

Loss of Astrocytic µ Opioid Receptors Exacerbates Aversion Associated with Morphine Withdrawal in Mice: Role of Mitochondrial Respiration

 , , , , ,
, , , , ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Mice
2.2. Tamoxifen Injections
2.3. Morphine and/or Naloxone Injections
2.4. Behavioral Testing
2.4.1. Open Field Test
2.4.2. Elevated Plus Maze
2.4.3. Novel Place Recognition
2.4.4. Morphine-Induced Sensitization
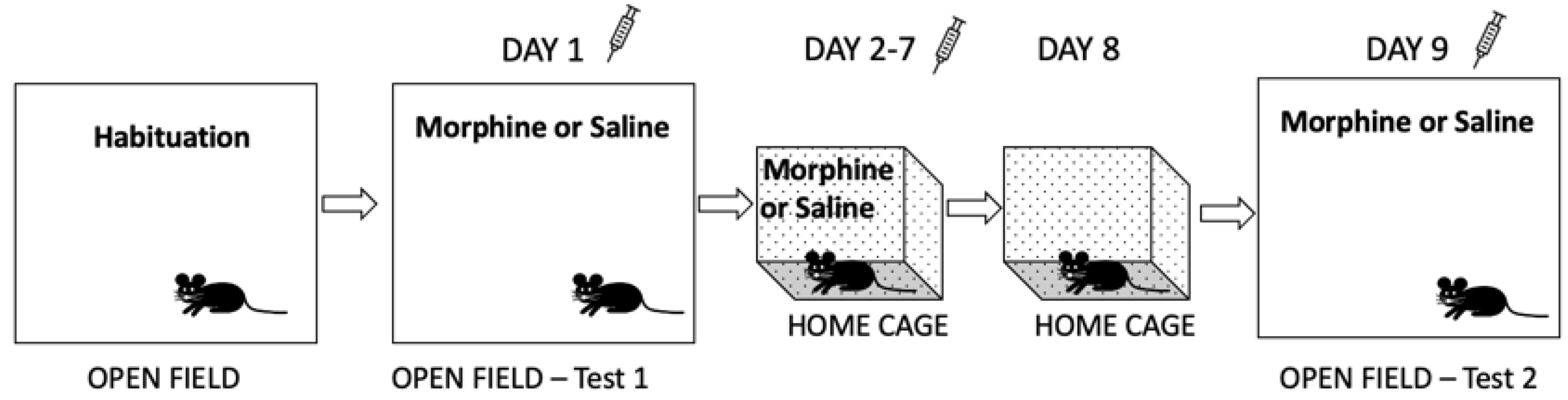
2.4.5. Conditioned Place Preference
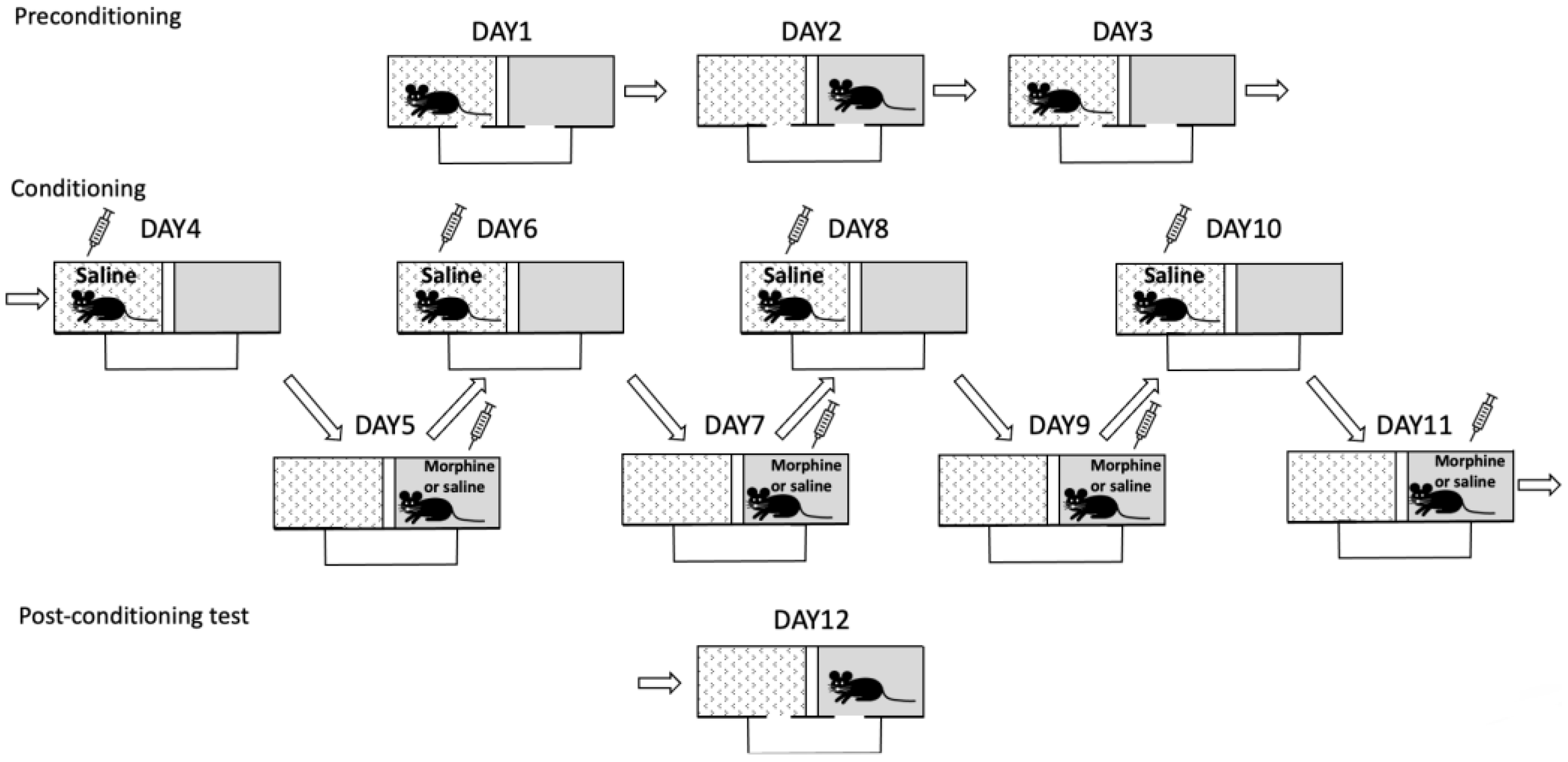
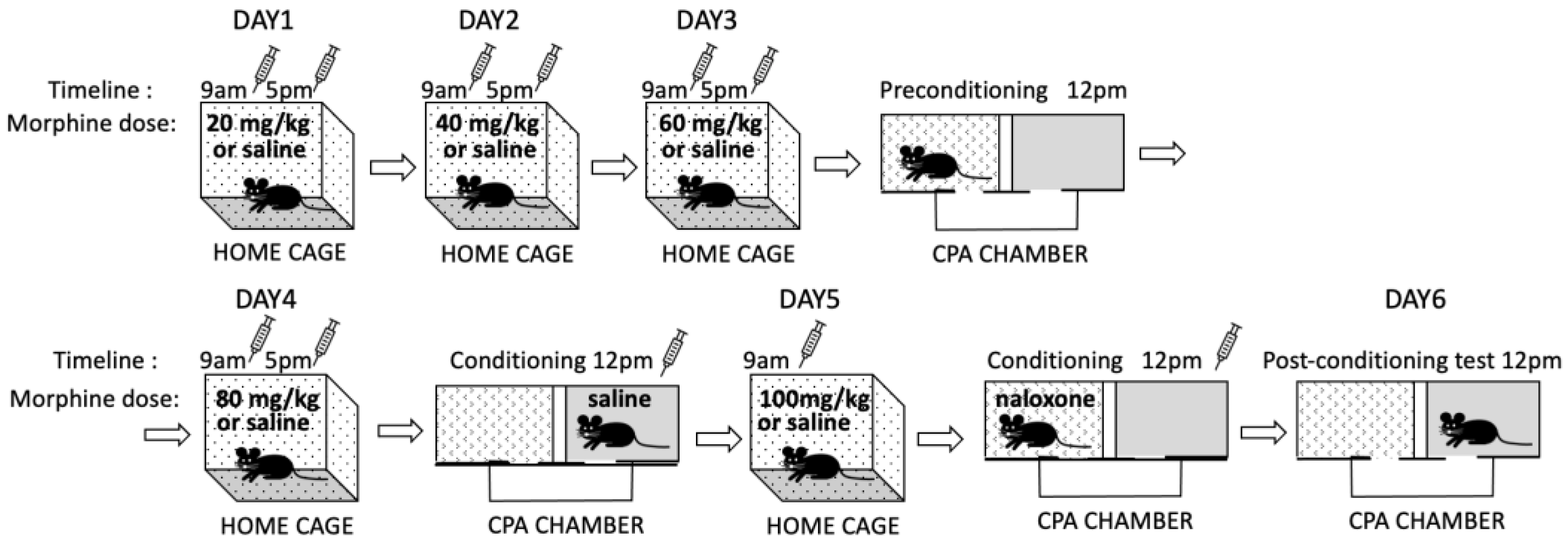
2.4.6. Conditioned Place Aversion
2.4.7. Analgesic Effects
2.5. RNA In Situ Hybridization
2.6. Biochemical Assays
2.6.1. Western Blotting
2.6.2. Metabolic Flux Assays
2.7. Statistical Analysis
3. Results
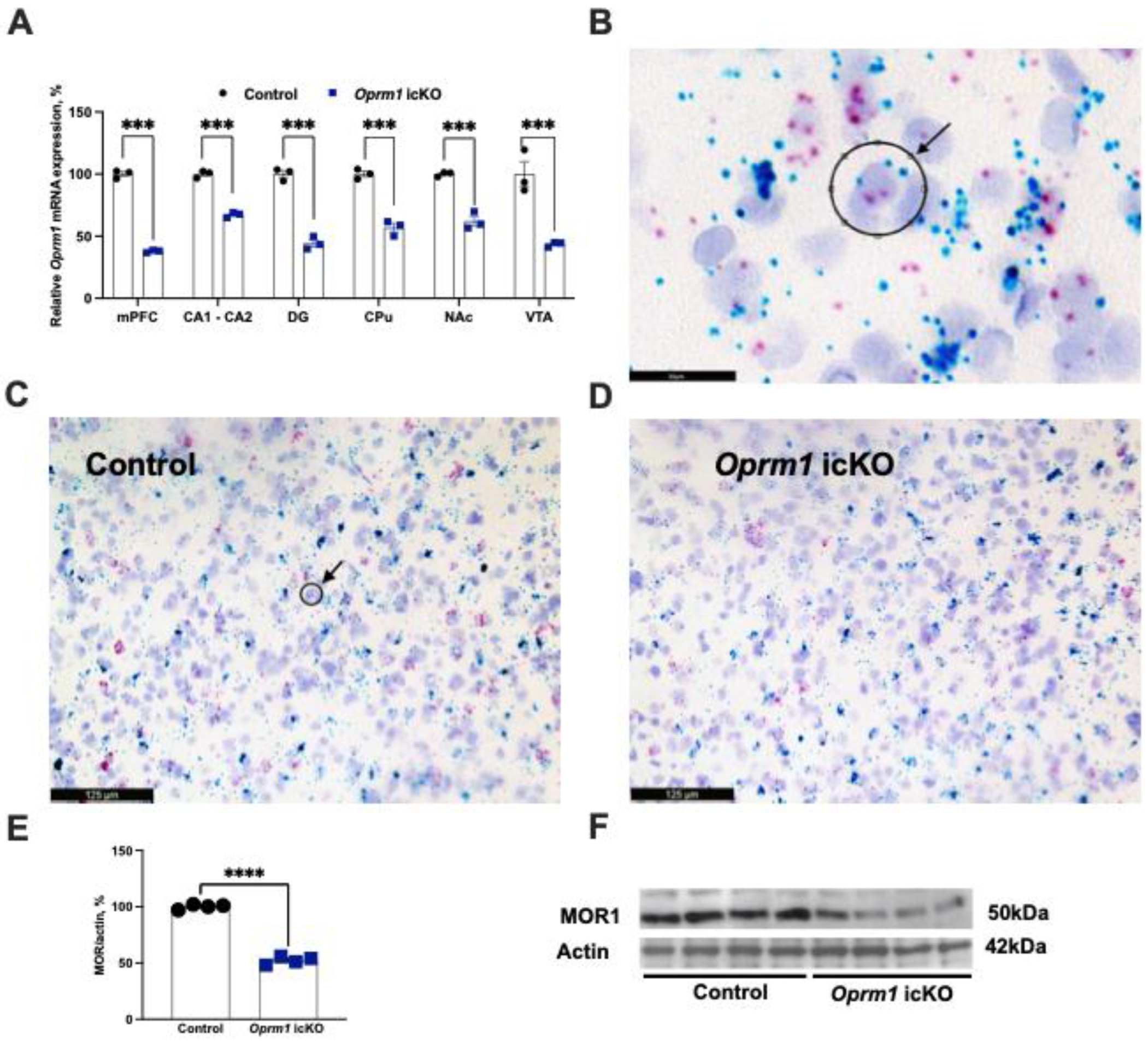
3.1. Decreased Levels of Oprm1 mRNA and Protein in Astrocytes from icKO Mice
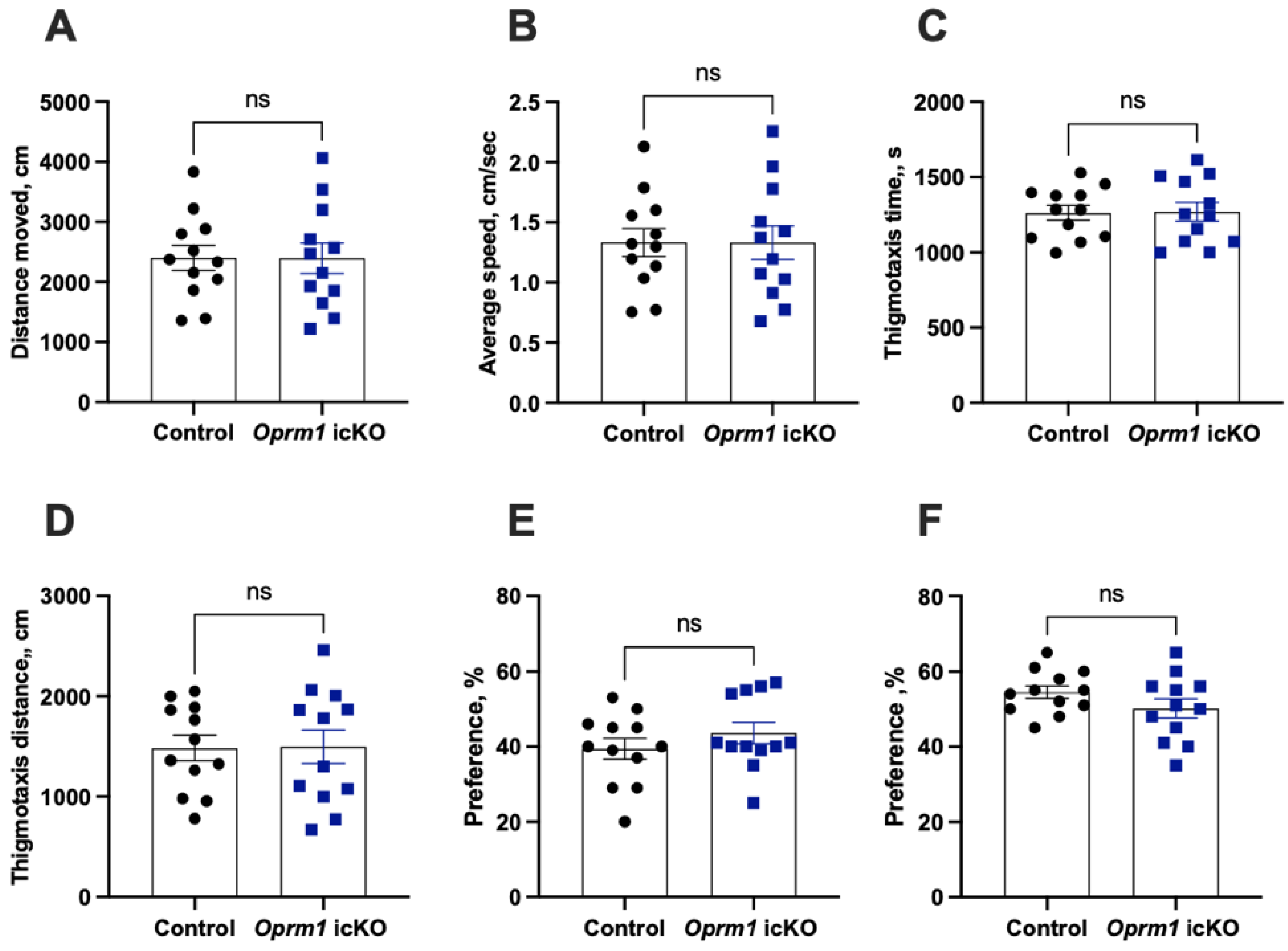
3.2. Normal Baseline Behavioral Phenotypes of Oprm1 icKO Mice
3.3. Effects of Oprm1 icKO on Drug-Induced Behaviors
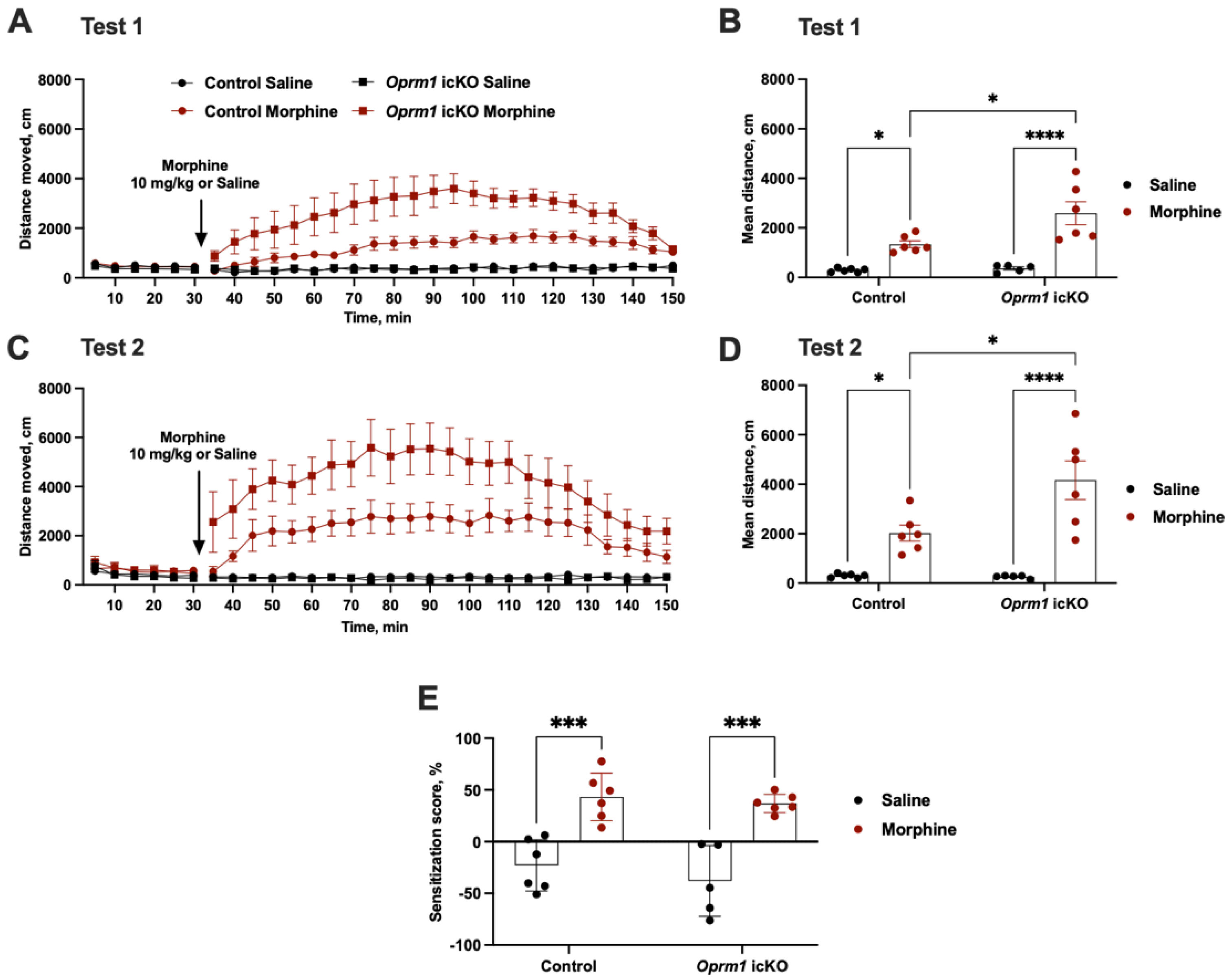
3.3.1. Increased Morphine-Induced Locomotor Activity but Not Locomotor Sensitization with Oprm1 icKO
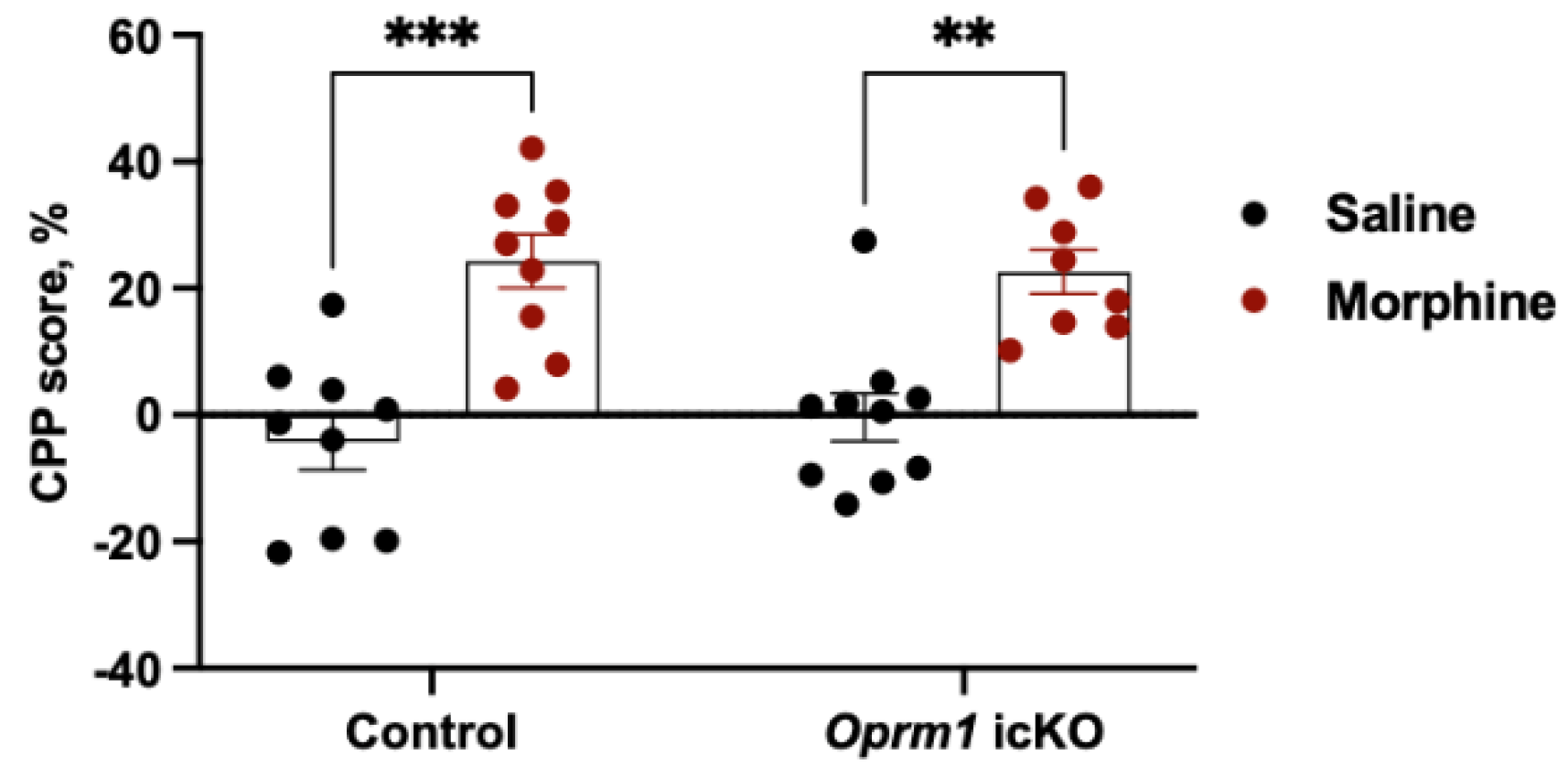
3.3.2. Opmr1 icKO Mice Exhibit Normal CPP to Morphine
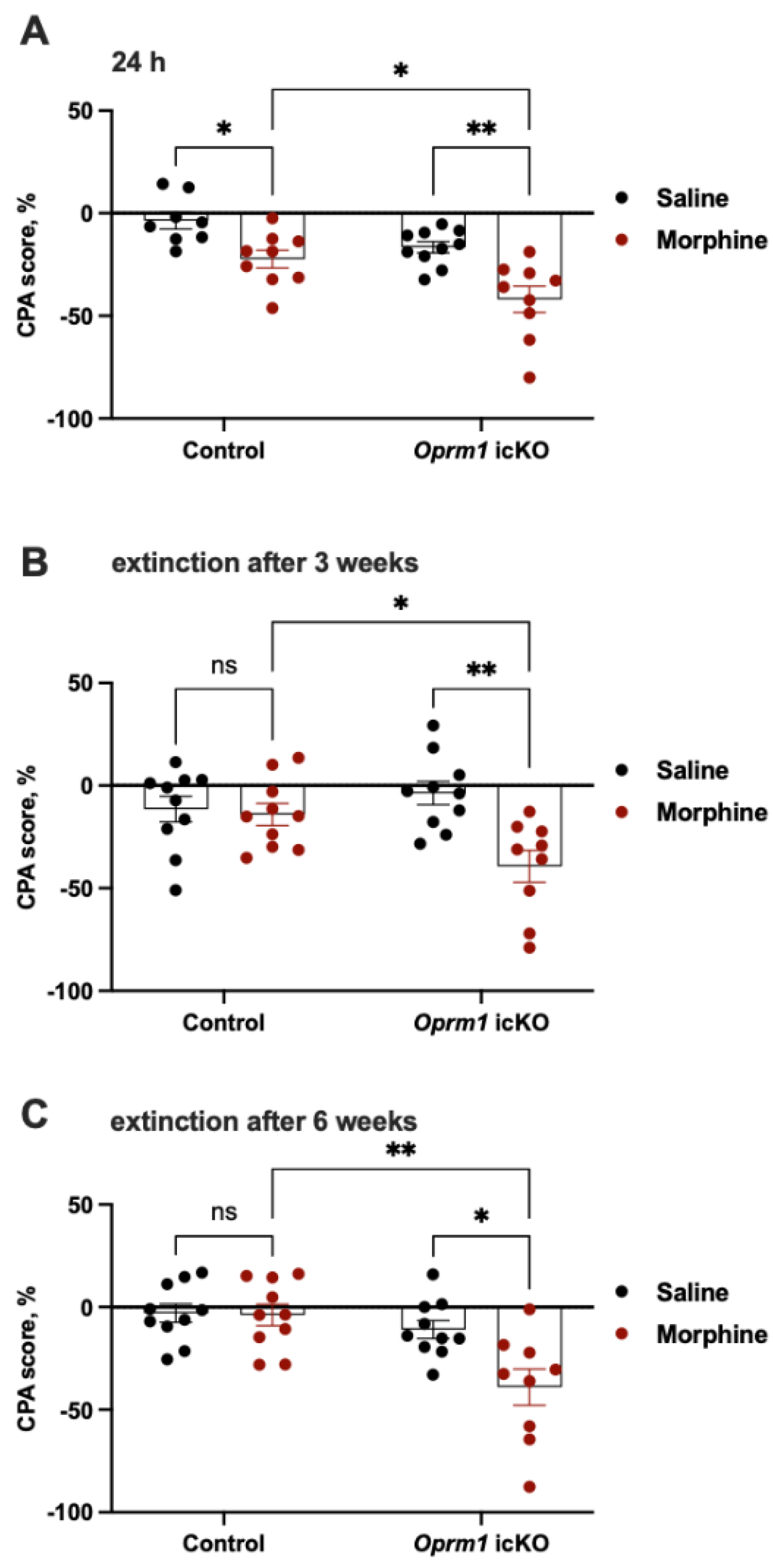
3.3.3. Oprm1 icKO Mice Exhibit Stronger CPA to Morphine Withdrawal
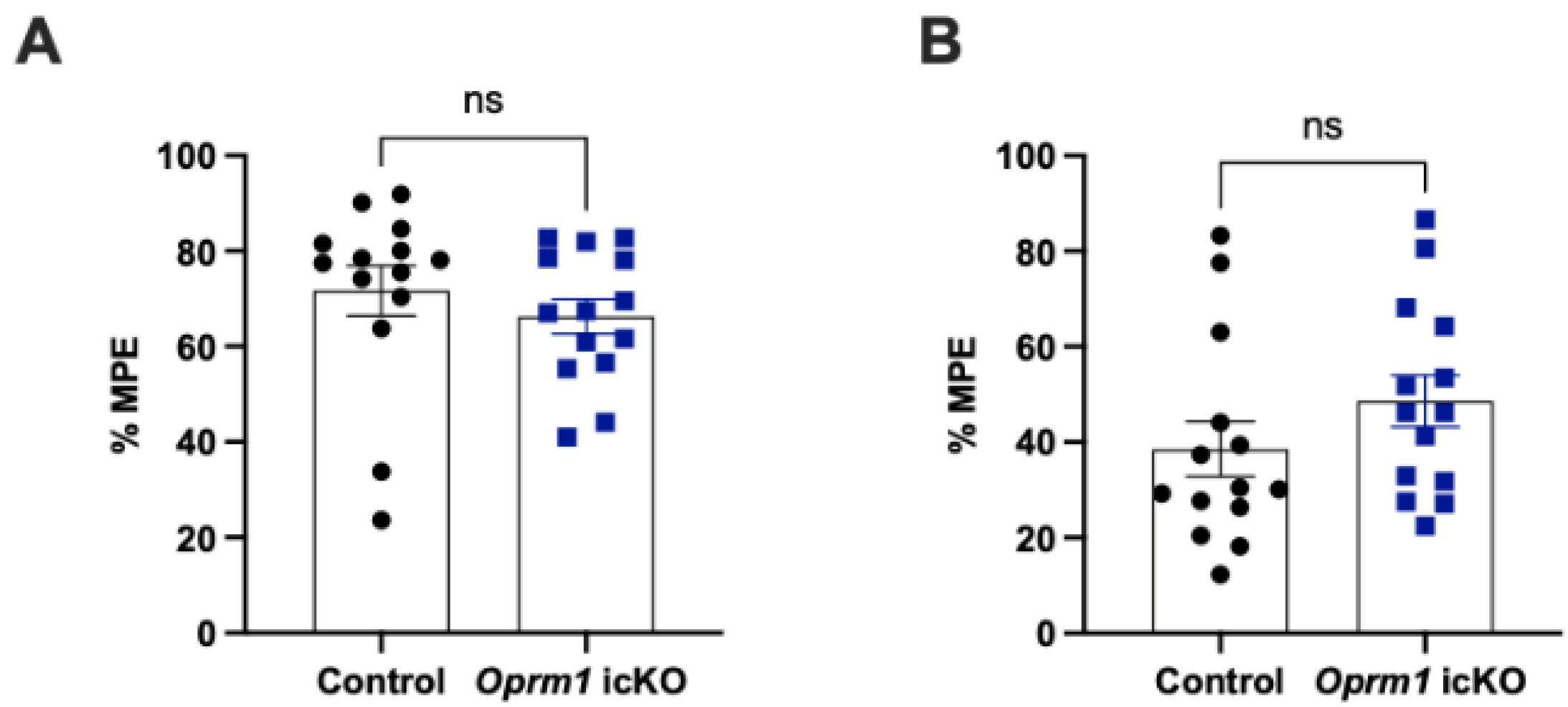
3.4. Oprm1 icKO Does Not Affect the Acute Analgesic Effects of Morphine
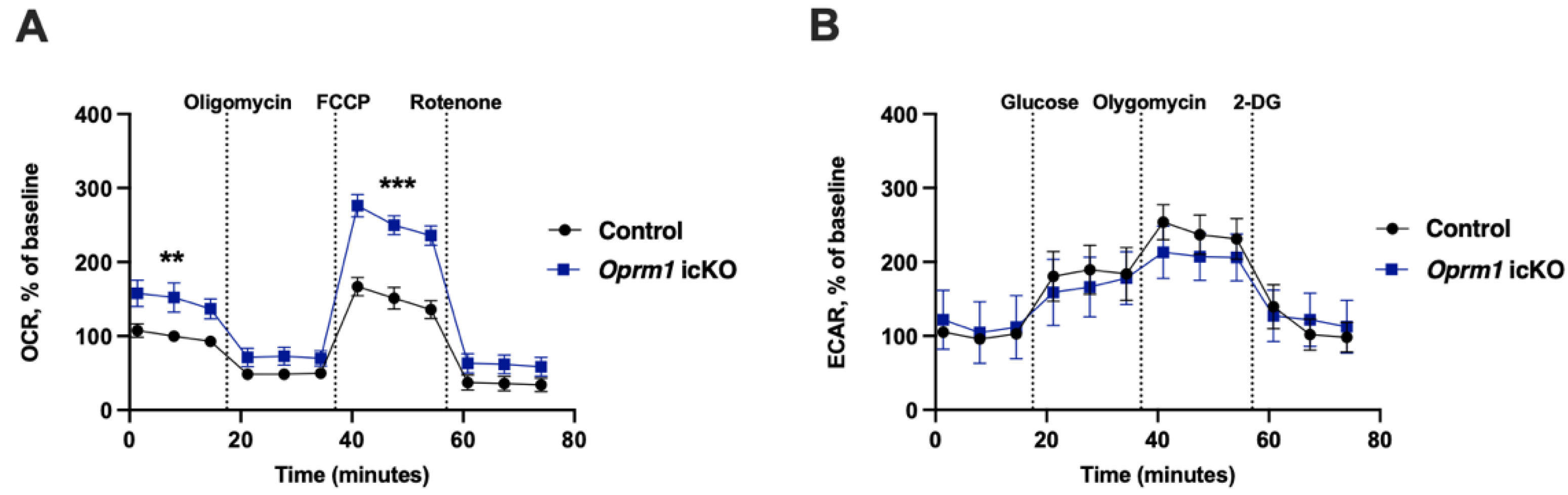
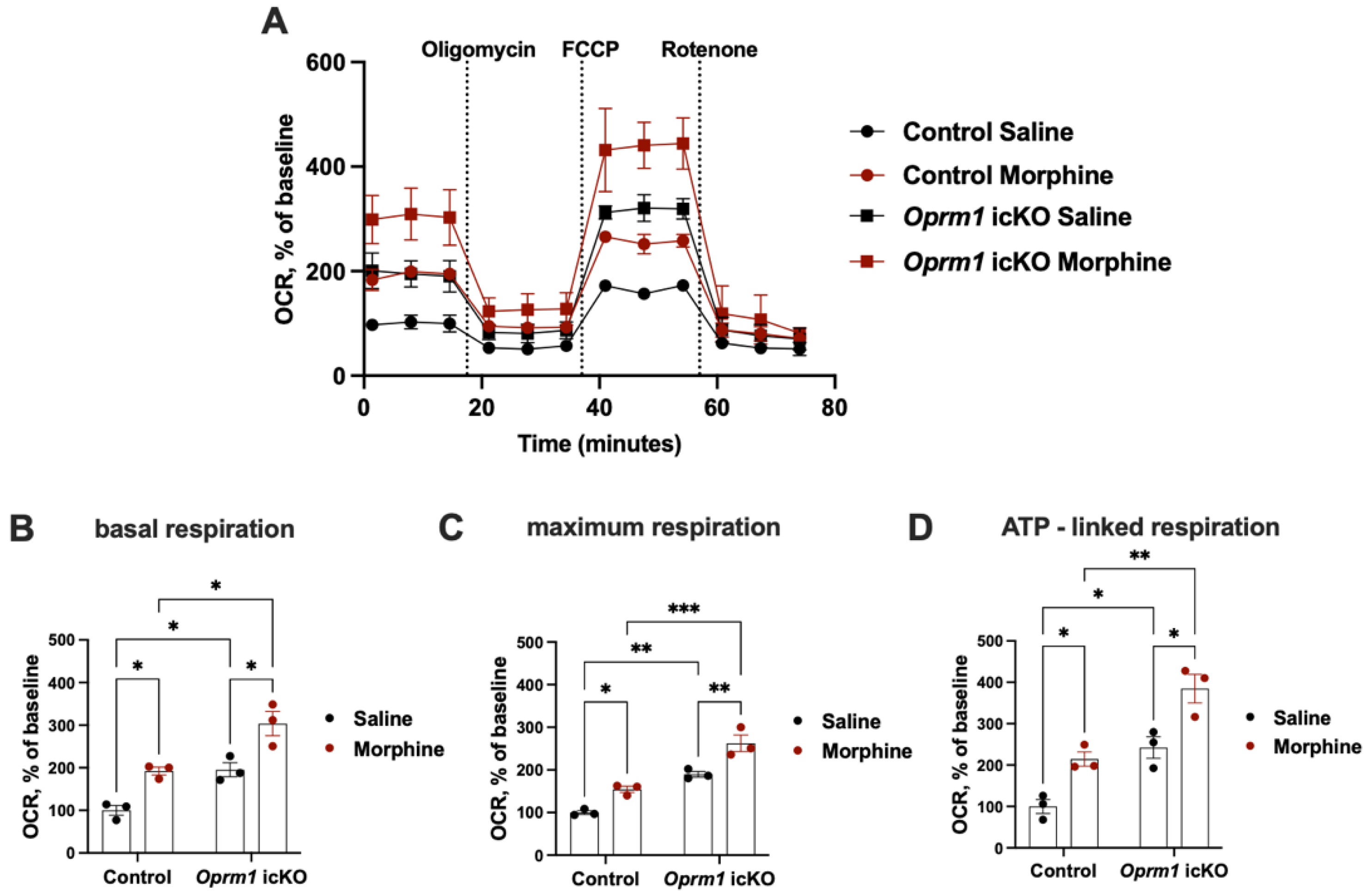
3.5. Increased OXPHOS in Oprm1-Deficient Astrocytes
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kruyer, A.; Scofield, M.D.; Wood, D.; Reissner, K.J.; Kalivas, P.W. Heroin Cue-Evoked Astrocytic Structural Plasticity at Nucleus Accumbens Synapses Inhibits Heroin Seeking. Biol. Psychiatry 2019, 86, 811–819. [Google Scholar] [CrossRef]
- Avey, D.; Sankararaman, S.; Yim, A.K.Y.; Barve, R.; Milbrandt, J.; Mitra, R.D. Single-Cell RNA-Seq Uncovers a Robust Transcriptional Response to Morphine by Glia. Cell Rep. 2018, 24, 3619–3629.e3614. [Google Scholar] [CrossRef]
