines for the Treatment of Latent Tuberculosis Infection: Recommendations from the National Tuberculosis Controllers Association and CDC, 2020. MMWR Recomm. Rep. 2020, 69, 1–11. [Google Scholar] [CrossRef]
- Gong, W.; Wu, X. Differential Diagnosis of Latent Tuberculosis Infection and Active Tuberculosis: A Key to a Successful Tuberculosis Control Strategy. Front. Microbiol. 2021, 12, 745592. [Google Scholar] [CrossRef]
- Jasmer, R.M.; Nahid, P.; Hopewell, P.C. Clinical practice. Latent tuberculosis infection. N. Engl. J. Med. 2002, 347, 1860–1866. [Google Scholar] [CrossRef] [PubMed]
- Pourakbari, B.; Mamishi, S.; Benvari, S.; Mahmoudi, S. Comparison of the QuantiFERON-TB Gold Plus and QuantiFERON-TB Gold In-Tube interferon-γ release assays: A systematic review and meta-analysis. Adv. Med. Sci. 2019, 64, 437–443. [Google Scholar] [CrossRef] [PubMed]
- Chapman, A.L.; Munkanta, M.; Wilkinson, K.A.; Pathan, A.A.; Ewer, K.; Ayles, H.; Reece, W.H.; Mwinga, A.; Godfrey-Faussett, P.; Lalvani, A. Rapid detection of active and latent tuberculosis infection in HIV-positive individuals by enumeration of Mycobacterium tuberculosis-specific T cells. AIDS 2002, 16, 2285–2293. [Google Scholar] [CrossRef]
- Dheda, K.; Lalvani, A.; Miller, R.F.; Scott, G.; Booth, H.; Johnson, M.A.; Zumla, A.; Rook, G.A. Performance of a T-cell-based diagnostic test for tuberculosis infection in HIV-infected individuals is independent of CD4 cell count. AIDS 2005, 19, 2038–2041. [Google Scholar] [CrossRef]
- Domínguez, J.; Latorre, I.; Altet, N.; Mateo, L.; De Souza-Galvão, M.; Ruiz-Manzano, J.; Ausina, V. IFN-gamma-release assays to diagnose TB infection in the immunocompromised individual. Expert Rev. Respir. Med. 2009, 3, 309–327. [Google Scholar] [CrossRef]
- Rangaka, M.X.; Wilkinson, K.A.; Seldon, R.; Van Cutsem, G.; Meintjes, G.A.; Morroni, C.; Mouton, P.; Diwakar, L.; Connell, T.G.; Maartens, G.; et al. Effect of HIV-1 infection on T-Cell-based and skin test detection of tuberculosis infection. Am. J. Respir. Crit. Care Med. 2007, 175, 514–520. [Google Scholar] [CrossRef]
- Richeldi, L.; Losi, M.; D’Amico, R.; Luppi, M.; Ferrari, A.; Mussini, C.; Codeluppi, M.; Cocchi, S.; Prati, F.; Paci, V.; et al. Performance of tests for latent tuberculosis in different groups of immunocompromised patients. Chest 2009, 136, 198–204. [Google Scholar] [CrossRef] [PubMed]
- Talati, N.J.; Seybold, U.; Humphrey, B.; Aina, A.; Tapia, J.; Weinfurter, P.; Albalak, R.; Blumberg, H.M. Poor concordance between interferon-gamma release assays and tuberculin skin tests in diagnosis of latent tuberculosis infection among HIV-infected individuals. BMC Infect. Dis. 2009, 9, 15. [Google Scholar] [CrossRef] [PubMed]
- Brock, I.; Ruhwald, M.; Lundgren, B.; Westh, H.; Mathiesen, L.R.; Ravn, P. Latent tuberculosis in HIV positive, diagnosed by the M. tuberculosis specific interferon-gamma test. Respir. Res. 2006, 7, 56. [Google Scholar] [CrossRef]
- Steffen, R.E.; Pinto, M.; Kritski, A.; Trajman, A. Cost-effectiveness of newer technologies for the diagnosis of Mycobacterium tuberculosis infection in Brazilian people living with HIV. Sci. Rep. 2020, 10, 21823. [Google Scholar] [CrossRef]
- Chongxing, Z.; Yuanchun, L.; Yan, H.; Dabin, L.; Zhezhe, C.; Liwen, H.; Huifang, Q.; Jing, Y.; Fengxue, L.; Xiaoyan, L.; et al. Evaluation of the Diagnostic Efficacy of EC-Test for Latent Tuberculosis Infection in Ambulatory People Living with HIV. AIDS 2023, 37, 1791–1797. [Google Scholar] [CrossRef]
- Chandra, P.; Grigsby, S.J.; Philips, J.A. Immune evasion and provocation by Mycobacterium tuberculosis. Nat. Rev. Microbiol. 2022, 20, 750–766. [Google Scholar] [CrossRef] [PubMed]
- Gideon, H.P.; Flynn, J.L. Latent tuberculosis: What the host “sees”? Immunol. Res. 2011, 50, 202–212. [Google Scholar] [CrossRef]
- Wong, N.S.; Leung, C.C.; Chan, K.C.W.; Chan, W.K.; Lin, A.W.C.; Lee, S.S. A longitudinal study on latent TB infection screening and its association with TB incidence in HIV patients. Sci. Rep. 2019, 9, 10093. [Google Scholar] [CrossRef]
