
Regulators of the Asexual Life Cycle of Aspergillus nidulans




Abstract
1. Introduction
2. Research on A. nidulans Asexual Spores
3. Conidiogenesis
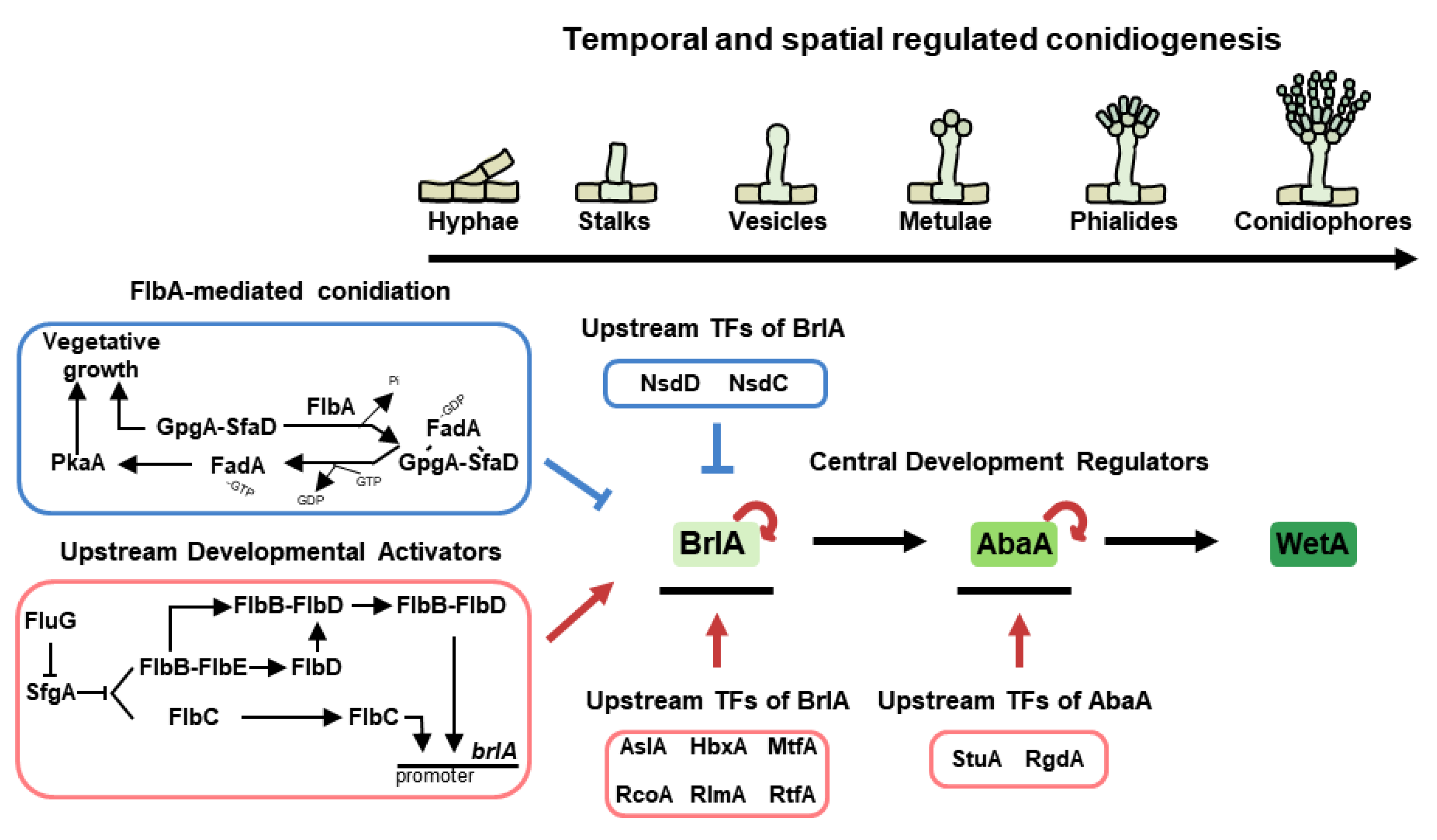
3.1. Temporal and Spatial Central Regulators of Conidiation

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Conidiogenesis | Description | Reference(s) |
|---|---|---|---|
| AclA/B | Activation | ATP-citrate lyase | [43] |
| AcoA~B | Activation | Aconidial genes, encoding putative aconitate hydratase | [44,45] |
| AflR | Activation | Sterigmatocystin/Aflatoxin Zn(II)2Cys6 transcriptional factor | [46] |
| AspE | Activation | Aspergillus septin E (septin protein) | [47] |
| BasA | Activation | Sphingolipid C4-hydroxylase, homolog of S.cerevisiae Sur2 | [48] |
| Bud4 | Activation | Bud site selection protein | [31] |
| CalB~H | Activation | Calcoflour hypersensitivity | [49] |
| CandA-C | Activation | Cullin-associated-nedd8-dissociated protein | [50] |
| ChsA | Activation | Chitin synthase encoding the fungal cell-wall integrity signaling (CWIS) pathway | [51] |
| FhbA/B | Activation | Flavohaemoglobins | [52,53] |
| GfsA | Activation | Galactofuranosyltransferase | [54] |
| GmcA | Activation | Glucose-methanol-choline oxidoreductase | [55] |
| KfsA | Activation | Kinase for septation | [56] |
| OdeA | Activation | Oleate Δ12 desaturases | [57] |
| PchA | Activation | Homolog of S. cerevisiae cyclin T | [58] |
| PclB | Activation | Homolog of S. cerevisiae pcl cyclins | [58] |
| PhnA | Repression | Phosducin-like protein (PhLP) | [59] |
| PhoA | Activation | PSTAIRE-containing kinase | [60] |
| PmtA/B | Activation | Protein O-mannosyltransferase involved in protein glycosylation | [61] |
| PmtC | Repression | Protein O-mannosyltransferase involved in protein glycosylation | [61,62] |
| PpoA/B | Repression | Psi-producing oxygenase involved in oxylipin biosynthesis | [63] |
| PpoC | Activation | Psi-producing oxygenase involved in oxylipin biosynthesis | [64] |
| PufA | Activation | Pumilio/fem-3 binding factor | [65] |
| SnaA~E | Activation | Suppressor of nudA1 | [66] |
| StcE/J/U | Repression | Sterigmatocystin biosynthetic gene cluster | [46] |
| SdeA/B | Activation | Δ9-stearic acid desaturases | [67] |
| SumO | Activation | Small ubiquitin-like modifier involved in SUMOylation | [68,69] |
| UgeA | Activation | UDP-glucose-4-epimerase | [70] |
| VapA | Activation | FYVE-like zinc finger protein, one of the VipC-associated protein | [71] |
| VapB | Activation | H3-K9 specific histone methyltransferase, one of the VipC-associated proteins | [71] |
| VipC | Repression | H3-K9 specific histone methyltransferase, one of the Velvet interacting proteins | [71] |
| WscA/B | Activation | Homolog of S. cerevisiae Wsc1 involved in the fungal cell-wall integrity signaling pathway | [72] |
3.2. Upstream Developmental Activators (UDAs)
3.2.1. Flb Protein-Mediated Asexual Development
3.2.2. Inhibition of FlbA-Mediated Stimulation of Vegetative Growth
3.3. Upstream Regulators of BrlA
3.4. Other Key Regulators of Asexual Development
4. Conidial Maturation and Dormancy
4.1. The Velvet Family
4.2. Transcription Factors Involved in Conidial Maturation and Dormancy
4.3. Genetic Regulators Related to Stress Tolerance in Conidia
4.3.1. Mitogen-Activated Protein Kinase (MAPK) Cascades
4.3.2. Other Transcription Factors Related to Stress Tolerance in Conidia
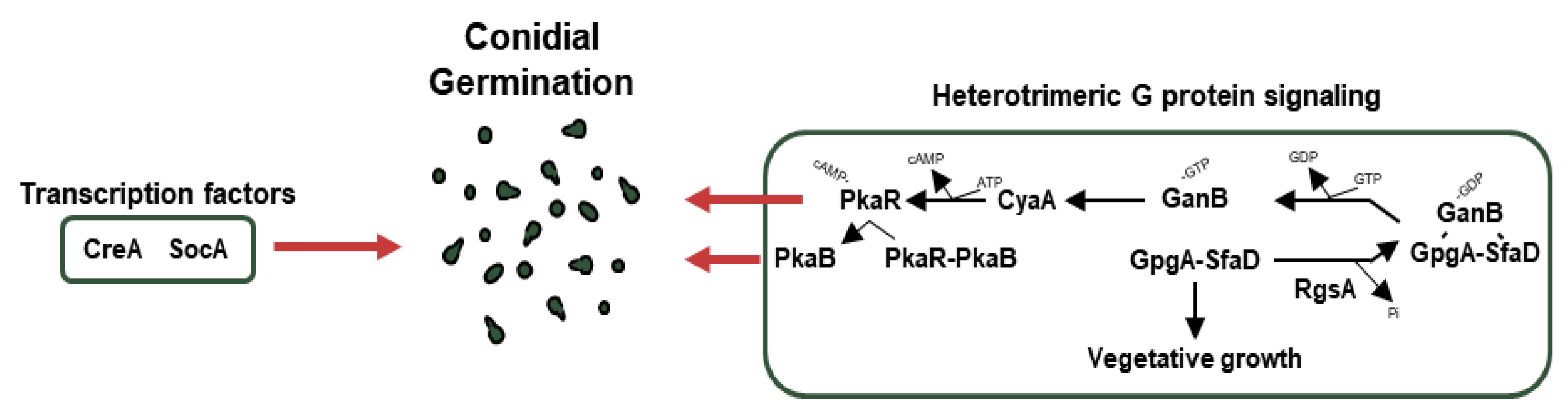
5. Conidial Germination
5.1. Transcription Factors Involved in Germination
5.2. Other Regulators Related to Germination
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Egbuta, M.A.; Mwanza, M.; Babalola, O.O. Health Risks Associated with Exposure to Filamentous Fungi. Int. J. Environ. Res. Public Health 2017, 14, 719. [Google Scholar] [CrossRef] [PubMed]
- Doehlemann, G.; Okmen, B.; Zhu, W.; Sharon, A. Plant Pathogenic Fungi. Microbiol. Spectr. 2017, 5, FUNK-0023-2016. [Google Scholar] [CrossRef] [PubMed]
- Fisher, M.C.; Henk, D.A.; Briggs, C.J.; Brownstein, J.S.; Madoff, L.C.; McCraw, S.L.; Gurr, S.J. Emerging fungal threats to animal, plant and ecosystem health. Nature 2012, 484, 186–194. [Google Scholar] [CrossRef] [PubMed]
- Alshannaq, A.; Yu, J.H. Occurrence, Toxicity, and Analysis of Major Mycotoxins in Food. Int. J. Environ. Res. Public Health 2017, 14, 632. [Google Scholar] [CrossRef] [PubMed]
- Sakekar, A.A.; Gaikwad, S.R.; Punekar, N.S. Protein expression and secretion by filamentous fungi. J. Biosci. 2021, 46, 5. [Google Scholar] [CrossRef]
- Wang, Q.; Zhong, C.; Xiao, H. Genetic Engineering of Filamentous Fungi for Efficient Protein Expression and Secretion. Front. Bioeng. Biotechnol. 2020, 8, 293. [Google Scholar] [CrossRef] [PubMed]
- Hedayati, M.T.; Pasqualotto, A.C.; Warn, P.A.; Bowyer, P.; Denning, D.W. Aspergillus flavus: Human pathogen, allergen and mycotoxin producer. Microbiology 2007, 153, 1677–1692. [Google Scholar] [CrossRef]
- Amaike, S.; Keller, N.P. Aspergillus flavus. Annu. Rev. Phytopathol. 2011, 49, 107–133. [Google Scholar] [CrossRef]
- Latge, J.P. Aspergillus fumigatus and aspergillosis. Clin. Microbiol. Rev. 1999, 12, 310–350. [Google Scholar] [CrossRef]
- Latge, J.P.; Chamilos, G. Aspergillus fumigatus and Aspergillosis in 2019. Clin. Microbiol. Rev. 2019, 33, e00140-18. [Google Scholar] [CrossRef]
- Cairns, T.C.; Nai, C.; Meyer, V. How a fungus shapes biotechnology: 100 years of Aspergillus niger research. Fungal Biol. Biotechnol. 2018, 5, 13. [Google Scholar] [CrossRef] [PubMed]
- Jin, F.J.; Hu, S.; Wang, B.T.; Jin, L. Advances in Genetic Engineering Technology and Its Application in the Industrial Fungus Aspergillus oryzae. Front. Microbiol. 2021, 12, 644404. [Google Scholar] [CrossRef] [PubMed]
- Bennett, J.W. Aspergillus: A primer for the novice. Med. Mycol. 2009, 47 (Suppl. 1), S5–S12. [Google Scholar] [CrossRef] [PubMed]
- Ni, M.; Yu, J.H. A novel regulator couples sporogenesis and trehalose biogenesis in Aspergillus nidulans. PLoS ONE 2007, 2, e970. [Google Scholar] [CrossRef]
- Etxebeste, O.; Garzia, A.; Espeso, E.A.; Ugalde, U. Aspergillus nidulans asexual development: Making the most of cellular modules. Trends Microbiol. 2010, 18, 569–576. [Google Scholar] [CrossRef]
- Calvo, A.M.; Wilson, R.A.; Bok, J.W.; Keller, N.P. Relationship between secondary metabolism and fungal development. Microbiol. Mol. Biol. Rev. 2002, 66, 447–459. [Google Scholar] [CrossRef]
- Adams, T.H.; Wieser, J.K.; Yu, J.H. Asexual sporulation in Aspergillus nidulans. Microbiol. Mol. Biol. Rev. 1998, 62, 35–54. [Google Scholar] [CrossRef]
- Park, H.S.; Yu, J.H. Genetic control of asexual sporulation in filamentous fungi. Curr. Opin. Microbiol. 2012, 15, 669–677. [Google Scholar] [CrossRef]
- Krijgsheld, P.; Bleichrodt, R.; van Veluw, G.J.; Wang, F.; Muller, W.H.; Dijksterhuis, J.; Wosten, H.A. Development in Aspergillus. Stud. Mycol. 2013, 74, 1–29. [Google Scholar] [CrossRef]