- Dozio, V.; Daali, Y.; Desmeules, J.; Sanchez, J.C. Deep proteomics and phosphoproteomics reveal novel biological pathways perturbed by morphine, morphine-3-glucuronide and morphine-6-glucuronide in human astrocytes. J. Neurosci. Res. 2022, 100, 220–236. [Google Scholar] [CrossRef] [PubMed]
- Liao, K.; Niu, F.; Hu, G.; Yang, L.; Dallon, B.; Villarreal, D.; Buch, S. Morphine-mediated release of miR-138 in astrocyte-derived extracellular vesicles promotes microglial activation. J. Extracell. Vesicles 2020, 10, e12027. [Google Scholar] [CrossRef]
- Bachtell, R.K.; Jones, J.D.; Heinzerling, K.G.; Beardsley, P.M.; Comer, S.D. Glial and neuroinflammatory targets for treating substance use disorders. Drug Alcohol Depend. 2017, 180, 156–170. [Google Scholar] [CrossRef] [PubMed]
- Rönnbäck, L.; Hansson, E. Are astroglial cells involved in morphine tolerance? Neurochem. Res. 1988, 13, 87–103. [Google Scholar] [CrossRef] [PubMed]
- Nam, M.H.; Han, K.S.; Lee, J.; Won, W.; Koh, W.; Bae, J.Y.; Woo, J.; Kim, J.; Kwong, E.; Choi, T.Y.; et al. Activation of Astrocytic μ-Opioid Receptor Causes Conditioned Place Preference. Cell Rep. 2019, 28, 1154–1166.e1155. [Google Scholar] [CrossRef]
- Kruyer, A.; Dixon, D.; Angelis, A.; Amato, D.; Kalivas, P.W. Astrocytes in the ventral pallidum extinguish heroin seeking through GAT-3 upregulation and morphological plasticity at D1-MSN terminals. Mol. Psychiatry 2022, 27, 855–864. [Google Scholar] [CrossRef]
- Lv, Y.; Jing, M.Y.; Li, P.Y.; Zhao, T.Y.; Pang, C.; Lu, G.Y.; Wang, Z.Y.; Wu, N.; Hu, G.; Song, R.; et al. Aquaporin-4 deletion attenuates opioid-induced addictive behaviours associated with dopamine levels in nucleus accumbens. Neuropharmacology 2022, 208, 108986. [Google Scholar] [CrossRef]
- Semyanov, A.; Verkhratsky, A. Astrocytic processes: From tripartite synapses to the active milieu. Trends Neurosci. 2021, 44, 781–792. [Google Scholar] [CrossRef]
- Murlanova, K.; Jouroukhin, Y.; Huseynov, S.; Pletnikova, O.; Morales, M.J.; Guan, Y.; Baraban, J.M.; Bergles, D.E.; Pletnikov, M.V. Deficient mitochondrial respiration in astrocytes impairs trace fear conditioning and increases naloxone-precipitated aversion in morphine-dependent mice. Glia 2022, 70, 1289–1300. [Google Scholar] [CrossRef]