- Liao, S.Y.; Atif, S.M.; Mould, K.; Konigsberg, I.R.; Fu, R.; Davidson, E.; Li, L.; Fontenot, A.P.; Maier, L.A.; Yang, I.V. Single-cell RNA sequencing identifies macrophage transcriptional heterogeneities in granulomatous diseases. Eur. Respir. J. 2021, 57, 2003794. [Google Scholar] [CrossRef] [PubMed]
- Avraham, R.; Haseley, N.; Brown, D.; Penaranda, C.; Jijon, H.B.; Trombetta, J.J.; Satija, R.; Shalek, A.K.; Xavier, R.J.; Regev, A.; et al. Pathogen Cell-to-Cell Variability Drives Heterogeneity in Host Immune Responses. Cell 2015, 162, 1309–1321. [Google Scholar] [CrossRef] [PubMed]
- Avraham, R.; Hung, D.T. A perspective on single cell behavior during infection. Gut Microbes 2016, 7, 518–525. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Gideon, H.P.; Hughes, T.K.; Tzouanas, C.N.; Wadsworth, M.H., 2nd; Tu, A.A.; Gierahn, T.M.; Peters, J.M.; Hopkins, F.F.; Wei, J.R.; Kummerlowe, C.; et al. Multimodal profiling of lung granulomas in macaques reveals cellular correlates of tuberculosis control. Immunity 2022, 55, 827–846. [Google Scholar] [CrossRef]
- Ma, F.; Hughes, T.K.; Teles, R.M.B.; Andrade, P.R.; de Andrade Silva, B.J.; Plazyo, O.; Tsoi, L.C.; Do, T.; Wadsworth, M.H., 2nd; Oulee, A.; et al. The cellular architecture of the antimicrobial response network in human leprosy granulomas. Nat. Immunol. 2021, 22, 839–850. [Google Scholar] [CrossRef]
- Cai, Y.; Dai, Y.; Wang, Y.; Yang, Q.; Guo, J.; Wei, C.; Chen, W.; Huang, H.; Zhu, J.; Zhang, C.; et al. Single-cell transcriptomics of blood reveals a natural killer cell subset depletion in tuberculosis. EBioMedicine 2020, 53, 102686. [Google Scholar] [CrossRef]
- Kulkarni, V.; Queiroz, A.T.L.; Sangle, S.; Kagal, A.; Salvi, S.; Gupta, A.; Ellner, J.; Kadam, D.; Rolla, V.C.; Andrade, B.B.; et al. A Two-Gene Signature for Tuberculosis Diagnosis in Persons with Advanced HIV. Front. Immunol. 2021, 12, 631165. [Google Scholar] [CrossRef]
- Hillman, H.; Khan, N.; Singhania, A.; Dubelko, P.; Soldevila, F.; Tippalagama, R.; DeSilva, A.D.; Gunasena, B.; Perera, J.; Scriba, T.J.; et al. Single-cell profiling reveals distinct subsets of CD14+ monocytes drive blood immune signatures of active tuberculosis. Front. Immunol. 2022, 13, 1087010. [Google Scholar] [CrossRef]
- Akter, S.; Chauhan, K.S.; Dunlap, M.D.; Choreño-Parra, J.A.; Lu, L.; Esaulova, E.; Zúñiga, J.; Artyomov, M.N.; Kaushal, D.; Khader, S.A. Mycobacterium tuberculosis infection drives a type I IFN signature in lung lymphocytes. Cell Rep. 2022, 39, 110983. [Google Scholar] [CrossRef]
- Hosmane, N.N.; Kwon, K.J.; Bruner, K.M.; Capoferri, A.A.; Beg, S.; Rosenbloom, D.I.; Keele, B.F.; Ho, Y.C.; Siliciano, J.D.; Siliciano, R.F. Proliferation of latently infected CD4+ T cells carrying replication-competent HIV-1: Potential role in latent reservoir dynamics. J. Exp. Med. 2017, 214, 959–972. [Google Scholar] [CrossRef]
- Lorenzi, J.C.; Cohen, Y.Z.; Cohn, L.B.; Kreider, E.F.; Barton, J.P.; Learn, G.H.; Oliveira, T.; Lavine, C.L.; Horwitz, J.A.; Settler, A.; et al. Paired quantitative and qualitative assessment of the replication-competent HIV-1 reservoir and comparison with integrated proviral DNA. Proc. Natl. Acad. Sci. USA 2016, 113, E7908–E7916. [Google Scholar] [CrossRef] [PubMed]
- Bui, J.K.; Sobolewski, M.D.; Keele, B.F.; Spindler, J.; Musick, A.; Wiegand, A.; Luke, B.T.; Shao, W.; Hughes, S.H.; Coffin, J.M.; et al. Proviruses with identical sequences comprise a large fraction of the replication-competent HIV reservoir. PLoS Pathog. 2017, 13, e1006283. [Google Scholar] [CrossRef] [PubMed]
- Bui, J.K.; Halvas, E.K.; Fyne, E.; Sobolewski, M.D.; Koontz, D.; Shao, W.; Luke, B.; Hong, F.F.; Kearney, M.F.; Mellors, J.W. Ex vivo activation of CD4+ T-cells from donors on suppressive ART can lead to sustained production of infectious HIV-1 from a subset of infected cells. PLoS Pathog. 2017, 13, e1006230. [Google Scholar] [CrossRef] [PubMed]
- Chomont, N.; El-Far, M.; Ancuta, P.; Trautmann, L.; Procopio, F.A.; Yassine-Diab, B.; Boucher, G.; Boulassel, M.R.; Ghattas, G.; Brenchley, J.M.; et al. HIV reservoir size and persistence are driven by T cell survival and homeostatic proliferation. Nat. Med. 2009, 15, 893–900. [Google Scholar] [CrossRef] [PubMed]