- Wang, F.; Sethiya, P.; Hu, X.H.; Guo, S.H.; Chen, Y.Y.; Li, A.; Tan, K.L.; Wong, K.H. Transcription in fungal conidia before dormancy produces phenotypically variable conidia that maximize survival in different environments. Nat. Microbiol. 2021, 6, 1066–1081. [Google Scholar] [CrossRef]
- Baltussen, T.J.H.; Zoll, J.; Verweij, P.E.; Melchers, W.J.G. Molecular Mechanisms of Conidial Germination in Aspergillus spp. Microbiol. Mol. Biol. Rev. 2020, 84, e00049-19. [Google Scholar] [CrossRef] [PubMed]
- Schultzhaus, Z.; Cunningham, G.A.; Mourino-Perez, R.R.; Shaw, B.D. The phospholipid flippase DnfD localizes to late Golgi and is involved in asexual differentiation in Aspergillus nidulans. Mycologia 2019, 111, 13–25. [Google Scholar] [CrossRef] [PubMed]
- Zhong, G.W.; Jiang, P.; Qiao, W.R.; Zhang, Y.W.; Wei, W.F.; Lu, L. Protein phosphatase 2A (PP2A) regulatory subunits ParA and PabA orchestrate septation and conidiation and are essential for PP2A activity in Aspergillus nidulans. Eukaryot. Cell 2014, 13, 1494–1506. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Ye, X.S.; Lee, S.L.; Wolkow, T.D.; McGuire, S.L.; Hamer, J.E.; Wood, G.C.; Osmani, S.A. Interaction between developmental and cell cycle regulators is required for morphogenesis in Aspergillus nidulans. EMBO J. 1999, 18, 6994–7001. [Google Scholar] [CrossRef] [PubMed]
- Kadry, A.A.; El-Ganiny, A.M.; Mosbah, R.A.; Kaminskyj, S.G.W. Deletion of Aspergillus nidulans GDP-mannose transporters affects hyphal morphometry, cell wall architecture, spore surface character, cell adhesion, and biofilm formation. Med. Mycol. 2018, 56, 621–630. [Google Scholar] [CrossRef]
- Karos, M.; Fischer, R. Molecular characterization of HymA, an evolutionarily highly conserved and highly expressed protein of Aspergillus nidulans. Mol. Gen. Genet. 1999, 260, 510–521. [Google Scholar] [CrossRef]
- Leeder, A.C.; Turner, G. Characterisation of Aspergillus nidulans polarisome component BemA. Fungal Genet. Biol. 2008, 45, 897–911. [Google Scholar] [CrossRef]
- Zhang, S.; Zheng, H.; Long, N.; Carbo, N.; Chen, P.; Aguilar, P.S.; Lu, L. FigA, a putative homolog of low-affinity calcium system member Fig1 in Saccharomyces cerevisiae, is involved in growth and asexual and sexual development in Aspergillus nidulans. Eukaryot. Cell 2014, 13, 295–303. [Google Scholar] [CrossRef]
- Canovas, D.; Marcos, A.T.; Gacek, A.; Ramos, M.S.; Gutierrez, G.; Reyes-Dominguez, Y.; Strauss, J. The histone acetyltransferase GcnE (GCN5) plays a central role in the regulation of Aspergillus asexual development. Genetics 2014, 197, 1175–1189. [Google Scholar] [CrossRef]
- El-Ganiny, A.M.; Sanders, D.A.; Kaminskyj, S.G. Aspergillus nidulans UDP-galactopyranose mutase, encoded by ugmA plays key roles in colony growth, hyphal morphogensis, and conidiation. Fungal Genet. Biol. 2008, 45, 1533–1542. [Google Scholar] [CrossRef]
- Si, H.; Rittenour, W.R.; Xu, K.; Nicksarlian, M.; Calvo, A.M.; Harris, S.D. Morphogenetic and developmental functions of the Aspergillus nidulans homologues of the yeast bud site selection proteins Bud4 and Axl2. Mol. Microbiol. 2012, 85, 252–270. [Google Scholar] [CrossRef] [PubMed]
- Schier, N.; Liese, R.; Fischer, R. A Pcl-like cyclin of Aspergillus nidulans is transcriptionally activated by developmental regulators and is involved in sporulation. Mol. Cell. Biol. 2001, 21, 4075–4088. [Google Scholar] [CrossRef]
- Liu, B.; Morris, N.R. A spindle pole body-associated protein, SNAD, affects septation and conidiation in Aspergillus nidulans. Mol. Gen. Genet. 2000, 263, 375–387. [Google Scholar] [CrossRef] [PubMed]
- Gems, D.H.; Clutterbuck, A.J. Enhancers of conidiation mutants in Aspergillus nidulans. Genetics 1994, 137, 79–85. [Google Scholar] [CrossRef] [PubMed]
- Appleyard, M.V.C.L.; McPheat, W.L.; Stark, M.J.R. A novel ‘two-component’ protein containing histidine kinase and response regulator domains required for sporulation in Aspergillus nidulans. Curr. Genet. 2000, 37, 364–372. [Google Scholar] [CrossRef]
- Clutterbuck, A.J. A mutational analysis of conidial development in Aspergillus nidulans. Genetics 1969, 63, 317–327. [Google Scholar] [CrossRef] [PubMed]
- Sewall, T.C. Cellular effects of misscheduled brlA, abaA, and wetA expression in Aspergillus nidulans. Can. J. Microbiol. 1994, 40, 1035–1042. [Google Scholar] [CrossRef]
- Adams, T.H.; Boylan, M.T.; Timberlake, W.E. brlA is necessary and sufficient to direct conidiophore development in Aspergillus nidulans. Cell 1988, 54, 353–362. [Google Scholar] [CrossRef]
- Chang, Y.C.; Timberlake, W.E. Identification of Aspergillus brlA response elements (BREs) by genetic selection in yeast. Genetics 1993, 133, 29–38. [Google Scholar] [CrossRef] [PubMed]
- Boylan, M.T.; Mirabito, P.M.; Willett, C.E.; Zimmerman, C.R.; Timberlake, W.E. Isolation and physical characterization of three essential conidiation genes from Aspergillus nidulans. Mol. Cell. Biol. 1987, 7, 3113–3118. [Google Scholar] [CrossRef]
- Sewall, T.C.; Mims, C.W.; Timberlake, W.E. abaA controls phialide differentiation in Aspergillus nidulans. Plant Cell 1990, 2, 731–739. [Google Scholar] [CrossRef] [PubMed]
- Andrianopoulos, A.; Timberlake, W.E. The Aspergillus nidulans abaA gene encodes a transcriptional activator that acts as a genetic switch to control development. Mol. Cell. Biol. 1994, 14, 2503–2515. [Google Scholar] [CrossRef] [PubMed]
- Hynes, M.J.; Murray, S.L. ATP-citrate lyase is required for production of cytosolic acetyl coenzyme A and development in Aspergillus nidulans. Eukaryot. Cell 2010, 9, 1039–1048. [Google Scholar] [CrossRef] [PubMed]
- Butnick, N.Z.; Yager, L.N.; Hermann, T.E.; Kurtz, M.B.; Champe, S.P. Mutants of Aspergillus nidulans blocked at an early stage of sporulation secrete an unusual metabolite. J. Bacteriol. 1984, 160, 533–540. [Google Scholar] [CrossRef]
- Butnick, N.Z.; Yager, L.N.; Kurtz, M.B.; Champe, S.P. Genetic analysis of mutants of Aspergillus nidulans blocked at an early stage of sporulation. J. Bacteriol. 1984, 160, 541–545. [Google Scholar] [CrossRef]
- Wilkinson, H.H.; Ramaswamy, A.; Sim, S.C.; Keller, N.P. Increased conidiation associated with progression along the sterigmatocystin biosynthetic pathway. Mycologia 2004, 96, 1190–1198. [Google Scholar] [CrossRef]
- Hernandez-Rodriguez, Y.; Masuo, S.; Johnson, D.; Orlando, R.; Smith, A.; Couto-Rodriguez, M.; Momany, M. Distinct septin heteropolymers co-exist during multicellular development in the filamentous fungus Aspergillus nidulans. PLoS ONE 2014, 9, e92819. [Google Scholar] [CrossRef]
- Li, S.; Bao, D.; Yuen, G.; Harris, S.D.; Calvo, A.M. basA regulates cell wall organization and asexual/sexual sporulation ratio in Aspergillus nidulans. Genetics 2007, 176, 243–253. [Google Scholar] [CrossRef][Green Version]
