


Effect of Vitamin D3 on Chemerin and Adiponectin Levels in Uterus of Polycystic Ovary Syndrome Rats

 , , and
, , and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. Animals and Experimental Protocol
2.3. Ovarian and Uterus Histology
2.4. ELISA
2.5. Colorimetric Assays
2.6. Real-Time PCR
2.7. Western Blotting
2.8. Immunohistochemistry
2.9. Statistical Analysis
3. Results
3.1. Plasma Levels of Hormonal and Metabolic Parameters, and Ovarian Morphology in PCOS Rats
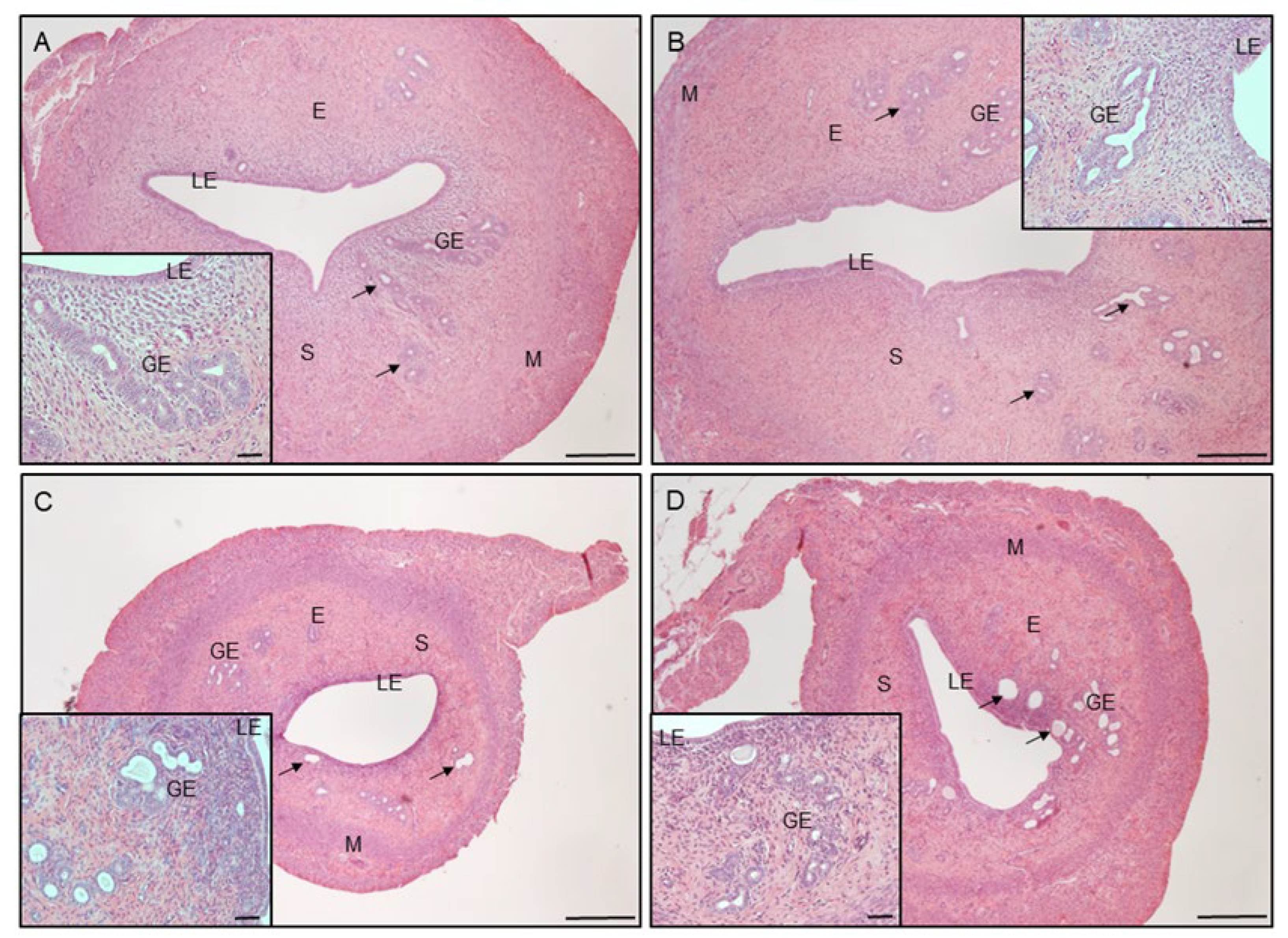
3.2. Uterine Histology
3.3. Effect of VD3 on Plasma Level of RARRES2, ADIPOQ and Steroids in PCOS Rats
3.4. Effect of VD3 on Gene and Protein Expression of RARRES2 and Its Receptors in the Uterus of PCOS Rats
3.5. Effect of VD3 on Cellular Immunolocalization of RARRES2 and CKMLR1 in the Uterus of PCOS Rats
3.6. Effect of VD3 on Gene and Protein Expression of ADIPOQ and Its Receptors in the Uterus of PCOS Rats
3.7. Effect of VD3 on Cellular Immunolocalization of ADIPOQ and AdipoR1 in the Uterus of PCOS Rats
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bruni, V.; Capozzi, A.; Lello, S. The Role of Genetics, Epigenetics and Lifestyle in Polycystic Ovary Syndrome Development: The State of the Art. Reprod. Sci. 2022, 29, 668–679. [Google Scholar] [CrossRef]
- De Leo, V.; Musacchio, M.C.; Cappelli, V.; Massaro, M.G.; Morgante, G.; Petraglia, F. Genetic, hormonal and metabolic aspects of PCOS: An update. Reprod. Biol. Endocrinol. 2016, 14, 38. [Google Scholar] [CrossRef]
- Palomba, S. Is fertility reduced in ovulatory women with polycystic ovary syndrome? An opinion paper. Hum. Reprod. 2021, 36, 2421–2428. [Google Scholar] [CrossRef] [PubMed]
- Palomba, S.; Daolio, J.; La Sala, G.B. Oocyte Competence in Women with Polycystic Ovary Syndrome. Trends Endocrinol. Metab. 2017, 28, 186–198. [Google Scholar] [CrossRef]
- Palomba, S.; de Wilde, M.A.; Falbo, A.; Koster, M.P.; La Sala, G.B.; Fauser, B.C. Pregnancy complications in women with polycystic ovary syndrome. Hum. Reprod. Update 2015, 21, 575–592. [Google Scholar] [CrossRef]
- Palomba, S.; Piltonen, T.T.; Giudice, L.C. Endometrial Function in Women with Polycystic Ovary Syndrome: A Comprehensive Review. Hum. Reprod. Update 2021, 27, 584–618. [Google Scholar] [CrossRef] [PubMed]
- Yan, L.; Wang, A.; Chen, L.; Shang, W.; Li, M.; Zhao, Y. Expression of Apoptosis-Related Genes in the Endometrium of Polycystic Ovary Syndrome Patients during the Window of Implantation. Gene 2012, 506, 350–354. [Google Scholar] [CrossRef]
- Oróstica, L.; Astorga, I.; Plaza-Parrochia, F.; Vera, C.; García, V.; Carvajal, R.; Gabler, F.; Romero, C.; Vega, M. Proinflammatory Environment and Role of TNF-α in Endometrial Function of Obese Women Having Polycystic Ovarian Syndrome. Int. J. Obes. 2016, 40, 1715–1722. [Google Scholar] [CrossRef]