- Henrich, T.J.; Hatano, H.; Bacon, O.; Hogan, L.E.; Rutishauser, R.; Hill, A.; Kearney, M.F.; Anderson, E.M.; Buchbinder, S.P.; Cohen, S.E.; et al. HIV-1 persistence following extremely early initiation of antiretroviral therapy (ART) during acute HIV-1 infection: An observational study. PLoS Med. 2017, 14, e1002417. [Google Scholar] [CrossRef]
- Gantner, P.; Pagliuzza, A.; Pardons, M.; Ramgopal, M.; Routy, J.P.; Fromentin, R.; Chomont, N. Single-cell TCR sequencing reveals phenotypically diverse clonally expanded cells harboring inducible HIV proviruses during ART. Nat. Commun. 2020, 11, 4089. [Google Scholar] [CrossRef]
- Simonetti, F.R.; Zhang, H.; Soroosh, G.P.; Duan, J.; Rhodehouse, K.; Hill, A.L.; Beg, S.A.; McCormick, K.; Raymond, H.E.; Nobles, C.L.; et al. Antigen-driven clonal selection shapes the persistence of HIV-1-infected CD4+ T cells in vivo. J. Clin. Investig. 2021, 131. [Google Scholar] [CrossRef]
- Douek, D.C.; Brenchley, J.M.; Betts, M.R.; Ambrozak, D.R.; Hill, B.J.; Okamoto, Y.; Casazza, J.P.; Kuruppu, J.; Kunstman, K.; Wolinsky, S.; et al. HIV preferentially infects HIV-specific CD4+ T cells. Nature 2002, 417, 95–98. [Google Scholar] [CrossRef]
- Mendoza, P.; Jackson, J.R.; Oliveira, T.Y.; Gaebler, C.; Ramos, V.; Caskey, M.; Jankovic, M.; Nussenzweig, M.C.; Cohn, L.B. Antigen-responsive CD4+ T cell clones contribute to the HIV-1 latent reservoir. J. Exp. Med. 2020, 217, e20200051. [Google Scholar] [CrossRef]
- Bradley, T.; Ferrari, G.; Haynes, B.F.; Margolis, D.M.; Browne, E.P. Single-Cell Analysis of Quiescent HIV Infection Reveals Host Transcriptional Profiles that Regulate Proviral Latency. Cell Rep. 2018, 25, 107–117. [Google Scholar] [CrossRef]
- Einkauf, K.B.; Osborn, M.R.; Gao, C.; Sun, W.; Sun, X.; Lian, X.; Parsons, E.M.; Gladkov, G.T.; Seiger, K.W.; Blackmer, J.E.; et al. Parallel analysis of transcription, integration, and sequence of single HIV-1 proviruses. Cell 2022, 185, 266–282 e215. [Google Scholar] [CrossRef] [PubMed]
- De Scheerder, M.A.; Vrancken, B.; Dellicour, S.; Schlub, T.; Lee, E.; Shao, W.; Rutsaert, S.; Verhofstede, C.; Kerre, T.; Malfait, T.; et al. HIV Rebound Is Predominantly Fueled by Genetically Identical Viral Expansions from Diverse Reservoirs. Cell Host Microbe 2019, 26, 347–358 e347. [Google Scholar] [CrossRef] [PubMed]
- Cohn, L.B.; Silva, I.T.; Oliveira, T.Y.; Rosales, R.A.; Parrish, E.H.; Learn, G.H.; Hahn, B.H.; Czartoski, J.L.; McElrath, M.J.; Lehmann, C.; et al. HIV-1 integration landscape during latent and active infection. Cell 2015, 160, 420–432. [Google Scholar] [CrossRef] [PubMed]
- Cho, A.; Gaebler, C.; Olveira, T.; Ramos, V.; Saad, M.; Lorenzi, J.C.C.; Gazumyan, A.; Moir, S.; Caskey, M.; Chun, T.W.; et al. Longitudinal clonal dynamics of HIV-1 latent reservoirs measured by combination quadruplex polymerase chain reaction and sequencing. Proc. Natl. Acad. Sci. USA 2022, 119, e2117630119. [Google Scholar] [CrossRef]
- Antar, A.A.; Jenike, K.M.; Jang, S.; Rigau, D.N.; Reeves, D.B.; Hoh, R.; Krone, M.R.; Keruly, J.C.; Moore, R.D.; Schiffer, J.T.; et al. Longitudinal study reveals HIV-1-infected CD4+ T cell dynamics during long-term antiretroviral therapy. J. Clin. Investig. 2020, 130, 3543–3559. [Google Scholar] [CrossRef] [PubMed]
- Huang, A.S.; Ramos, V.; Oliveira, T.Y.; Gaebler, C.; Jankovic, M.; Nussenzweig, M.C.; Cohn, L.B. Integration features of intact latent HIV-1 in CD4+ T cell clones contribute to viral persistence. J. Exp. Med. 2021, 218, e20211427. [Google Scholar] [CrossRef]
- Cohn, L.B.; da Silva, I.T.; Valieris, R.; Huang, A.S.; Lorenzi, J.C.C.; Cohen, Y.Z.; Pai, J.A.; Butler, A.L.; Caskey, M.; Jankovic, M.; et al. Clonal CD4+ T cells in the HIV-1 latent reservoir display a distinct gene profile upon reactivation. Nat. Med. 2018, 24, 604–609. [Google Scholar] [CrossRef]