- Agarwal, A.; Wu, P.H.; Hughes, E.G.; Fukaya, M.; Tischfield, M.A.; Langseth, A.J.; Wirtz, D.; Bergles, D.E. Transient Opening of the Mitochondrial Permeability Transition Pore Induces Microdomain Calcium Transients in Astrocyte Processes. Neuron 2017, 93, 587–605.e587. [Google Scholar] [CrossRef]
- Rose, J.; Brian, C.; Pappa, A.; Panayiotidis, M.I.; Franco, R. Mitochondrial Metabolism in Astrocytes Regulates Brain Bioenergetics, Neurotransmission and Redox Balance. Front. Neurosci. 2020, 14, 536682. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Wang, C.; Ren, J.; Guo, X.; Yun, K. Morphine Protects Spinal Cord Astrocytes from Glutamate-Induced Apoptosis via Reducing Endoplasmic Reticulum Stress. Int. J. Mol. Sci. 2016, 17, 1523. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Kim, M.S.; Park, C.; Jung, E.B.; Choi, D.H.; Kim, T.Y.; Moon, S.K.; Park, R. Morphine prevents glutamate-induced death of primary rat neonatal astrocytes through modulation of intracellular redox. Immunopharmacol. Immunotoxicol. 2004, 26, 17–28. [Google Scholar] [CrossRef] [PubMed]
- Ravindranathan, A.; Joslyn, G.; Robertson, M.; Schuckit, M.A.; Whistler, J.L.; White, R.L. Functional characterization of human variants of the mu-opioid receptor gene. Proc. Natl. Acad. Sci. USA 2009, 106, 10811–10816. [Google Scholar] [CrossRef]
- Srinivasan, R.; Lu, T.Y.; Chai, H.; Xu, J.; Huang, B.S.; Golshani, P.; Coppola, G.; Khakh, B.S. New Transgenic Mouse Lines for Selectively Targeting Astrocytes and Studying Calcium Signals in Astrocyte Processes In Situ and In Vivo. Neuron 2016, 92, 1181–1195. [Google Scholar] [CrossRef]
- Weibel, R.; Reiss, D.; Karchewski, L.; Gardon, O.; Matifas, A.; Filliol, D.; Becker, J.A.; Wood, J.N.; Kieffer, B.L.; Gaveriaux-Ruff, C. Mu opioid receptors on primary afferent nav1.8 neurons contribute to opiate-induced analgesia: Insight from conditional knockout mice. PLoS ONE 2013, 8, e74706. [Google Scholar] [CrossRef]
- Bohn, L.M.; Gainetdinov, R.R.; Sotnikova, T.D.; Medvedev, I.O.; Lefkowitz, R.J.; Dykstra, L.A.; Caron, M.G. Enhanced rewarding properties of morphine, but not cocaine, in beta(arrestin)-2 knock-out mice. J. Neurosci. 2003, 23, 10265–10273. [Google Scholar] [CrossRef]
