
The Biological Functions and Intestinal Inflammation Regulation of IL-21 in Grass Carp (Ctenopharyngodon idella) during Infection with Aeromonas hydrophila


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Fish, Cells and Bacteria
2.2. Plasmids and Reagents
2.3. Bacterial Challenge
2.4. Hematoxylin and Eosin (H&E) Staining of Grass Carp Intestine
2.5. Expression Analysis of il21 in the Primary HKLs after Stimulation with Inactivated A. hydrophila and LPS
2.6. RNA Extraction, cDNA Synthesis and qRT-PCR
2.7. Western Blotting
2.8. Immunofluorescent Microscopy
2.9. Production and Bioactivity Testing of Recombinant IL-21
2.10. Luciferase Promoter Reporter Assay
2.11. Statistical Analysis
3. Results
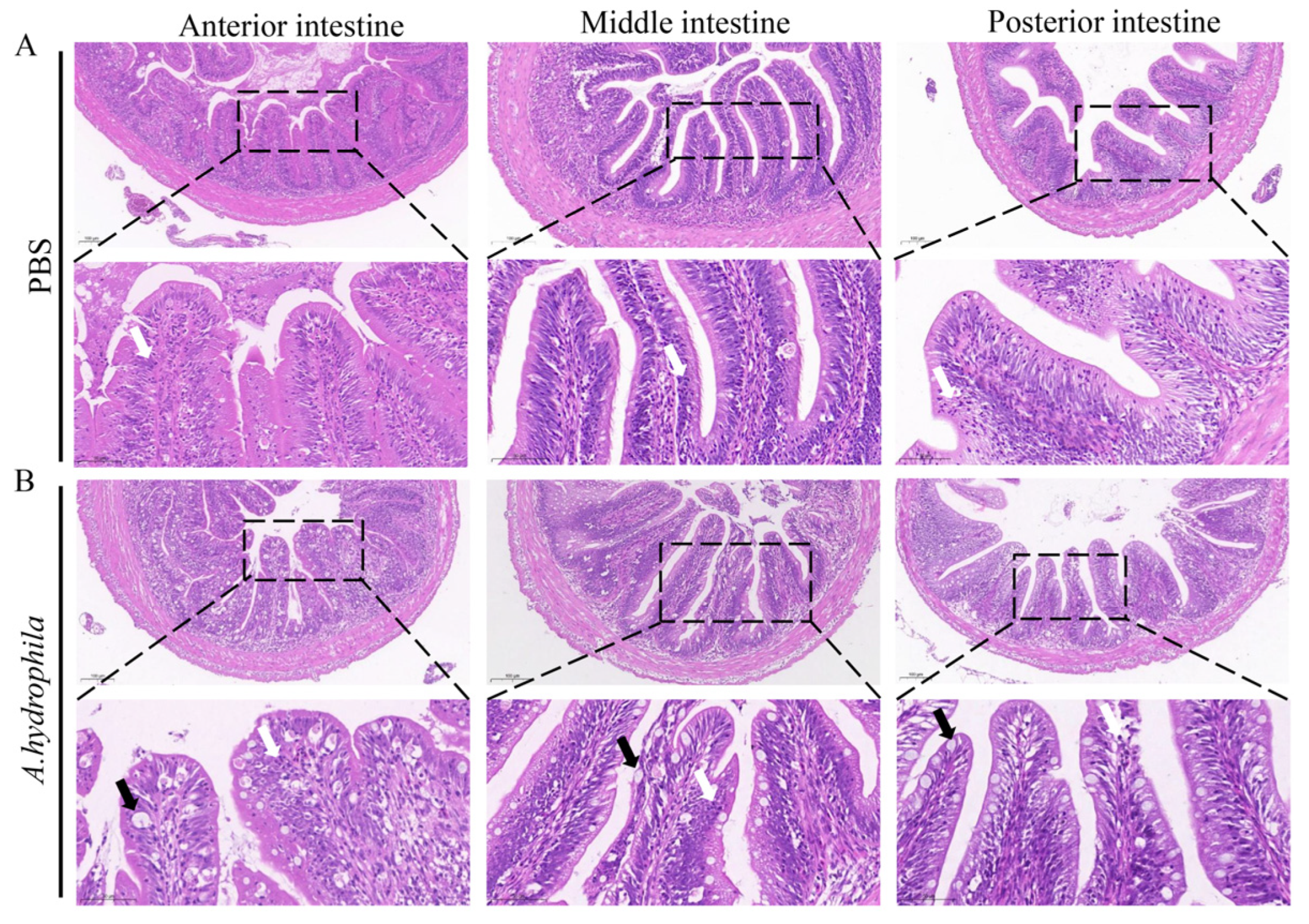
3.1. Pathological Changes in Different Segments of Intestine after Infection with A. hydrophila
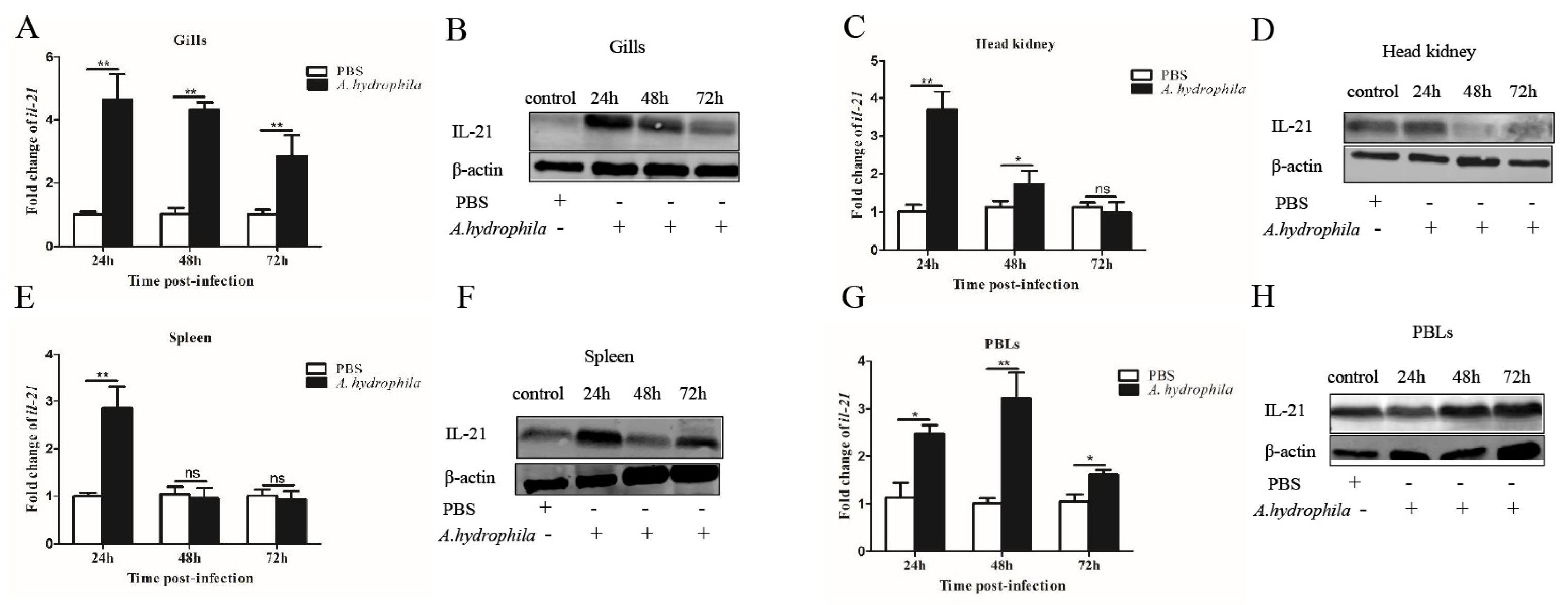
3.2. Tissue Expression Patterns of il21 after A. hydrophila Infection
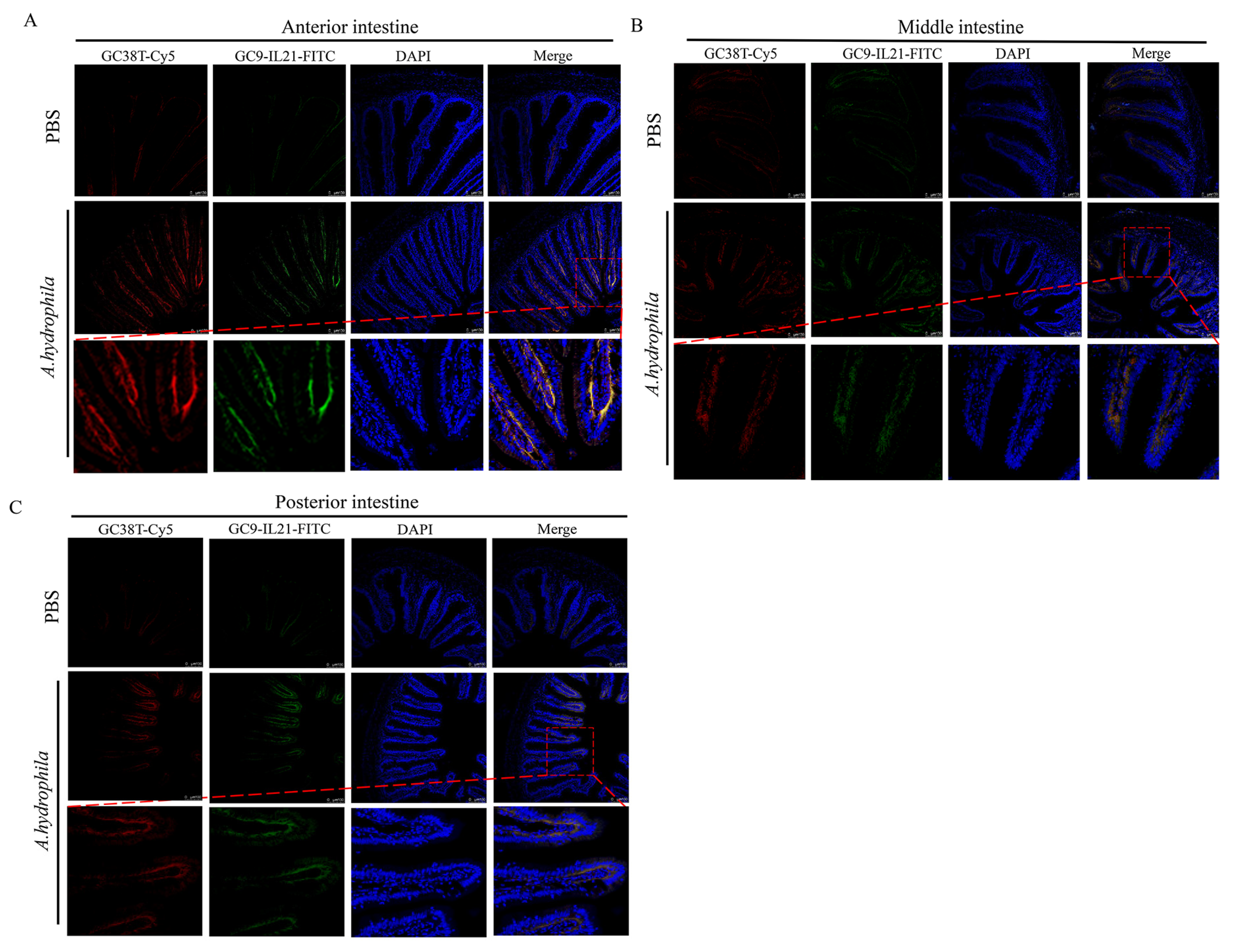
3.3. Analysis of IL-21 Producing CD3γ/δ T Cells in Different Segments of Intestine after A. hydrophila Infection
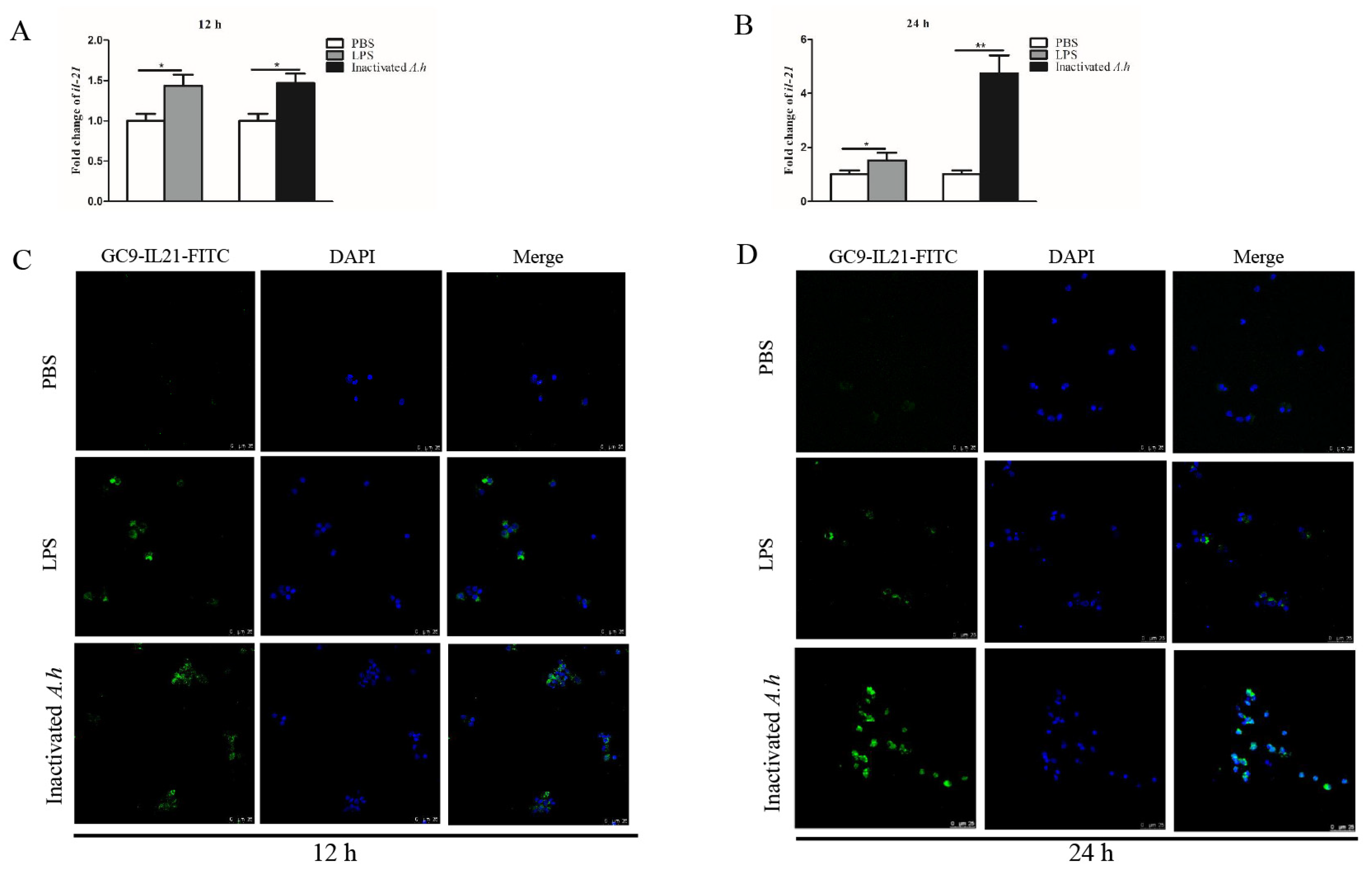
3.4. Expression Analysis of il21 in the Primary HKLs after Stimulation with LPS and Inactivated A. hydrophila
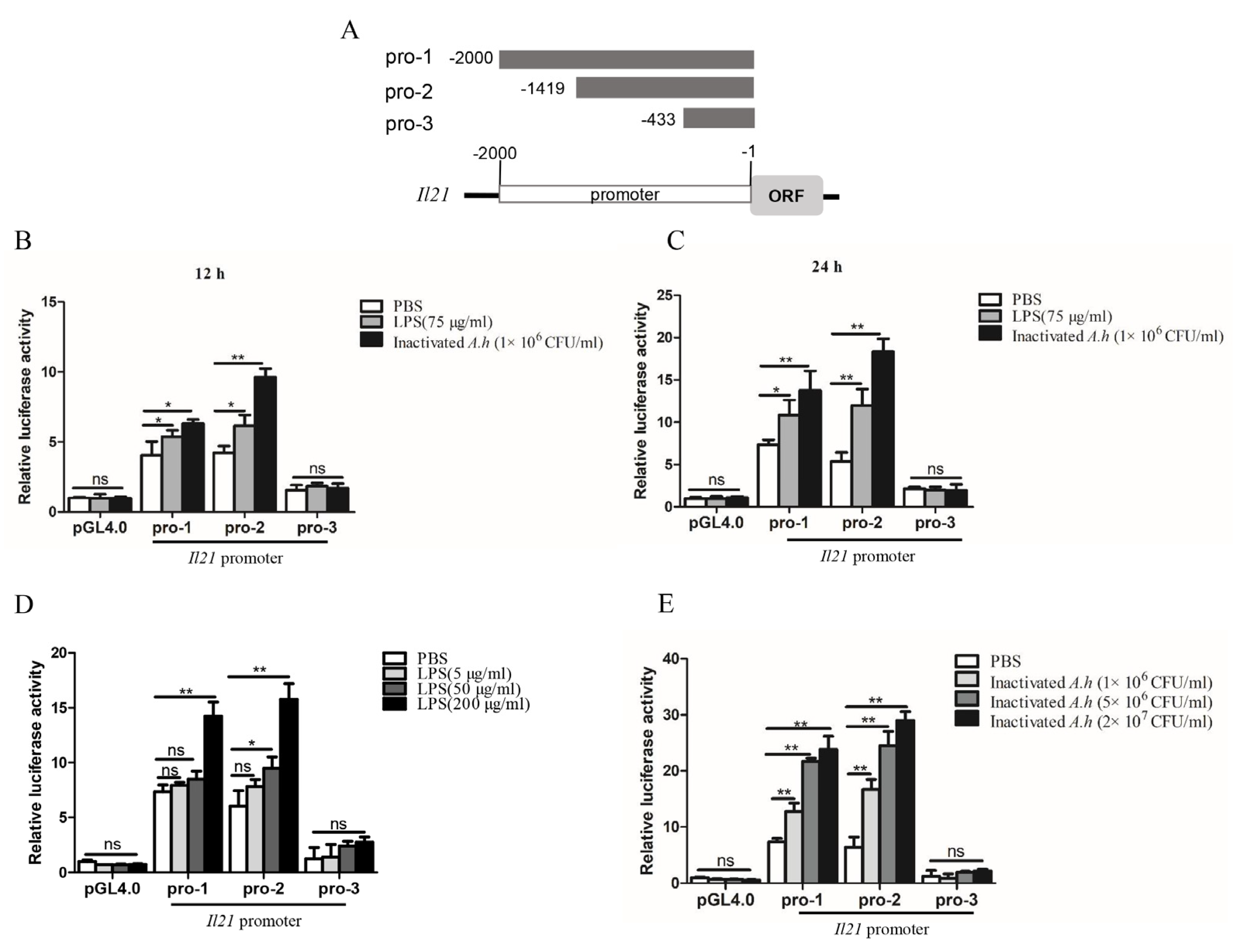
3.5. Analysis of Promoter Activity of il21
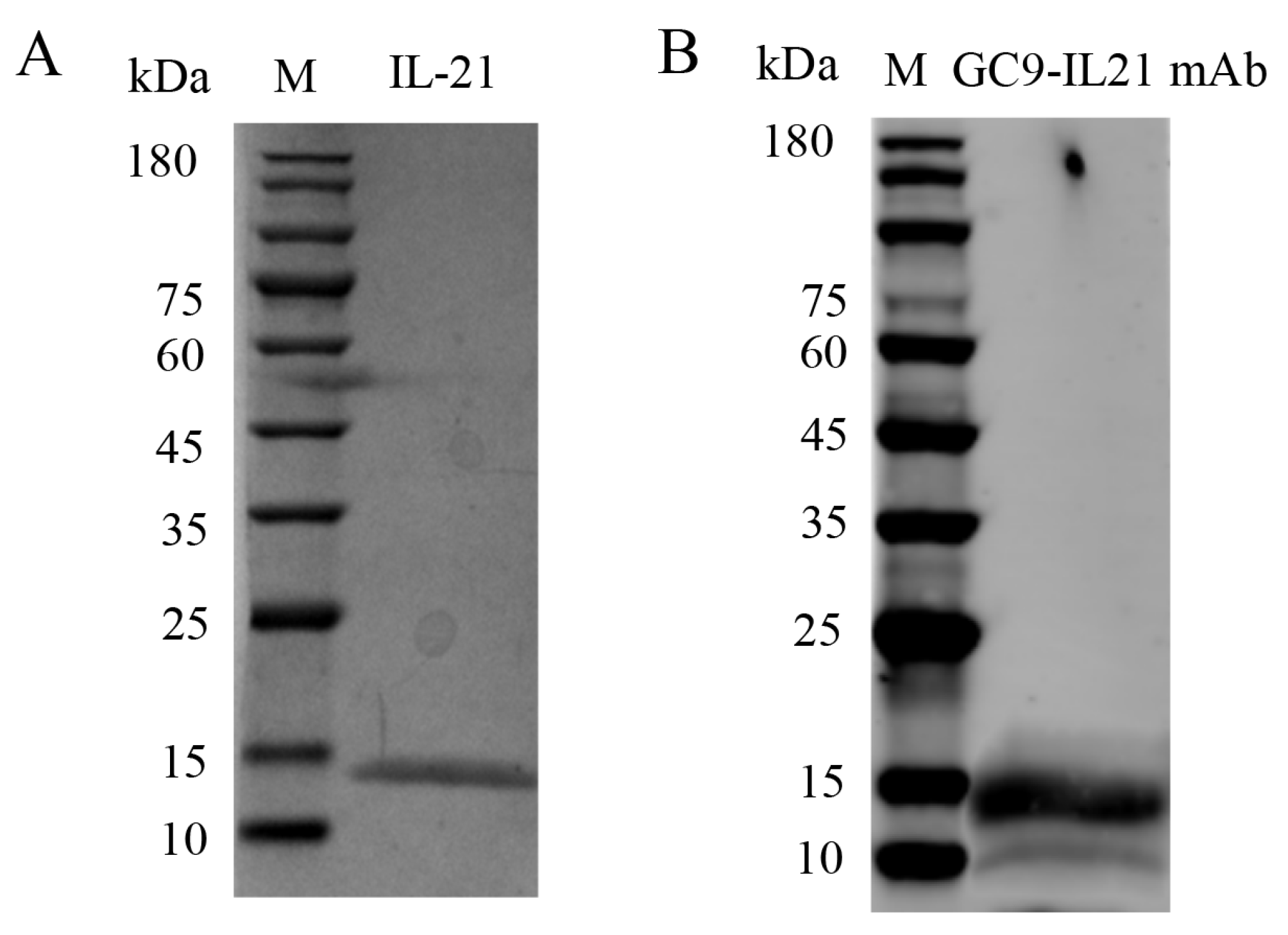
3.6. Biotesting of Bioactivities of rIL-21 Protein
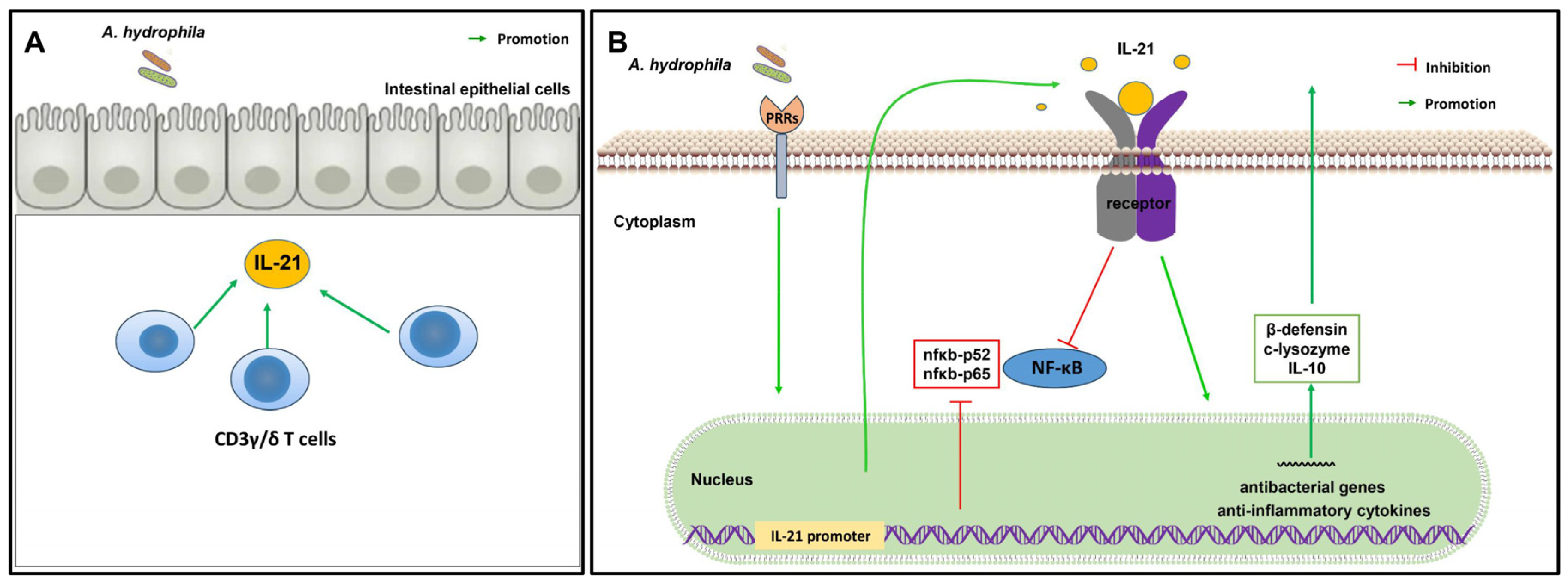
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Parrish, N.J.; Dillon, S.R.; Nelson, A.; Hammond, A.; Sprecher, C.; Gross, J.A.; Johnston, J.; Madden, K.; Xu, W.; West, J.; et al. Interleukin 21 and its receptor are involved in NK cell expansion and regulation of lymphocyte function. Nature 2000, 408, 57–63. [Google Scholar] [CrossRef]
