
Prognostic Role of Prolactin-Induced Protein (PIP) in Breast Cancer

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
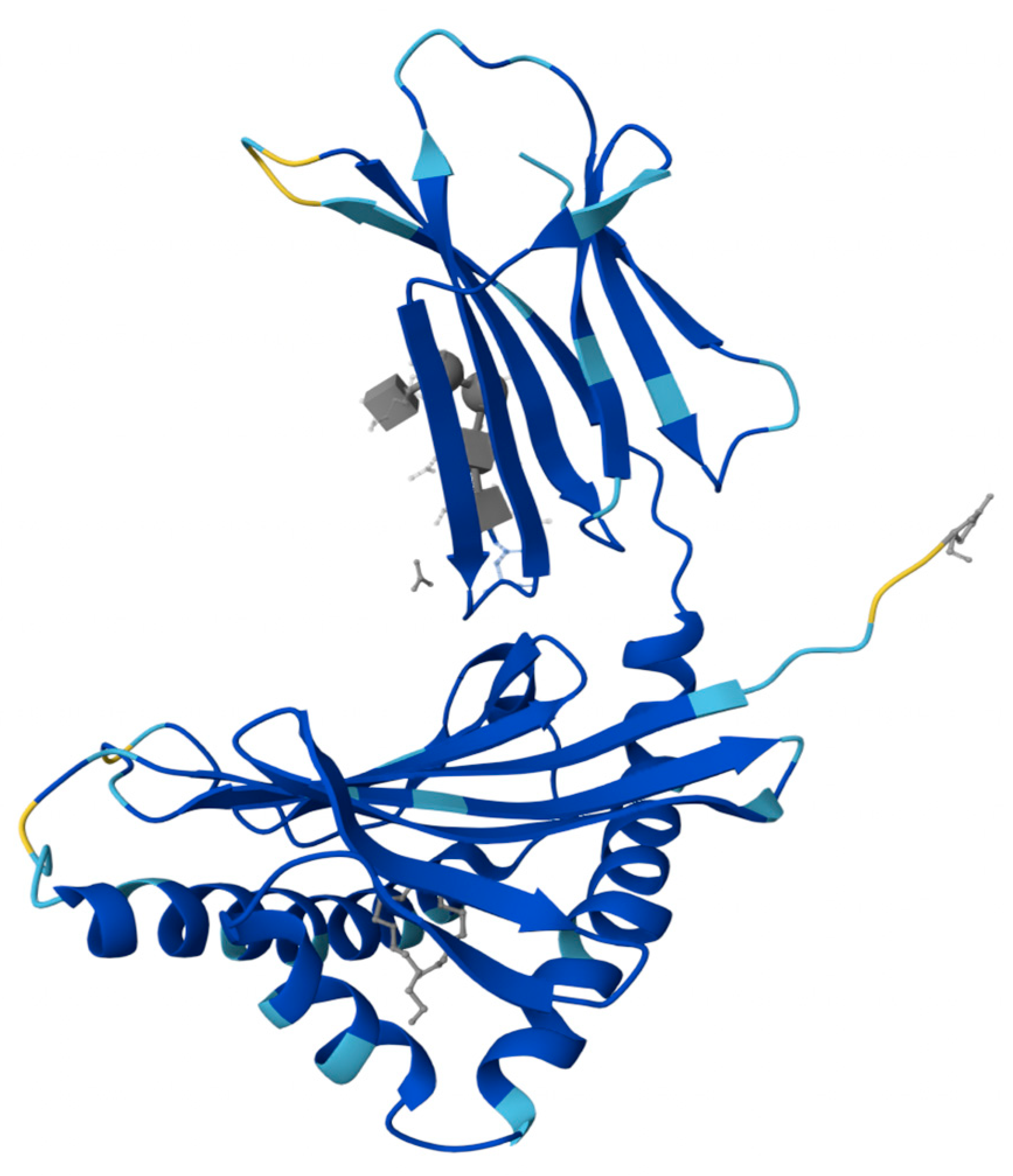
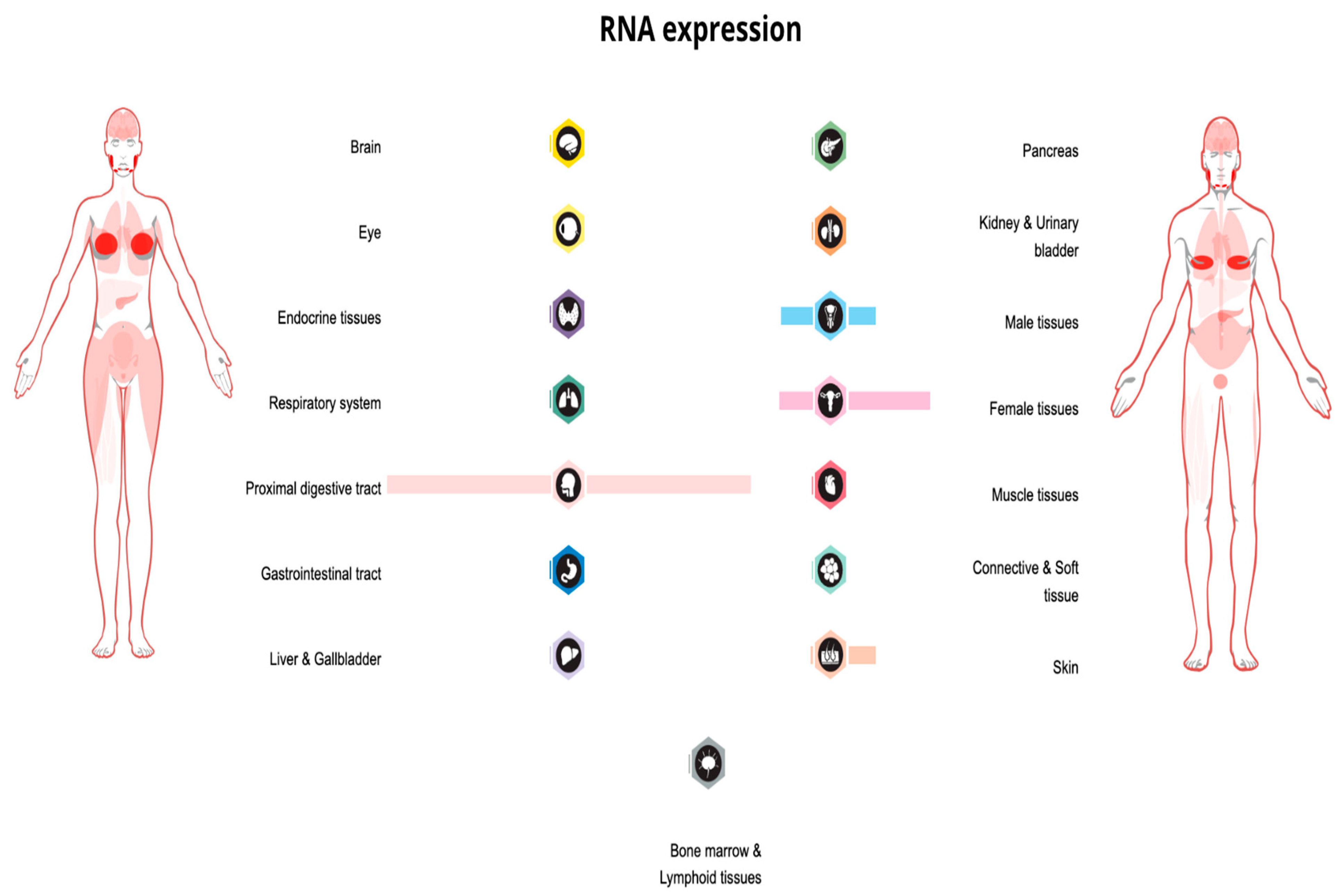
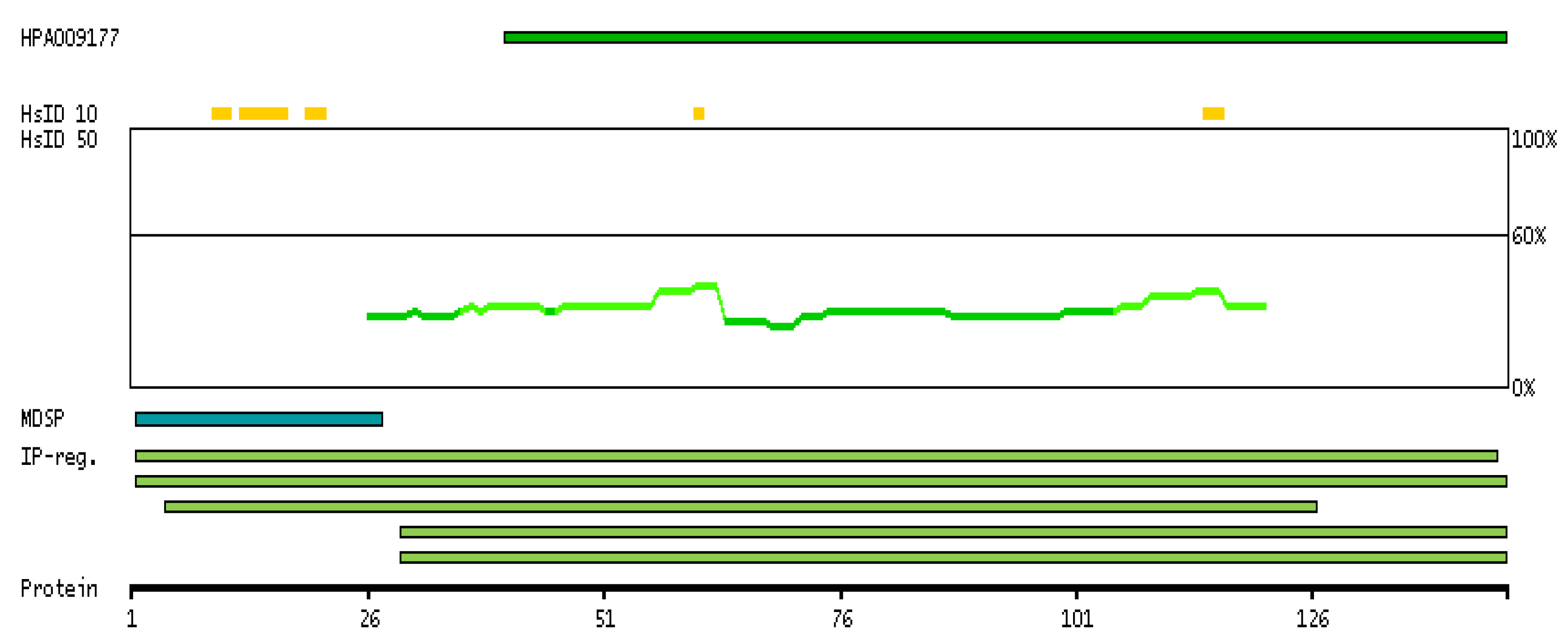
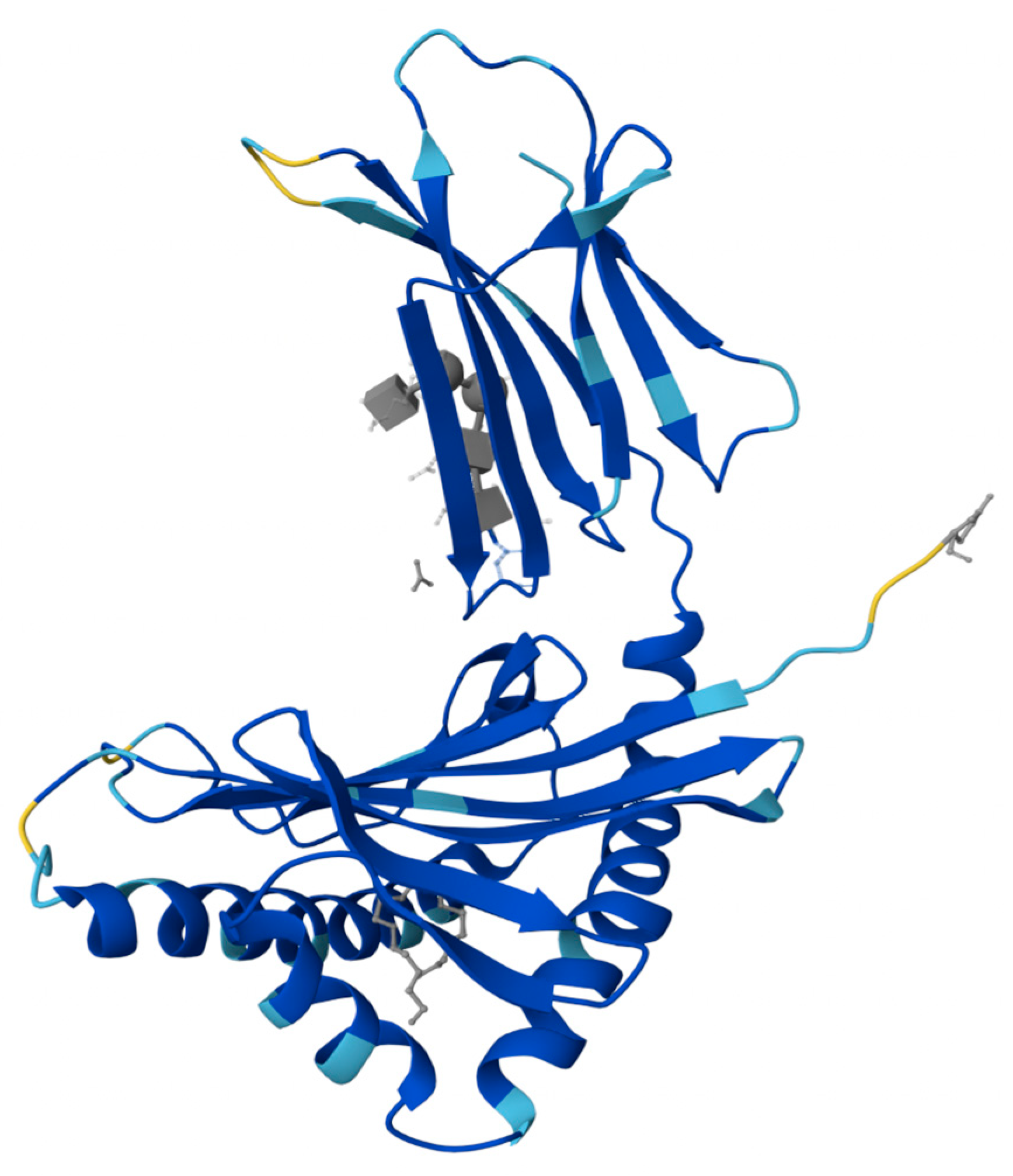
2. PIP: Structure, Expression, and Homeostatic Function
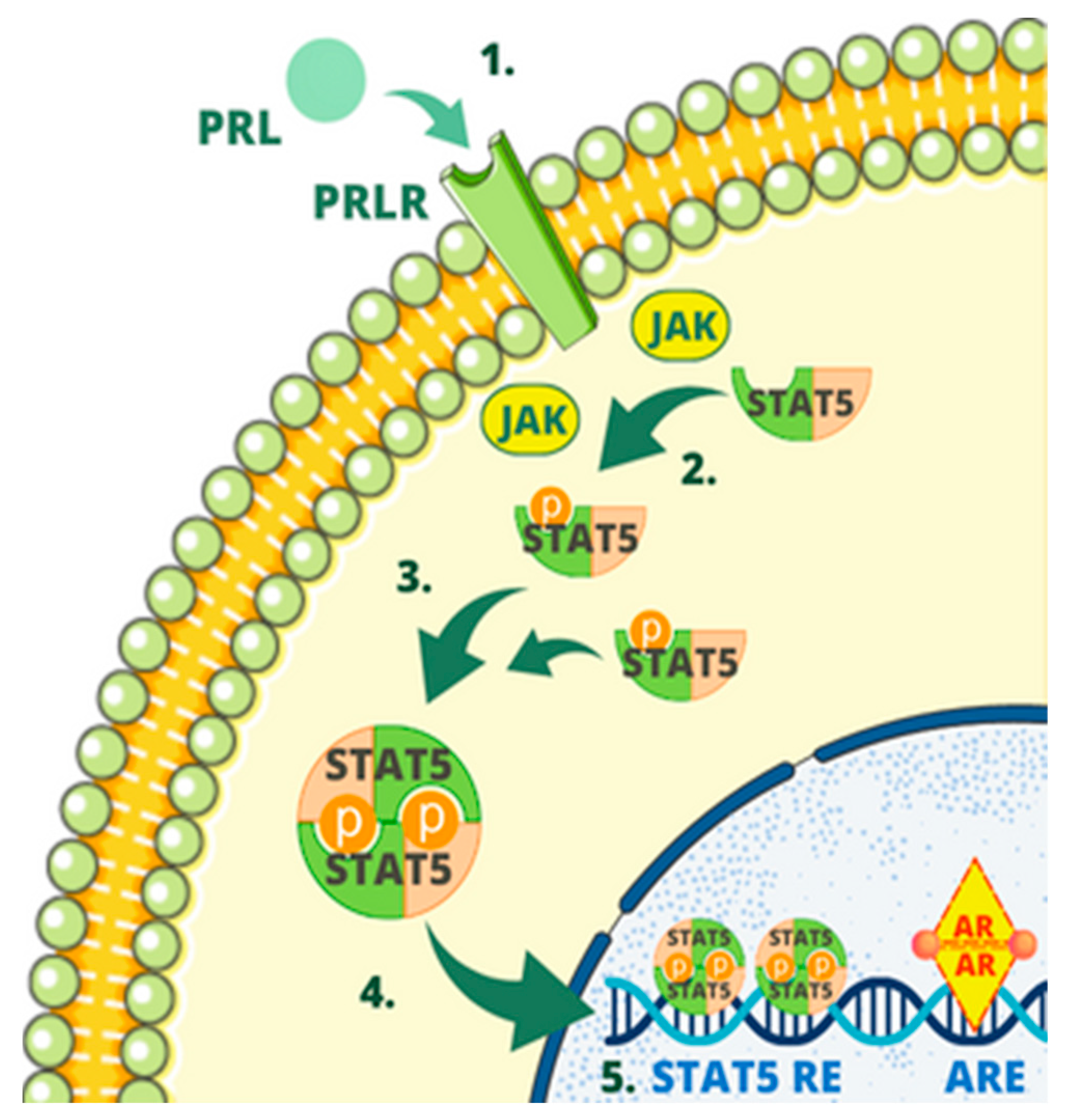
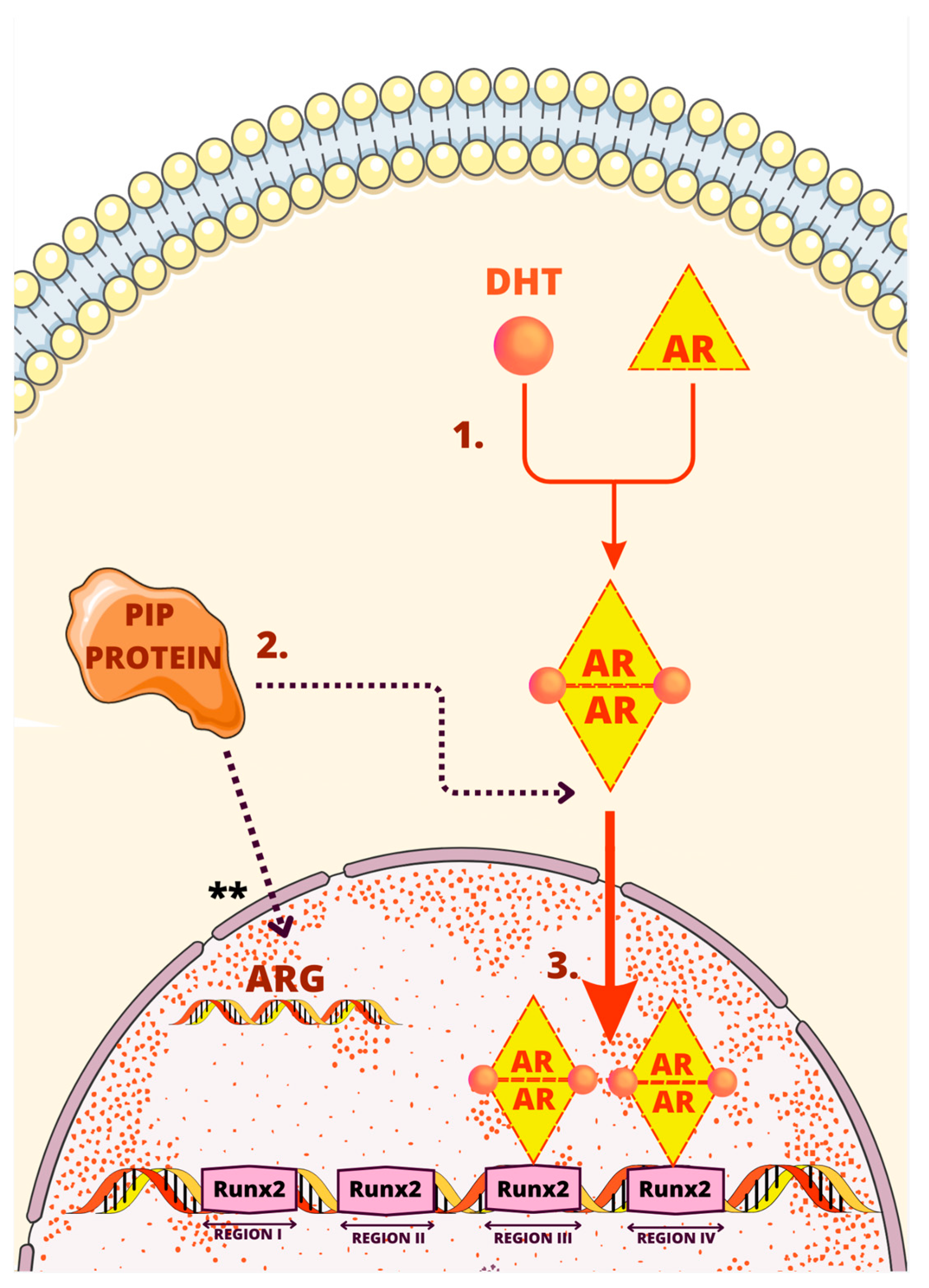
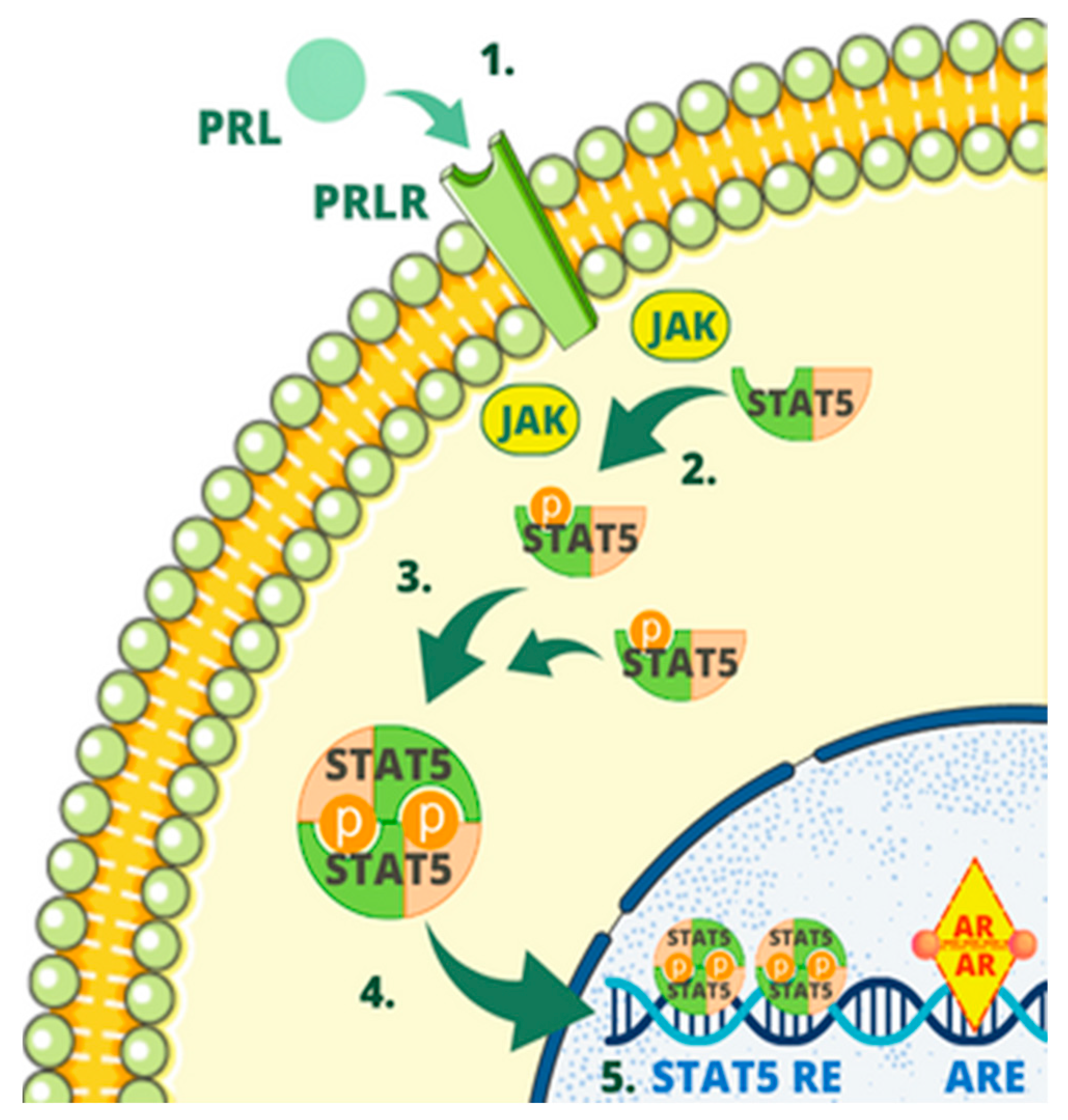
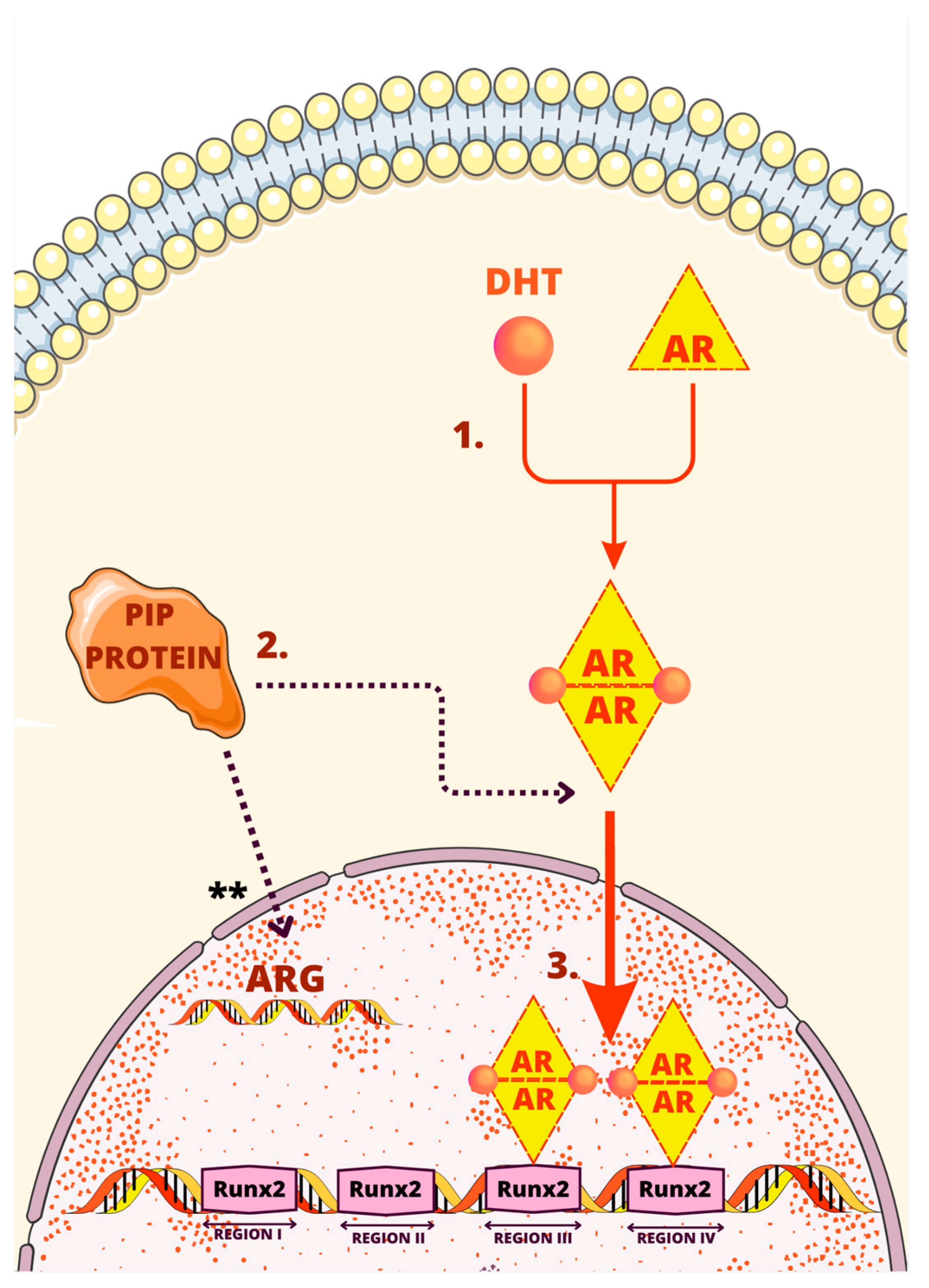
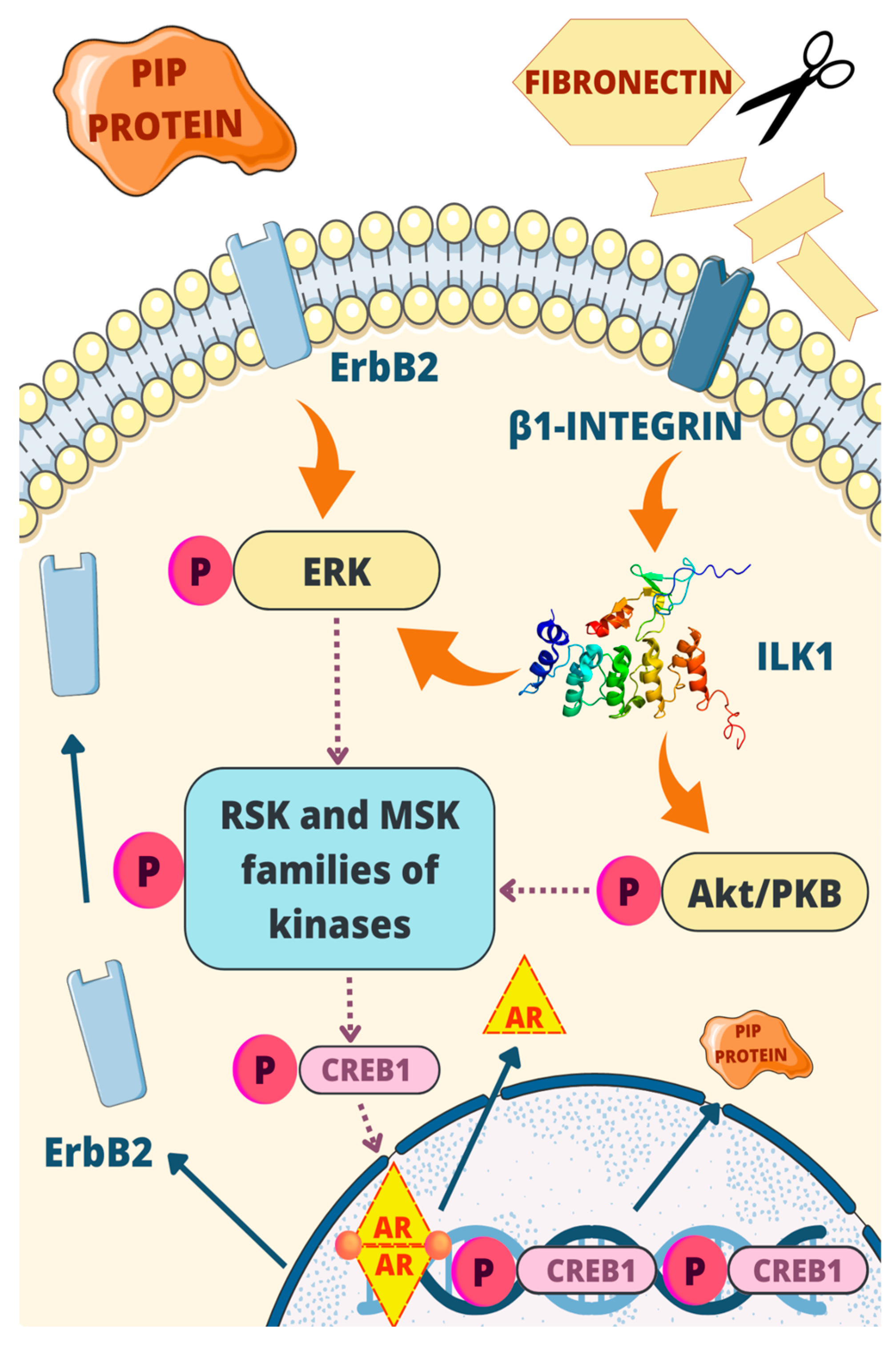
3. Regulatory Mechanisms of the PIP Gene
4. Prognostic Significance of PIP Expression in Breast Cancer
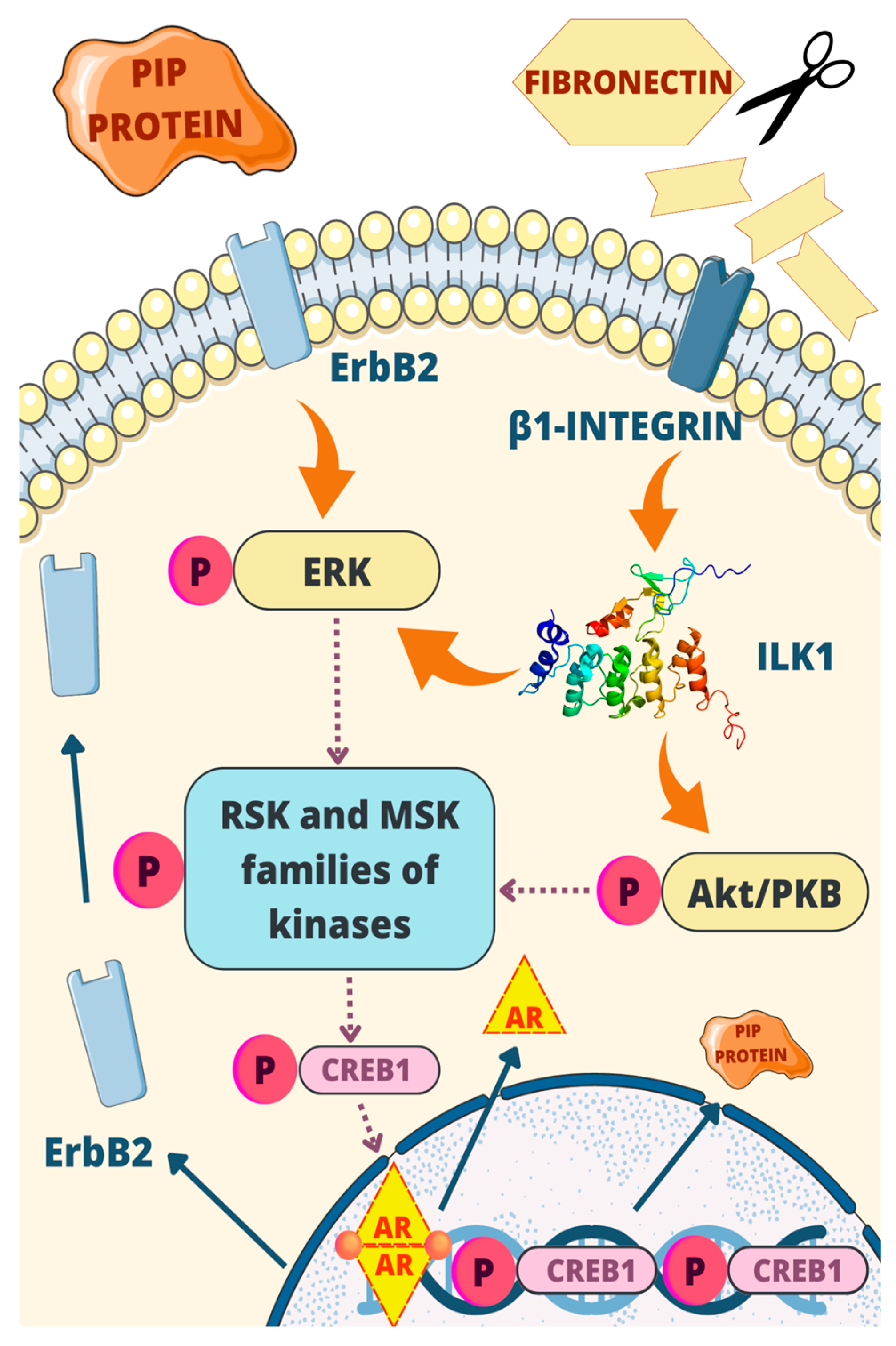
5. Exploring the Impact of PIP in Cell-Based Therapies: Insights from Mouse Models and Tumor Growth Studies
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Umadat, V.; Ihedioha, O.; Shiu, R.; Uzonna, J.; Myal, Y. The prolactin-inducible-protein (PIP): A regulatory molecule in adaptive and innate immunity. Open J. Immunol. 2013, 2013, 210–217. [Google Scholar] [CrossRef]
- Haagensen, J.D.E.; Mazoujian, G. Biochemistry and immunochemistry of gross cystic disease fluid proteins of the breast. Dis. Breast 1986, 474. [Google Scholar]
- Urbaniak, A.; Jablonska, K.; Suchanski, J.; Partynska, A.; Szymczak-Kulus, K.; Matkowski, R.; Maciejczyk, A.; Ugorski, M.; Dziegiel, P. Prolactin-induced protein (PIP) increases the sensitivity of breast cancer cells to drug-induced apoptosis. Sci. Rep. 2023, 13, 6574. [Google Scholar] [CrossRef] [PubMed]
- Jablonska, K.; Grzegrzolka, J.; Podhorska-Okolow, M.; Stasiolek, M.; Pula, B.; Olbromski, M.; Gomulkiewicz, A.; Piotrowska, A.; Rys, J.; Ambicka, A.; et al. Prolactin-induced protein as a potential therapy response marker of adjuvant chemotherapy in breast cancer patients. Am. J. Cancer Res. 2016, 6, 878. [Google Scholar] [PubMed]
- Darb-Esfahani, S.; von Minckwitz, G.; Denkert, C.; Ataseven, B.; Högel, B.; Mehta, K.; Kaltenecker, G.; Rüdiger, T.; Pfitzner, B.; Kittel, K.; et al. Gross cystic disease fluid protein 15 (GCDFP-15) expression in breast cancer subtypes. BMC Cancer 2014, 14, 546. [Google Scholar] [CrossRef]
- Haagensen, D.E.J.; Mazoujian, G.; Dilley, W.G.; Pedersen, C.E.; Kister, S.J.; Wells, S.A.J. Breast gross cystic disease fluid analysis. I. Isolation and radioimmunoassay for a major component protein. J. Natl. Cancer Inst. 1979, 62, 239–247. [Google Scholar] [PubMed]
- Murphy, L.C.; Tsuyuki, D.; Myal, Y.; Shiu, R.P. Isolation and sequencing of a cDNA clone for a prolactin-inducible protein (PIP). Regulation of PIP gene expression in the human breast cancer cell line, T-47D. J. Biol. Chem. 1987, 262, 15236–15241. [Google Scholar] [CrossRef]
- Schaller, J.; Akiyama, K.; Kimura, H.; Hess, D.; Affolter, M.; Rickli, E.E. Primary structure of a new actin-binding protein from human seminal plasma. Eur. J. Biochem. 1991, 196, 743–750. [Google Scholar] [CrossRef]