- Ding, D.-C.; Chen, W.; Wang, J.-H.; Lin, S.-Z. Association between Polycystic Ovarian Syndrome and Endometrial, Ovarian, and Breast Cancer: A Population-Based Cohort Study in Taiwan. Medicine 2018, 97, e12608. [Google Scholar] [CrossRef] [PubMed]
- Grzesiak, M.; Waszkiewicz, E.; Wojtas, M.; Kowalik, K.; Franczak, A. Expression of Vitamin D Receptor in the Porcine Uterus and Effect of 1,25(OH)2D3 on Progesterone and Estradiol-17β Secretion by Uterine Tissues in Vitro. Theriogenology 2019, 125, 102–108. [Google Scholar] [CrossRef] [PubMed]
- Ingles, S.A.; Wu, L.; Liu, B.T.; Chen, Y.; Wang, C.-Y.; Templeman, C.; Brueggmann, D. Differential Gene Expression by 1,25(OH) 2D3 in an Endometriosis Stromal Cell Line. J. Steroid Biochem. Mol. Biol. 2017, 173, 223–227. [Google Scholar] [CrossRef]
- Grzesiak, M.; Burzawa, G.; Kurowska, P.; Blaszczyk, K.; Szlaga, A.; Blasiak, A.; Sechman, A.; Rak, A. Altered vitamin D3 metabolism in the ovary and periovarian adipose tissue of rats with letrozole-induced PCOS. Histochem. Cell Biol. 2021, 155, 101–116. [Google Scholar] [CrossRef]
- Dravecká, I.; Figurová, J.; Javorský, M.; Petríková, J.; Vaľková, M.; Lazúrová, I. The Effect of Alfacalcidiol and Metformin on Phenotype Manifestations in Women with Polycystic Ovary Syndrome—A Preliminary Study. Physiol. Res. 2016, 815–822. [Google Scholar] [CrossRef] [PubMed]
- Krul-Poel, Y.H.M.; Koenders, P.P.; Steegers-Theunissen, R.P.; ten Boekel, E.; ter Wee, M.M.; Louwers, Y.; Lips, P.; Laven, J.S.E.; Simsek, S. Vitamin D and Metabolic Disturbances in Polycystic Ovary Syndrome (PCOS): A Cross-Sectional Study. PLoS ONE 2018, 13, e0204748. [Google Scholar] [CrossRef] [PubMed]
- Kuyucu, Y.; Çelik, L.S.; Kendirlinan, Ö.; Tap, Ö.; Mete, U.Ö. Investigation of the uterine structural changes in the experimental model with polycystic ovary syndrome and effects of vitamin D treatment: An ultrastructural and immunohistochemical study. Reprod. Biol. 2018, 18, 53–59. [Google Scholar] [CrossRef]
- Mu, Y.; Cheng, D.; Yin, T.; Yang, J. Vitamin D and Polycystic Ovary Syndrome: A Narrative Review. Reprod. Sci. 2021, 28, 2110–2117. [Google Scholar] [CrossRef]
- Nassar, S.Z.; Badae, N.M. Protective Effect of Vitamin D Supplementation in a Rat Modal of Preeclampsia: A Possible Implication of Chemerin. Hypertens. Pregnancy 2019, 38, 149–156. [Google Scholar] [CrossRef] [PubMed]
- Mohammadi, S.M.; Eghbali, S.A.; Soheilikhah, S.; Jam Ashkezari, S.; Salami, M.; Afkhami-Ardekani, M.; Afkhami-Ardekani, A. The Effects of Vitamin D Supplementation on Adiponectin Level and Insulin Resistance in First-Degree Relatives of Subjects with Type 2 Diabetes: A Randomized Double-Blinded Controlled Trial. Electron Physician 2016, 8, 2849–2854. [Google Scholar] [CrossRef] [PubMed]