- Nurieva, R.; Yang, X.; Martinez, G.; Zhang, Y.; Panopoulos, A.D.; Ma, L.; Schluns, K.; Tian, Q.; Watowich, S.S.; Jetten, A.M.; et al. Essential autocrine regulation by IL-21 in the generation of inflammatory T cells. Nature 2007, 448, 480–483. [Google Scholar] [CrossRef] [PubMed]
- Spolski, R.; Leonard, W.J. IL-21 and T follicular helper cells. Int. Immunol. 2010, 22, 7–12. [Google Scholar] [CrossRef] [PubMed]
- Ghalamfarsa, G.; Mahmoudi, M.; Mohammadnia, A.M.; Yazdani, Y.; Anvari, E.; Hadinia, A.; Ghanbari, A.; Setayesh, M.; Yousefi, M.; Jadidi, N.F. IL-21 and IL-21 receptor in the immunopathogenesis of multiple sclerosis. J. Immunotoxicol. 2016, 13, 274–285. [Google Scholar] [CrossRef] [PubMed]
- Parrish, N.J.; Foster, D.C.; Holly, R.D.; Clegg, C.H. Interleukin-21 and the IL-21 receptor: Novel effectors of NK and T cell responses. J. Leukoc. Biol. 2002, 72, 856–863. [Google Scholar] [CrossRef]
- Collins, M.; Whitters, M.J.; Young, D.A. IL-21 and IL-21 Receptor: A New Cytokine Pathway Modulates Innate and Adaptive Immunity. Immunol. Res. 2003, 28, 131–140. [Google Scholar] [CrossRef]
- Yi, J.S.; Cox, M.A.; Zajac, A.J. Interleukin-21: A multifunctional regulator of immunity to infections. Microbes Infect. 2010, 12, 1111–1119. [Google Scholar] [CrossRef]
- Rückert, R.; Bulfone, P.S.; Brandt, K. Interleukin-21 stimulates antigen uptake, protease activity, survival and induction of CD4+ T cell proliferation by murine macrophages. Clin. Exp. Immunol. 2008, 151, 487–495. [Google Scholar] [CrossRef]
- Li, S.; Wang, W.; Fu, S.; Wang, J.; Liu, H.; Xie, S.; Liu, B.; Li, Y.; Lv, Q.; Li, Z.; et al. IL-21 Modulates release of proinflammatory cytokines in LPS-Stimulated macrophages through distinct signaling pathways. Mediat. Inflamm. 2013, 2013, 548073. [Google Scholar] [CrossRef]
- Coquet, J.M.; Kyparissoudis, K.; Pellicci, D.G.; Besra, G.; Berzins, S.P.; Smyth, M.J.; Godfrey, D.I. IL-21 Is produced by NKT cells and nodulates NKT cell activation and cytokine production. J. Immunol. 2007, 178, 2827–2834. [Google Scholar] [CrossRef]