- Rathman, W.M.; Van Zeyl, M.J.; Van den Keybus, P.A.; Bank, R.A.; Veerman, E.C.; Nieuw Amerongen, A.V. Isolation and characterization of three non-mucinous human salivary proteins with affinity for hydroxyapatite. J. Biol. Buccale 1989, 17, 199–208. [Google Scholar]
- Toth, C.A.; Haagensen, D.E.; Davis, S.; Zamcheck, N.; Thomas, P. Hepatic clearance and metabolism in the rat of a human breast cancer associated glycoprotein (GCDFP-15). Breast Cancer Res. Treat. 1988, 12, 235–243. [Google Scholar] [CrossRef]
- Hassan, M.I.; Bilgrami, S.; Kumar, V.; Singh, N.; Yadav, S.; Kaur, P.; Singh, T.P. Crystal structure of the novel complex formed between zinc alpha2-glycoprotein (ZAG) and prolactin-inducible protein (PIP) from human seminal plasma. J. Mol. Biol. 2008, 384, 663–672. [Google Scholar] [CrossRef] [PubMed]
- Schenkels, L.C.P.M.; Rathman, W.M.; Veerman, E.C.I.; Amerongen, A.V.N. Detection of proteins related to a salivary glycoprotein (EP-GP). Concentrations in human secretions (saliva, sweat, tears, nasal mucus, cerumen, seminal plasma). Biol. Chem. Hoppe. Seyler. 1991, 372, 325–330. [Google Scholar] [CrossRef] [PubMed]
- Mazoujian, G.; Pinkus, G.S.; Davis, S.; Haagensen, D.E. Immunohistochemistry of a gross cystic disease fluid protein (GCDFP-15) of the breast. A marker of apocrine epithelium and breast carcinomas with apocrine features. Am. J. Pathol. 1983, 110, 105. [Google Scholar] [PubMed]
- Jüngert, K.; Paulsen, F.; Jacobi, C.; Horwath-Winter, J.; Garreis, F. Prolactin Inducible Protein, but Not Prolactin, Is Present in Human Tears, Is Involved in Tear Film Quality, and Influences Evaporative Dry Eye Disease. Front. Med. 2022, 9, 892831. [Google Scholar] [CrossRef]
- Haagensen, D.E.; Dilley, W.G.; Mazoujian, G.; Wells, S.A. Review of GCDFP-15. An apocrine marker protein. Ann. N. Y. Acad. Sci. 1990, 586, 161–173. [Google Scholar] [CrossRef]
- Haagensen, D.E.; Gall, S.A.; Brazy, J.E.; Giannola, J.; Wells, S.A. Analysis of amniotic fluid, maternal plasma, and cord blood for a human breast gross cystic disease fluid protein. Am. J. Obstet. Gynecol. 1980, 138, 25–32. [Google Scholar] [CrossRef]
- Chivasso, C.; Nesverova, V.; Järvå, M.; Blanchard, A.; Rose, K.L.; Öberg, F.K.; Wang, Z.; Martin, M.; Lhotellerie, F.; Zindy, E.; et al. Unraveling Human AQP5-PIP Molecular Interaction and Effect on AQP5 Salivary Glands Localization in SS Patients. Cells 2021, 10, 2108. [Google Scholar] [CrossRef]
- Tsubota, K.; Hirai, S.; King, L.S.; Agre, P.; Ishida, N. Defective cellular trafficking of lacrimal gland aquaporin-5 in Sjögren’s syndrome. Lancet 2001, 357, 688–689. [Google Scholar] [CrossRef]
- Terceiro, L.E.L.; Blanchard, A.A.A.; Edechi, C.A.; Freznosa, A.; Triggs-Raine, B.; Leygue, E.; Myal, Y. Generation of prolactin-inducible protein (Pip) knockout mice by CRISPR/Cas9-mediated gene engineering. Can. J. Physiol. Pharmacol. 2022, 100, 86–91. [Google Scholar] [CrossRef]
- Chng, C.L.; Seah, L.L.; Yang, M.; Shen, S.Y.; Koh, S.K.; Gao, Y.; Deng, L.; Tong, L.; Beuerman, R.W.; Zhou, L. Tear Proteins Calcium binding protein A4 (S100A4) and Prolactin Induced Protein (PIP) are Potential Biomarkers for Thyroid Eye Disease. Sci. Rep. 2018, 8, 16936. [Google Scholar] [CrossRef]