- Seyyed Abootorabi, M.; Ayremlou, P.; Behroozi-Lak, T.; Nourisaeidlou, S. The Effect of Vitamin D Supplementation on Insulin Resistance, Visceral Fat and Adiponectin in Vitamin D Deficient Women with Polycystic Ovary Syndrome: A Randomized Placebo-Controlled Trial. Gynecol. Endocrinol. 2018, 34, 489–494. [Google Scholar] [CrossRef] [PubMed]
- Recinella, L.; Orlando, G.; Ferrante, C.; Chiavaroli, A.; Brunetti, L.; Leone, S. Adipokines: New Potential Therapeutic Target for Obesity and Metabolic, Rheumatic, and Cardiovascular Diseases. Front. Physiol. 2020, 11, 578966. [Google Scholar] [CrossRef] [PubMed]
- Mehrabani, S.; Arab, A.; Karimi, E.; Nouri, M.; Mansourian, M. Blood Circulating Levels of Adipokines in Polycystic Ovary Syndrome Patients: A Systematic Review and Meta-analysis. Reprod. Sci. 2021, 28, 3032–3050. [Google Scholar] [CrossRef] [PubMed]
- Smolińska, N.; Maleszka, A.; Dobrzyn, K.; Kiezun, M.; Szeszko, K.; Kaminski, T. Expression of adiponectin and adiponectin receptors 1 and 2 in the porcine uterus, conceptus, and trophoblast during early pregnancy. Theriogenology 2014, 82, 951–965. [Google Scholar] [CrossRef] [PubMed]
- Gudelska, M.; Dobrzyn, K.; Kiezun, M.; Rytelewska, E.; Kisielewska, K.; Kaminska, B.; Kaminski, T.; Smolinska, N. The Expression of Chemerin and Its Receptors (CMKLR1, GPR1, CCRL2) in the Porcine Uterus during the Oestrous Cycle and Early Pregnancy and in Trophoblasts and Conceptuses. Animal 2020, 14, 2116–2128. [Google Scholar] [CrossRef] [PubMed]
- Takemura, Y.; Osuga, Y.; Yamauchi, T.; Kobayashi, M.; Harada, M.; Hirata, T.; Morimoto, C.; Hirota, Y.; Yoshino, O.; Koga, K.; et al. Expression of Adiponectin Receptors and Its Possible Implication in the Human Endometrium. Endocrinology 2006, 147, 3203–3210. [Google Scholar] [CrossRef] [PubMed]
- Schaffer, A.; Ducza, E.; Bódi, N.; Bagyánszki, M.; Szalai, Z.; Mirdamadi, M.; Barna, T.; Szűcs, K.F.; Gáspár, R. The ontogenies of endometrial and myometrial leptin and adiponectin receptors in pregnant rats: Their putative impact on uterine contractility. Life Sci. 2022, 297, 120465. [Google Scholar] [CrossRef]
- Smolińska, N.; Szeszko, K.; Dobrzyn, K.; Kiezun, M.; Rytelewska, E.; Kisielewska, K.; Gudelska, M.; Bors, K.; Wyrebek, J.; Kopij, G.; et al. Transcriptomic Analysis of Porcine Endometrium during Implantation after In Vitro Stimulation by Adiponectin. Int. J. Mol. Sci. 2019, 20, 1335. [Google Scholar] [CrossRef]
- Orzechowska, K.; Kopij, G.; Paukszto, L.; Dobrzyn, K.; Kiezun, M.; Jastrzebski, J.; Kaminski, T.; Smolinska, N. Chemerin Effect on Transcriptome of the Porcine Endometrium during Implantation Determined by RNA-Sequencing. Biol. Reprod. 2022, 107, 557–573. [Google Scholar] [CrossRef]
- Franks, S. Do animal models of polycystic ovary syndrome help to understand its pathogenesis and management? Yes, but their limitations should be recognized. Endocrinology 2009, 150, 3983–3985. [Google Scholar] [CrossRef]