- Dvorscek, A.R.; McKenzie, C.I.; Robinson, M.J.; Ding, Z.; Pitt, C.; O’Donnell, K.; Zotos, D.; Brink, R.; Tarlinton, D.M.; Quast, I. IL-21 has a critical role in establishing germinal centers by amplifying early B cell proliferation. EMBO Rep. 2022, 23, e54677. [Google Scholar] [CrossRef] [PubMed]
- Quast, I.; Dvorscek, A.R.; Pattaroni, C.; Steiner, T.M.; McKenzie, C.I.; Pitt, C.; O’Donnell, K.; Ding, Z.; Hill, D.L.; Brink, R.; et al. Interleukin-21, acting beyond the immunological synapse, independently controls T follicular helper and germinal center B cells. Immunity 2022, 55, 1414–1430.e5. [Google Scholar] [CrossRef] [PubMed]
- Brandt, K.; Bulfone, P.S.; Foster, D.C.; Rückert, R. Interleukin-21 inhibits dendritic cell activation and maturation. Blood. 2003, 102, 4090–4098. [Google Scholar] [CrossRef] [PubMed]
- Bird, S.; Zou, J.; Kono, T.; Sakai, M.; Dijkstra, J.M.; Secombes, C. Characterisation and expression analysis of interleukin 2 (IL-2) and IL-21 homologues in the Japanese pufferfish, Fugu rubripes, following their discovery by synteny. Immunogenetics 2005, 56, 909–923. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Xiang, L.; Shao, J.; Jia, S. Molecular cloning, characterization and expression analysis of an IL-21 homologue from Tetraodon nigroviridis. Cytokine 2006, 35, 126–134. [Google Scholar] [CrossRef] [PubMed]
- Gunimaladevi, I.; Savan, R.; Sato, K.; Yamaguchi, R.; Sakai, M. Characterization of an interleukin-15 like (IL-15L) gene from zebrafish (Danio rerio). Fish Shellfish Immunol. 2007, 22, 351–362. [Google Scholar] [CrossRef]
- Wang, T.; Diaz, R.P.; Costa, M.M.; Campbell, S.; Snow, M.; Collet, B.; Martin, S.A.M.; Secombes, C.J. Functional characterization of a nonmammalian IL-21: Rainbow trout Oncorhynchus mykiss IL-21 upregulates the expression of the Th cell signature cytokines IFN-γ, IL-10, and IL-22. J. Immunol. 2011, 186, 708–721. [Google Scholar] [CrossRef]