- Liu, R.; Yeh, Y.J.; Varabyou, A.; Collora, J.A.; Sherrill-Mix, S.; Talbot, C.C., Jr.; Mehta, S.; Albrecht, K.; Hao, H.; Zhang, H.; et al. Single-cell transcriptional landscapes reveal HIV-1-driven aberrant host gene transcription as a potential therapeutic target. Sci. Transl. Med. 2020, 12, eaaz0802. [Google Scholar] [CrossRef]
- Collora, J.A.; Liu, R.; Pinto-Santini, D.; Ravindra, N.; Ganoza, C.; Lama, J.R.; Alfaro, R.; Chiarella, J.; Spudich, S.; Mounzer, K.; et al. Single-cell multiomics reveals persistence of HIV-1 in expanded cytotoxic T cell clones. Immunity 2022, 55, 1013–1031.e7. [Google Scholar] [CrossRef]
- Lee, G.Q.; Orlova-Fink, N.; Einkauf, K.; Chowdhury, F.Z.; Sun, X.; Harrington, S.; Kuo, H.H.; Hua, S.; Chen, H.R.; Ouyang, Z.; et al. Clonal expansion of genome-intact HIV-1 in functionally polarized Th1 CD4+ T cells. J. Clin. Investig. 2017, 127, 2689–2696. [Google Scholar] [CrossRef]
- Horsburgh, B.A.; Lee, E.; Hiener, B.; Eden, J.S.; Schlub, T.E.; von Stockenstrom, S.; Odevall, L.; Milush, J.M.; Liegler, T.; Sinclair, E.; et al. High levels of genetically intact HIV in HLA-DR+ memory T cells indicates their value for reservoir studies. AIDS 2020, 34, 659–668. [Google Scholar] [CrossRef] [PubMed]
- Hiener, B.; Horsburgh, B.A.; Eden, J.S.; Barton, K.; Schlub, T.E.; Lee, E.; von Stockenstrom, S.; Odevall, L.; Milush, J.M.; Liegler, T.; et al. Identification of Genetically Intact HIV-1 Proviruses in Specific CD4+ T Cells from Effectively Treated Participants. Cell Rep. 2017, 21, 813–822. [Google Scholar] [CrossRef] [PubMed]
- Cockerham, L.R.; Siliciano, J.D.; Sinclair, E.; O’Doherty, U.; Palmer, S.; Yukl, S.A.; Strain, M.C.; Chomont, N.; Hecht, F.M.; Siliciano, R.F.; et al. CD4+ and CD8+ T cell activation are associated with HIV DNA in resting CD4+ T cells. PLoS ONE 2014, 9, e110731. [Google Scholar] [CrossRef]
- Weymar, G.H.J.; Bar-On, Y.; Oliveira, T.Y.; Gaebler, C.; Ramos, V.; Hartweger, H.; Breton, G.; Caskey, M.; Cohn, L.B.; Jankovic, M.; et al. Distinct gene expression by expanded clones of quiescent memory CD4+ T cells harboring intact latent HIV-1 proviruses. Cell Rep. 2022, 40, 111311. [Google Scholar] [CrossRef] [PubMed]
- Doitsh, G.; Greene, W.C. Dissecting How CD4 T Cells Are Lost During HIV Infection. Cell Host Microbe 2016, 19, 280–291. [Google Scholar] [CrossRef]
- Clark, I.C.; Wheeler, M.A.; Lee, H.G.; Li, Z.; Sanmarco, L.M.; Thaploo, S.; Polonio, C.M.; Shin, S.W.; Scalisi, G.; Henry, A.R.; et al. Identification of astrocyte regulators by nucleic acid cytometry. Nature 2023, 614, 326–333. [Google Scholar] [CrossRef] [PubMed]
- Clark, I.C.; Mudvari, P.; Thaploo, S.; Smith, S.; Abu-Laban, M.; Hamouda, M.; Theberge, M.; Shah, S.; Ko, S.H.; Perez, L.; et al. HIV silencing and cell survival signatures in infected T cell reservoirs. Nature 2023, 614, 318–325. [Google Scholar] [CrossRef] [PubMed]
- Swirski, F.K.; Nahrendorf, M.; Etzrodt, M.; Wildgruber, M.; Cortez-Retamozo, V.; Panizzi, P.; Figueiredo, J.L.; Kohler, R.H.; Chudnovskiy, A.; Waterman, P.; et al. Identification of splenic reservoir monocytes and their deployment to inflammatory sites. Science 2009, 325, 612–616. [Google Scholar] [CrossRef]
- Lund, H.; Boysen, P.; Akesson, C.P.; Lewandowska-Sabat, A.M.; Storset, A.K. Transient Migration of Large Numbers of CD14++ CD16+ Monocytes to the Draining Lymph Node after Onset of Inflammation. Front. Immunol. 2016, 7, 322. [Google Scholar] [CrossRef]
- Micci, L.; Alvarez, X.; Iriele, R.I.; Ortiz, A.M.; Ryan, E.S.; McGary, C.S.; Deleage, C.; McAtee, B.B.; He, T.; Apetrei, C.; et al. CD4 depletion in SIV-infected macaques results in macrophage and microglia infection with rapid turnover of infected cells. PLoS Pathog. 2014, 10, e1004467. [Google Scholar] [CrossRef]
- Burdo, T.H.; Soulas, C.; Orzechowski, K.; Button, J.; Krishnan, A.; Sugimoto, C.; Alvarez, X.; Kuroda, M.J.; Williams, K.C. Increased monocyte turnover from bone marrow correlates with severity of SIV encephalitis and CD163 levels in plasma. PLoS Pathog. 2010, 6, e1000842. [Google Scholar] [CrossRef] [PubMed]