- Gómez-Milanés, I.; Almela, P.; García-Carmona, J.A.; García-Gutiérrez, M.S.; Aracil-Fernández, A.; Manzanares, J.; Milanés Maquilón, M.V.; Laorden, M.L. Accumbal dopamine, noradrenaline and serotonin activity after naloxone-conditioned place aversion in morphine-dependent mice. Neurochem. Int. 2012, 61, 433–440. [Google Scholar] [CrossRef]
- Song, X.; Li, W.; Shi, Y.; Zhang, J.; Li, Y. Expression of protein kinase A and the kappa opioid receptor in selected brain regions and conditioned place aversion in morphine-dependent rats. Oncotarget 2017, 8, 82632–82642. [Google Scholar] [CrossRef]
- Selvaraj, S.; Mezzano, V.; Loomis, C.; Hanniford, D. RNAscope® and BaseScope™: In-situ RNA analysis for formalin-fixed paraffin-embedded tissues and beyond. J. Biomol. Tech. 2020, 31, S31. [Google Scholar]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef] [PubMed]
- Faul, F.; Erdfelder, E.; Lang, A.G.; Buchner, A. G*Power 3: A flexible statistical power analysis program for the social, behavioral, and biomedical sciences. Behav. Res. Methods 2007, 39, 175–191. [Google Scholar] [CrossRef] [PubMed]
- Nam, M.H.; Han, K.S.; Lee, J.; Bae, J.Y.; An, H.; Park, S.; Oh, S.J.; Kim, E.; Hwang, E.; Bae, Y.C.; et al. Expression of µ-Opioid Receptor in CA1 Hippocampal Astrocytes. Exp. Neurobiol. 2018, 27, 120–128. [Google Scholar] [CrossRef]
- Corkrum, M.; Rothwell, P.E.; Thomas, M.J.; Kofuji, P.; Araque, A. Opioid-Mediated Astrocyte-Neuron Signaling in the Nucleus Accumbens. Cells 2019, 8, 586. [Google Scholar] [CrossRef] [PubMed]
- Winchenbach, J.; Düking, T.; Berghoff, S.A.; Stumpf, S.K.; Hülsmann, S.; Nave, K.A.; Saher, G. Inducible targeting of CNS astrocytes in Aldh1l1-CreERT2 BAC transgenic mice. F1000Research 2016, 5, 2934. [Google Scholar] [CrossRef]
- Le Merrer, J.; Becker, J.A.; Befort, K.; Kieffer, B.L. Reward processing by the opioid system in the brain. Physiol. Rev. 2009, 89, 1379–1412. [Google Scholar] [CrossRef]
- Buenaventura, R.G.; Harvey, A.C.; Burns, M.P.; Main, B.S. Sequential Isolation of Microglia and Astrocytes from Young and Aged Adult Mouse Brains for Downstream Transcriptomic Analysis. Methods Protoc. 2022, 5, 77. [Google Scholar] [CrossRef] [PubMed]
- Holt, L.M.; Olsen, M.L. Novel Applications of Magnetic Cell Sorting to Analyze Cell-Type Specific Gene and Protein Expression in the Central Nervous System. PLoS ONE 2016, 11, e0150290. [Google Scholar] [CrossRef]