- Yu, C.; Zhang, P.; Li, X.; Sun, L. Japanese flounder Paralichthys olivaceus interleukin 21 induces inflammatory response and plays a vital role in the immune defense against bacterial pathogen. Fish Shellfish Immunol. 2020, 98, 364–373. [Google Scholar] [CrossRef]
- Zhang, A.; Jian, X.; Wang, D.; Ren, J.; Wang, X.; Zhou, H. Characterization and bioactivity of grass carp (Ctenopharyngodon idella) interleukin-21: Inducible production and involvement in inflammatory regulation. Fish Shellfish Immunol. 2020, 99, 19–26. [Google Scholar] [CrossRef]
- Cui, Z.; Zhang, X.; Chen, X.; Zhang, X.; Zhang, Y. Splicing variants of grass carp (Ctenopharyngodon idellus) IL-21: Functions in IgM+ B cell proliferation and IgM secretion. Dev. Comp. Immunol. 2020, 110, 103728. [Google Scholar] [CrossRef]
- Xu, J.; Wang, J.; Min, Q.; Wang, W.; Qin, Y.; Lei, L.; Gao, Q.; Zou, J. Characterisation of IL-21 and IL-21Rα in grass carp: IL-21-producing cells are upregulated during Flavobacterium columnare infection. Aquac. Fish. 2021. [Google Scholar] [CrossRef]
- Li, D.; Cui, Z.; Zhao, F.; Zhu, X.; Tan, A.; Deng, Y.; Lai, Y.; Huang, Z. Characterization of snakehead (Channa argus) interleukin-21: Involvement in immune defense against two pathogenic bacteria, in leukocyte proliferation, and in activation of JAK–STAT signaling pathway. Fish Shellfish Immunol. 2022, 123, 207–217. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Su, J. Interleukin-2 family cytokines: An overview of genes, expression, signaling and functional roles in teleost. Dev. Comp. Immunol. 2023, 141, 104645. [Google Scholar] [CrossRef]
- Rauta, P.R.; Nayak, B.; Das, S. Immune system and immune responses in fish and their role in comparative immunity study: A model for higher organisms. Immunol. Lett. 2012, 148, 23–33. [Google Scholar] [CrossRef] [PubMed]