- Osuka, S.; Nakanishi, N.; Murase, T.; Nakamura, T.; Goto, M.; Iwase, A.; Kikkawa, F. Animal models of polycystic ovary syndrome: A review of hormone-induced rodent models focused on hypothalamus-pituitary-ovary axis and neuropeptides. Reprod. Med. Biol. 2018, 18, 151–160. [Google Scholar] [CrossRef] [PubMed]
- Brock, B.J.; Waterman, M.R. Biochemical differences between rat and human cytochrome P450c17 support the different steroidogenic needs of these two species. Biochemistry 1999, 38, 1598–1606. [Google Scholar] [CrossRef] [PubMed]
- Stener-Victorin, E.; Padmanabhan, V.; Walters, K.A.; Campbell, R.E.; Benrick, A.; Giacobini, P.; Dumesic, D.A.; Abbott, D.H. Animal Models to Understand the Etiology and Pathophysiology of Polycystic Ovary Syndrome. Endocr. Rev. 2020, 41, bnaa010. [Google Scholar] [CrossRef]
- Malik, M.; Roh, M.; England, S.K. Uterine contractions in rodent models and humans. Acta Physiol. 2021, 231, e13607. [Google Scholar] [CrossRef] [PubMed]
- Kafali, H.; Iriadam, M.; Ozardalı, I.; Demir, N. Letrozole-Induced Polycystic Ovaries in the Rat: A New Model for Cystic Ovarian Disease. Arch. Med. Res. 2004, 35, 103–108. [Google Scholar] [CrossRef] [PubMed]
- Lajtai, K.; Nagy, C.T.; Tarszabó, R.; Benkő, R.; Hadjadj, L.; Sziva, R.E.; Gerszi, D.; Bányai, B.; Ferdinandy, P.; Nádasy, G.L.; et al. Effects of Vitamin D deficiency on proliferation and autophagy of ovarian and liver tissues in a rat model of polycystic ovary syndrome. Biomolecules 2019, 9, 471. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Rak, A.; Drwal, E.; Rame, C.; Knapczyk-Stwora, K.; Słomczyńska, M.; Dupont, J.; Gregoraszczuk, E.L. Expression of apelin and apelin receptor (APJ) in porcine ovarian follicles and in vitro effect of apelin on steroidogenesis and proliferation through APJ activation and different signaling pathways. Theriogenology 2017, 96, 126–135. [Google Scholar] [CrossRef] [PubMed]
- Asadi, M.; Matin, N.; Frootan, M.; Mohamadpour, J.; Qorbani, M.; Tanha, F.D. Vitamin D Improves Endometrial Thickness in PCOS Women Who Need Intrauterine Insemination: A Randomized Double-Blind Placebo-Controlled Trial. Arch. Gynecol. Obs. 2014, 289, 865–870. [Google Scholar] [CrossRef] [PubMed]
- Thys-Jacobs, S.; Donovan, D.; Papadopoulos, A.; Sarrel, P.; Bilezikian, J.P. Vitamin D and calcium dysregulation in the polycystic ovarian syndrome. Steroids. 1999, 64, 430–435. [Google Scholar] [CrossRef]
- Wehr, E.; Pieber, T.R.; Obermayer-Pietsch, B. Effect of vitamin D3 treatment on glucose metabolism and menstrual frequency in polycystic ovary syndrome women: A pilot study. J. Endocrinol. Invest. 2011, 34, 757–763. [Google Scholar] [CrossRef] [PubMed]
- Behmanesh, N.; Abedelahi, A.; Charoudeh, H.N.; Alihemmati, A. Effects of Vitamin D Supplementation on Follicular Development, Gonadotropins and Sex Hormone Concentrations, and Insulin Resistance in Induced Polycystic Ovary Syndrome. Turk. J. Obstet. Gynecol. 2019, 16, 143–150. [Google Scholar] [CrossRef] [PubMed]