- Brase, D.A.; Loh, H.H.; Way, E.L. Comparison of the effects of morphine on locomotor activity, analgesia and primary and protracted physical dependence in six mouse strains. J. Pharmacol. Exp. Ther. 1977, 201, 368–374. [Google Scholar] [PubMed]
- Robinson, T.E.; Berridge, K.C. The psychology and neurobiology of addiction: An incentive-sensitization view. Addiction 2000, 95 (Suppl. S2), S91–S117. [Google Scholar] [CrossRef] [PubMed]
- De Vries, T.J.; Shippenberg, T.S. Neural systems underlying opiate addiction. J. Neurosci. 2002, 22, 3321–3325. [Google Scholar] [CrossRef]
- Supplie, L.M.; Düking, T.; Campbell, G.; Diaz, F.; Moraes, C.T.; Götz, M.; Hamprecht, B.; Boretius, S.; Mahad, D.; Nave, K.A. Respiration-Deficient Astrocytes Survive as Glycolytic Cells In Vivo. J. Neurosci. 2017, 37, 4231–4242. [Google Scholar] [CrossRef]
- Kruyer, A.; Scofield, M.D. Astrocytes in Addictive Disorders. Adv. Neurobiol. 2021, 26, 231–254. [Google Scholar] [CrossRef]
- Valentino, R.J.; Volkow, N.D. Untangling the complexity of opioid receptor function. Neuropsychopharmacology 2018, 43, 2514–2520. [Google Scholar] [CrossRef]
- Darcq, E.; Kieffer, B.L. Opioid receptors: Drivers to addiction? Nat. Rev. Neurosci. 2018, 19, 499–514. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Zeng, F.; Hu, Y.; Li, X. A Mini-Review of the Role of Glutamate Transporter in Drug Addiction. Front. Neurol. 2019, 10, 1123. [Google Scholar] [CrossRef] [PubMed]
- Roberts-Wolfe, D.J.; Kalivas, P.W. Glutamate Transporter GLT-1 as a Therapeutic Target for Substance Use Disorders. CNS Neurol. Disord. Drug. Targets 2015, 14, 745–756. [Google Scholar] [CrossRef] [PubMed]
- Shevelkin, A.V.; Terrillion, C.E.; Hasegawa, Y.; Mychko, O.A.; Jouroukhin, Y.; Sawa, A.; Kamiya, A.; Pletnikov, M.V. Astrocyte DISC1 contributes to cognitive function in a brain region-dependent manner. Hum. Mol. Genet. 2020, 29, 2936–2950. [Google Scholar] [CrossRef]
- Robinson, M.B.; Lee, M.L.; DaSilva, S. Glutamate Transporters and Mitochondria: Signaling, Co-compartmentalization, Functional Coupling, and Future Directions. Neurochem. Res. 2020, 45, 526–540. [Google Scholar] [CrossRef] [PubMed]