- Urbaniak, A.; Jablonska, K.; Podhorska-Okolow, M.; Ugorski, M.; Dziegiel, P. Prolactin-induced protein (PIP)-characterization and role in breast cancer progression. Am. J. Cancer Res. 2018, 8, 2150. [Google Scholar] [PubMed]
- Shiu, R.P.; Iwasiow, B.M. Prolactin-inducible proteins in human breast cancer cells. J. Biol. Chem. 1985, 260, 11307–11313. [Google Scholar] [CrossRef] [PubMed]
- Naderi, A. Prolactin-induced protein in breast cancer. Adv. Exp. Med. Biol. 2015, 846, 189–200. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Zhang, Y.; Jia, L.; Xing, Y.; Zhao, B.; Sui, L.; Liu, D.; Xu, X. Downregulation of Prolactin-Induced Protein Promotes Osteogenic Differentiation of Periodontal Ligament Stem Cells. Med. Sci. Monit. Int. Med. J. Exp. Clin. Res. 2021, 27, e930610. [Google Scholar] [CrossRef] [PubMed]
- Priyadarsini, S.; Hjortdal, J.; Sarker-Nag, A.; Sejersen, H.; Asara, J.M.; Karamichos, D. Gross cystic disease fluid protein-15/prolactin-inducible protein as a biomarker for keratoconus disease. PLoS ONE 2014, 9, e113310. [Google Scholar] [CrossRef]
- Kamiya, K.; Sakabe, J.I.; Yamaguchi, H.; Suzuki, T.; Yatagai, T.; Aoshima, M.; Ito, T.; Tokura, Y. Gross cystic disease fluid protein 15 in stratum corneum is a potential marker of decreased eccrine sweating for atopic dermatitis. PLoS ONE 2015, 10, e0125082. [Google Scholar] [CrossRef] [PubMed]
- Gonda, X.; Eszlari, N.; Torok, D.; Gal, Z.; Bokor, J.; Millinghoffer, A.; Baksa, D.; Petschner, P.; Antal, P.; Breen, G.; et al. Genetic underpinnings of affective temperaments: A pilot GWAS investigation identifies a new genome-wide significant SNP for anxious temperament in ADGRB3 gene. Transl. Psychiatry 2021, 11, 337. [Google Scholar] [CrossRef]
- Caputo, E.; Manco, G.; Mandrich, L.; Guardiola, J. A novel aspartyl proteinase from apocrine epithelia and breast tumors. J. Biol. Chem. 2000, 275, 7935–7941. [Google Scholar] [CrossRef]
- Naderi, A.; Meyer, M. Prolactin-induced protein mediates cell invasion and regulates integrin signaling in estrogen receptor-negative breast cancer. Breast Cancer Res. 2012, 14, R111. [Google Scholar] [CrossRef]
- Schenkels, L.C.P.M.; Walgreen-Weterings, E.; Veerman, E.C.I.; Nieuw Amerongen, A.V.; Schaller, J.; Schadee-Eestermans, I.L. Identity of human extra parotid glycoprotein (EP-GP) with secretory actin binding protein (SABP) and its biological properties. Biol. Chem. Hoppe. Seyler. 1994, 375, 609–616. [Google Scholar] [CrossRef]
- Schenkels, L.C.; Ligtenberg, A.J.; Veerman, E.C.; Van Nieuw Amerongen, A. Interaction of the salivary glycoprotein EP-GP with the bacterium Streptococcus salivarius HB. J. Dent. Res. 1993, 72, 1559–1565. [Google Scholar] [CrossRef] [PubMed]
- Schenkels, L.C.P.M.; Walgreen-Weterings, E.; Oomen, L.C.J.M.; Bolscher, J.G.M.; Veerman, E.C.I.; Nieuw Amerongen, A.V. In vivo binding of the salivary glycoprotein EP-GP (identical to GCDFP-15) to oral and non-oral bacteria detection and identification of EP-GP binding species. Biol. Chem. 1997, 378, 83–88. [Google Scholar] [CrossRef] [PubMed]
