
Down-Regulation of ABCA7 in Human Microglia, Astrocyte and THP-1 Cell Lines by Cholesterol Depletion, IL-1β and TNFα, or PMA

 ,
, 
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Lines and Culture
2.2. Cell Cholesterol Depletion
2.3. Modified Amplex Red Assay for Measuring Cell Cholesterol
2.4. Cell Treatment with Mifepristone, LXR Agonist and U18666A
2.5. Cell Treatment with Pro-Inflammatory Cytokines
2.6. Western Immunoblotting
2.7. IL-1β and TNFα ELISA
2.8. Extraction and Re-Analysis of Publicly Available Microarray and RNA Sequencing Data
2.9. Statistical Analysis and Figure Assembly
3. Results
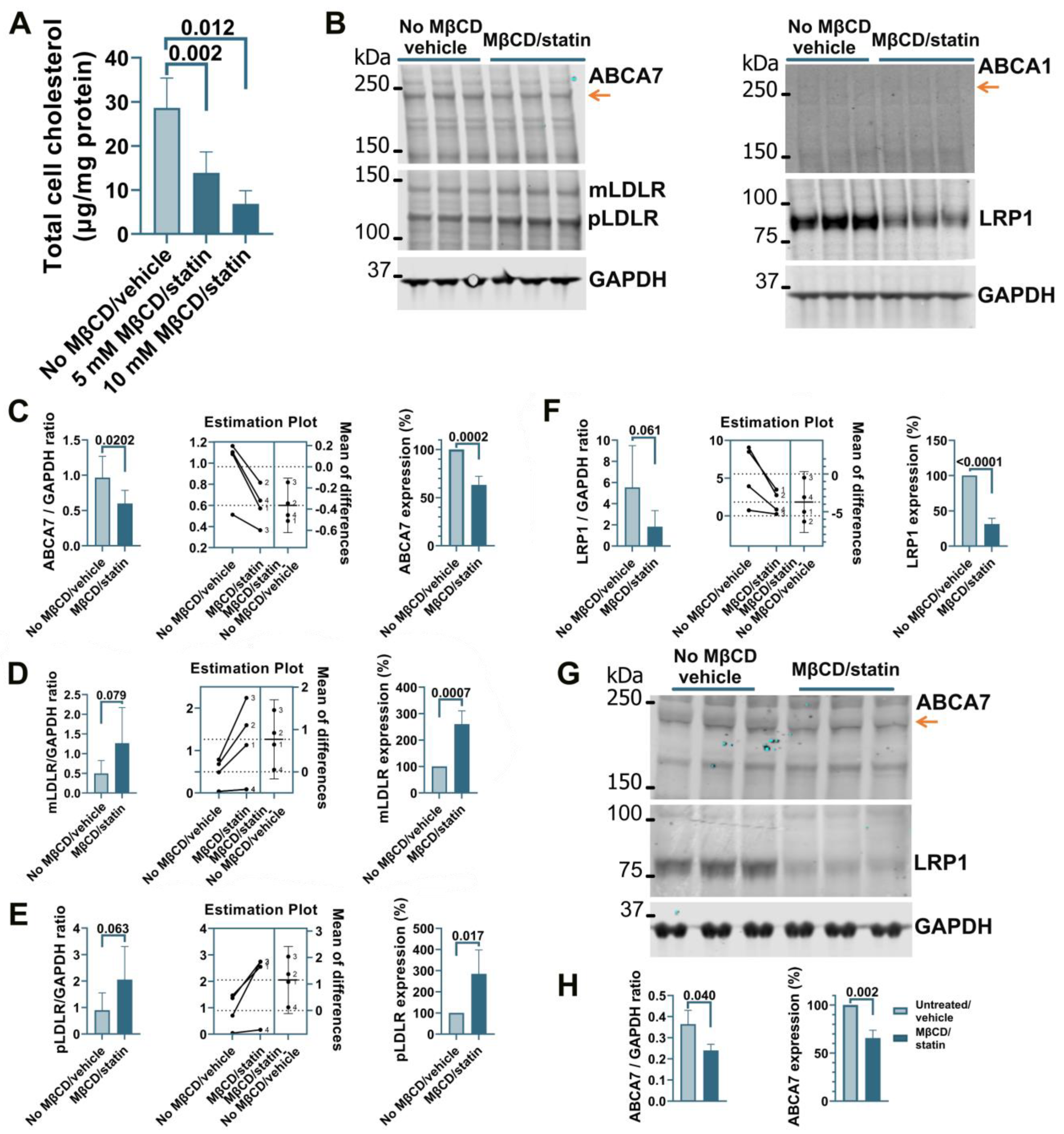
3.1. Cholesterol Depletion Down-Regulates ABCA7 Expression in Human Microglia and Astrocyte but Not Neuronal Cells
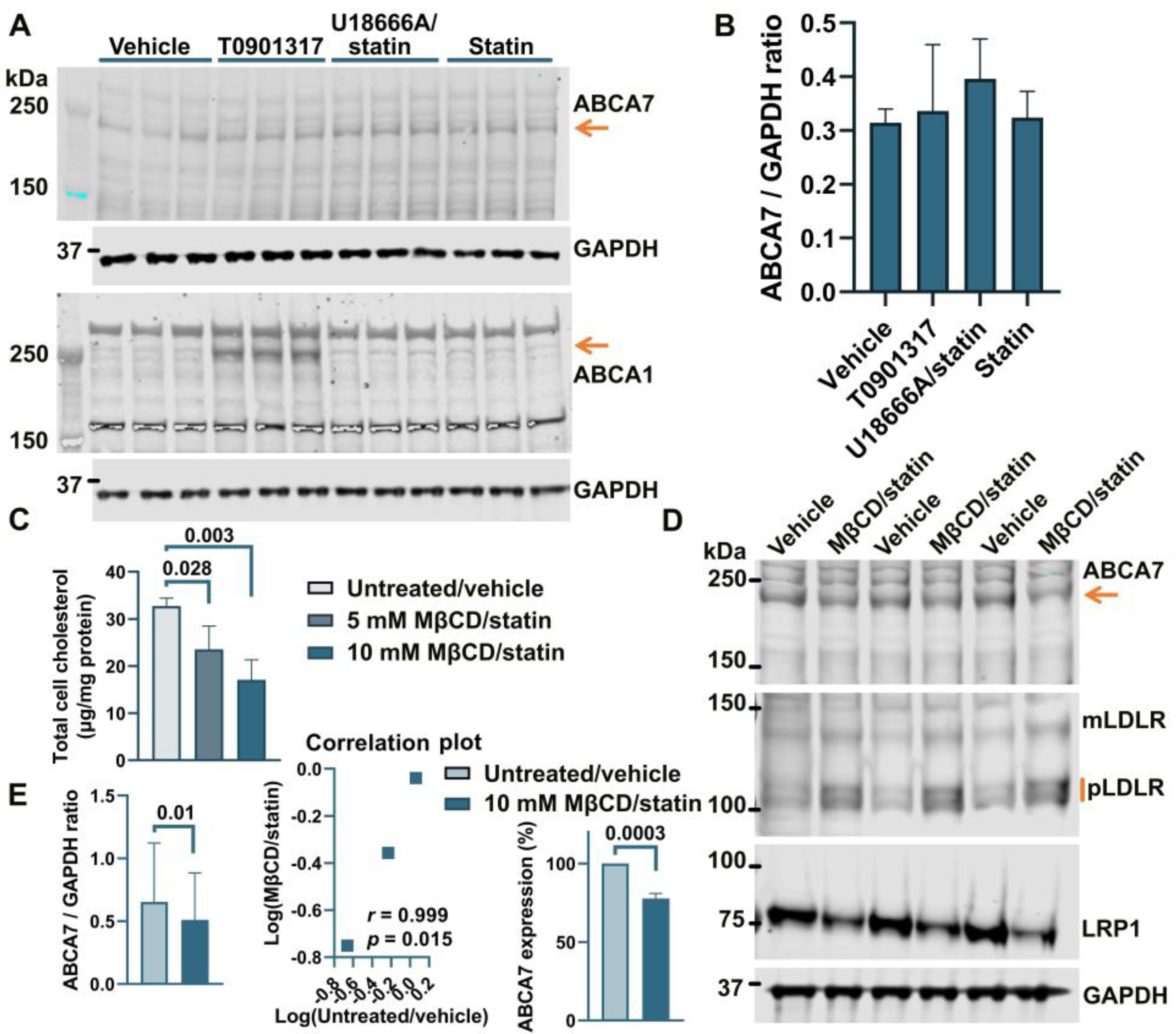
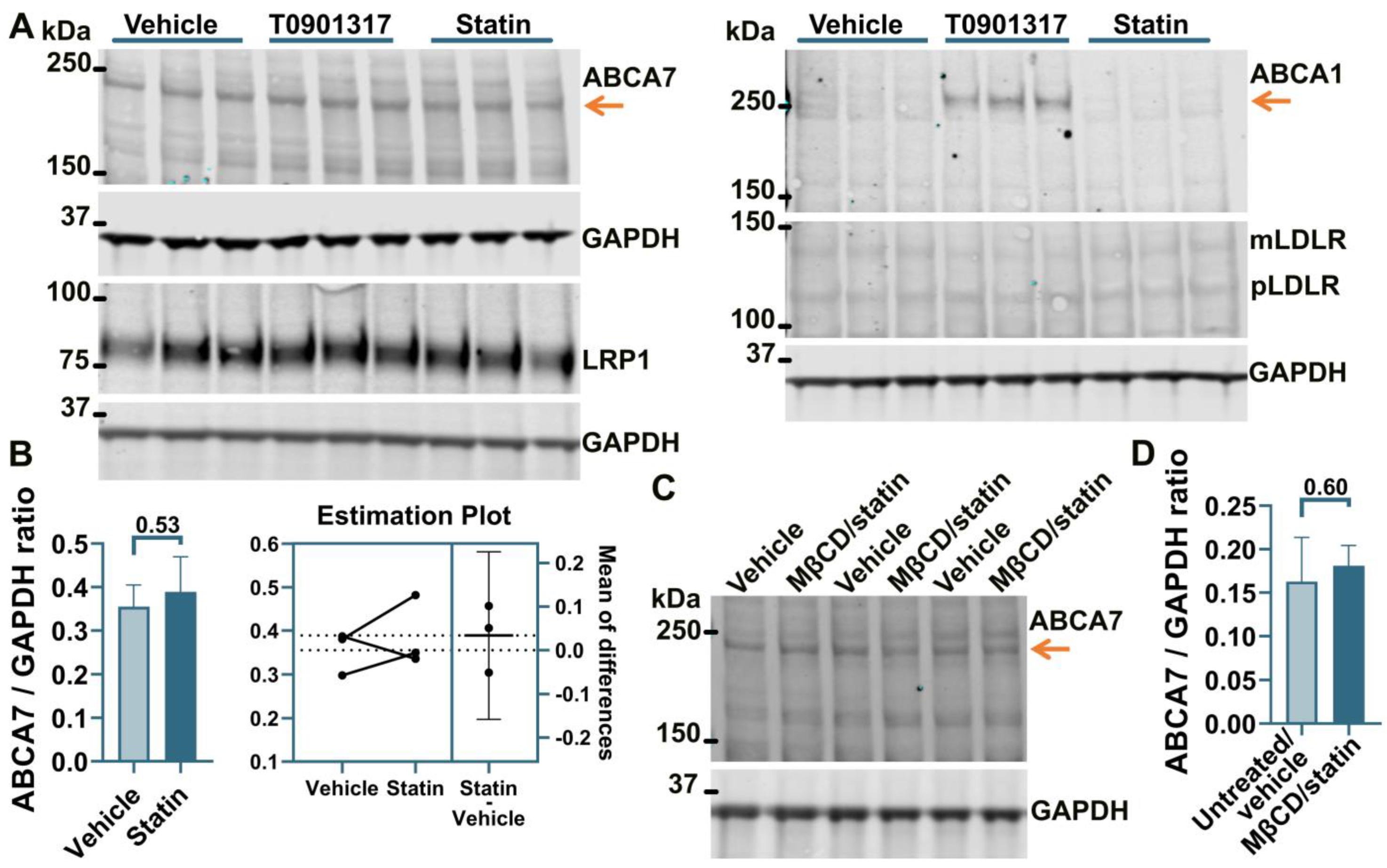
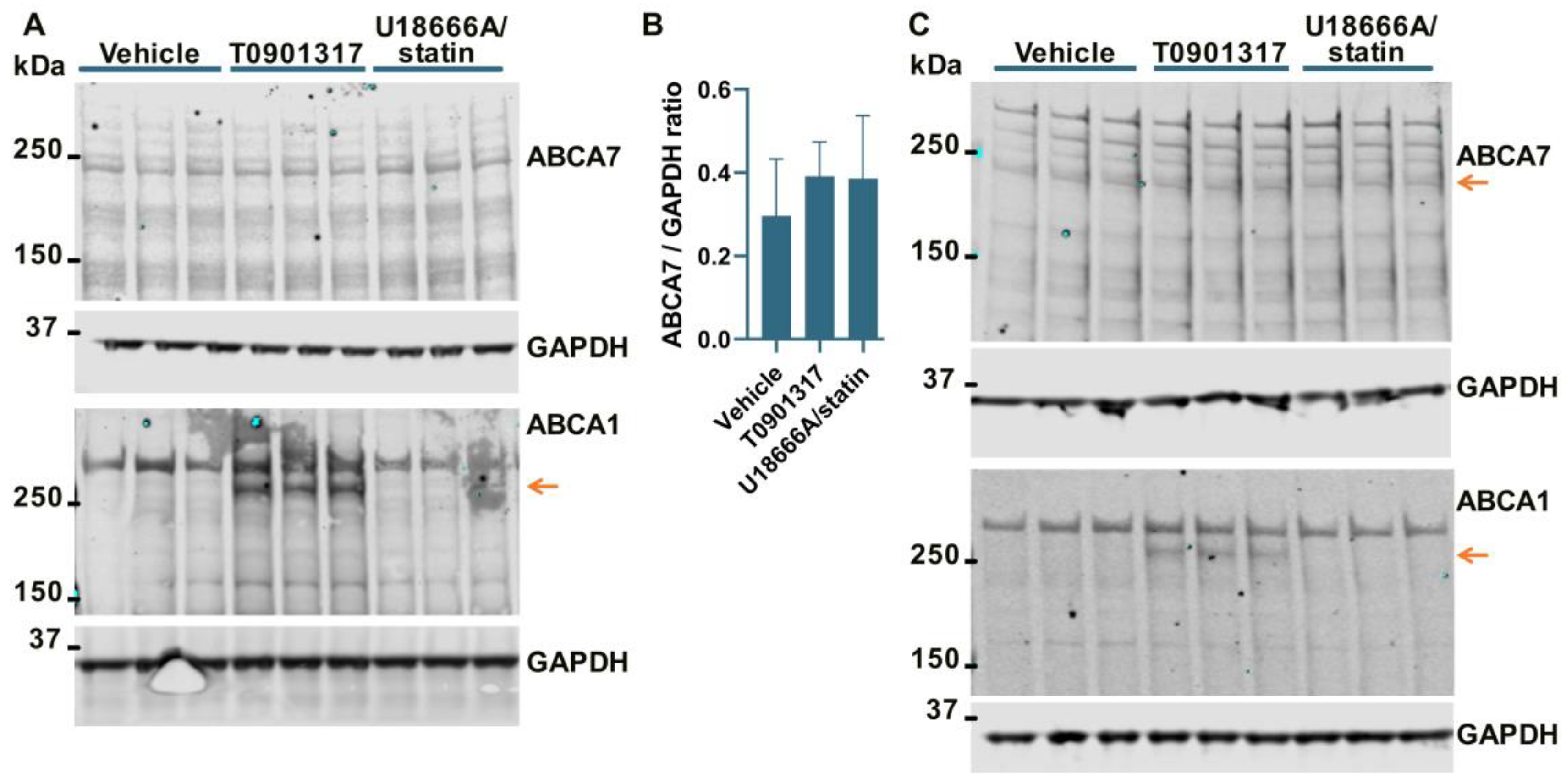
3.2. No Effect of an LXR Agonist or the U18666A/Statin Co-Treatment on ABCA7 Expression in Human Neural Cell Lines
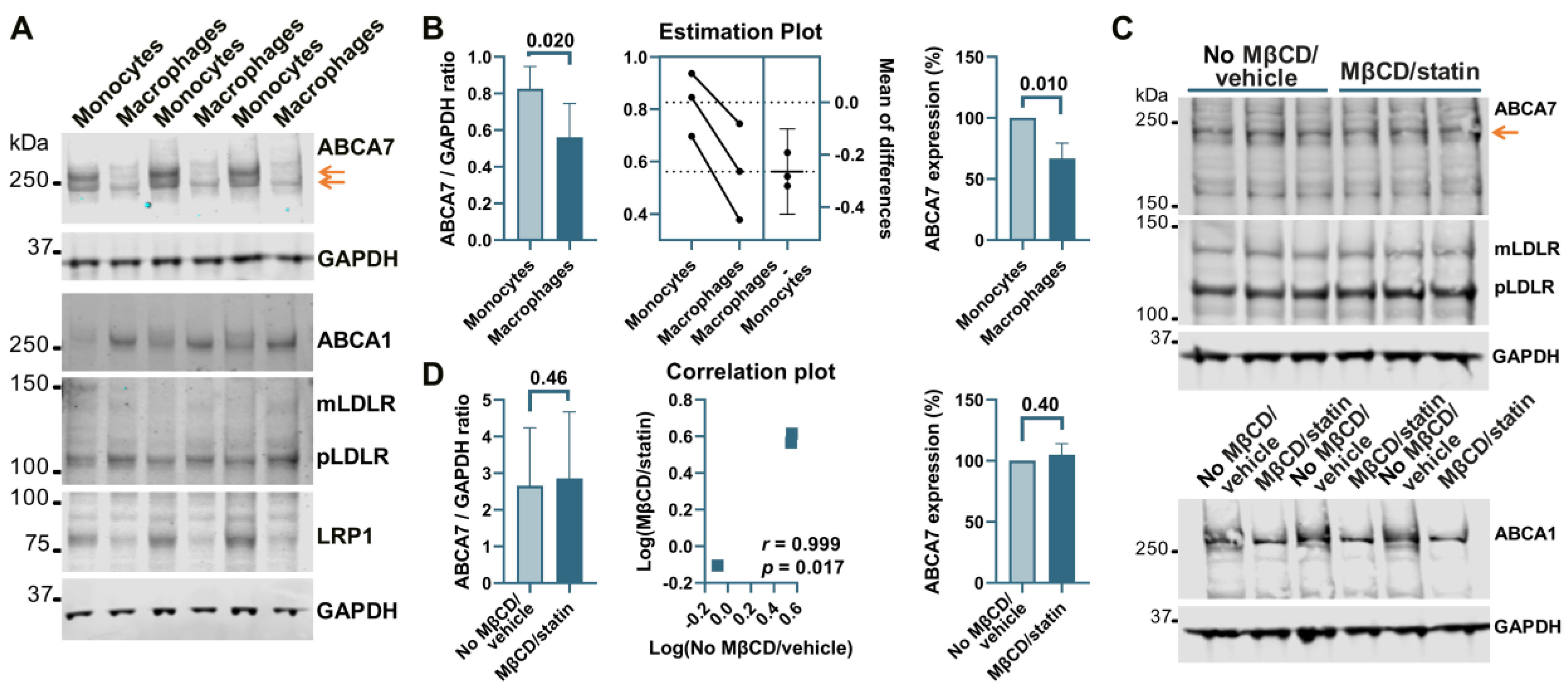
3.3. PMA Treatment to Differentiate THP-1 Monocytes to Macrophages Induces a Reduction in ABCA7 Levels, but Subsequent Cholesterol Depletion in THP-1 Macrophages Has No Effect on ABCA7 Expression
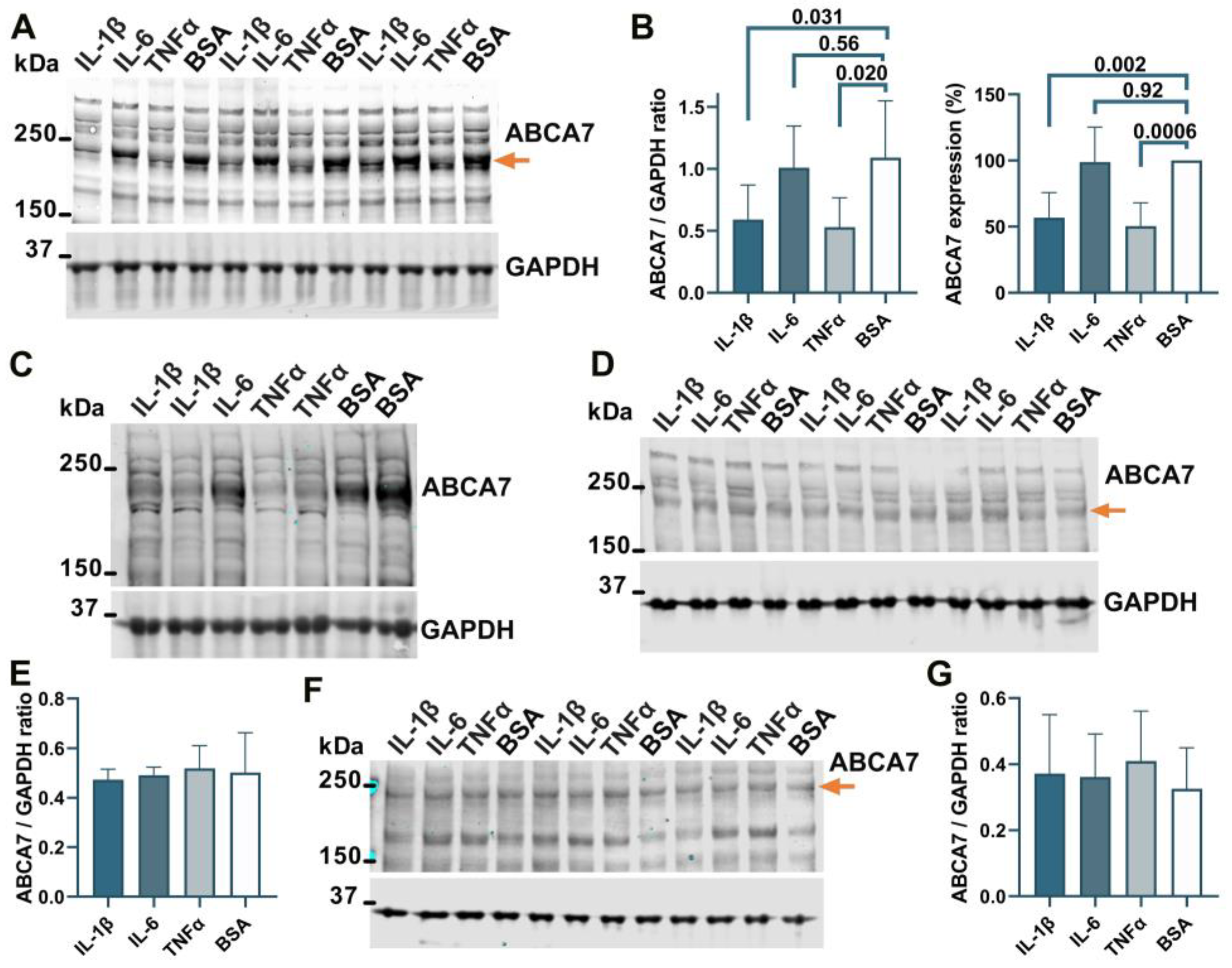
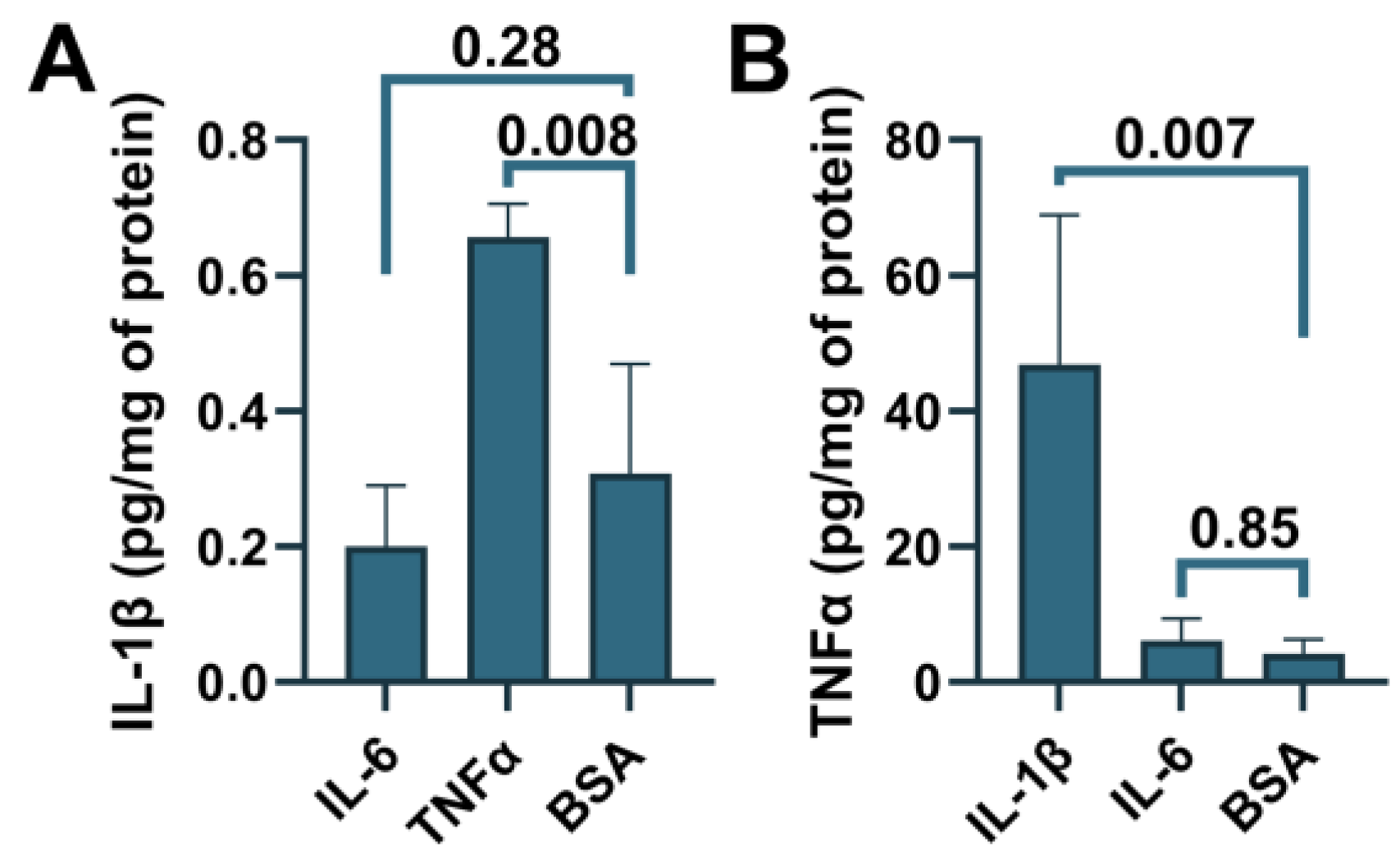
3.4. Suppression of the ABCA7 Protein by IL-1β and TNFα in Microglia but Not Astrocyte or Neuronal Cells
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
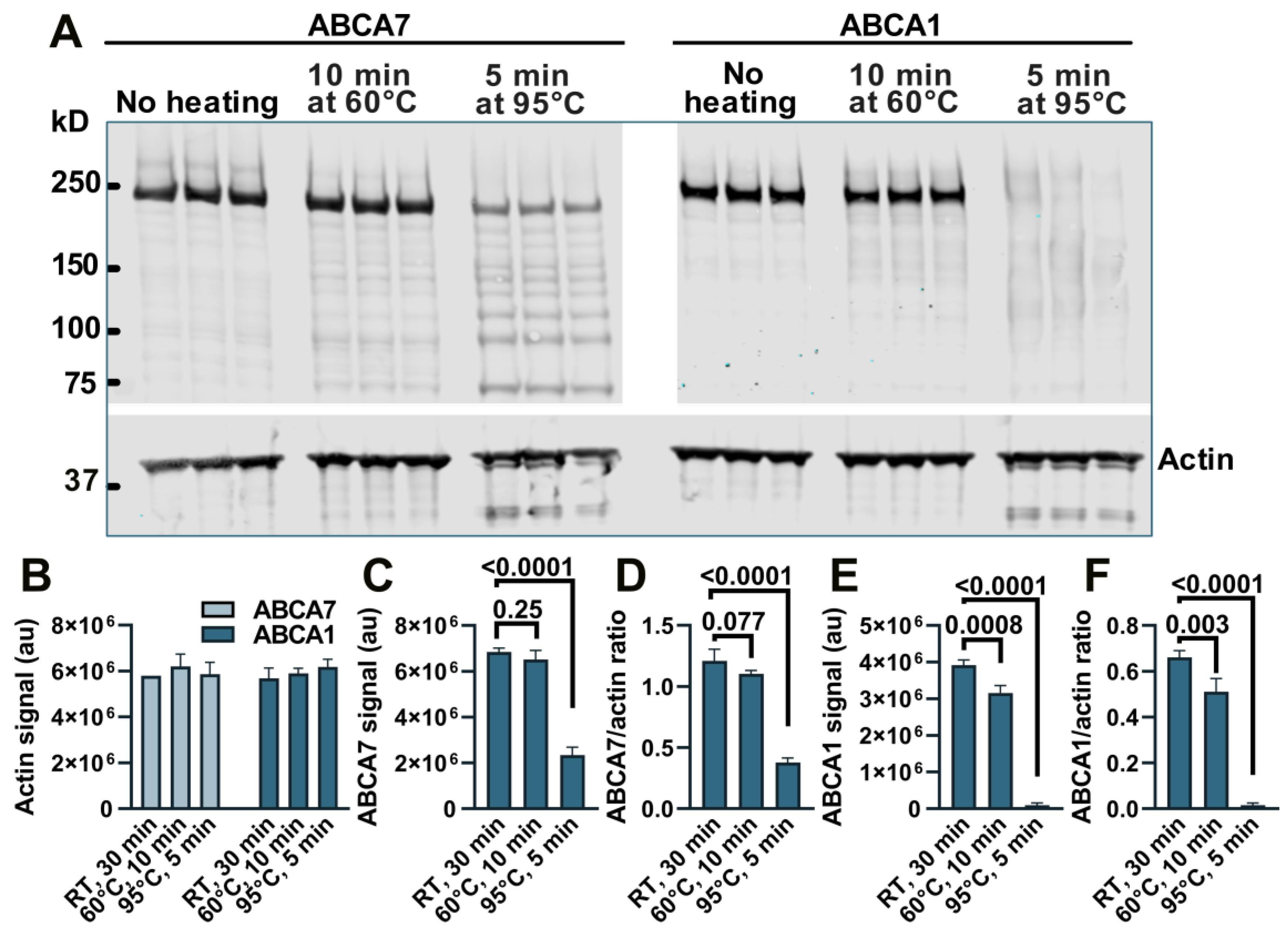
Appendix A. Loss of Immunodetectable ABCA7 and ABCA1 during Heating of Cell Lysates

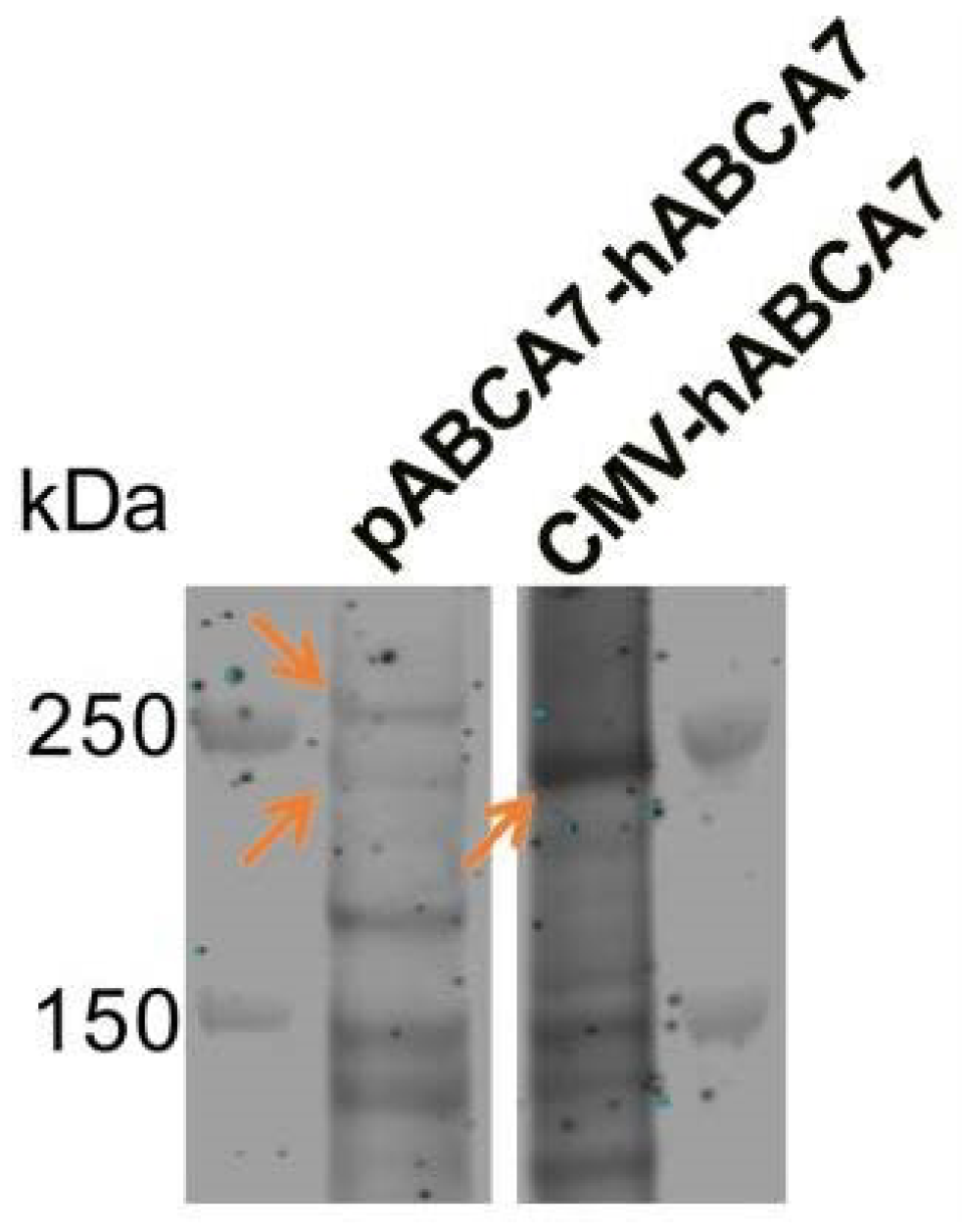
Appendix B. Existence of Two ABCA7 Isoforms

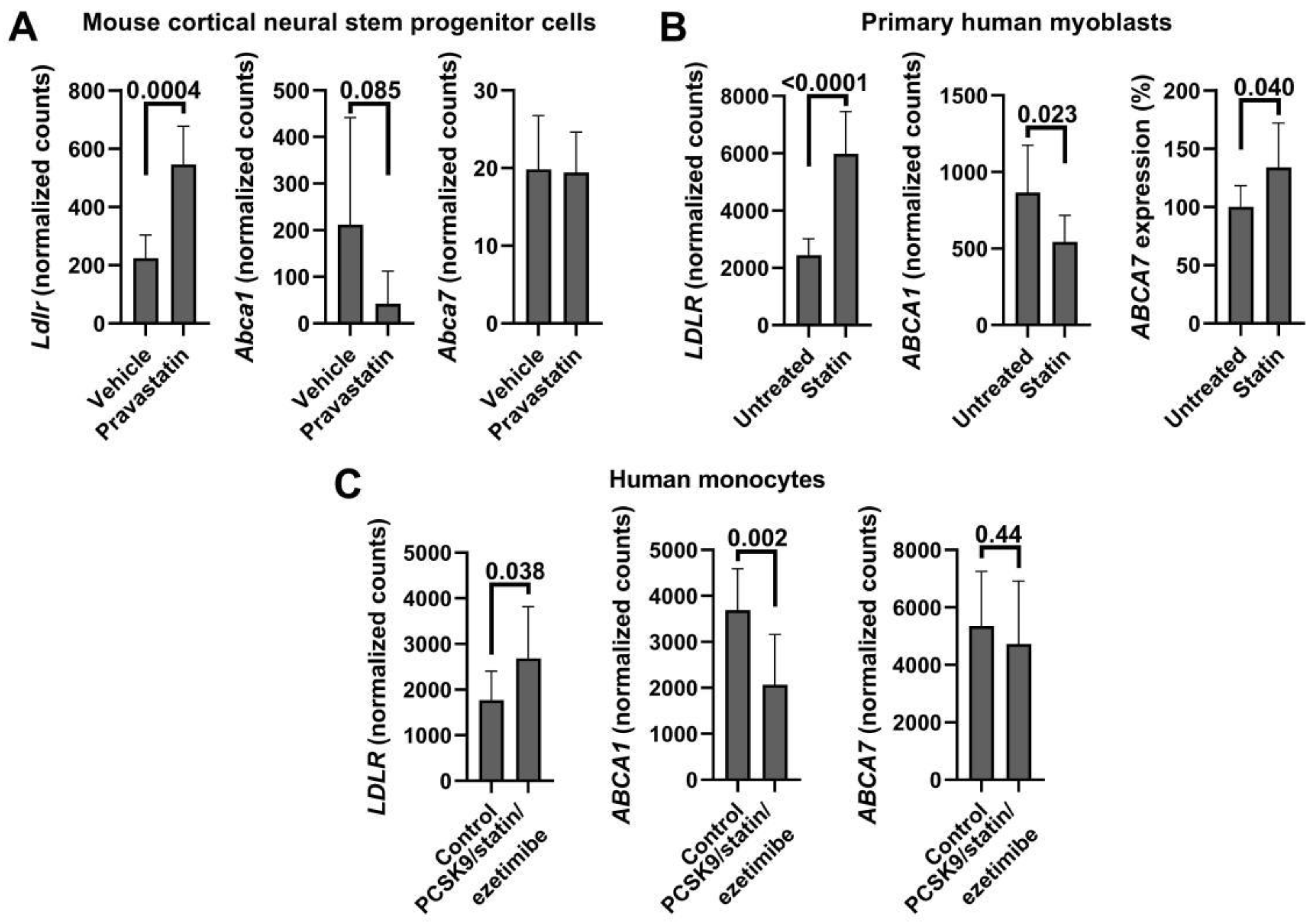
Appendix C. Cholesterol Depletion and ABCA7/Abca7 Expression in Public Datasets

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Reference GEO Accession Number | Tissue Treatment (n) | Effect (Fold Change, p-Value) * | ||
|---|---|---|---|---|
| LDLR/Ldlr | ABCA1/Abca1 | ABCA7/Abca7 | ||
| Lu et al. [18] Not deposited | Human aortic smooth muscle cells 2-Hydroxypropyl-β-cyclodextrin 10 mg/mL vs. vehicle for 24 h (~53% reduction in cholesterol) (4 repeats) | 3.14 0 | −3.57 0 | −0.12 0.80 |
| Seo et al. [19] GSE28084 | C57BL/6 mouse liver Lovastatin 100 mg and ezetimibe 21 mg per 100 g chow vs. powdered cholesterol 1% w/w of chow for one week (3 mice per group) | 1.42 1.02 × 10−7 | −0.28 6.71 × 10−4 | 0.024 0.67 |
| Carson et al. [20] GSE111945 | Mouse cortical neural stem progenitor cells 10 μ pravastatin vs. vehicle for 24 h (6 mice per group) | 2.4 1.9 × 10−7 | −5.0 0.0079 | −1.02 0.89 |
| Chamaria et al. [57] GSE86216 | Human peripheral blood mononuclear cells Baseline vs. 8–12 weeks on 40 mg rosuvastatin daily (72 individuals) | 0.30 5.41 × 10−6 | −0.20 1.59 × 10−5 | −0.028 0.42 |