- Fischer-Smith, T.; Croul, S.; Sverstiuk, A.E.; Capini, C.; L’Heureux, D.; Regulier, E.G.; Richardson, M.W.; Amini, S.; Morgello, S.; Khalili, K.; et al. CNS invasion by CD14+/CD16+ peripheral blood-derived monocytes in HIV dementia: Perivascular accumulation and reservoir of HIV infection. J. Neurovirol. 2001, 7, 528–541. [Google Scholar] [CrossRef] [PubMed]
- Leon-Rivera, R.; Morsey, B.; Niu, M.; Fox, H.S.; Berman, J.W. Interactions of Monocytes, HIV, and ART Identified by an Innovative scRNAseq Pipeline: Pathways to Reservoirs and HIV-Associated Comorbidities. mBio 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- Plaza-Jennings, A.L.; Valada, A.; O’Shea, C.; Iskhakova, M.; Hu, B.; Javidfar, B.; Ben Hutta, G.; Lambert, T.Y.; Murray, J.; Kassim, B.; et al. HIV integration in the human brain is linked to microglial activation and 3D genome remodeling. Mol. Cell 2022, 82, 4647–4663.e8. [Google Scholar] [CrossRef] [PubMed]
- Rane, L.; Rahman, S.; Magalhaes, I.; Ahmed, R.; Spangberg, M.; Kondova, I.; Verreck, F.; Andersson, J.; Brighenti, S.; Maeurer, M.J. Increased (6 exon) interleukin-7 production after M. tuberculosis infection and soluble interleukin-7 receptor expression in lung tissue. Genes Immun. 2011, 12, 513–522. [Google Scholar] [CrossRef] [PubMed]
- Abrahams, M.R.; Joseph, S.B.; Garrett, N.; Tyers, L.; Moeser, M.; Archin, N.; Council, O.D.; Matten, D.; Zhou, S.; Doolabh, D.; et al. The replication-competent HIV-1 latent reservoir is primarily established near the time of therapy initiation. Sci. Transl. Med. 2019, 11, eaaw5589. [Google Scholar] [CrossRef]
- Vandergeeten, C.; Fromentin, R.; DaFonseca, S.; Lawani, M.B.; Sereti, I.; Lederman, M.M.; Ramgopal, M.; Routy, J.P.; Sekaly, R.P.; Chomont, N. Interleukin-7 promotes HIV persistence during antiretroviral therapy. Blood 2013, 121, 4321–4329. [Google Scholar] [CrossRef]
- Renault, C.; Veyrenche, N.; Mennechet, F.; Bedin, A.S.; Routy, J.P.; Van de Perre, P.; Reynes, J.; Tuaillon, E. Th17 CD4+ T-Cell as a Preferential Target for HIV Reservoirs. Front. Immunol. 2022, 13, 822576. [Google Scholar] [CrossRef]
- Jurado, J.O.; Pasquinelli, V.; Alvarez, I.B.; Pena, D.; Rovetta, A.I.; Tateosian, N.L.; Romeo, H.E.; Musella, R.M.; Palmero, D.; Chuluyan, H.E.; et al. IL-17 and IFN-gamma expression in lymphocytes from patients with active tuberculosis correlates with the severity of the disease. J. Leukoc. Biol. 2012, 91, 991–1002. [Google Scholar] [CrossRef]
- Chen, X.; Zhang, M.; Liao, M.; Graner, M.W.; Wu, C.; Yang, Q.; Liu, H.; Zhou, B. Reduced Th17 response in patients with tuberculosis correlates with IL-6R expression on CD4+ T Cells. Am. J. Respir. Crit. Care Med. 2010, 181, 734–742. [Google Scholar] [CrossRef]
- Xu, L.; Cui, G.; Jia, H.; Zhu, Y.; Ding, Y.; Chen, J.; Lu, C.; Ye, P.; Gao, H.; Li, L.; et al. Decreased IL-17 during treatment of sputum smear-positive pulmonary tuberculosis due to increased regulatory T cells and IL-10. J. Transl. Med. 2016, 14, 179. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Xun, J.; Yang, J.; Ji, Y.; Liu, L.; Qi, T.; Wang, Z.; Zhang, R.; Shen, Y.; Ponte, R.; et al. Plasma Indoleamine 2,3-Dioxygenase Activity Is Associated with the Size of the Human Immunodeficiency Virus Reservoir in Patients Receiving Antiretroviral Therapy. Clin. Infect. Dis. 2019, 68, 1274–1281. [Google Scholar] [CrossRef] [PubMed]
- Adu-Gyamfi, C.G.; Snyman, T.; Hoffmann, C.J.; Martinson, N.A.; Chaisson, R.E.; George, J.A.; Suchard, M.S. Plasma Indoleamine 2, 3-Dioxygenase, a Biomarker for Tuberculosis in Human Immunodeficiency Virus-Infected Patients. Clin. Infect. Dis. 2017, 65, 1356–1358. [Google Scholar] [CrossRef] [PubMed]
- Estévez, O.; Anibarro, L.; Garet, E.; Pallares, Á.; Barcia, L.; Calviño, L.; Maueia, C.; Mussá, T.; Fdez-Riverola, F.; Glez-Peña, D.; et al. An RNA-seq Based Machine Learning Approach Identifies Latent Tuberculosis Patients with an Active Tuberculosis Profile. Front. Immunol. 2020, 11, 1470. [Google Scholar] [CrossRef]