- Genda, E.N.; Jackson, J.G.; Sheldon, A.L.; Locke, S.F.; Greco, T.M.; O’Donnell, J.C.; Spruce, L.A.; Xiao, R.; Guo, W.; Putt, M.; et al. Co-compartmentalization of the astroglial glutamate transporter, GLT-1, with glycolytic enzymes and mitochondria. J. Neurosci. 2011, 31, 18275–18288. [Google Scholar] [CrossRef] [PubMed]
- Bauer, D.E.; Jackson, J.G.; Genda, E.N.; Montoya, M.M.; Yudkoff, M.; Robinson, M.B. The glutamate transporter, GLAST, participates in a macromolecular complex that supports glutamate metabolism. Neurochem. Int. 2012, 61, 566–574. [Google Scholar] [CrossRef]
- Rose, E.M.; Koo, J.C.; Antflick, J.E.; Ahmed, S.M.; Angers, S.; Hampson, D.R. Glutamate transporter coupling to Na, K-ATPase. J. Neurosci. 2009, 29, 8143–8155. [Google Scholar] [CrossRef]
- Voloboueva, L.A.; Suh, S.W.; Swanson, R.A.; Giffard, R.G. Inhibition of mitochondrial function in astrocytes: Implications for neuroprotection. J. Neurochem. 2007, 102, 1383–1394. [Google Scholar] [CrossRef] [PubMed]
- Woo, D.H.; Bae, J.Y.; Nam, M.H.; An, H.; Ju, Y.H.; Won, J.; Choi, J.H.; Hwang, E.M.; Han, K.S.; Bae, Y.C.; et al. Activation of Astrocytic μ-opioid Receptor Elicits Fast Glutamate Release Through TREK-1-Containing K2P Channel in Hippocampal Astrocytes. Front. Cell. Neurosci. 2018, 12, 319. [Google Scholar] [CrossRef]
- Saeedi, N.; Darvishmolla, M.; Tavassoli, Z.; Davoudi, S.; Heysieattalab, S.; Hosseinmardi, N.; Janahmadi, M.; Behzadi, G. The role of hippocampal glial glutamate transporter (GLT-1) in morphine-induced behavioral responses. Brain Behav. 2021, 11, e2323. [Google Scholar] [CrossRef]
- O’Sullivan, S.J.; Malahias, E.; Park, J.; Srivastava, A.; Reyes, B.A.S.; Gorky, J.; Vadigepalli, R.; Van Bockstaele, E.J.; Schwaber, J.S. Single-Cell Glia and Neuron Gene Expression in the Central Amygdala in Opioid Withdrawal Suggests Inflammation With Correlated Gut Dysbiosis. Front. Neurosci. 2019, 13, 665. [Google Scholar] [CrossRef]
- Zhang, Z.; Park, J.W.; Ahn, I.S.; Diamante, G.; Sivakumar, N.; Arneson, D.; Yang, X.; van Veen, J.E.; Correa, S.M. Estrogen receptor alpha in the brain mediates tamoxifen-induced changes in physiology in mice. eLife 2021, 10, e63333. [Google Scholar] [CrossRef]