| Grunwald et al. [58] GSE107998 | Primary human myoblasts 5 µM rosuvastatin vs. vehicle (4 cell populations) | 2.45 9.25 × 10−8 | 0.74 0.094 | 1.16 0.302 |
| 5 µM simvastatin vs. vehicle (4 cell populations) | 2.39 5.04 × 10−7 | 0.53 0.008 | 1.5 0.015 | |
| Willemsen et al. [59] GSE192709 * | Human monocytes Patients with familial hypercholesterolemia treated with a combination of PCSK9 inhibitor, statin and ezetimibe vs. normolipidemic controls (10 per group) | N/A See Figure A3C | ||

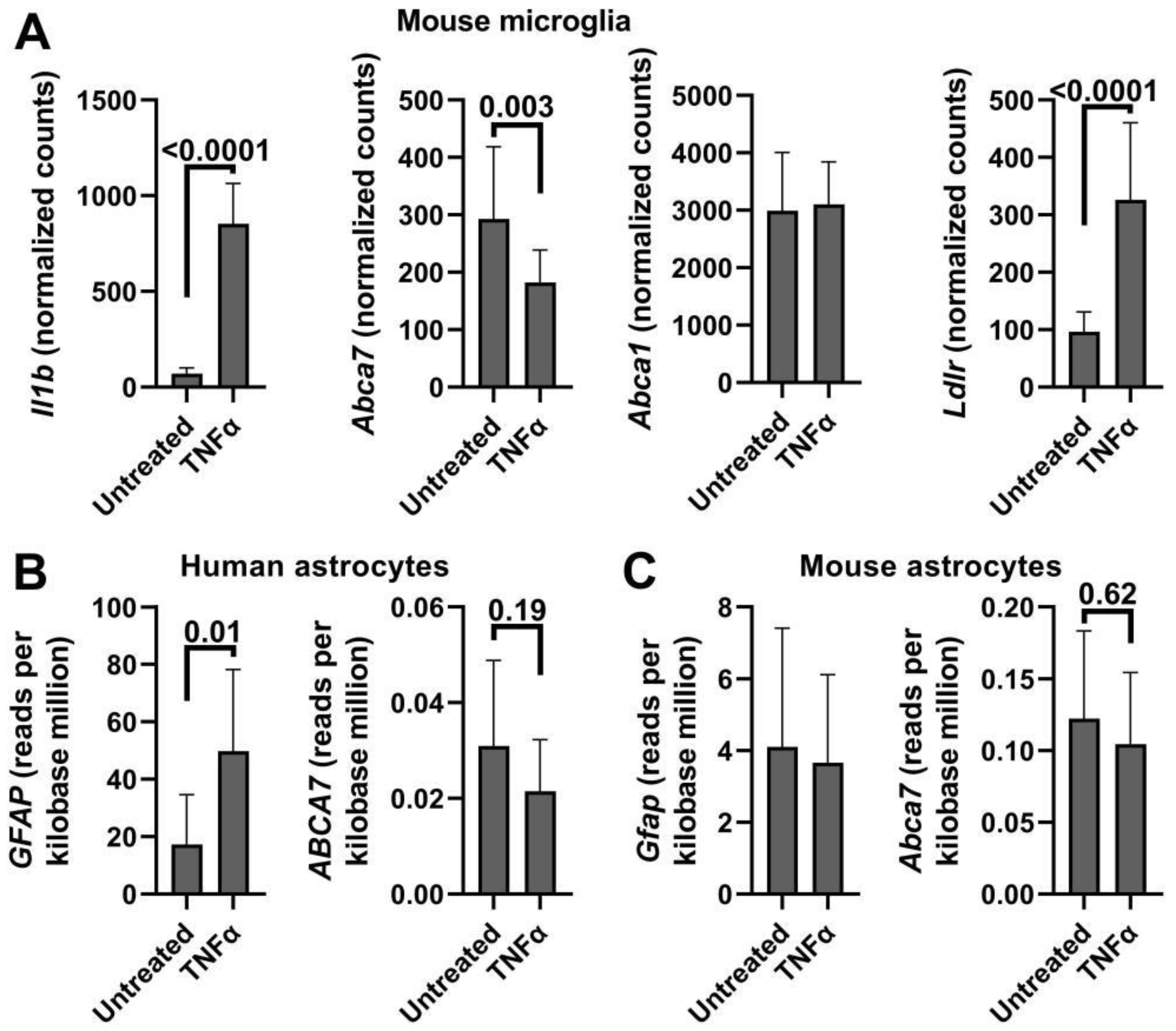
Appendix D. ABCA7/Abca7 Expression in IL-1β- or TNFα-Treated Microglia or Astroglia in Public Gene Expression Datasets
| Reference GEO Accession Number | Tissue Treatment (n) | Effect (Fold Change, p-Value) * | |
|---|---|---|---|
| GFAP/Gfap | ABCA7/Abca7 | ||
| Delbridge et al. [48] GSE154853 | Mouse microglia Untreated vs. 50 ng/mL TNFα for 24 h (n = 9 and 11) | Not applicable | −0.38 0.003 |
| Schneider et al. [61] GSE205610 | Mouse enteric glial cells (a type of astrocyte) Untreated vs. 10 ng/mL IL-1β for 24 h (n = 3) | −1.1 0.87 | −0.04 0.72 |
| Lemaître et al. [62] GSE201555 | Human fetal astrocytes Untreated vs. 20 ng/mL IL-1β for 48 h (n = 4 donors) | 1.1 0.00001 | −0.34 0.041 |
| Li et al. [63] GSE147870 | Human fetal astrocytes Untreated vs. 30 ng/mL TNFα for 48 h (n = 9 donors) | 1.9 0.01 | −0.11 0.62 |
| Mouse astrocytes Untreated vs. 30 ng/mL TNFα for 48 h (n = 6) | −0.11 0.8 | −0.29 0.35 | |
| Birck et al. [64] GSE117736 | Neurospheres obtained from embryonic murine neural stem cells and then differentiated to astrocytes Untreated vs. 50 ng/mL TNFα for 24 h (n = 3) | 0.55 0.013 | 0.24 0.048 |
| Gabel et al. [65] GSE73022 | Primary mouse astrocytes Untreated vs. 50 ng/mL TNFα for 24 h (n = 3) | 0.51 0.13 | 0.34 0.037 |

References
- Bossaerts, L.; Cacace, R.; Van Broeckhoven, C. The role of ATP-binding cassette subfamily A in the etiology of Alzheimer’s disease. Mol. Neurodegener. 2022, 17, 31. [Google Scholar] [CrossRef]
- De Roeck, A.; Van Broeckhoven, C.; Sleegers, K. The role of ABCA7 in Alzheimer’s disease: Evidence from genomics, transcriptomics and methylomics. Acta Neuropathol. 2019, 138, 201–220. [Google Scholar] [CrossRef] [PubMed]
- Stage, E.; Risacher, S.L.; Lane, K.A.; Gao, S.; Nho, K.; Saykin, A.J.; Apostolova, L.G.; Alzheimer’s Disease Neuroimaging Initiative. Association of the top 20 Alzheimer’s disease risk genes with [18F]flortaucipir PET. Alzheimer’s Dement. 2022, 14, e12308. [Google Scholar] [CrossRef] [PubMed]
- Apostolova, L.G.; Risacher, S.L.; Duran, T.; Stage, E.C.; Goukasian, N.; West, J.D.; Do, T.M.; Grotts, J.; Wilhalme, H.; Nho, K.; et al. Associations of the top 20 Alzheimer disease risk variants with brain amyloidosis. JAMA Neurol. 2018, 75, 328–341. [Google Scholar] [CrossRef] [PubMed]
- Lyssenko, N.N.; Praticò, D. ABCA7 and the altered lipidostasis hypothesis of Alzheimer’s disease. Alzheimer’s Dement. 2021, 17, 164–174. [Google Scholar] [CrossRef] [PubMed]