- Nathan, A.; Beynor, J.I.; Baglaenko, Y.; Suliman, S.; Ishigaki, K.; Asgari, S.; Huang, C.C.; Luo, Y.; Zhang, Z.; Lopez, K.; et al. Multimodally profiling memory T cells from a tuberculosis cohort identifies cell state associations with demographics, environment and disease. Nat. Immunol. 2021, 22, 781–793. [Google Scholar] [CrossRef]
- Boisson-Dupuis, S.; Bustamante, J.; El-Baghdadi, J.; Camcioglu, Y.; Parvaneh, N.; El Azbaoui, S.; Agader, A.; Hassani, A.; El Hafidi, N.; Mrani, N.A.; et al. Inherited and acquired immunodeficiencies underlying tuberculosis in childhood. Immunol. Rev. 2015, 264, 103–120. [Google Scholar] [CrossRef]
- Scriba, T.J.; Kalsdorf, B.; Abrahams, D.A.; Isaacs, F.; Hofmeister, J.; Black, G.; Hassan, H.Y.; Wilkinson, R.J.; Walzl, G.; Gelderbloem, S.J.; et al. Distinct, specific IL-17- and IL-22-producing CD4+ T cell subsets contribute to the human anti-mycobacterial immune response. J. Immunol. 2008, 180, 1962–1970. [Google Scholar] [CrossRef]
- Okada, S.; Markle, J.G.; Deenick, E.K.; Mele, F.; Averbuch, D.; Lagos, M.; Alzahrani, M.; Al-Muhsen, S.; Halwani, R.; Ma, C.S.; et al. Impairment of immunity to Candida and Mycobacterium in humans with bi-allelic RORC mutations. Science 2015, 349, 606–613. [Google Scholar] [CrossRef]
- Bossel Ben-Moshe, N.; Hen-Avivi, S.; Levitin, N.; Yehezkel, D.; Oosting, M.; Joosten, L.A.B.; Netea, M.G.; Avraham, R. Predicting bacterial infection outcomes using single cell RNA-sequencing analysis of human immune cells. Nat. Commun. 2019, 10, 3266. [Google Scholar] [CrossRef]
- Oelen, R.; de Vries, D.H.; Brugge, H.; Gordon, M.G.; Vochteloo, M.; Ye, C.J.; Westra, H.J.; Franke, L.; van der Wijst, M.G.P. Single-cell RNA-sequencing of peripheral blood mononuclear cells reveals widespread, context-specific gene expression regulation upon pathogenic exposure. Nat. Commun. 2022, 13, 3267. [Google Scholar] [CrossRef]
- Thobakgale, C.; Naidoo, K.; McKinnon, L.R.; Werner, L.; Samsunder, N.; Karim, S.A.; Ndung’u, T.; Altfeld, M.; Naidoo, K. Interleukin 1-Beta (IL-1β) Production by Innate Cells Following TLR Stimulation Correlates with TB Recurrence in ART-Treated HIV-Infected Patients. J. Acquir. Immune Defic. Syndr. 2017, 74, 213–220. [Google Scholar] [CrossRef]
- McLaren, P.J.; Fellay, J. HIV-1 and human genetic variation. Nat. Rev. Genet. 2021, 22, 645–657. [Google Scholar] [CrossRef]
- Horton, R.E.; McLaren, P.J.; Fowke, K.; Kimani, J.; Ball, T.B. Cohorts for the study of HIV-1-exposed but uninfected individuals: Benefits and limitations. J. Infect. Dis. 2010, 202 (Suppl. S3), S377–S381. [Google Scholar] [CrossRef] [PubMed]
- Kulkarni, P.S.; Butera, S.T.; Duerr, A.C. Resistance to HIV-1 infection: Lessons learned from studies of highly exposed persistently seronegative (HEPS) individuals. AIDS Rev. 2003, 5, 87–103. [Google Scholar] [PubMed]
- Sabin, C.A.; Lundgren, J.D. The natural history of HIV infection. Curr. Opin. HIV AIDS 2013, 8, 311–317. [Google Scholar] [CrossRef] [PubMed]
- Ciuffi, A.; Bleiber, G.; Munoz, M.; Martinez, R.; Loeuillet, C.; Rehr, M.; Fischer, M.; Gunthard, H.F.; Oxenius, A.; Meylan, P.; et al. Entry and transcription as key determinants of differences in CD4 T-cell permissiveness to human immunodeficiency virus type 1 infection. J. Virol. 2004, 78, 10747–10754. [Google Scholar] [CrossRef] [PubMed]
- Mohammadi, P.; Desfarges, S.; Bartha, I.; Joos, B.; Zangger, N.; Munoz, M.; Gunthard, H.F.; Beerenwinkel, N.; Telenti, A.; Ciuffi, A. 24 hours in the life of HIV-1 in a T cell line. PLoS Pathog. 2013, 9, e1003161. [Google Scholar] [CrossRef]
- Rausell, A.; Munoz, M.; Martinez, R.; Roger, T.; Telenti, A.; Ciuffi, A. Innate immune defects in HIV permissive cell lines. Retrovirology 2016, 13, 43. [Google Scholar] [CrossRef]
- Rato, S.; Rausell, A.; Munoz, M.; Telenti, A.; Ciuffi, A. Single-cell analysis identifies cellular markers of the HIV permissive cell. PLoS Pathog. 2017, 13, e1006678. [Google Scholar] [CrossRef]
- Lim, A.L.; Moos, P.; Pond, C.D.; Larson, E.C.; Martins, L.J.; Szaniawski, M.A.; Planelles, V.; Barrows, L.R. HIV-1 provirus transcription and translation in macrophages differs from pre-integrated cDNA complexes and requires E2F transcriptional programs. Virulence 2022, 13, 386–413. [Google Scholar] [CrossRef]