- Liao, Z.; Su, J. Progresses on three pattern recognition receptor families (TLRs, RLRs and NLRs) in teleost. Dev. Comp. Immunol. 2021, 122, 104131. [Google Scholar] [CrossRef] [PubMed]
- Tang, L.; Wang, G.; Jiang, J.; Feng, L.; Yang, L.; Li, S.; Kuang, S.; Zhou, X. Effect of methionine on intestinal enzymes activities, microflora and humoral immune of juvenile Jian carp (Cyprinus carpio var. Jian). Aquac. Nutr. 2009, 15, 477–483. [Google Scholar] [CrossRef]
- Stosik, M.; Tokarz, D.B.; Deptuła, W. Immunity of the intestinal mucosa in teleost fish. Fish Shellfish Immunol. 2023, 133, 108572. [Google Scholar] [CrossRef]
- Lü, A.; Hu, X.; Zheng, L.; Zhu, A.; Cao, C.; Jiang, J. Isolation and characterization of Citrobacter spp. from the intestine of grass carp Ctenopharyngodon idellus. Aquaculture. 2011, 313, 156–160. [Google Scholar] [CrossRef]
- Song, X.; Zhao, J.; Bo, Y.; Liu, Z.; Wu, K.; Gong, C. Aeromonas hydrophila induces intestinal inflammation in grass carp (Ctenopharyngodon idella): An experimental model. Aquaculture. 2014, 434, 171–178. [Google Scholar] [CrossRef]
- Zhang, C.; Yuan, X.; Xu, R.; Qi, Q.; Wang, Y. The intestinal histopathology, innate immune response and antioxidant capacity of blunt snout bream (Megalobrama amblycephala) in response to Aeromonas hydrophila. Fish Shellfish Immunol. 2022, 124, 525–533. [Google Scholar] [CrossRef]
- Schroers, V.; Marel, D.V.M.; Neuhaus, H.; Steinhagen, D. Changes of intestinal mucus glycoproteins after peroral application of Aeromonas hydrophila to common carp (Cyprinus carpio). Aquaculture 2009, 288, 184–189. [Google Scholar] [CrossRef]