- Abe-Dohmae, S.; Ikeda, Y.; Matsuo, M.; Hayashi, M.; Okuhira, K.; Ueda, K.; Yokoyama, S. Human ABCA7 supports apolipoprotein-mediated release of cellular cholesterol and phospholipid to generate high density lipoprotein. J. Biol. Chem. 2004, 279, 604–611. [Google Scholar] [CrossRef]
- Hayashi, M.; Abe-Dohmae, S.; Okazaki, M.; Ueda, K.; Yokoyama, S. Heterogeneity of high density lipoprotein generated by ABCA1 and ABCA7. J. Lipid Res. 2005, 46, 1703–1711. [Google Scholar] [CrossRef] [PubMed]
- Iwamoto, N.; Abe-Dohmae, S.; Sato, R.; Yokoyama, S. ABCA7 expression is regulated by cellular cholesterol through the SREBP2 pathway and associated with phagocytosis. J. Lipid Res. 2006, 47, 1915–1927. [Google Scholar] [CrossRef]
- Tanaka, N.; Abe-Dohmae, S.; Iwamoto, N.; Fitzgerald, M.L.; Yokoyama, S. HMG-CoA reductase inhibitors enhance phagocytosis by upregulating ATP-binding cassette transporter A7. Atherosclerosis 2011, 217, 407–414. [Google Scholar] [CrossRef]
- Picataggi, A.; Rodrigues, A.; Cromley, D.A.; Wang, H.; Wiener, J.P.; Garliyev, V.; Billheimer, J.T.; Grabiner, B.C.; Hurt, J.A.; Chen, A.C.; et al. Specificity of ABCA7-mediated cell lipid efflux. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2022, 1867, 159157. [Google Scholar] [CrossRef]
- Trigueros-Motos, L.; van Capelleveen, J.C.; Torta, F.; Castaño, D.; Zhang, L.H.; Chai, E.C.; Kang, M.; Dimova, L.G.; Schimmel, A.W.M.; Tietjen, I.; et al. ABCA8 regulates cholesterol efflux and high-density lipoprotein cholesterol levels. Arterioscler. Thromb. Vasc. Biol. 2017, 37, 2147–2155. [Google Scholar] [CrossRef]
- Kanoni, S.; Graham, S.E.; Wang, Y.; Surakka, I.; Ramdas, S.; Zhu, X.; Clarke, S.L.; Bhatti, K.F.; Vedantam, S.; Winkler, T.W.; et al. Implicating genes, pleiotropy, and sexual dimorphism at blood lipid loci through multi-ancestry meta-analysis. Genome Biol. 2022, 23, 268. [Google Scholar] [CrossRef] [PubMed]
- Dong, W.; Wong, K.H.Y.; Liu, Y.; Levy-Sakin, M.; Hung, W.C.; Li, M.; Li, B.; Jin, S.C.; Choi, J.; Lopez-Giraldez, F.; et al. Whole-exome sequencing reveals damaging gene variants associated with hypoalphalipoproteinemia. J. Lipid Res. 2022, 63, 100209. [Google Scholar] [CrossRef] [PubMed]
- Richardson, T.G.; Sanderson, E.; Palmer, T.M.; Ala-Korpela, M.; Ference, B.A.; Davey Smith, G.; Holmes, M.V. Evaluating the relationship between circulating lipoprotein lipids and apolipoproteins with risk of coronary heart disease: A multivariable Mendelian randomisation analysis. PLoS Med. 2020, 17, e1003062. [Google Scholar] [CrossRef]
- Graham, S.E.; Clarke, S.L.; Wu, K.H.; Kanoni, S.; Zajac, G.J.M.; Ramdas, S.; Surakka, I.; Ntalla, I.; Vedantam, S.; Winkler, T.W.; et al. The power of genetic diversity in genome-wide association studies of lipids. Nature 2021, 600, 675–679. [Google Scholar] [CrossRef] [PubMed]