- Hill, T.W.; Loprete, D.M.; Momany, M.; Ha, Y.; Harsch, L.M.; Livesay, J.A.; Mirchandani, A.; Murdock, J.J.; Vaughan, M.J.; Watt, M.B. Isolation of cell wall mutants in Aspergillus nidulans by screening for hypersensitivity to Calcofluor White. Mycologia 2006, 98, 399–409. [Google Scholar] [CrossRef]
- Kohler, A.M.; Harting, R.; Langeneckert, A.E.; Valerius, O.; Gerke, J.; Meister, C.; Strohdiek, A.; Braus, G.H. Integration of Fungus-Specific CandA-C1 into a Trimeric CandA Complex Allowed Splitting of the Gene for the Conserved Receptor Exchange Factor of CullinA E3 Ubiquitin Ligases in Aspergilli. mBio 2019, 10, e01094-19. [Google Scholar] [CrossRef]
- Culp, D.W.; Dodge, C.L.; Miao, Y.; Li, L.; Sag-Ozkal, D.; Borgia, P.T. The chsA gene from Aspergillus nidulans is necessary for maximal conidiation. FEMS Microbiol. Lett. 2000, 182, 349–353. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Marcos, A.T.; Ramos, M.S.; Marcos, J.F.; Carmona, L.; Strauss, J.; Canovas, D. Nitric oxide synthesis by nitrate reductase is regulated during development in Aspergillus. Mol. Microbiol. 2016, 99, 15–33. [Google Scholar] [CrossRef] [PubMed]
- Marcos, A.T.; Ramos, M.S.; Schinko, T.; Strauss, J.; Canovas, D. Nitric oxide homeostasis is required for light-dependent regulation of conidiation in Aspergillus. Fungal Genet. Biol. 2020, 137, 103337. [Google Scholar] [CrossRef] [PubMed]
- Komachi, Y.; Hatakeyama, S.; Motomatsu, H.; Futagami, T.; Kizjakina, K.; Sobrado, P.; Ekino, K.; Takegawa, K.; Goto, M.; Nomura, Y.; et al. GfsA encodes a novel galactofuranosyltransferase involved in biosynthesis of galactofuranose antigen of O-glycan in Aspergillus nidulans and Aspergillus fumigatus. Mol. Microbiol. 2013, 90, 1054–1073. [Google Scholar] [CrossRef]
- Etxebeste, O.; Herrero-García, E.; Cortese, M.S.; Garzia, A.; Oiartzabal-Arano, E.; Ríos, V.D.L.; Ugalde, U.; Espeso, E.A. GmcA is a putative glucose-methanol-choline oxidoreductase required for the induction of asexual development in Aspergillus nidulans. PLoS ONE 2012, 7, e40292. [Google Scholar] [CrossRef] [PubMed]
- Takeshita, N.; Vienken, K.; Rolbetzki, A.; Fischer, R. The Aspergillus nidulans putative kinase, KfsA (kinase for septation), plays a role in septation and is required for efficient asexual spore formation. Fungal Genet. Biol. 2007, 44, 1205–1214. [Google Scholar] [CrossRef]
- Shimizu, K.; Keller, N.P. Genetic involvement of a cAMP-dependent protein kinase in a G protein signaling pathway regulating morphological and chemical transitions in Aspergillus nidulans. Genetics 2001, 157, 591–600. [Google Scholar] [CrossRef]
- Kempf, C.; Bathe, F.; Fischer, R. Evidence that two Pcl-like cyclins control Cdk9 activity during cell differentiation in Aspergillus nidulans asexual development. Eukaryot. Cell 2013, 12, 23–36. [Google Scholar] [CrossRef]
- Seo, J.A.; Yu, J.H. The phosducin-like protein PhnA is required for Gβγ-mediated signaling for vegetative growth, developmental control, and toxin biosynthesis in Aspergillus nidulans. Eukaryot. Cell 2006, 5, 400–410. [Google Scholar] [CrossRef]
- Bussink, H.J.; Osmani, S.A. A cyclin-dependent kinase family member (PHOA) is required to link developmental fate to environmental conditions in Aspergillus nidulans. EMBO J. 1998, 17, 3990–4003. [Google Scholar] [CrossRef]
- Le, T.H.T.; Oki, A.; Goto, M.; Shimizu, K. Protein O-mannosyltransferases are required for sterigmatocystin production and developmental processes in Aspergillus nidulans. Curr. Genet. 2018, 64, 1043–1056. [Google Scholar] [CrossRef] [PubMed]
- Otamendi, A.; Espeso, E.A.; Etxebeste, O. Identification and Characterization of Aspergillus nidulans Mutants Impaired in Asexual Development under Phosphate Stress. Cells 2019, 8, 1520. [Google Scholar] [CrossRef] [PubMed]
- Tsitsigiannis, D.I.; Zarnowski, R.; Keller, N.P. The lipid body protein, PpoA, coordinates sexual and asexual sporulation in Aspergillus nidulans. J. Biol. Chem. 2004, 279, 11344–11353. [Google Scholar] [CrossRef]
- Tsitsigiannis, D.I.; Kowieski, T.M.; Zarnowski, R.; Keller, N.P. Endogenous lipogenic regulators of spore balance in Aspergillus nidulans. Eukaryot. Cell 2004, 3, 1398–1411. [Google Scholar] [CrossRef]
- Son, S.H.; Jang, S.Y.; Park, H.S. Functions of PUF Family RNA-Binding Proteins in Aspergillus nidulans. J. Microbiol. Biotechnol. 2021, 31, 676–685. [Google Scholar] [CrossRef]
- Goldman, G.H.; Morris, N.R. Extragenic suppressors of a dynein mutation that blocks nuclear migration in Aspergillus nidulans. Genetics 1995, 139, 1223–1232. [Google Scholar] [CrossRef] [PubMed]
- Wilson, R.A.; Chang, P.K.; Dobrzyn, A.; Ntambi, J.M.; Zarnowski, R.; Keller, N.P. Two Δ9-stearic acid desaturases are required for Aspergillus nidulans growth and development. Fungal Genet. Biol. 2004, 41, 501–509. [Google Scholar] [CrossRef]
- Wong, K.H.; Todd, R.B.; Oakley, B.R.; Oakley, C.E.; Hynes, M.J.; Davis, M.A. Sumoylation in Aspergillus nidulans: sumO inactivation, overexpression and live-cell imaging. Fungal Genet. Biol. 2008, 45, 728–737. [Google Scholar] [CrossRef]
- Harting, R.; Bayram, O.; Laubinger, K.; Valerius, O.; Braus, G.H. Interplay of the fungal sumoylation network for control of multicellular development. Mol. Microbiol. 2013, 90, 1125–1145. [Google Scholar] [CrossRef]
- El-Ganiny, A.M.; Sheoran, I.; Sanders, D.A.; Kaminskyj, S.G. Aspergillus nidulans UDP-glucose-4-epimerase UgeA has multiple roles in wall architecture, hyphal morphogenesis, and asexual development. Fungal Genet. Biol. 2010, 47, 629–635. [Google Scholar] [CrossRef]
- Sarikaya-Bayram, O.; Bayram, O.; Feussner, K.; Kim, J.H.; Kim, H.S.; Kaever, A.; Feussner, I.; Chae, K.S.; Han, D.M.; Han, K.H.; et al. Membrane-bound methyltransferase complex VapA-VipC-VapB guides epigenetic control of fungal development. Dev. Cell 2014, 29, 406–420. [Google Scholar] [CrossRef] [PubMed]
- Futagami, T.; Nakao, S.; Kido, Y.; Oka, T.; Kajiwara, Y.; Takashita, H.; Omori, T.; Furukawa, K.; Goto, M. Putative stress sensors WscA and WscB are involved in hypo-osmotic and acidic pH stress tolerance in Aspergillus nidulans. Eukaryot. Cell 2011, 10, 1504–1515. [Google Scholar] [CrossRef] [PubMed]
- Sewall, T.C.; Mims, C.W.; Timberlake, W.E. Conidium differentiation in Aspergillus nidulans wild-type and wet-white (wetA) mutant strains. Dev. Biol. 1990, 138, 499–508. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.Y.; Mead, M.E.; Lee, M.K.; Loss, E.M.O.; Kim, S.C.; Rokas, A.; Yu, J.H. Systematic Dissection of the Evolutionarily Conserved WetA Developmental Regulator across a Genus of Filamentous Fungi. mBio 2018, 9, e01130-18. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.Y.; Mead, M.E.; Lee, M.K.; Neuhaus, G.F.; Adpressa, D.A.; Martien, J.I.; Son, Y.E.; Moon, H.; Amador-Noguez, D.; Han, K.H.; et al. Transcriptomic, Protein-DNA Interaction, and Metabolomic Studies of VosA, VelB, and WetA in Aspergillus nidulans Asexual Spores. mBio 2021, 12, e03128-20. [Google Scholar] [CrossRef] [PubMed]