- Caputo, E.; Camarca, A.; Moharram, R.; Tornatore, P.; Thatcher, B.; Guardiola, J.; Martin, B.M. Structural study of GCDFP-15/gp17 in disease versus physiological conditions using a proteomic approach. Biochemistry 2003, 42, 6169–6178. [Google Scholar] [CrossRef] [PubMed]
- Bergamo, P.; Balestrieri, M.; Cammarota, G.; Guardiola, J.; Abrescia, P. CD4-mediated anchoring of the seminal antigen gp17 onto the spermatozoon surface. Hum. Immunol. 1997, 58, 30–41. [Google Scholar] [CrossRef] [PubMed]
- Autiero, M.; Abrescia, P.; Guardiola, J. Interaction of seminal plasma proteins with cell surface antigens: Presence of a CD4-binding glycoprotein in human seminal plasma. Exp. Cell Res. 1991, 197, 268–271. [Google Scholar] [CrossRef]
- Kitano, T.; Tian, W.; Umetsu, K.; Yuasa, I.; Yamazaki, K.; Saitou, N.; Osawa, M. Origin and evolution of gene for prolactin-induced protein. Gene 2006, 383, 64–70. [Google Scholar] [CrossRef]
- Basmaciogullari, S.; Autiero, M.; Culerrier, R.; Mani, J.C.; Gaubin, M.; Mishal, Z.; Guardiola, J.; Granier, C.; Piatier-Tonneau, D. Mapping the CD4 binding domain of gp17, a glycoprotein secreted from seminal vesicles and breast carcinomas. Biochemistry 2000, 39, 5332–5340. [Google Scholar] [CrossRef]
- Gaubin, M.; Autiero, M.; Basmaciogullari, S.; Métivier, D.; Misëhal, Z.; Culerrier, R.; Oudin, A.; Guardiola, J.; Piatier-Tonneau, D. Potent inhibition of CD4/TCR-mediated T cell apoptosis by a CD4-binding glycoprotein secreted from breast tumor and seminal vesicle cells. J. Immunol. 1999, 162, 2631–2638. [Google Scholar] [CrossRef]
- Li, J.; Liu, D.; Mou, Z.; Ihedioha, O.C.; Blanchard, A.; Jia, P.; Myal, Y.; Uzonna, J.E. Deficiency of prolactin-inducible protein leads to impaired Th1 immune response and susceptibility to Leishmania major in mice. Eur. J. Immunol. 2015, 45, 1082–1091. [Google Scholar] [CrossRef]
- Blanchard, A.; Nistor, A.; Castaneda, F.E.; Martin, D.; Hicks, G.G.; Amara, F.; Shiu, R.P.C.; Myal, Y. Generation and initial characterization of the prolactin-inducible protein (PIP) null mouse: Accompanying global changes in gene expression in the submandibular gland. Can. J. Physiol. Pharmacol. 2009, 87, 859–872. [Google Scholar] [CrossRef]
- Sharif, R.; Bak-Nielsen, S.; Hjortdal, J.; Karamichos, D. Pathogenesis of Keratoconus: The intriguing therapeutic potential of Prolactin-inducible protein. Prog. Retin. Eye Res. 2018, 67, 150–167. [Google Scholar] [CrossRef] [PubMed]
- Hassan, M.I.; Waheed, A.; Yadav, S.; Singh, T.P.; Ahmad, F. Prolactin inducible protein in cancer, fertility and immunoregulation: Structure, function and its clinical implications. Cell. Mol. Life Sci. 2009, 66, 447–459. [Google Scholar] [CrossRef] [PubMed]
- Witkin, S.S.; Richards, J.M.; Bongiovanni, A.M.; Zelikovsky, G. An IgG-Fc binding protein in seminal fluid. Am. J. Reprod. Immunol. 1983, 3, 23–27. [Google Scholar] [CrossRef] [PubMed]