- Iaea, D.B.; Spahr, Z.R.; Singh, R.K.; Chan, R.B.; Zhou, B.; Bareja, R.; Elemento, O.; Di Paolo, G.; Zhang, X.; Maxfield, F.R. Stable reduction of STARD4 alters cholesterol regulation and lipid homeostasis. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2020, 1865, 158609. [Google Scholar] [CrossRef]
- Kálmán, S.; Garbett, K.A.; Vereczkei, A.; Shelton, R.C.; Korade, Z.; Mirnics, K. Metabolic stress-induced microRNA and mRNA expression profiles of human fibroblasts. Exp. Cell Res. 2014, 320, 343–353. [Google Scholar] [CrossRef][Green Version]
- Lu, H.; Sun, J.; Liang, W.; Chang, Z.; Rom, O.; Zhao, Y.; Zhao, G.; Xiong, W.; Wang, H.; Zhu, T.; et al. Cyclodextrin Prevents Abdominal Aortic Aneurysm via Activation of Vascular Smooth Muscle Cell Transcription Factor EB. Circulation 2020, 142, 483–498. [Google Scholar] [CrossRef]
- Seo, Y.K.; Jeon, T.I.; Chong, H.K.; Biesinger, J.; Xie, X.; Osborne, T.F. Genome-wide localization of SREBP-2 in hepatic chromatin predicts a role in autophagy. Cell Metab. 2011, 13, 367–375. [Google Scholar] [CrossRef]
- Carson, R.A.; Rudine, A.C.; Tally, S.J.; Franks, A.L.; Frahm, K.A.; Waldman, J.K.; Silswal, N.; Burale, S.; Phan, J.V.; Chandran, U.R.; et al. Statins impact primary embryonic mouse neural stem cell survival, cell death, and fate through distinct mechanisms. PLoS ONE 2018, 13, e0196387. [Google Scholar] [CrossRef]
- Kusnadi, A.; Park, S.H.; Yuan, R.; Pannellini, T.; Giannopoulou, E.; Oliver, D.; Lu, T.; Park-Min, K.H.; Ivashkiv, L.B. The Cytokine TNF promotes transcription factor SREBP activity and binding to inflammatory genes to activate macrophages and limit tissue repair. Immunity 2019, 51, 241–257.e9. [Google Scholar] [CrossRef] [PubMed]
- Jehle, A.W.; Gardai, S.J.; Li, S.; Linsel-Nitschke, P.; Morimoto, K.; Janssen, W.J.; Vandivier, R.W.; Wang, N.; Greenberg, S.; Dale, B.M.; et al. ATP-binding cassette transporter A7 enhances phagocytosis of apoptotic cells and associated ERK signaling in macrophages. J. Cell Biol. 2006, 174, 547–556. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, N.; Abe-Dohmae, S.; Iwamoto, N.; Fitzgerald, M.L.; Yokoyama, S. Helical apolipoproteins of high-density lipoprotein enhance phagocytosis by stabilizing ATP-binding cassette transporter A7. J. Lipid Res. 2010, 51, 2591–2599. [Google Scholar] [CrossRef]
- Abe-Dohmae, S.; Yokoyama, S. ABCA7 links sterol metabolism to the host defense system: Molecular background for potential management measure of Alzheimer’s disease. Gene 2021, 768, 145316. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Mesa, Y.; Jay, T.R.; Checkley, M.A.; Luttge, B.; Dobrowolski, C.; Valadkhan, S.; Landreth, G.E.; Karn, J.; Alvarez-Carbonell, D. Immortalization of primary microglia: A new platform to study HIV regulation in the central nervous system. J. Neurovirol. 2017, 23, 47–66. [Google Scholar] [CrossRef] [PubMed]