- Huhn, A.S.; Berry, M.S.; Dunn, K.E. Systematic review of sex-based differences in opioid-based effects. Int. Rev. Psychiatry 2018, 30, 107–116. [Google Scholar] [CrossRef]
- Rocks, D.; Cham, H.; Kundakovic, M. Why the estrous cycle matters for neuroscience. Biol. Sex. Differ. 2022, 13, 62. [Google Scholar] [CrossRef]
- Crespo-Castrillo, A.; Arevalo, M.A. Microglial and Astrocytic Function in Physiological and Pathological Conditions: Estrogenic Modulation. Int. J. Mol. Sci. 2020, 21, 3219. [Google Scholar] [CrossRef]
- Martin-Jiménez, C.; Gaitán-Vaca, D.M.; Areiza, N.; Echeverria, V.; Ashraf, G.M.; González, J.; Sahebkar, A.; Garcia-Segura, L.M.; Barreto, G.E. Astrocytes Mediate Protective Actions of Estrogenic Compounds after Traumatic Brain Injury. Neuroendocrinology 2019, 108, 142–160. [Google Scholar] [CrossRef] [PubMed]
- Chisholm, N.C.; Sohrabji, F. Astrocytic response to cerebral ischemia is influenced by sex differences and impaired by aging. Neurobiol. Dis. 2016, 85, 245–253. [Google Scholar] [CrossRef]
- Wang, C.; Jie, C.; Dai, X. Possible roles of astrocytes in estrogen neuroprotection during cerebral ischemia. Rev. Neurosci. 2014, 25, 255–268. [Google Scholar] [CrossRef]
- Kim, R.; Testen, A.; Harder, E.V.; Brown, N.E.; Witt, E.A.; Bellinger, T.J.; Franklin, J.P.; Reissner, K.J. Abstinence-Dependent Effects of Long-Access Cocaine Self-Administration on Nucleus Accumbens Astrocytes Are Observed in Male, But Not Female, Rats. eNeuro 2022, 9, 1–14. [Google Scholar] [CrossRef]
- Hadschieff, V.; Drdla-Schutting, R.; Springer, D.N.; Siegert, A.S.M.; Schroeder, H.; Sandkühler, J. Fundamental sex differences in morphine withdrawal-induced neuronal plasticity. Pain 2020, 161, 2022–2034. [Google Scholar] [CrossRef]
- Serdarevic, M.; Striley, C.W.; Cottler, L.B. Sex differences in prescription opioid use. Curr. Opin. Psychiatry 2017, 30, 238–246. [Google Scholar] [CrossRef] [PubMed]
- Foo, L.C.; Dougherty, J.D. Aldh1L1 is expressed by postnatal neural stem cells in vivo. Glia 2013, 61, 1533–1541. [Google Scholar] [CrossRef] [PubMed]
- Beyer, F.; Lüdje, W.; Karpf, J.; Saher, G.; Beckervordersandforth, R. Distribution of Aldh1L1-CreER(T2) Recombination in Astrocytes Versus Neural Stem Cells in the Neurogenic Niches of the Adult Mouse Brain. Front. Neurosci. 2021, 15, 713077. [Google Scholar] [CrossRef]