- Kalamon, N.; Błaszczyk, K.; Szlaga, A.; Billert, M.; Skrzypski, M.; Pawlicki, P.; Górowska-Wójtowicz, E.; Kotula-Balak, M.; Błasiak, A.; Rak, A. Levels of the Neuropeptide Phoenixin-14 and Its Receptor GRP173 in the Hypothalamus, Ovary and Periovarian Adipose Tissue in Rat Model of Polycystic Ovary Syndrome. Biochem. Biophys. Res. Commun. 2020, 528, 628–635. [Google Scholar] [CrossRef] [PubMed]
- Shang, K.; Jia, X.; Qiao, J.; Kang, J.; Guan, Y. Endometrial Abnormality in Women with Polycystic Ovary Syndrome. Reprod. Sci. 2012, 19, 674–683. [Google Scholar] [CrossRef] [PubMed]
- Lin, K.; Sun, X.; Wang, X.; Wang, H.; Chen, X. Circulating Adipokine Levels in Nonobese Women With Polycystic Ovary Syndrome and in Nonobese Control Women: A Systematic Review and Meta-Analysis. Front Endocrinol. 2021, 11, 537809. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Zhu, Q.; Wang, W.; Qi, J.; He, Y.; Wang, Y.; Lu, Y.; Wu, H.; Ding, Y.; Sun, Y. Elevated chemerin induces insulin resistance in human granulosa-lutein cells from polycystic ovary syndrome patients. FASEB J. 2019, 33, 11303–11313. [Google Scholar] [CrossRef]
- Yang, S.; Wang, Q.; Huang, W.; Song, Y.; Feng, G.; Zhou, L.; Tan, J. Are Serum Chemerin Levels Different between Obese and Non-Obese Polycystic Ovary Syndrome Women? Gynecol. Endocrinol. 2016, 32, 38–41. [Google Scholar] [CrossRef] [PubMed]
- Bobjer, J.; Katrinaki, M.; Dermitzaki, E.; Margioris, A.N.; Giwercman, A.; Tsatsanis, C. Serum Chemerin Levels Are Negatively Associated with Male Fertility and Reproductive Hormones. Hum. Reprod. 2018, 33, 2168–2174. [Google Scholar] [CrossRef] [PubMed]
- Abeer, A.; Said, M.D.; Suzan, M.M.; Moursi, M.D. Vitamin D Supplementation Reduces Serum Chemerin Level in Gestational Diabetes Mellitus Rat Model. Med. J. Cairo Univ. 2019, 87, 3069–3080. [Google Scholar] [CrossRef][Green Version]
- Schwanhäusser, B.; Busse, D.; Li, N.; Dittmar, G.; Schuchhardt, J.; Wolf, J.; Chen, W.; Selbach, M. Global Quantification of Mammalian Gene Expression Control. Nature 2011, 473, 337–342. [Google Scholar] [CrossRef] [PubMed]
- Vogel, C.; Marcotte, E.M. Insights into the Regulation of Protein Abundance from Proteomic and Transcriptomic Analyses. Nat. Rev. Genet. 2012, 13, 227–232. [Google Scholar] [CrossRef]
- Roman, A.A.; Sinal, C. Vitamin D regulation of chemerin in human bone marrow adipogenesis. FASEB J. 2009, 23. [Google Scholar] [CrossRef]
- Nagpal, S.; Patel, S.; Jacobe, H.; DiSepio, D.; Ghosn, C.; Malhotra, M.; Teng, M.; Duvic, M.; Chandraratna, R.A.S. Tazarotene-Induced Gene 2 (TIG2), a Novel Retinoid-Responsive Gene in Skin. J. Investig. Dermatol. 1997, 109, 91–95. [Google Scholar] [CrossRef] [PubMed]