- Oram, J.F.; Vaughan, A.M.; Stocker, R. ATP-binding cassette transporter A1 mediates cellular secretion of alpha-tocopherol. J. Biol. Chem. 2001, 276, 39898–39902. [Google Scholar] [CrossRef]
- Robinet, P.; Wang, Z.; Hazen, S.L.; Smith, J.D. A simple and sensitive enzymatic method for cholesterol quantification in macrophages and foam cells. J. Lipid Res. 2010, 51, 3364–3369. [Google Scholar] [CrossRef]
- Cinar, O.; Viechtbauer, W. The poolr package for combining independent and dependent p values. J. Stat. Softw. 2022, 101, 1–42. [Google Scholar] [CrossRef]
- Goldstein, J.L.; Brown, M.S. The LDL receptor. Arterioscler. Thromb. Vasc. Biol. 2009, 29, 431–438. [Google Scholar] [CrossRef]
- Cummings, R.D.; Kornfeld, S.; Schneider, W.J.; Hobgood, K.K.; Tolleshaug, H.; Brown, M.S.; Goldstein, J.L. Biosynthesis of N- and O-linked oligosaccharides of the low density lipoprotein receptor. J. Biol. Chem. 1983, 258, 15261–15273. [Google Scholar] [CrossRef]
- Terrand, J.; Bruban, V.; Zhou, L.; Gong, W.; El Asmar, Z.; May, P.; Zurhove, K.; Haffner, P.; Philippe, C.; Woldt, E.; et al. LRP1 controls intracellular cholesterol storage and fatty acid synthesis through modulation of Wnt signaling. J. Biol. Chem. 2009, 284, 381–388. [Google Scholar] [CrossRef] [PubMed]
- Selvais, C.; D’Auria, L.; Tyteca, D.; Perrot, G.; Lemoine, P.; Troeberg, L.; Dedieu, S.; Noël, A.; Nagase, H.; Henriet, P.; et al. Cell cholesterol modulates metalloproteinase-dependent shedding of low-density lipoprotein receptor-related protein-1 (LRP-1) and clearance function. FASEB J. 2011, 25, 2770–2781. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Yeung, A.W.K.; Atanasov, A.G. A review: Molecular mechanism of regulation of ABCA1 expression. Curr. Protein Pept. Sci. 2022, 23, 170–191. [Google Scholar] [CrossRef] [PubMed]
- Kim, W.S.; Fitzgerald, M.L.; Kang, K.; Okuhira, K.; Bell, S.A.; Manning, J.J.; Koehn, S.L.; Lu, N.; Moore, K.J.; Freeman, M.W. Abca7 null mice retain normal macrophage phosphatidylcholine and cholesterol efflux activity despite alterations in adipose mass and serum cholesterol levels. J. Biol. Chem. 2005, 280, 3989–3995. [Google Scholar] [CrossRef]
- Kuzu, O.F.; Toprak, M.; Noory, M.A.; Robertson, G.P. Effect of lysosomotropic molecules on cellular homeostasis. Pharmacol. Res. 2017, 117, 177–184. [Google Scholar] [CrossRef]
- Matejuk, A.; Ransohoff, R.M. Crosstalk between astrocytes and microglia: An overview. Front. Immunol. 2020, 11, 1416. [Google Scholar] [CrossRef]
- Lee, Y.B.; Nagai, A.; Kim, S.U. Cytokines, chemokines, and cytokine receptors in human microglia. J. Neurosci. Res. 2002, 69, 94–103. [Google Scholar] [CrossRef]
- Lee, S.C.; Liu, W.; Dickson, D.W.; Brosnan, C.F.; Berman, J.W. Cytokine production by human fetal microglia and astrocytes. Differential induction by lipopolysaccharide and IL-1 beta. J. Immunol. 1993, 150, 2659–2667. [Google Scholar] [CrossRef]