- Pan, C.; Zhu, Y.; Cao, K.; Li, J.; Wang, S.; Zhu, J.; Zeng, X.; Zhang, H.; Qin, Z. Transcriptome, intestinal microbiome and histomorphology profiling of differences in the response of Chinese sea bass (Lateolabrax maculatus) to Aeromonas hydrophila infection. Front. Microbiol. 2023, 14, 1103412. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Wang, J.; Jia, Z.; Feng, J.; Wang, B.; Wang, Z.; Liu, Q.; Wu, K.; Huang, W.; Zhao, X.; et al. Novel dimeric architecture of an IFN-γ–related cytokine provides insights into subfunctionalization of type II IFNs in teleost fish. J. Immunol. 2022, 209, 2203–2214. [Google Scholar] [CrossRef] [PubMed]
- Qin, Y.; Sun, Z.; Wang, W.; Xu, J.; Wang, B.; Jia, Z.; Li, X.; Wang, J.; Gao, Q.; Chen, X.; et al. Characterization of CD3γ/δ+ cells in grass carp (Ctenopharyngodon idella). Dev. Comp. Immunol. 2021, 114, 103791. [Google Scholar] [CrossRef]
- Feng, J.; Jia, Z.; Yuan, G.; Zhu, X.; Liu, Q.; Wu, K.; Wang, J.; Zou, J. Expression and functional characterization of three β-defensins in grass carp (Ctenopharyngodon idella). Dev. Comp. Immunol. 2023, 140, 104616. [Google Scholar] [CrossRef]
- Yuan, G.; Zhu, L.; Jiang, X.; Zhang, J.; Pei, C.; Zhao, X.; Li, L.; Kong, X. Diagnosis of co-infection with white spot syndrome virus and Aeromonas veronii in red swamp crayfish Procambarus clarkii. Aquaculture 2021, 532, 736010. [Google Scholar] [CrossRef]
- Jiang, X.; Gao, J.; Xue, Y.; Qin, Y.; Li, X.; Sun, Z.; Xie, H.; Chang, M.; Nie, P.; Zou, J.; et al. Identification and expression analysis of IL-4/13 receptors in grass carp Ctenopharyngodon idella. Fish Shellfish Immunol. 2019, 87, 254–264. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-Time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Jia, Z.; Feng, J.; Yuan, G.; Xiao, H.; Dang, H.; Zhang, Y.; Chen, K.; Zou, J.; Wang, J. The Meteorin-like cytokine is upregulated in grass carp after infection with Aeromonas hydrophila. Dev. Comp. Immunol. 2023, 141, 104632. [Google Scholar] [CrossRef]
- Wang, J.; Jia, Z.; Dang, H.; Zou, J. Meteorin-like/Meteorin-β upregulates proinflammatory cytokines via NF-κB pathway in grass carp Ctenopharyngodon idella. Dev. Comp. Immunol. 2022, 127, 104289. [Google Scholar] [CrossRef]