- Lander, E.S.; Linton, L.M.; Birren, B.; Nusbaum, C.; Zody, M.C.; Baldwin, J.; Devon, K.; Dewar, K.; Doyle, M.; Fitzhugh, W.; et al. Initial sequencing and analysis of the human genome. Nature 2001, 409, 860–921. [Google Scholar] [CrossRef]
- Myal, Y.; Gregory, C.; Wang, H.; Hamerton, J.L.; Shiu, R.P.C. The gene for prolactin-inducible protein (PIP), uniquely expressed in exocrine organs, maps to chromosome 7. Somat. Cell Mol. Genet. 1989, 15, 265–270. [Google Scholar] [CrossRef]
- Simard, J.; Hatton, A.C.; Labriet, C.; Dauvoist, S.; Zhao, H.F.; Haagensen, D.E.; Labrie, F. Inhibitory effect of estrogens on GCDFP-15 mRNA levels and secretion in ZR-75-1 human breast cancer cells. Mol. Endocrinol. 1989, 3, 694–702. [Google Scholar] [CrossRef]
- Haagensen, D.E.; Stewart, P.; Dilley, W.G.; Wells, S.A. Secretion of breast gross cystic disease fluid proteins by T47D breast cancer cells in culture--modulation by steroid hormones. Breast Cancer Res. Treat. 1992, 23, 77–86. [Google Scholar] [CrossRef]
- Loos, S.; Schulz, K.D.; Hackenberg, R. Regulation of GCDFP-15 expression in human mammary cancer cells. Int. J. Mol. Med. 1999, 4, 135–140. [Google Scholar] [CrossRef]
- Liu, X.; Robinson, G.W.; Gouilleux, F.; Groner, B.; Hennighausen, L. Cloning and expression of Stat5 and an additional homologue (Stat5b) involved in prolactin signal transduction in mouse mammary tissue. Proc. Natl. Acad. Sci. USA 1995, 92, 8831–8835. [Google Scholar] [CrossRef]
- Carsol, J.L.; Gingras, S.; Simard, J. Synergistic action of prolactin (PRL) and androgen on PRL-inducible protein gene expression in human breast cancer cells: A unique model for functional cooperation between signal transducer and activator of transcription-5 and androgen receptor. Mol. Endocrinol. 2002, 16, 1696–1710. [Google Scholar] [CrossRef]
- Mundlos, S.; Otto, F.; Mundlos, C.; Mulliken, J.B.; Aylsworth, A.S.; Albright, S.; Lindhout, D.; Cole, W.G.; Henn, W.; Knoll, J.H.M.; et al. Mutations involving the transcription factor CBFA1 cause cleidocranial dysplasia. Cell 1997, 89, 773–779. [Google Scholar] [CrossRef] [PubMed]
- Lucero, C.M.J.; Vega, O.A.; Osorio, M.M.; Tapia, J.C.; Antonelli, M.; Stein, G.S.; van Wijnen, A.J.; Galindo, M.A. The Cancer-Related Transcription Factor Runx2 Modulates Cell Proliferation in Human Osteosarcoma Cell Lines. J. Cell. Physiol. 2013, 228, 714–723. [Google Scholar] [CrossRef] [PubMed]
- Inman, C.K.; Shore, P. The osteoblast transcription factor Runx2 is expressed in mammary epithelial cells and mediates osteopontin expression. J. Biol. Chem. 2003, 278, 48684–48689. [Google Scholar] [CrossRef] [PubMed]
- Selvamurugan, N.; Kwok, S.; Partridge, N.C. Smad3 interacts with JunB and Cbfa1/Runx2 for transforming growth factor-beta1-stimulated collagenase-3 expression in human breast cancer cells. J. Biol. Chem. 2004, 279, 27764–27773. [Google Scholar] [CrossRef]
- Onodera, Y.; Miki, Y.; Suzuki, T.; Takagi, K.; Akahira, J.; Sakyu, T.; Watanabe, M.; Inoue, S.; Ishida, T.; Ohuchi, N.; et al. Runx2 in human breast carcinoma: Its potential roles in cancer progression. Cancer Sci. 2010, 101, 2670–2675. [Google Scholar] [CrossRef]