- Basu, A.; Krady, J.K.; Enterline, J.R.; Levison, S.W. Transforming growth factor beta1 prevents IL-1beta-induced microglial activation, whereas TNFalpha- and IL-6-stimulated activation are not antagonized. Glia 2002, 40, 109–120. [Google Scholar] [CrossRef]
- Medzhitov, R. The spectrum of inflammatory responses. Science 2021, 374, 1070–1075. [Google Scholar] [CrossRef]
- Meizlish, M.L.; Franklin, R.A.; Zhou, X.; Medzhitov, R. Tissue homeostasis and inflammation. Annu. Rev. Immunol. 2021, 39, 557–581. [Google Scholar] [CrossRef]
- Bernier, L.P.; York, E.M.; MacVicar, B.A. Immunometabolism in the brain: How metabolism shapes microglial function. Trends Neurosci. 2020, 43, 854–869. [Google Scholar] [CrossRef]
- Lee, M.S.; Bensinger, S.J. Reprogramming cholesterol metabolism in macrophages and its role in host defense against cholesterol-dependent cytolysins. Cell. Mol. Immunol. 2022, 19, 327–336. [Google Scholar] [CrossRef]
- Hayakawa, S.; Tamura, A.; Nikiforov, N.; Koike, H.; Kudo, F.; Cheng, Y.; Miyazaki, T.; Kubekina, M.; Kirichenko, T.V.; Orekhov, A.N.; et al. Activated cholesterol metabolism is integral for innate macrophage responses by amplifying Myd88 signaling. JCI Insight 2022, 7, e138539. [Google Scholar] [CrossRef]
- Zhou, Q.D.; Chi, X.; Lee, M.S.; Hsieh, W.Y.; Mkrtchyan, J.J.; Feng, A.C.; He, C.; York, A.G.; Bui, V.L.; Kronenberger, E.B.; et al. Interferon-mediated reprogramming of membrane cholesterol to evade bacterial toxins. Nat. Immunol. 2020, 21, 746–755. [Google Scholar] [CrossRef] [PubMed]
- Guo, C.; Chi, Z.; Jiang, D.; Xu, T.; Yu, W.; Wang, Z.; Chen, S.; Zhang, L.; Liu, Q.; Guo, X.; et al. Cholesterol homeostatic regulator SCAP-SREBP2 integrates NLRP3 inflammasome activation and cholesterol biosynthetic signaling in macrophages. Immunity 2018, 49, 842–856.e7. [Google Scholar] [CrossRef] [PubMed]
- Vitali, C.; Wellington, C.L.; Calabresi, L. HDL and cholesterol handling in the brain. Cardiovasc. Res. 2014, 103, 405–413. [Google Scholar] [CrossRef] [PubMed]
- Delbridge, A.R.D.; Huh, D.; Brickelmaier, M.; Burns, J.C.; Roberts, C.; Challa, R.; Raymond, N.; Cullen, P.; Carlile, T.M.; Ennis, K.A.; et al. Organotypic brain slice culture microglia exhibit molecular similarity to acutely-isolated adult microglia and provide a platform to study neuroinflammation. Front. Cell. Neurosci. 2020, 14, 592005. [Google Scholar] [CrossRef] [PubMed]
- Giambelluca, S.; Ochs, M.; Lopez-Rodriguez, E. Resting time after phorbol 12-myristate 13-acetate in THP-1 derived macrophages provides a non-biased model for the study of NLRP3 inflammasome. Front. Immunol. 2022, 13, 958098. [Google Scholar] [CrossRef]
- Tcw, J.; Qian, L.; Pipalia, N.H.; Chao, M.J.; Liang, S.A.; Shi, Y.; Jain, B.R.; Bertelsen, S.E.; Kapoor, M.; Marcora, E.; et al. Cholesterol and matrisome pathways dysregulated in astrocytes and microglia. Cell 2022, 185, 2213–2233.e25. [Google Scholar] [CrossRef]