- Machelska, H.; Celik, M. Opioid Receptors in Immune and Glial Cells-Implications for Pain Control. Front. Immunol. 2020, 11, 300. [Google Scholar] [CrossRef] [PubMed]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}

| Cohort | Test | PND |
|---|---|---|
| Cohort 1 | Open field test | 60 |
| Elevated plus maze | 80 | |
| Novel place recognition test | 90–92 | |
| Cohort 2 | Morphine-induced sensitization | 90–102 |
| Cohort 3 | Conditioned place preference | 60–71 |
| Cohort 4 | Conditioned place aversion | 105–110; 130; 150 |
| Cohort 5 | Hargreaves test | 70 |
| Cohort 6 | Hot plate test | 70 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Murlanova, K.; Jouroukhin, Y.; Novototskaya-Vlasova, K.; Huseynov, S.; Pletnikova, O.; Morales, M.J.; Guan, Y.; Kamiya, A.; Bergles, D.E.; Dietz, D.M.; et al. Loss of Astrocytic µ Opioid Receptors Exacerbates Aversion Associated with Morphine Withdrawal in Mice: Role of Mitochondrial Respiration. Cells 2023, 12, 1412. https://doi.org/10.3390/cells12101412
Murlanova K, Jouroukhin Y, Novototskaya-Vlasova K, Huseynov S, Pletnikova O, Morales MJ, Guan Y, Kamiya A, Bergles DE, Dietz DM, et al. Loss of Astrocytic µ Opioid Receptors Exacerbates Aversion Associated with Morphine Withdrawal in Mice: Role of Mitochondrial Respiration. Cells. 2023; 12(10):1412. https://doi.org/10.3390/cells12101412
Chicago/Turabian StyleMurlanova, Kateryna, Yan Jouroukhin, Ksenia Novototskaya-Vlasova, Shovgi Huseynov, Olga Pletnikova, Michael J. Morales, Yun Guan, Atsushi Kamiya, Dwight E. Bergles, David M. Dietz, and et al. 2023. "Loss of Astrocytic µ Opioid Receptors Exacerbates Aversion Associated with Morphine Withdrawal in Mice: Role of Mitochondrial Respiration" Cells 12, no. 10: 1412. https://doi.org/10.3390/cells12101412
APA StyleMurlanova, K., Jouroukhin, Y., Novototskaya-Vlasova, K., Huseynov, S., Pletnikova, O., Morales, M. J., Guan, Y., Kamiya, A., Bergles, D. E., Dietz, D. M., & Pletnikov, M. V. (2023). Loss of Astrocytic µ Opioid Receptors Exacerbates Aversion Associated with Morphine Withdrawal in Mice: Role of Mitochondrial Respiration. Cells, 12(10), 1412. https://doi.org/10.3390/cells12101412