- De Henau, O.; Degroot, G.N.; Imbault, V.; Robert, V.; De Poorter, C.; Mcheik, S.; Galés, C.; Parmentier, M.; Springael, J.Y. Signaling Properties of Chemerin Receptors CMKLR1, GPR1 and CCRL2. PLoS ONE 2016, 11, e0164179. [Google Scholar] [CrossRef]
- Monnier, J.; Lewén, S.; O.’Hara, E.; Huang, K.; Tu, H.; Butcher, E.C.; Zabel, B.A. Expression, Regulation, and Function of Atypical Chemerin Receptor CCRL2 on Endothelial Cells. J. Immunol. 2012, 189, 956–967. [Google Scholar] [CrossRef]
- Gudelska, M.; Dobrzyn, K.; Kiezun, M.; Kisielewska, K.; Rytelewska, E.; Kaminski, T.; Smolinska, N. Chemerin Affects P4 and E2 Synthesis in the Porcine Endometrium during Early Pregnancy. Int. J. Mol. Sci. 2022, 23, 945. [Google Scholar] [CrossRef] [PubMed]
- Luo, X.; Gong, Y.; Cai, L.; Zhang, L.; Dong, X. Chemerin Regulates Autophagy to Participate in Polycystic Ovary Syndrome. J. Int. Med. Res. 2021, 49, 030006052110583. [Google Scholar] [CrossRef] [PubMed]
- Estienne, A.; Mellouk, N.; Bongrani, A.; Plotton, I.; Langer, I.; Ramé, C.; Petit, C.; Guérif, F.; Froment, P.; Dupont, J. Involvement of Chemerin and CMKLR1 in the Progesterone Decrease by PCOS Granulosa Cells. Reproduction 2021, 162, 427–436. [Google Scholar] [CrossRef]
- Yamamoto, Y.; Hirose, H.; Saito, I.; Tomita, M.; Taniyama, M.; Matsubara, K.; Okazaki, Y.; Ishii, T.; Nishikai, K.; Saruta, T. Correlation of the Adipocyte-Derived Protein Adiponectin with Insulin Resistance Index and Serum High-Density Lipoprotein-Cholesterol, Independent of Body Mass Index, in the Japanese Population. Clin. Sci. 2002, 103, 137. [Google Scholar] [CrossRef]
- Nikooyeh, B.; Neyestani, T.R. Can Vitamin D Be Considered an Adiponectin Secretagogue? A Systematic Review and Meta-Analysis. J. Steroid Biochem. Mol. Biol. 2021, 212, 105925. [Google Scholar] [CrossRef]
- Walker, G.E.; Ricotti, R.; Roccio, M.; Moia, S.; Bellone, S.; Prodam, F.; Bona, G. Pediatric Obesity and Vitamin D Deficiency: A Proteomic Approach Identifies Multimeric Adiponectin as a Key Link between These Conditions. PLoS ONE 2014, 9, e83685. [Google Scholar] [CrossRef] [PubMed]
- Lorente-Cebrián, S.; Eriksson, A.; Dunlop, T.; Mejhert, N.; Dahlman, I.; Åström, G.; Sjölin, E.; Wåhlén, K.; Carlberg, C.; Laurencikiene, J.; et al. Differential effects of 1α,25-dihydroxycholecalciferol on MCP-1 and adiponectin production in human white adipocytes. Eur. J. Nutr. 2012, 51, 335–342. [Google Scholar] [CrossRef]
- Vaidya, A.; Williams, J.S. The relationship between vitamin D and the renin-angiotensin system in the pathophysiology of hypertension, kidney disease, and diabetes. Metabolism 2012, 61, 450–458. [Google Scholar] [CrossRef] [PubMed]