- Wiehler, A.; Branzoli, F.; Adanyeguh, I.; Mochel, F.; Pessiglione, M. A neuro-metabolic account of why daylong cognitive work alters the control of economic decisions. Curr. Biol. 2022, 32, 3564–3575.e5. [Google Scholar] [CrossRef] [PubMed]
- Aikawa, T.; Ren, Y.; Yamazaki, Y.; Tachibana, M.; Johnson, M.R.; Anderson, C.T.; Martens, Y.A.; Holm, M.L.; Asmann, Y.W.; Saito, T.; et al. ABCA7 haplodeficiency disturbs microglial immune responses in the mouse brain. Proc. Natl. Acad. Sci. USA 2019, 116, 23790–23796. [Google Scholar] [CrossRef] [PubMed]
- Mahmoudi, E.; Sadaghiyani, S.; Lin, P.; Kamdar, N.; Norcott, A.; Peterson, M.D.; Meade, M.A. Diagnosis of Alzheimer’s disease and related dementia among people with multiple sclerosis: Large cohort study, USA. Mult. Scler. Relat. Disord. 2022, 57, 103351. [Google Scholar] [CrossRef]
- Lamartinière, Y.; Boucau, M.C.; Dehouck, L.; Krohn, M.; Pahnke, J.; Candela, P.; Gosselet, F.; Fenart, L. ABCA7 downregulation modifies cellular cholesterol homeostasis and decreases amyloid-β peptide efflux in an in vitro model of the blood-brain barrier. J. Alzheimer’s Dis. 2018, 64, 1195–1211. [Google Scholar] [CrossRef]
- Satoh, K.; Abe-Dohmae, S.; Yokoyama, S.; St George-Hyslop, P.; Fraser, P.E. ATP-binding cassette transporter A7 (ABCA7) loss of function alters Alzheimer amyloid processing. J. Biol. Chem. 2015, 290, 24152–24165. [Google Scholar] [CrossRef] [PubMed]
- Ikeda, Y.; Abe-Dohmae, S.; Munehira, Y.; Aoki, R.; Kawamoto, S.; Furuya, A.; Shitara, K.; Amachi, T.; Kioka, N.; Matsuo, M.; et al. Posttranscriptional regulation of human ABCA7 and its function for the apoA-I-dependent lipid release. Biochem. Biophys. Res. Commun. 2003, 311, 313–318. [Google Scholar] [CrossRef]
- Chamaria, S.; Johnson, K.W.; Vengrenyuk, Y.; Baber, U.; Shameer, K.; Divaraniya, A.A.; Glicksberg, B.S.; Li, L.; Bhatheja, S.; Moreno, P.; et al. Intracoronary imaging, cholesterol efflux, and transcriptomics after intensive statin treatment in diabetes. Sci. Rep. 2017, 7, 7001. [Google Scholar] [CrossRef]
- Grunwald, S.A.; Popp, O.; Haafke, S.; Jedraszczak, N.; Grieben, U.; Saar, K.; Patone, G.; Kress, W.; Steinhagen-Thiessen, E.; Dittmar, G.; et al. Statin-induced myopathic changes in primary human muscle cells and reversal by a prostaglandin F2 alpha analogue. Sci. Rep. 2020, 10, 2158. [Google Scholar] [CrossRef]
- Willemsen, L.; Chen, H.J.; van Roomen, C.P.A.A.; Griffith, G.R.; Siebeler, R.; Neele, A.E.; Kroon, J.; Hoeksema, M.A.; de Winther, M.P.J. Monocyte and macrophage lipid accumulation results in down-regulated type-I interferon responses. Front. Cardiovasc. Med. 2022, 9, 829877. [Google Scholar] [CrossRef]