- Ellison, G.; Klinowska, T.; Westwood, R.F.R.; Docter, E.; French, T.; Fox, J.C. Further evidence to support the melanocytic origin of MDA-MB-435. Mol. Pathol. 2002, 55, 294–299. [Google Scholar] [CrossRef]
- Baniwal, S.K.; Little, G.H.; Chimge, N.O.; Frenkel, B. Runx2 Controls a Feed-forward loop between Androgen and Prolactin-induced Protein (PIP) in Stimulating T47D Cell Proliferation. J. Cell. Physiol. 2012, 227, 2276. [Google Scholar] [CrossRef]
- Sandelin, A.; Alkema, W.; Engström, P.; Wasserman, W.W.; Lenhard, B. JASPAR: An open-access database for eukaryotic transcription factor binding profiles. Nucleic Acids Res. 2004, 32, D91. [Google Scholar] [CrossRef]
- Ning, Y.M.; Robins, D.M. AML3/CBFalpha1 is required for androgen-specific activation of the enhancer of the mouse sex-limited protein (Slp) gene. J. Biol. Chem. 1999, 274, 30624–30630. [Google Scholar] [CrossRef]
- Baniwal, S.K.; Khalid, O.; Sir, D.; Buchanan, G.; Coetzee, G.A.; Frenkel, B. Repression of Runx2 by androgen receptor (AR) in osteoblasts and prostate cancer cells: AR binds Runx2 and abrogates its recruitment to DNA. Mol. Endocrinol. 2009, 23, 1203–1214. [Google Scholar] [CrossRef]
- Tabe, Y.; Jin, L.; Tsutsumi-Ishii, Y.; Xu, Y.; McQueen, T.; Priebe, W.; Mills, G.B.; Ohsaka, A.; Nagaoka, I.; Andreeff, M.; et al. Activation of integrin-linked kinase is a critical prosurvival pathway induced in leukemic cells by bone marrow-derived stromal cells. Cancer Res. 2007, 67, 684–694. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.; Dedhar, S. Integrin-linked kinase (ILK) and its interactors: A new paradigm for the coupling of extracellular matrix to actin cytoskeleton and signaling complexes. J. Cell Biol. 2001, 155, 505–510. [Google Scholar] [CrossRef] [PubMed]
- Legate, K.R.; Montañez, E.; Kudlacek, O.; Fässler, R. ILK, PINCH and parvin: The tIPP of integrin signalling. Nat. Rev. Mol. Cell Biol. 2006, 7, 20–31. [Google Scholar] [CrossRef] [PubMed]
- Falcioni, R.; Antonini, A.; Nisticò, P.; Di Stefano, S.; Crescenzi, M.; Natali, P.G.; Sacchi, A. α6β4 and α6β1 integrins associate with ErbB-2 in human carcinoma cell lines. Exp. Cell Res. 1997, 236, 76–85. [Google Scholar] [CrossRef]
- Yu, X.; Miyamoto, S.; Mekada, E. Integrin alpha 2 beta 1-dependent EGF receptor activation at cell-cell contact sites. J. Cell Sci. 2000, 113, 2139–2147. [Google Scholar] [CrossRef]
- Chia, K.M.; Liu, J.; Francis, G.D.; Naderi, A. A feedback loop between androgen receptor and ERK signaling in estrogen receptor-negative breast cancer. Neoplasia 2011, 13, 154–166. [Google Scholar] [CrossRef]
- Clark, J.W.; Snell, L.; Shiu, R.P.C.; Orr, F.W.; Maitre, N.; Vary, C.P.H.; Cole, D.J.; Watson, P.H. The potential role for prolactin-inducible protein (PIP) as a marker of human breast cancer micrometastasis. Br. J. Cancer 1999, 81, 1002–1008. [Google Scholar] [CrossRef]
- Edechi, C.A.; Ikeogu, N.M.; Akaluka, G.N.; Terceiro, L.E.L.; Machado, M.; Salako, E.S.; Barazandeh, A.F.; Kung, S.K.P.; Uzonna, J.E.; Myal, Y. The Prolactin Inducible Protein Modulates Antitumor Immune Responses and Metastasis in a Mouse Model of Triple Negative Breast Cancer. Front. Oncol. 2021, 11, 639859. [Google Scholar] [CrossRef]