- Wieser, J.; Lee, B.N.; Fondon, J., 3rd; Adams, T.H. Genetic requirements for initiating asexual development in Aspergillus nidulans. Curr. Genet. 1994, 27, 62–69. [Google Scholar] [CrossRef]
- Lee, B.N.; Adams, T.H. FluG and flbA function interdependently to initiate conidiophore development in Aspergillus nidulans through brlA β activation. EMBO J. 1996, 15, 299–309. [Google Scholar] [CrossRef]
- Seo, J.A.; Guan, Y.; Yu, J.H. FluG-dependent asexual development in Aspergillus nidulans occurs via derepression. Genetics 2006, 172, 1535–1544. [Google Scholar] [CrossRef]
- Etxebeste, O.; Ni, M.; Garzia, A.; Kwon, N.J.; Fischer, R.; Yu, J.H.; Espeso, E.A.; Ugalde, U. Basic-zipper-type transcription factor FlbB controls asexual development in Aspergillus nidulans. Eukaryot. Cell 2008, 7, 38–48. [Google Scholar] [CrossRef]
- Garzia, A.; Etxebeste, O.; Herrero-Garcia, E.; Fischer, R.; Espeso, E.A.; Ugalde, U. Aspergillus nidulans FlbE is an upstream developmental activator of conidiation functionally associated with the putative transcription factor FlbB. Mol. Microbiol. 2009, 71, 172–184. [Google Scholar] [CrossRef]
- Kwon, N.J.; Shin, K.S.; Yu, J.H. Characterization of the developmental regulator FlbE in Aspergillus fumigatus and Aspergillus nidulans. Fungal Genet. Biol. 2010, 47, 981–993. [Google Scholar] [CrossRef] [PubMed]
- Garzia, A.; Etxebeste, O.; Herrero-Garcia, E.; Ugalde, U.; Espeso, E.A. The concerted action of bZip and cMyb transcription factors FlbB and FlbD induces brlA expression and asexual development in Aspergillus nidulans. Mol. Microbiol. 2010, 75, 1314–1324. [Google Scholar] [CrossRef] [PubMed]
- Wieser, J.; Adams, T.H. flbD encodes a Myb-like DNA-binding protein that coordinates initiation of Aspergillus nidulans conidiophore development. Genes Dev. 1995, 9, 491–502. [Google Scholar] [CrossRef]
- Kwon, N.J.; Garzia, A.; Espeso, E.A.; Ugalde, U.; Yu, J.H. FlbC is a putative nuclear C2H2 transcription factor regulating development in Aspergillus nidulans. Mol. Microbiol. 2010, 77, 1203–1219. [Google Scholar] [CrossRef] [PubMed]
- Hicks, J.K.; Yu, J.H.; Keller, N.P.; Adams, T.H. Aspergillus sporulation and mycotoxin production both require inactivation of the FadA G alpha protein-dependent signaling pathway. EMBO J. 1997, 16, 4916–4923. [Google Scholar] [CrossRef] [PubMed]
- Lee, B.N.; Adams, T.H. Overexpression of flbA, an early regulator of Aspergillus asexual sporulation, leads to activation of brlA and premature initiation of development. Mol. Microbiol. 1994, 14, 323–334. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.H.; Wieser, J.; Adams, T.H. The Aspergillus FlbA RGS domain protein antagonizes G protein signaling to block proliferation and allow development. EMBO J. 1996, 15, 5184–5190. [Google Scholar] [CrossRef]
- Wieser, J.; Yu, J.H.; Adams, T.H. Dominant mutations affecting both sporulation and sterigmatocystin biosynthesis in Aspergillus nidulans. Curr. Genet. 1997, 32, 218–224. [Google Scholar] [CrossRef]
- Calvo, A.M.; Gardner, H.W.; Keller, N.P. Genetic connection between fatty acid metabolism and sporulation in Aspergillus nidulans. J. Biol. Chem. 2001, 276, 25766–25774. [Google Scholar] [CrossRef]
- Seo, J.A.; Han, K.H.; Yu, J.H. Multiple roles of a heterotrimeric G-protein γ-subunit in governing growth and development of Aspergillus nidulans. Genetics 2005, 171, 81–89. [Google Scholar] [CrossRef]
- Rosen, S.; Yu, J.H.; Adams, T.H. The Aspergillus nidulans sfaD gene encodes a G protein β subunit that is required for normal growth and repression of sporulation. EMBO J. 1999, 18, 5592–5600. [Google Scholar] [CrossRef]
- Miller, K.Y.; Wu, J.; Miller, B.L. StuA is required for cell pattern formation in Aspergillus. Genes Dev. 1992, 6, 1770–1782. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.Y.; Kim, L.H.; Kim, H.E.; Park, J.S.; Han, K.H.; Han, D.M. A putative APSES transcription factor is necessary for normal growth and development of Aspergillus nidulans. J. Microbiol. 2013, 51, 800–806. [Google Scholar] [CrossRef] [PubMed]
- Kovacs, Z.; Szarka, M.; Kovacs, S.; Boczonadi, I.; Emri, T.; Abe, K.; Pocsi, I.; Pusztahelyi, T. Effect of cell wall integrity stress and RlmA transcription factor on asexual development and autolysis in Aspergillus nidulans. Fungal Genet. Biol. 2013, 54, 1–14. [Google Scholar] [CrossRef]
- Kim, Y.J.; Yu, Y.M.; Maeng, P.J. Differential Control of Asexual Development and Sterigmatocystin Biosynthesis by a Novel Regulator in Aspergillus nidulans. Sci. Rep. 2017, 7, 46340. [Google Scholar] [CrossRef]
- Ramamoorthy, V.; Dhingra, S.; Kincaid, A.; Shantappa, S.; Feng, X.; Calvo, A.M. The putative C2H2 transcription factor MtfA is a novel regulator of secondary metabolism and morphogenesis in Aspergillus nidulans. PLoS ONE 2013, 8, e74122. [Google Scholar] [CrossRef]
- Hicks, J.; Lockington, R.A.; Strauss, J.; Dieringer, D.; Kubicek, C.P.; Kelly, J.; Keller, N. RcoA has pleiotropic effects on Aspergillus cellular development. Mol. Microbiol. 2001, 39, 1482–1493. [Google Scholar] [CrossRef] [PubMed]
- Ramamoorthy, V.; Shantappa, S.; Dhingra, S.; Calvo, A.M. veA-dependent RNA-pol II transcription elongation factor-like protein, RtfA, is associated with secondary metabolism and morphological development in Aspergillus nidulans. Mol. Microbiol. 2012, 85, 795–814. [Google Scholar] [CrossRef]
- Son, S.H.; Son, Y.E.; Cho, H.J.; Chen, W.; Lee, M.K.; Kim, L.H.; Han, D.M.; Park, H.S. Homeobox proteins are essential for fungal differentiation and secondary metabolism in Aspergillus nidulans. Sci. Rep. 2020, 10, 6094. [Google Scholar] [CrossRef]
- Kim, H.R.; Chae, K.S.; Han, K.H.; Han, D.M. The nsdC gene encoding a putative C2H2-type transcription factor is a key activator of sexual development in Aspergillus nidulans. Genetics 2009, 182, 771–783. [Google Scholar] [CrossRef]
- Lee, M.K.; Kwon, N.J.; Lee, I.S.; Jung, S.; Kim, S.C.; Yu, J.H. Negative regulation and developmental competence in Aspergillus. Sci. Rep. 2016, 6, 28874. [Google Scholar] [CrossRef] [PubMed]
- Busby, T.M.; Miller, K.Y.; Miller, B.L. Suppression and enhancement of the Aspergillus nidulans medusa mutation by altered dosage of the bristle and stunted genes. Genetics 1996, 143, 155–163. [Google Scholar] [CrossRef] [PubMed]
- Chung, D.W.; Greenwald, C.; Upadhyay, S.; Ding, S.; Wilkinson, H.H.; Ebbole, D.J.; Shaw, B.D. acon-3, the Neurospora crassa ortholog of the developmental modifier, medA, complements the conidiation defect of the Aspergillus nidulans mutant. Fungal Genet. Biol. 2011, 48, 370–376. [Google Scholar] [CrossRef] [PubMed]