- Mahnke, Y.D.; Fletez-Brant, K.; Sereti, I.; Roederer, M. Reconstitution of Peripheral T Cells by Tissue-Derived CCR4+ Central Memory Cells Following HIV-1 Antiretroviral Therapy. Pathog. Immun. 2016, 1, 260–290. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.M.; Zhang, J.Y.; Xing, X.; Huang, H.H.; Xia, P.; Dai, X.P.; Hu, W.; Zhang, C.; Song, J.W.; Fan, X.; et al. Global transcriptomic characterization of T cells in individuals with chronic HIV-1 infection. Cell Discov. 2022, 8, 29. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Zhang, Q.; Hui, H.; Agrawal, K.; Karris, M.A.Y.; Rana, T.M. An atlas of immune cell exhaustion in HIV-infected individuals revealed by single-cell transcriptomics. Emerg. Microbes Infect. 2020, 9, 2333–2347. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Tang, Y.; Wang, Y.; Li, Y.; Yang, Y.; Liao, K.; Song, F.; Deng, S.; Chen, Y. Single-cell sequencing resolves the landscape of immune cells and regulatory mechanisms in HIV-infected immune non-responders. Cell Death Dis. 2022, 13, 849. [Google Scholar] [CrossRef]
- Niu, M.; Morsey, B.; Lamberty, B.G.; Emanuel, K.; Yu, F.; Leon-Rivera, R.; Berman, J.W.; Gaskill, P.J.; Matt, S.M.; Ciborowski, P.S.; et al. Methamphetamine Increases the Proportion of SIV-Infected Microglia/Macrophages, Alters Metabolic Pathways, and Elevates Cell Death Pathways: A Single-Cell Analysis. Viruses 2020, 12, 1297. [Google Scholar] [CrossRef]
- Tiwari, S.; van Tonder, A.J.; Vilchèze, C.; Mendes, V.; Thomas, S.E.; Malek, A.; Chen, B.; Chen, M.; Kim, J.; Blundell, T.L.; et al. Arginine-deprivation-induced oxidative damage sterilizes Mycobacterium tuberculosis. Proc. Natl. Acad. Sci. USA 2018, 115, 9779–9784. [Google Scholar] [CrossRef]
- Farhadian, S.F.; Mehta, S.S.; Zografou, C.; Robertson, K.; Price, R.W.; Pappalardo, J.; Chiarella, J.; Hafler, D.A.; Spudich, S.S. Single-cell RNA sequencing reveals microglia-like cells in cerebrospinal fluid during virologically suppressed HIV. JCI Insight 2018, 3, e121718. [Google Scholar] [CrossRef]
- Foreman, T.W.; Mehra, S.; Lackner, A.A.; Kaushal, D. Translational Research in the Nonhuman Primate Model of Tuberculosis. ILAR J. 2017, 58, 151–159. [Google Scholar] [CrossRef]
- Scanga, C.A.; Flynn, J.L. Modeling tuberculosis in nonhuman primates. Cold Spring Harb. Perspect. Med. 2014, 4, a018564. [Google Scholar] [CrossRef]
- Cooper, E.B.; Brent, L.J.N.; Snyder-Mackler, N.; Singh, M.; Sengupta, A.; Khatiwada, S.; Malaivijitnond, S.; Qi Hai, Z.; Higham, J.P. The rhesus macaque as a success story of the Anthropocene. Elife 2022, 11, e78169. [Google Scholar] [CrossRef]
- Sharan, R.; Ganatra, S.R.; Singh, D.K.; Cole, J.; Foreman, T.W.; Thippeshappa, R.; Peloquin, C.A.; Shivanna, V.; Gonzalez, O.; Day, C.L.; et al. Isoniazid and rifapentine treatment effectively reduces persistent M. tuberculosis infection in macaque lungs. J. Clin. Investig. 2022, 132, e121718. [Google Scholar] [CrossRef]
- Esaulova, E.; Das, S.; Singh, D.K.; Choreño-Parra, J.A.; Swain, A.; Arthur, L.; Rangel-Moreno, J.; Ahmed, M.; Singh, B.; Gupta, A.; et al. The immune landscape in tuberculosis reveals populations linked to disease and latency. Cell Host Microbe 2021, 29, 165–178. [Google Scholar] [CrossRef] [PubMed]
- Han, L.; Wei, X.; Liu, C.; Volpe, G.; Zhuang, Z.; Zou, X.; Wang, Z.; Pan, T.; Yuan, Y.; Zhang, X.; et al. Cell transcriptomic atlas of the non-human primate Macaca fascicularis. Nature 2022, 604, 723–731. [Google Scholar] [CrossRef]
- Qu, J.; Yang, F.; Zhu, T.; Wang, Y.; Fang, W.; Ding, Y.; Zhao, X.; Qi, X.; Xie, Q.; Chen, M.; et al. A reference single-cell regulomic and transcriptomic map of cynomolgus monkeys. Nat. Commun. 2022, 13, 4069. [Google Scholar] [CrossRef]
- Pisu, D.; Huang, L.; Narang, V.; Theriault, M.; Lê-Bury, G.; Lee, B.; Lakudzala, A.E.; Mzinza, D.T.; Mhango, D.V.; Mitini-Nkhoma, S.C.; et al. Single cell analysis of M. tuberculosis phenotype and macrophage lineages in the infected lung. J. Exp. Med. 2021, 218, e20210615. [Google Scholar] [CrossRef] [PubMed]