- Shirian, F.; Kheradmand, P.; Ranjbari, N.; Shahbazian, H.; Latifi, S.M. Immunoexpression of the GCDFP-15 Marker in Different Grades of Breast Carcinoma. Iran. J. Pathol. 2023, 18, 75–81. [Google Scholar] [CrossRef]
- Ilhan, B.; Emiroğlu, S.; Türkay, R.; Ilhan, R. The role of histopathologic testing on apocrine carcinoma of the breast. Curr. Probl. Cancer 2020, 44, 100501. [Google Scholar] [CrossRef]
- Gangadharan, A.; Nyirenda, T.; Patel, K.; Jaimes-Delgadillo, N.; Coletta, D.; Tanaka, T.; Walland, A.C.; Jameel, Z.; Vedantam, S.; Tang, S.; et al. Prolactin Induced Protein (PIP) is a potential biomarker for early stage and malignant breast cancer. Breast 2018, 39, 101–109. [Google Scholar] [CrossRef] [PubMed]
- Lehmann-Che, J.; Hamy, A.S.; Porcher, R.; Barritault, M.; Bouhidel, F.; Habuellelah, H.; Leman-Detours, S.; de Roquancourt, A.; Cahen-Doidy, L.; Bourstyn, E.; et al. Molecular apocrine breast cancers are aggressive estrogen receptor negative tumors overexpressing either HER2 or GCDFP15. Breast Cancer Res. 2013, 15, R37. [Google Scholar] [CrossRef] [PubMed]
- Debily, M.A.; El Marhomy, S.; Boulanger, V.; Eveno, E.; Mariage-Samson, R.; Camarca, A.; Auffray, C.; Piatier-Tonneau, D.; Imbeaud, S. A functional and regulatory network associated with PIP expression in human breast cancer. PLoS ONE 2009, 4, e4696. [Google Scholar] [CrossRef] [PubMed]
- Baniwal, S.K.; Chimge, N.O.; Jordan, V.C.; Tripathy, D.; Frenkel, B. Prolactin-Induced Protein (PIP) Regulates Proliferation of Luminal A Type Breast Cancer Cells in an Estrogen-Independent Manner. PLoS ONE 2013, 8, e62361. [Google Scholar] [CrossRef] [PubMed]
- Luo, M.H.; Huang, Y.H.; Ni, Y.B.; Tsang, J.Y.S.; Chan, S.K.; Shao, M.M.; Tse, G.M. Expression of mammaglobin and gross cystic disease fluid protein-15 in breast carcinomas. Hum. Pathol. 2013, 44, 1241–1250. [Google Scholar] [CrossRef]
- DeNardo, D.G.; Brennan, D.J.; Rexhepaj, E.; Ruffell, B.; Shiao, S.L.; Madden, S.F.; Gallagher, W.M.; Wadhwani, N.; Keil, S.D.; Junaid, S.A.; et al. Leukocyte complexity predicts breast cancer survival and functionally regulates response to chemotherapy. Cancer Discov. 2011, 1, 54–67. [Google Scholar] [CrossRef]
- Emens, L.A. Breast cancer immunobiology driving immunotherapy: Vaccines and immune checkpoint blockade. Expert Rev. Anticancer Ther. 2012, 12, 1597–1611. [Google Scholar] [CrossRef]
- Ihedioha, O.C.; Shiu, R.P.C.; Uzonna, J.E.; Myal, Y. Prolactin-Inducible Protein: From Breast Cancer Biomarker to Immune Modulator—Novel Insights from Knockout Mice. DNA Cell Biol. 2016, 35, 537–541. [Google Scholar] [CrossRef]
- Kinoshita, M.; Sawabe, M.; Soejima, Y.; Mieno, M.N.; Arai, T.; Honma, N. Gross Cystic Disease Fluid Protein-15 (GCDFP-15) Expression Characterizes Breast Mucinous Carcinomas in Older Women. Diagnostics 2022, 12, 3129. [Google Scholar] [CrossRef]