- Alkahyyat, F.; Ni, M.; Kim, S.C.; Yu, J.H. The WOPR Domain Protein OsaA Orchestrates Development in Aspergillus nidulans. PLoS ONE 2015, 10, e0137554. [Google Scholar] [CrossRef]
- Navarro, R.E.; Hansberg, W.; Timberlake, W.E.; Stringer, M.A. catA, a new Aspergillus nidulans gene encoding a developmentally regulated catalase. Curr. Genet. 1996, 29, 352–359. [Google Scholar] [CrossRef]
- Kawasaki, L.; Wysong, D.; Diamond, R.; Aguirre, J. Two divergent catalase genes are differentially regulated during Aspergillus nidulans development and oxidative stress. J. Bacteriol. 1997, 179, 3284–3292. [Google Scholar] [CrossRef]
- Wang, S.; Cao, J.; Liu, X.; Hu, H.; Shi, J.; Zhang, S.; Keller, N.P.; Lu, L. Putative calcium channels CchA and MidA play the important roles in conidiation, hyphal polarity and cell wall components in Aspergillus nidulans. PLoS ONE 2012, 7, e46564. [Google Scholar] [CrossRef]
- Feng, X.; Ramamoorthy, V.; Pandit, S.S.; Prieto, A.; Espeso, E.A.; Calvo, A.M. cpsA regulates mycotoxin production, morphogenesis and cell wall biosynthesis in the fungus Aspergillus nidulans. Mol. Microbiol. 2017, 105, 1–24. [Google Scholar] [CrossRef]
- Stringer, M.A.; Timberlake, W.E. dewA encodes a fungal hydrophobin component of the Aspergillus spore wall. Mol. Microbiol. 1995, 16, 33–44. [Google Scholar] [CrossRef]
- Grunbacher, A.; Throm, T.; Seidel, C.; Gutt, B.; Rohrig, J.; Strunk, T.; Vincze, P.; Walheim, S.; Schimmel, T.; Wenzel, W.; et al. Six hydrophobins are involved in hydrophobin rodlet formation in Aspergillus nidulans and contribute to hydrophobicity of the spore surface. PLoS ONE 2014, 9, e94546. [Google Scholar] [CrossRef]
- Wartenberg, D.; Vodisch, M.; Kniemeyer, O.; Albrecht-Eckardt, D.; Scherlach, K.; Winkler, R.; Weide, M.; Brakhage, A.A. Proteome analysis of the farnesol-induced stress response in Aspergillus nidulans--The role of a putative dehydrin. J. Proteom. 2012, 75, 4038–4049. [Google Scholar] [CrossRef]
- Son, Y.E.; Cho, H.J.; Chen, W.; Son, S.H.; Lee, M.K.; Yu, J.H.; Park, H.S. The role of the VosA-repressed dnjA gene in development and metabolism in Aspergillus species. Curr. Genet. 2020, 66, 621–633. [Google Scholar] [CrossRef]
- Karacsony, Z.; Gacser, A.; Vagvolgyi, C.; Scazzocchio, C.; Hamari, Z. A dually located multi-HMG-box protein of Aspergillus nidulans has a crucial role in conidial and ascospore germination. Mol. Microbiol. 2014, 94, 383–402. [Google Scholar] [CrossRef] [PubMed]
- Donnelly, E.; Barnett, Y.A.; McCullough, W. Germinating conidiospores of Aspergillus amino acid auxotrophs are hypersensitive to heat shock, oxidative stress and DNA damage. FEBS Lett. 1994, 355, 201–204. [Google Scholar] [CrossRef] [PubMed]
- Son, Y.E.; Park, H.S. Conserved Roles of MonA in Fungal Growth and Development in Aspergillus Species. Mycobiology 2019, 47, 457–465. [Google Scholar] [CrossRef]
- Lim, J.Y.; Jang, S.H.; Park, H.M. Mannitol-1-phosphate dehydrogenase, MpdA, is required for mannitol production in vegetative cells and involved in hyphal branching, heat resistance of conidia and sexual development in Aspergillus nidulans. Curr. Genet. 2021, 67, 613–630. [Google Scholar] [CrossRef]
- Futagami, T.; Seto, K.; Kajiwara, Y.; Takashita, H.; Omori, T.; Takegawa, K.; Goto, M. The putative stress sensor protein MtlA is required for conidia formation, cell wall stress tolerance, and cell wall integrity in Aspergillus nidulans. Biosci. Biotechnol. Biochem. 2014, 78, 326–335. [Google Scholar] [CrossRef] [PubMed]
- Fillinger, S.; Chaveroche, M.K.; van Dijck, P.; de Vries, R.; Ruijter, G.; Thevelein, J.; d’Enfert, C. Trehalose is required for the acquisition of tolerance to a variety of stresses in the filamentous fungus Aspergillus nidulans. Microbiology 2001, 147, 1851–1862. [Google Scholar] [CrossRef]
- Ahmed, Y.L.; Gerke, J.; Park, H.S.; Bayram, O.; Neumann, P.; Ni, M.; Dickmanns, A.; Kim, S.C.; Yu, J.H.; Braus, G.H.; et al. The velvet family of fungal regulators contains a DNA-binding domain structurally similar to NF-κB. PLoS Biol. 2013, 11, e1001750. [Google Scholar] [CrossRef]
- Park, H.S.; Ni, M.; Jeong, K.C.; Kim, Y.H.; Yu, J.H. The role, interaction and regulation of the velvet regulator VelB in Aspergillus nidulans. PLoS ONE 2012, 7, e45935. [Google Scholar] [CrossRef]
- Park, H.-S.; Yu, Y.M.; Lee, M.-K.; Maeng, P.J.; Kim, S.C.; Yu, J.-H. Velvet-mediated repression of β-glucan synthesis in Aspergillus nidulans spores. Sci. Rep. 2015, 5, 10199. [Google Scholar] [CrossRef] [PubMed]
- Park, H.S.; Lee, M.K.; Kim, S.C.; Yu, J.H. The role of VosA/VelB-activated developmental gene vadA in Aspergillus nidulans. PLoS ONE 2017, 12, e0177099. [Google Scholar] [CrossRef] [PubMed]
- Son, Y.E.; Park, H.S. Genome Wide Analysis Reveals the Role of VadA in Stress Response, Germination, and Sterigmatocystin Production in Aspergillus nidulans Conidia. Microorganisms 2020, 8, 1319. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Lee, M.K.; Lim, J.; Moon, H.; Park, H.S.; Zheng, W.; Yu, J.H. The putative sensor histidine kinase VadJ coordinates development and sterigmatocystin production in Aspergillus nidulans. J. Microbiol. 2021, 59, 746–752. [Google Scholar] [CrossRef]
- Zhao, Y.; Lee, M.K.; Lim, J.; Moon, H.; Park, H.S.; Zheng, W.; Yu, J.H. The velvet-activated putative C6 transcription factor VadZ regulates development and sterigmatocystin production in Aspergillus nidulans. Fungal Biol. 2022, 126, 421–428. [Google Scholar] [CrossRef]
- Kim, M.J.; Jung, W.H.; Son, Y.E.; Yu, J.H.; Lee, M.K.; Park, H.S. The velvet repressed vidA gene plays a key role in governing development in Aspergillus nidulans. J. Microbiol. 2019, 57, 893–899. [Google Scholar] [CrossRef]
- Son, Y.E.; Park, H.S. Unveiling the Functions of the VosA-VelB Target Gene vidD in Aspergillus nidulans. Mycobiology 2021, 49, 258–266. [Google Scholar] [CrossRef]
- Lee, M.K.; Son, Y.E.; Park, H.S.; Alshannaq, A.; Han, K.H.; Yu, J.H. Velvet activated McrA plays a key role in cellular and metabolic development in Aspergillus nidulans. Sci. Rep. 2020, 10, 15075. [Google Scholar] [CrossRef]
- Thieme, K.G.; Gerke, J.; Sasse, C.; Valerius, O.; Thieme, S.; Karimi, R.; Heinrich, A.K.; Finkernagel, F.; Smith, K.; Bode, H.B.; et al. Velvet domain protein VosA represses the zinc cluster transcription factor SclB regulatory network for Aspergillus nidulans asexual development, oxidative stress response and secondary metabolism. PLoS Genet. 2018, 14, e1007511. [Google Scholar] [CrossRef]
- Son, Y.E.; Cho, H.J.; Lee, M.K.; Park, H.S. Characterizing the role of Zn cluster family transcription factor ZcfA in governing development in two Aspergillus species. PLoS ONE 2020, 15, e0228643. [Google Scholar] [CrossRef]
- Son, S.H.; Lee, M.K.; Son, Y.E.; Park, H.S. HbxB Is a Key Regulator for Stress Response and β-Glucan Biogenesis in Aspergillus nidulans. Microorganisms 2021, 9, 144. [Google Scholar] [CrossRef]