- Liu, E.; Meigs, J.B.; Pittas, A.G.; McKeown, N.M.; Economos, C.D.; Booth, S.L.; Jacques, P.F. Plasma 25-hydroxyvitamin d is associated with markers of the insulin resistant phenotype in nondiabetic adults. J. Nutr. 2009, 139, 329–334. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.H.; Wang, Y.C.; Wu, S.C.; Mersmann, H.J.; Cheng, W.T.K.; Ding, S.T. Insulin regulates the expression of adiponectin and adiponectin receptors in porcine adipocytes. Domest. Anim. Endocrinol. 2008, 34, 352–359. [Google Scholar] [CrossRef] [PubMed]
- Comim, F.V.; Hardy, K.; Franks, S. Adiponectin and Its Receptors in the Ovary: Further Evidence for a Link between Obesity and Hyperandrogenism in Polycystic Ovary Syndrome. PLoS ONE 2013, 8, e80416. [Google Scholar] [CrossRef] [PubMed]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Symbol | Gene Name | Catalog Number | Reference Sequence |
|---|---|---|---|
| Rarres2 | retinoic acid 109 receptor responder protein 2 | Rn01451853_m1 | NM_001013427.1 |
| Ccrl2 | chemokine (C-C motif) receptor-like 2 | Rn01746782_g1 | NM_001108191.1 |
| Cmklr1 | chemokine-like receptor 1 | Rn00573616_s1 | NM_022218.2 |
| Gpr1 | G protein-coupled receptor 1 | Rn00564179_s1 | NM_012961.1 |
| Adipoq | adiponectin | Rn00595250_m1 | NM_144744 |
| Adipor1 | adiponectin receptor-1 | Rn01483784_m1 | NM_207587.1 |
| Adipor2 | adiponectin receptor-2 | Rn01463173_m1 | NM_001037979.1 |
| Gapdh | Glyceraldehyde-3-phosphate dehydrogenase | Rn01775763_g1 | NM_017008.4 |
| Hormone and Metabolic Parameters | Control | PCOS Rats |
|---|---|---|
| Testosterone (ng/mL) | 0.268 a | 2.471 b |
| Estradiol (pg/mL) | 28.918 b | 22.575 a |
| Glucose (mg/dL) | 186.685 a | 281.391 b |
| Triglyceride (mg/dL) | 59.490 a | 98.668 b |
| HDL | 26.303 a | 28.067 b |
| LDL | 3.950 a | 17.735 b |
| Cholesterol (mg/dL) | 42.615 a | 66.538 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pich, K.; Rajewska, J.; Kamińska, K.; Tchurzyk, M.; Szlaga, A.; Sambak, P.; Błasiak, A.; Grzesiak, M.; Rak, A. Effect of Vitamin D3 on Chemerin and Adiponectin Levels in Uterus of Polycystic Ovary Syndrome Rats. Cells 2023, 12, 2026. https://doi.org/10.3390/cells12162026
Pich K, Rajewska J, Kamińska K, Tchurzyk M, Szlaga A, Sambak P, Błasiak A, Grzesiak M, Rak A. Effect of Vitamin D3 on Chemerin and Adiponectin Levels in Uterus of Polycystic Ovary Syndrome Rats. Cells. 2023; 12(16):2026. https://doi.org/10.3390/cells12162026
Chicago/Turabian StylePich, Karolina, Jesika Rajewska, Kinga Kamińska, Marcelina Tchurzyk, Agata Szlaga, Patryk Sambak, Anna Błasiak, Małgorzata Grzesiak, and Agnieszka Rak. 2023. "Effect of Vitamin D3 on Chemerin and Adiponectin Levels in Uterus of Polycystic Ovary Syndrome Rats" Cells 12, no. 16: 2026. https://doi.org/10.3390/cells12162026
APA StylePich, K., Rajewska, J., Kamińska, K., Tchurzyk, M., Szlaga, A., Sambak, P., Błasiak, A., Grzesiak, M., & Rak, A. (2023). Effect of Vitamin D3 on Chemerin and Adiponectin Levels in Uterus of Polycystic Ovary Syndrome Rats. Cells, 12(16), 2026. https://doi.org/10.3390/cells12162026