- de Andrade Natal, R.; Derchain, S.F.; Pavanello, M.; Paiva, G.R.; Sarian, L.O.; Vassallo, J. Expression of unusual immunohistochemical markers in mucinous breast carcinoma. Acta Histochem. 2017, 119, 327–336. [Google Scholar] [CrossRef]
- Provenzano, E.; Gatalica, Z.; Vranic, S. Carcinoma with Apocrine Differentiation; World Health Organization: Geneva, Switzerland, 2019; pp. 172–176. [Google Scholar]
- Bhargava, R.; Beriwal, S.; Dabbs, D.J. Mammaglobin vs. GCDFP-15An Immunohistologic Validation Survey for Sensitivity and Specificity. Am. J. Clin. Pathol. 2007, 127, 103–113. [Google Scholar] [CrossRef]
- Sapino, A.; Papotti, M.; Righi, L.; Cassoni, P.; Chiusa, L.; Bussolati, G. Clinical significance of neuroendocrine carcinoma of the breast. Ann. Oncol. 2001, 12, 115–117. [Google Scholar] [CrossRef]
- Fritzsche, F.R.; Thomas, A.; Winzer, K.J.; Beyer, B.; Dankof, A.; Bellach, J.; Dahl, E.; Dietel, E.; Kristiansen, G. Co-expression and prognostic value of gross cystic disease fluid protein 15 and mammaglobin in primary breast cancer. Histol. Histopathol. 2007, 22, 1221–1230. [Google Scholar] [CrossRef] [PubMed]
- Haagensen, D.E.; Kelly, D.; Bodian, C.A. GCDFP-15 blood levels for stratification of risk of breast cancer development in women with active breast gross cystic disease. Breast 1997, 6, 113–119. [Google Scholar] [CrossRef]
- Park, K.W.; Im, Y.H.; Lee, J.; Kim, E.; Lee, H.; Song, B.G.; Park, J.O.; Kim, K.; Jung, C.W.; Park, Y.S.; et al. Use of GCDFP-15 (BRST-2) as a Specific Immunocytochemical Marker for Diagnosis of Gastric Metastasis of Breast Carcinoma. Cancer Res. Treat. 2003, 35, 460–464. [Google Scholar] [CrossRef] [PubMed]
- Pasquinelli, R.; Barba, P.; Capasso, I.; D’aiuto, M.; D’aiuto, G.; Anzisi, A.M.; De Berardinis, P.; Guardiola, J. Circulating antibodies against the breast tumor marker GCDFP-15/gp17 in mammary carcinoma patients and in patients carrying benign breast conditions. Int. J. Cancer (Pred. Oncol.) 1999, 84, 568–572. [Google Scholar] [CrossRef]
- Bashor, C.J.; Hilton, I.B.; Bandukwala, H.; Smith, D.M.; Veiseh, O. Engineering the next generation of cell-based therapeutics. Nat. Rev. Drug Discov. 2022, 21, 655–675. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sauer, N.; Matkowski, I.; Bodalska, G.; Murawski, M.; Dzięgiel, P.; Calik, J. Prognostic Role of Prolactin-Induced Protein (PIP) in Breast Cancer. Cells 2023, 12, 2252. https://doi.org/10.3390/cells12182252
Sauer N, Matkowski I, Bodalska G, Murawski M, Dzięgiel P, Calik J. Prognostic Role of Prolactin-Induced Protein (PIP) in Breast Cancer. Cells. 2023; 12(18):2252. https://doi.org/10.3390/cells12182252
Chicago/Turabian StyleSauer, Natalia, Igor Matkowski, Grażyna Bodalska, Marek Murawski, Piotr Dzięgiel, and Jacek Calik. 2023. "Prognostic Role of Prolactin-Induced Protein (PIP) in Breast Cancer" Cells 12, no. 18: 2252. https://doi.org/10.3390/cells12182252
APA StyleSauer, N., Matkowski, I., Bodalska, G., Murawski, M., Dzięgiel, P., & Calik, J. (2023). Prognostic Role of Prolactin-Induced Protein (PIP) in Breast Cancer. Cells, 12(18), 2252. https://doi.org/10.3390/cells12182252