- Cho, H.J.; Park, H.S. The function of a conidia specific transcription factor CsgA in Aspergillus nidulans. Sci. Rep. 2022, 12, 15588. [Google Scholar] [CrossRef]
- Hagiwara, D.; Matsubayashi, Y.; Marui, J.; Furukawa, K.; Yamashino, T.; Kanamaru, K.; Kato, M.; Abe, K.; Kobayashi, T.; Mizuno, T. Characterization of the NikA histidine kinase implicated in the phosphorelay signal transduction of Aspergillus nidulans, with special reference to fungicide responses. Biosci. Biotechnol. Biochem. 2007, 71, 844–847. [Google Scholar] [CrossRef] [PubMed]
- Jaimes-Arroyo, R.; Lara-Rojas, F.; Bayram, O.; Valerius, O.; Braus, G.H.; Aguirre, J. The SrkA Kinase Is Part of the SakA Mitogen-Activated Protein Kinase Interactome and Regulates Stress Responses and Development in Aspergillus nidulans. Eukaryot. Cell 2015, 14, 495–510. [Google Scholar] [CrossRef] [PubMed]
- Garrido-Bazan, V.; Jaimes-Arroyo, R.; Sanchez, O.; Lara-Rojas, F.; Aguirre, J. SakA and MpkC Stress MAPKs Show Opposite and Common Functions During Stress Responses and Development in Aspergillus nidulans. Front. Microbiol. 2018, 9, 2518. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.Y.; Chun, J.; Jun, S.C.; Han, D.M.; Chae, K.S.; Jahng, K.Y. The MpkB MAP kinase plays a role in autolysis and conidiation of Aspergillus nidulans. Fungal Genet. Biol. 2013, 61, 42–49. [Google Scholar] [CrossRef] [PubMed]
- Fujioka, T.; Mizutani, O.; Furukawa, K.; Sato, N.; Yoshimi, A.; Yamagata, Y.; Nakajima, T.; Abe, K. MpkA-Dependent and -independent cell wall integrity signaling in Aspergillus nidulans. Eukaryot. Cell 2007, 6, 1497–1510. [Google Scholar] [CrossRef] [PubMed]
- Vargas-Perez, I.; Sanchez, O.; Kawasaki, L.; Georgellis, D.; Aguirre, J. Response regulators SrrA and SskA are central components of a phosphorelay system involved in stress signal transduction and asexual sporulation in Aspergillus nidulans. Eukaryot. Cell 2007, 6, 1570–1583. [Google Scholar] [CrossRef]
- Lara-Rojas, F.; Sanchez, O.; Kawasaki, L.; Aguirre, J. Aspergillus nidulans transcription factor AtfA interacts with the MAPK SakA to regulate general stress responses, development and spore functions. Mol. Microbiol. 2011, 80, 436–454. [Google Scholar] [CrossRef]
- Kocsis, B.; Lee, M.K.; Yu, J.H.; Nagy, T.; Daroczi, L.; Batta, G.; Pocsi, I.; Leiter, E. Functional analysis of the bZIP-type transcription factors AtfA and AtfB in Aspergillus nidulans. Front. Microbiol. 2022, 13, 1003709. [Google Scholar] [CrossRef]
- Mendoza-Martinez, A.E.; Lara-Rojas, F.; Sanchez, O.; Aguirre, J. NapA Mediates a Redox Regulation of the Antioxidant Response, Carbon Utilization and Development in Aspergillus nidulans. Front. Microbiol. 2017, 8, 516. [Google Scholar] [CrossRef]
- Bok, J.W.; Wiemann, P.; Garvey, G.S.; Lim, F.Y.; Haas, B.; Wortman, J.; Keller, N.P. Illumina identification of RsrA, a conserved C2H2 transcription factor coordinating the NapA mediated oxidative stress signaling pathway in Aspergillus. BMC Genom. 2014, 15, 1011. [Google Scholar] [CrossRef]
- Fischer, R.; Timberlake, W.E. Aspergillus nidulans apsA (anucleate primary sterigmata) encodes a coiled-coil protein required for nuclear positioning and completion of asexual development. J. Cell Biol. 1995, 128, 485–498. [Google Scholar] [CrossRef]
- Clutterbuck, A.J. Mutants of Aspergillus nidulans deficient in nuclear migration during hyphal growth and conidiation. Microbiology 1994, 140 Pt 5, 1169–1174. [Google Scholar] [CrossRef][Green Version]
- Lindsey, R.; Cowden, S.; Hernandez-Rodriguez, Y.; Momany, M. Septins AspA and AspC are important for normal development and limit the emergence of new growth foci in the multicellular fungus Aspergillus nidulans. Eukaryot. Cell 2010, 9, 155–163. [Google Scholar] [CrossRef]
- Hernandez-Rodriguez, Y.; Hastings, S.; Momany, M. The septin AspB in Aspergillus nidulans forms bars and filaments and plays roles in growth emergence and conidiation. Eukaryot. Cell 2012, 11, 311–323. [Google Scholar] [CrossRef] [PubMed]
- Belaish, R.; Sharon, H.; Levdansky, E.; Greenstein, S.; Shadkchan, Y.; Osherov, N. The Aspergillus nidulans cetA and calA genes are involved in conidial germination and cell wall morphogenesis. Fungal Genet. Biol. 2008, 45, 232–242. [Google Scholar] [CrossRef] [PubMed]
- Rasmussen, C.D.; Lu, K.P.; Means, R.L.; Means, A.R. Calmodulin and cell cycle control. J. Physiol. 1992, 86, 83–88. [Google Scholar] [CrossRef]
- Yamazaki, H.; Tanaka, A.; Kaneko, J.; Ohta, A.; Horiuchi, H. Aspergillus nidulans ChiA is a glycosylphosphatidylinositol (GPI)-anchored chitinase specifically localized at polarized growth sites. Fungal Genet. Biol. 2008, 45, 963–972. [Google Scholar] [CrossRef] [PubMed]
- Joseph, J.D.; Means, A.R. Identification and characterization of two Ca2+/CaM-dependent protein kinases required for normal nuclear division in Aspergillus nidulans. J. Biol. Chem. 2000, 275, 38230–38238. [Google Scholar] [CrossRef]
- Suzuki, S.; Bayram, O.S.; Bayram, O.; Braus, G.H. conF and conJ contribute to conidia germination and stress response in the filamentous fungus Aspergillus nidulans. Fungal Genet. Biol. 2013, 56, 42–53. [Google Scholar] [CrossRef]
- Johns, S.A.; Leeder, A.C.; Safaie, M.; Turner, G. Depletion of Aspergillus nidulans cotA causes a severe polarity defect which is not suppressed by the nuclear migration mutation nudA2. Mol. Genet. Genom. 2006, 275, 593–604. [Google Scholar] [CrossRef]
- Shi, J.; Chen, W.; Liu, Q.; Chen, S.; Hu, H.; Turner, G.; Lu, L. Depletion of the MobB and CotA complex in Aspergillus nidulans causes defects in polarity maintenance that can be suppressed by the environment stress. Fungal Genet. Biol. 2008, 45, 1570–1581. [Google Scholar] [CrossRef] [PubMed]
- Kong, Q.; Wang, L.; Liu, Z.; Kwon, N.J.; Kim, S.C.; Yu, J.H. Gβ-like CpcB plays a crucial role for growth and development of Aspergillus nidulans and Aspergillus fumigatus. PLoS ONE 2013, 8, e70355. [Google Scholar] [CrossRef] [PubMed]
- Schultzhaus, Z.; Yan, H.; Shaw, B.D. Aspergillus nidulans flippase DnfA is cargo of the endocytic collar and plays complementary roles in growth and phosphatidylserine asymmetry with another flippase, DnfB. Mol. Microbiol. 2015, 97, 18–32. [Google Scholar] [CrossRef] [PubMed]
- Rohrig, J.; Kastner, C.; Fischer, R. Light inhibits spore germination through phytochrome in Aspergillus nidulans. Curr. Genet. 2013, 59, 55–62. [Google Scholar] [CrossRef] [PubMed]
- Harispe, L.; Portela, C.; Scazzocchio, C.; Penalva, M.A.; Gorfinkiel, L. Ras GTPase-activating protein regulation of actin cytoskeleton and hyphal polarity in Aspergillus nidulans. Eukaryot. Cell 2008, 7, 141–153. [Google Scholar] [CrossRef]
- Bakti, F.; Kiraly, A.; Orosz, E.; Miskei, M.; Emri, T.; Leiter, E.; Pocsi, I. Study on the glutathione metabolism of the filamentous fungus Aspergillus nidulans. Acta Microbiol. Immunol. Hung. 2017, 64, 255–272. [Google Scholar] [CrossRef][Green Version]