- Rombout, J.H.W.M.; Abelli, L.; Picchietti, S.; Scapigliati, G.; Kiron, V. Teleost intestinal immunology. Fish Shellfish Immunol. 2011, 31, 616–626. [Google Scholar] [CrossRef] [PubMed]
- Xiao, H.; Yun, S.; Huang, W.; Dang, H.; Jia, Z.; Chen, K.; Zhao, X.; Wu, Y.; Shi, Y.; Wang, J.; et al. IL-4/13 expressing CD3γ/δ+ T cells regulate mucosal immunity in response to Flavobacterium columnare infection in grass carp. Fish Shellfish Immunol. 2023, 134, 108586. [Google Scholar] [CrossRef] [PubMed]
- Spolski, R.; Kim, H.P.; Zhu, W.; Levy, D.E.; Leonard, W.J. IL-21 Mediates suppressive effects via its induction of IL-10. J. Immunol. 2009, 182, 2859–2867. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Cardoso, J.C.R.; Félix, R.C.; Mateus, A.P.; Canário, A.V.M.; Power, D.M. Fish lysozyme gene family evolution and divergent function in early development. Dev. Comp. Immunol. 2021, 114, 103772. [Google Scholar] [CrossRef]
- Mokhtar, D.M.; Abdelhafez, E.A. An overview of the structural and functional aspects of immune cells in teleosts. Histol. Histopathol. 2021, 36, 399–414. [Google Scholar] [CrossRef]
- FAO. The State of World Fisheries and Aquaculture-Sustainability in Action; Food and Agriculture Organization of the United Nations: Rome, Italy, 2022. [Google Scholar]
- Wang, M.; Guo, J.; Zhao, Y.; Wang, J. IL-21 mediates microRNA-423-5p/claudin-5 signal pathway and intestinal barrier function in inflammatory bowel disease. Aging 2020, 12, 16099–16110. [Google Scholar] [CrossRef]
- Brandt, K.; Singh, P.; Bulfonepaus, S.; Ruckert, R. Interleukin-21: A new modulator of immunity, infection, and cancer. Cytokine Growth Factor Rev. 2007, 18, 223–232. [Google Scholar] [CrossRef]
- Poholek, C.H.; Dulson, S.J.; Zajac, A.J.; Harrington, L.E. IL-21 Controls ILC3 cytokine production and promotes a protective phenotype in a mouse model of colitis. ImmunoHorizons 2019, 3, 194–202. [Google Scholar] [CrossRef]