- Berry, M.P.; Graham, C.M.; McNab, F.W.; Xu, Z.; Bloch, S.A.; Oni, T.; Wilkinson, K.A.; Banchereau, R.; Skinner, J.; Wilkinson, R.J.; et al. An interferon-inducible neutrophil-driven blood transcriptional signature in human tuberculosis. Nature 2010, 466, 973–977. [Google Scholar] [CrossRef]
- Pan, J.; Zhang, X.; Xu, J.; Chang, Z.; Xin, Z.; Wang, G. Landscape of Exhausted T Cells in Tuberculosis Revealed by Single-Cell Sequencing. Microbiol. Spectr. 2023, 11, e02839-22. [Google Scholar] [CrossRef] [PubMed]
- Sannier, G.; Dubé, M.; Kaufmann, D.E. Single-Cell Technologies Applied to HIV-1 Research: Reaching Maturity. Front. Microbiol. 2020, 11, 297. [Google Scholar] [CrossRef]
- Mbonye, U.; Karn, J. The Molecular Basis for Human Immunodeficiency Virus Latency. Annu. Rev. Virol. 2017, 4, 261–285. [Google Scholar] [CrossRef]
- Trease, A.J.; Niu, M.; Morsey, B.; Guda, C.; Byrareddy, S.N.; Buch, S.; Fox, H.S. Antiretroviral therapy restores the homeostatic state of microglia in SIV-infected rhesus macaques. J. Leukoc. Biol. 2022, 112, 969–981. [Google Scholar] [CrossRef]
- Han, Q.; Bradley, T.; Williams, W.B.; Cain, D.W.; Montefiori, D.C.; Saunders, K.O.; Parks, R.J.; Edwards, R.W.; Ferrari, G.; Mueller, O.; et al. Neonatal Rhesus Macaques Have Distinct Immune Cell Transcriptional Profiles following HIV Envelope Immunization. Cell Rep. 2020, 30, 1553–1569. [Google Scholar] [CrossRef]
- Marsden, M.D. Benefits and limitations of humanized mice in HIV persistence studies. Retrovirology 2020, 17, 7. [Google Scholar] [CrossRef]
- Victor Garcia, J. Humanized mice for HIV and AIDS research. Curr. Opin. Virol. 2016, 19, 56–64. [Google Scholar] [CrossRef]
- Cheng, L.; Yu, H.; Wrobel, J.A.; Li, G.; Liu, P.; Hu, Z.; Xu, X.N.; Su, L. Identification of pathogenic TRAIL-expressing innate immune cells during HIV-1 infection in humanized mice by scRNA-Seq. JCI Insight 2020, 5, e135344. [Google Scholar] [CrossRef]
- Aso, H.; Nagaoka, S.; Kawakami, E.; Ito, J.; Islam, S.; Tan, B.J.Y.; Nakaoka, S.; Ashizaki, K.; Shiroguchi, K.; Suzuki, Y.; et al. Multiomics Investigation Revealing the Characteristics of HIV-1-Infected Cells In Vivo. Cell Rep. 2020, 32, 107887. [Google Scholar] [CrossRef] [PubMed]
- Nusbaum, R.J.; Calderon, V.E.; Huante, M.B.; Sutjita, P.; Vijayakumar, S.; Lancaster, K.L.; Hunter, R.L.; Actor, J.K.; Cirillo, J.D.; Aronson, J.; et al. Pulmonary Tuberculosis in Humanized Mice Infected with HIV-1. Sci. Rep. 2016, 6, 21522. [Google Scholar] [CrossRef] [PubMed]
- Huante, M.B.; Saito, T.B.; Nusbaum, R.J.; Naqvi, K.F.; Chauhan, S.; Hunter, R.L.; Actor, J.K.; Rudra, J.S.; Endsley, M.A.; Lisinicchia, J.G.; et al. Small Animal Model of Post-chemotherapy Tuberculosis Relapse in the Setting of HIV Co-infection. Front. Cell. Infect. Microbiol. 2020, 10, 150. [Google Scholar] [CrossRef] [PubMed]
- Endsley, J.J.; Huante, M.B.; Naqvi, K.F.; Gelman, B.B.; Endsley, M.A. Advancing our understanding of HIV co-infections and neurological disease using the humanized mouse. Retrovirology 2021, 18, 14. [Google Scholar] [CrossRef]
- Cevaal, P.M.; Bekker, L.G.; Hermans, S. TB-IRIS pathogenesis and new strategies for intervention: Insights from related inflammatory disorders. Tuberculosis 2019, 118, 101863. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kulkarni, S.; Endsley, J.J.; Lai, Z.; Bradley, T.; Sharan, R. Single-Cell Transcriptomics of Mtb/HIV Co-Infection. Cells 2023, 12, 2295. https://doi.org/10.3390/cells12182295
Kulkarni S, Endsley JJ, Lai Z, Bradley T, Sharan R. Single-Cell Transcriptomics of Mtb/HIV Co-Infection. Cells. 2023; 12(18):2295. https://doi.org/10.3390/cells12182295
Chicago/Turabian StyleKulkarni, Smita, Janice J. Endsley, Zhao Lai, Todd Bradley, and Riti Sharan. 2023. "Single-Cell Transcriptomics of Mtb/HIV Co-Infection" Cells 12, no. 18: 2295. https://doi.org/10.3390/cells12182295
APA StyleKulkarni, S., Endsley, J. J., Lai, Z., Bradley, T., & Sharan, R. (2023). Single-Cell Transcriptomics of Mtb/HIV Co-Infection. Cells, 12(18), 2295. https://doi.org/10.3390/cells12182295