- Escartin, C.; Galea, E.; Lakatos, A.; O’Callaghan, J.P.; Petzold, G.C.; Serrano-Pozo, A.; Steinhäuser, C.; Volterra, A.; Carmignoto, G.; Agarwal, A.; et al. Reactive astrocyte nomenclature, definitions, and future directions. Nat. Neurosci. 2021, 24, 312–325. [Google Scholar] [CrossRef]
- Schneider, R.; Leven, P.; Mallesh, S.; Breßer, M.; Schneider, L.; Mazzotta, E.; Fadda, P.; Glowka, T.; Vilz, T.O.; Lingohr, P.; et al. IL-1-dependent enteric gliosis guides intestinal inflammation and dysmotility and modulates macrophage function. Commun. Biol. 2022, 5, 811. [Google Scholar] [CrossRef] [PubMed]
- Lemaître, F.; Farzam-Kia, N.; Carmena Moratalla, A.; Carpentier Solorio, Y.; Clenet, M.L.; Tastet, O.; Cleret-Buhot, A.; Guimond, J.V.; Haddad, E.; Duquette, P.; et al. IL-27 shapes the immune properties of human astrocytes and their impact on encountered human T lymphocytes. J. Neuroinflamm. 2022, 19, 212. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Pan, L.; Pembroke, W.G.; Rexach, J.E.; Godoy, M.I.; Condro, M.C.; Alvarado, A.G.; Harteni, M.; Chen, Y.W.; Stiles, L.; et al. Conservation and divergence of vulnerability and responses to stressors between human and mouse astrocytes. Nat. Commun. 2021, 12, 3958. [Google Scholar] [CrossRef] [PubMed]
- Birck, C.; Ginolhac, A.; Pavlou, M.A.S.; Michelucci, A.; Heuschling, P.; Grandbarbe, L. NF-κB and TNF affect the astrocytic differentiation from neural stem cells. Cells 2021, 10, 840. [Google Scholar] [CrossRef]
- Gabel, S.; Koncina, E.; Dorban, G.; Heurtaux, T.; Birck, C.; Glaab, E.; Michelucci, A.; Heuschling, P.; Grandbarbe, L. Inflammation promotes a conversion of astrocytes into neural progenitor cells via NF-κB activation. Mol. Neurobiol. 2016, 53, 5041–5055. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wiener, J.P.; Desire, S.; Garliyev, V.; Lyssenko III, N.; Praticò, D.; Lyssenko, N.N. Down-Regulation of ABCA7 in Human Microglia, Astrocyte and THP-1 Cell Lines by Cholesterol Depletion, IL-1β and TNFα, or PMA. Cells 2023, 12, 2143. https://doi.org/10.3390/cells12172143
Wiener JP, Desire S, Garliyev V, Lyssenko III N, Praticò D, Lyssenko NN. Down-Regulation of ABCA7 in Human Microglia, Astrocyte and THP-1 Cell Lines by Cholesterol Depletion, IL-1β and TNFα, or PMA. Cells. 2023; 12(17):2143. https://doi.org/10.3390/cells12172143
Chicago/Turabian StyleWiener, Joel P., Sindy Desire, Viktor Garliyev, Nicholas Lyssenko III, Domenico Praticò, and Nicholas N. Lyssenko. 2023. "Down-Regulation of ABCA7 in Human Microglia, Astrocyte and THP-1 Cell Lines by Cholesterol Depletion, IL-1β and TNFα, or PMA" Cells 12, no. 17: 2143. https://doi.org/10.3390/cells12172143
APA StyleWiener, J. P., Desire, S., Garliyev, V., Lyssenko III, N., Praticò, D., & Lyssenko, N. N. (2023). Down-Regulation of ABCA7 in Human Microglia, Astrocyte and THP-1 Cell Lines by Cholesterol Depletion, IL-1β and TNFα, or PMA. Cells, 12(17), 2143. https://doi.org/10.3390/cells12172143