- Dos Reis, T.F.; Mellado, L.; Lohmar, J.M.; Silva, L.P.; Zhou, J.J.; Calvo, A.M.; Goldman, G.H.; Brown, N.A. GPCR-mediated glucose sensing system regulates light-dependent fungal development and mycotoxin production. PLoS Genet. 2019, 15, e1008419. [Google Scholar] [CrossRef]
- Amon, J.; Varga, G.; Pfeiffer, I.; Farkas, Z.; Karacsony, Z.; Hegedus, Z.; Vagvolgyi, C.; Hamari, Z. The role of the Aspergillus nidulans high mobility group B protein HmbA, the orthologue of Saccharomyces cerevisiae Nhp6p. Sci. Rep. 2022, 12, 17334. [Google Scholar] [CrossRef]
- Kang, E.H.; Kim, J.A.; Oh, H.W.; Park, H.M. LAMMER Kinase LkhA plays multiple roles in the vegetative growth and asexual and sexual development of Aspergillus nidulans. PLoS ONE 2013, 8, e58762. [Google Scholar] [CrossRef]
- Harris, S.D. Morphogenesis is coordinated with nuclear division in germinating Aspergillus nidulans conidiospores. Microbiology 1999, 145 Pt 10, 2747–2756. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Kim, J.M.; Song, H.Y.; Choi, H.J.; So, K.K.; Kim, D.H.; Chae, K.S.; Han, D.M.; Jahng, K.Y. Characterization of NpgA, a 4′-phosphopantetheinyl transferase of Aspergillus nidulans, and evidence of its involvement in fungal growth and formation of conidia and cleistothecia for development. J. Microbiol. 2015, 53, 21–31. [Google Scholar] [CrossRef]
- Hynes, M.J.; Murray, S.L.; Khew, G.S.; Davis, M.A. Genetic analysis of the role of peroxisomes in the utilization of acetate and fatty acids in Aspergillus nidulans. Genetics 2008, 178, 1355–1369. [Google Scholar] [CrossRef]
- Ahn, C.S.; Oh, Y.; Kim, J.G.; Han, K.H.; Lee, C.W.; Kim, J.W. The observation of plcA mutation and localization in Aspergillus nidulans. J. Microbiol. 2014, 52, 590–596. [Google Scholar] [CrossRef]
- Jiang, P.; Wei, W.F.; Zhong, G.W.; Zhou, X.G.; Qiao, W.R.; Fisher, R.; Lu, L. The function of the three phosphoribosyl pyrophosphate synthetase (Prs) genes in hyphal growth and conidiation in Aspergillus nidulans. Microbiology 2017, 163, 218–232. [Google Scholar] [CrossRef] [PubMed]
- Osherov, N.; May, G. Conidial germination in Aspergillus nidulans requires RAS signaling and protein synthesis. Genetics 2000, 155, 647–656. [Google Scholar] [CrossRef]
- Kwon, N.J.; Park, H.S.; Jung, S.; Kim, S.C.; Yu, J.H. The putative guanine nucleotide exchange factor RicA mediates upstream signaling for growth and development in Aspergillus. Eukaryot. Cell 2012, 11, 1399–1412. [Google Scholar] [CrossRef] [PubMed]
- Fillinger, S.; Chaveroche, M.K.; Shimizu, K.; Keller, N.; d’Enfert, C. cAMP and ras signalling independently control spore germination in the filamentous fungus Aspergillus nidulans. Mol. Microbiol. 2002, 44, 1001–1016. [Google Scholar] [CrossRef]
- Harris, S.D.; Hofmann, A.F.; Tedford, H.W.; Lee, M.P. Identification and characterization of genes required for hyphal morphogenesis in the filamentous fungus Aspergillus nidulans. Genetics 1999, 151, 1015–1025. [Google Scholar] [CrossRef]
- Whittaker, S.L.; Lunness, P.; Milward, K.J.; Doonan, J.H.; Assinder, S.J. sodVIC is an α-COP-related gene which is essential for establishing and maintaining polarized growth in Aspergillus nidulans. Fungal Genet. Biol. 1999, 26, 236–252. [Google Scholar] [CrossRef]
- Lim, J.Y.; Kang, E.H.; Park, Y.H.; Kook, J.H.; Park, H.M. Survival factor SvfA plays multiple roles in differentiation and is essential for completion of sexual development in Aspergillus nidulans. Sci. Rep. 2020, 10, 5586. [Google Scholar] [CrossRef]
- Upadhyay, S.; Shaw, B.D. A phosphoglucose isomerase mutant in Aspergillus nidulans is defective in hyphal polarity and conidiation. Fungal Genet. Biol. 2006, 43, 739–751. [Google Scholar] [CrossRef]
- Momany, M.; Westfall, P.J.; Abramowsky, G. Aspergillus nidulans swo mutants show defects in polarity establishment, polarity maintenance and hyphal morphogenesis. Genetics 1999, 151, 557–567. [Google Scholar] [CrossRef] [PubMed]
- Higashitsuji, Y.; Herrero, S.; Takeshita, N.; Fischer, R. The cell end marker protein TeaC is involved in growth directionality and septation in Aspergillus nidulans. Eukaryot. Cell 2009, 8, 957–967. [Google Scholar] [CrossRef] [PubMed]
- Afroz, S.; El-Ganiny, A.M.; Sanders, D.A.; Kaminskyj, S.G. Roles of the Aspergillus nidulans UDP-galactofuranose transporter, UgtA in hyphal morphogenesis, cell wall architecture, conidiation, and drug sensitivity. Fungal Genet. Biol. 2011, 48, 896–903. [Google Scholar] [CrossRef] [PubMed]
- Hoshi, H.O.; Zheng, L.; Ohta, A.; Horiuchi, H. A Wiskott-Aldrich syndrome protein is involved in endocytosis in Aspergillus nidulans. Biosci. Biotechnol. Biochem. 2016, 80, 1802–1812. [Google Scholar] [CrossRef] [PubMed]
- Colabardini, A.C.; Brown, N.A.; Savoldi, M.; Goldman, M.H.; Goldman, G.H. Functional characterization of Aspergillus nidulans ypkA, a homologue of the mammalian kinase SGK. PLoS ONE 2013, 8, e57630. [Google Scholar] [CrossRef]
- Ries, L.N.; Beattie, S.R.; Espeso, E.A.; Cramer, R.A.; Goldman, G.H. Diverse Regulation of the CreA Carbon Catabolite Repressor in Aspergillus nidulans. Genetics 2016, 203, 335–352. [Google Scholar] [CrossRef]
- Dowzer, C.E.; Kelly, J.M. Analysis of the creA gene, a regulator of carbon catabolite repression in Aspergillus nidulans. Mol. Cell. Biol. 1991, 11, 5701–5709. [Google Scholar] [CrossRef]
- Ni, M.; Rierson, S.; Seo, J.A.; Yu, J.H. The pkaB gene encoding the secondary protein kinase A catalytic subunit has a synthetic lethal interaction with pkaA and plays overlapping and opposite roles in Aspergillus nidulans. Eukaryot. Cell 2005, 4, 1465–1476. [Google Scholar] [CrossRef] [PubMed]
- Han, K.H.; Seo, J.A.; Yu, J.H. Regulators of G-protein signalling in Aspergillus nidulans: RgsA downregulates stress response and stimulates asexual sporulation through attenuation of GanB (Gα) signalling. Mol. Microbiol. 2004, 53, 529–540. [Google Scholar] [CrossRef] [PubMed]




| Name | Stages | Description | Reference(s) |
|---|---|---|---|
| DnfD | Vesicles | Phospholipid flippase, homolog of S. cerevisiae Drs2-Neo1-Family Neo1 | [22] |
| PabA | Vesicles | Putative regulatory subunit of protein phosphatase 2A (PP2A) | [23] |
| DopA | Vesicles, Metulae | Dopey protein | [22] |
| NimX | Vesicles, Metulae | Cyclin-dependent kinase involved in cell cycle control | [24] |
| GmtA/B | Metulae | Putative GDP-mannose transporter | [25] |
| HymA | Metulae | Hypha-like metulae | [26] |
| BemA | Metulae, Phialides | Protein kinase activator | [27] |
| FigA | Metulae, Phialides | Putative low-affinity calcium family protein | [28] |
| GcnE | Metulae, Phialides | SAGA complex histone H3K9 acetyltransferase catalytic subunit | [29] |
| ParA | Metulae, Phialides | Putative regulatory subunit of protein phosphatase 2A (PP2A) | [23] |