- Wang, Y.; Jiang, X.; Zhu, J.; Dan, Y.; Zhang, X.; Wang, X.; You, Y.; Wang, B.; Xu, Y.; Lu, C.; et al. IL-21/IL-21R signaling suppresses intestinal inflammation induced by DSS through regulation of Th responses in lamina propria in mice. Sci. Rep. 2016, 6, 31881. [Google Scholar] [CrossRef]
- Fina, D.; Sarra, M.; Fantini, M.C.; Rizzo, A.; Caruso, R.; Caprioli, F.; Stolfi, C.; Cardolini, I.; Dottori, M.; Boirivant, M.; et al. Regulation of gut inflammation and Th17 cell response by interleukin-21. Gastroenterology 2008, 134, 1038–1048.e2. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yuan, G.; Zhao, W.; Zhang, Y.; Jia, Z.; Chen, K.; Wang, J.; Feng, H.; Zou, J. The Biological Functions and Intestinal Inflammation Regulation of IL-21 in Grass Carp (Ctenopharyngodon idella) during Infection with Aeromonas hydrophila. Cells 2023, 12, 2276. https://doi.org/10.3390/cells12182276
Yuan G, Zhao W, Zhang Y, Jia Z, Chen K, Wang J, Feng H, Zou J. The Biological Functions and Intestinal Inflammation Regulation of IL-21 in Grass Carp (Ctenopharyngodon idella) during Infection with Aeromonas hydrophila. Cells. 2023; 12(18):2276. https://doi.org/10.3390/cells12182276
Chicago/Turabian StyleYuan, Gaoliang, Weihua Zhao, Yanwei Zhang, Zhao Jia, Kangyong Chen, Junya Wang, Hao Feng, and Jun Zou. 2023. "The Biological Functions and Intestinal Inflammation Regulation of IL-21 in Grass Carp (Ctenopharyngodon idella) during Infection with Aeromonas hydrophila" Cells 12, no. 18: 2276. https://doi.org/10.3390/cells12182276
APA StyleYuan, G., Zhao, W., Zhang, Y., Jia, Z., Chen, K., Wang, J., Feng, H., & Zou, J. (2023). The Biological Functions and Intestinal Inflammation Regulation of IL-21 in Grass Carp (Ctenopharyngodon idella) during Infection with Aeromonas hydrophila. Cells, 12(18), 2276. https://doi.org/10.3390/cells12182276