| UgmA | Metulae, Phialides | UDP-galactopyranose mutase | [30] |
| Axl2 | Phialides | Axial budding positional marker | [31] |
| PclA | Phialides | Homolog of S. cerevisiae pcl cyclins | [32] |
| SnaD | Phialides | Coiled-coil protein associated with the spindle pole body, named after the suppressor of nudA1 | [33] |
| SthA/B | Phialides | Sthenyo | [34] |
| TcsA | Phialides | Two-component signaling protein involved in oxidative/osmotic stress pathway | [35] |
| Name | Conidiogenesis | Conidial Viability | Conidial Stress Response | Description | Reference(s) |
|---|---|---|---|---|---|
| CatA | Resistant to H2O2 | Spore-specific catalase A, involved in oxidative/osmotic stress | [105] | ||
| CatB | Resistant to H2O2 | Catalase B, involved in oxidative/osmotic stress | [106] | ||
| CchA | Activation | Sensitive to CFW and CR | Calcium channel | [107] | |
| CpsA | Activation | Resistant to MSB, SDS and CFW | Capsule polysaccharide synthase | [108] | |
| DewA | Spore-wall fungal hydrophobin, named after detergent wettable | [109,110] | |||
| DlpA | Resistant to heat and H2O2 | Dehydrin-like protein | [111] | ||
| DnjA | Activation | Maintenance | Resistant to heat | Putative DnaJ proteinRegulation of trehalose biosynthesis | [112] |
| HmbB | Activation | Maintenance | Putative high-mobility group box proteinRegulation of trehalose biosynthesis and proper germination | [113] | |
| LysB/D | Resistant to heat, UV and H2O2 | Homoisocitrate dehydrogenase/synthase | [114] | ||
| MidA | Activation | Sensitive to CFW and CR | Stretch-activated calcium channels, named after mating-induced death | [107] | |
| MonA | Activation | Maintenance | Resistant to heat | A subunit of a guanine nucleotide exchange factor Regulation of trehalose biosynthesis | [115] |
| MpdA | Resistant to heat Sensitive to benomyl | Mannitol-1-phosphate 5-dehydrogenase | [116] | ||
| MtlA | Activation | Resistant to CFW and CR | Mid2-like protein | [117] | |
| PufE | Repression | Maintenance | Resistant to heat | Pumilio/fem-3 binding factor Regulation of trehalose biosynthesis | [65] |
| RodA | Rodlet protein composed of fungal spore wall | [110] | |||
| TpsA | Maintenance | Resistant to heat and H2O2 | Trehalose-6-phosphate synthase Regulation of trehalose biosynthesis | [118] |
| Name | Conidiogenesis | Germination | Description | Reference(s) |
|---|---|---|---|---|
| ArgB | Activation | Activation | Ornithine carbamoyl transferase, homolog of S. cerevisiae Arg3 Conidial resistant to heat, UV, and H2O2 | [114] |
| ApsA | Activation | Activation | Anucleate primary sterigmata protein A | [143,144] |
| ApsB | Activation | Activation | Anucleate primary sterigmata protein B | [144] |
| AspA | Activation | Activation | Aspergillus septin A | [145] |
| AspB | Activation | Activation | Aspergillus septin B | [146] |
| AspC | Activation | Activation | Aspergillus septin C | [145] |
| CalA | Activation | Activation | Fungal thaumatin-like proteins, named after calcoflour hypersensitivity | [49,147] |
| CaM | Activation | Calmodulin involved in calcium-calcineurin signaling | [148] | |
| CetA | Activation | Fungal thaumatin-like proteins, named after conidial-enriched transcripts | [147] | |
| ChiA | Activation | Chitinase involved in fungal cell-wall integrity signaling (CWIS) | [149] | |
| CmkA/B | Activation | Ca2+/caM-dependent protein kinase involved in calcium-calcineurin signaling | [150] | |
| ConF/J | Activation | Conidiation-specific gene Conidial sensitive to polyol | [151] | |
| CotA | Activation | Activation | NDR protein kinase, homolog of S. cerevisiae Cbk1 | [152,153] |
| CpcB | Activation | Activation | Gβ-like protein, named after Cross pathway control WD repeat protein B | [154] |
| DnfA | Activation | Activation | Phospholipid flippases, homolog of S. cerevisiae Drs2-Neo1-Family Dnf1/2 | [22,155] |
| DnfB | Activation | Activation | Phospholipid flippases | [22,155] |
| FphA | Activation | Fungal phytochrome (red light-sensing photoreceptor) | [156] | |
| GapA | Activation | Activation | Ras GTPase-activating protein | [157] |
| GcsA | Activation | Glucosylceramide synthase | [158] | |
| GlrA | Activation | Glutathione reductase | [158] | |
| GprH | Activation | G protein-coupled receptor | [159] | |
| HmbA | Activation | Activation | High-mobility-group B protein A | [160] |
| LkhA | Activation | Activation | LAMMER kinase, homolog of S. pombe Lkh1 | [161] |
| MobB | Activation | Activation | Homolog of S. cerevisiae Mob2 | [153] |
| NimA | Activation | Cell-cycle regulated serine/threonine protein kinase, homolog of S. pombe Kin3 | [162] | |
| NpgA | Activation | Activation | 4′-phosphopantetheinyl transferase, named after the null pigmentation mutant | [163] |
| PexC PexE~G | Activation | Activation | Peroxisome biogenesis protein (peroxin) | [164] |
| PclA | Activation | Phosphatidylinositol phospholipase, homolog of S. cerevisiae Pcl1 Conidial resistant to cold | [165] | |
| PrsB/C | Activation | Phosphoribosyl pyrophosphate synthetase | [166] | |
| RasA | Activation | Ras-like protein | [167] | |
| RicA | Activation | Activation | GDP/GTP exchange factor in Heterotrimeric G protein signaling | [168] |
| SchA | Activation | Sch9-like kinase | [169] | |
| SepA | Activation | Formin protein involved in the formation of an actin ring at the septation site | [170] | |
| SodVIC | Activation | α-COP-like protein, named after the stabilization of disomy | [171] | |
| SvfA | Activation | Activation | Homolog of S. cerevisiae survival factor 1 Conidial resistant to cold, MSB and H2O2 | [172] |
| PmtA (=SwoA) | Activation | Protein O-mannosyltransferase (Swollen cells) Conidial resistant to CFW | [173,174] | |
| SwoB | Activation | Activation | Swollen cells Conidial resistant to CFW | [174] |
| TeaC | Activation | Cell end marker protein | [175] | |
| UgtA | Activation | Activation | UDP-Galf transporter | [176] |
| WspA | Activation | Activation | Wiskott–Aldrich syndrome protein | [177] |
| YpkA | Activation | Activation | Polyphosphate kinase, homolog of S. cerevisiae Ypk1 | [178] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Son, Y.-E.; Yu, J.-H.; Park, H.-S. Regulators of the Asexual Life Cycle of Aspergillus nidulans. Cells 2023, 12, 1544. https://doi.org/10.3390/cells12111544
Son Y-E, Yu J-H, Park H-S. Regulators of the Asexual Life Cycle of Aspergillus nidulans. Cells. 2023; 12(11):1544. https://doi.org/10.3390/cells12111544
Chicago/Turabian StyleSon, Ye-Eun, Jae-Hyuk Yu, and Hee-Soo Park. 2023. "Regulators of the Asexual Life Cycle of Aspergillus nidulans" Cells 12, no. 11: 1544. https://doi.org/10.3390/cells12111544
APA StyleSon, Y.-E., Yu, J.-H., & Park, H.-S. (2023). Regulators of the Asexual Life Cycle of Aspergillus nidulans. Cells, 12(11), 1544. https://doi.org/10.3390/cells12111544