
Rhodnius prolixus Hemolymph Immuno-Physiology: Deciphering the Systemic Immune Response Triggered by Trypanosoma cruzi Establishment in the Vector Using Quantitative Proteomics



Abstract
:1. Introduction
2. Materials and Methods
2.1. Samples Preparation
2.2. Hemolymph Preparation
2.3. Sample Preparation Prior to Liquid Chromatography Tandem Mass Spectrometry (LC-MS/MS)
2.4. Mass Spectrometric Data Analysis
2.5. Prediction of Protein Localization
2.6. Functional Characterization and Protein Classification
2.7. Phenoloxidase and Prophenoloxidase Activity
2.8. Western Blotting
3. Results
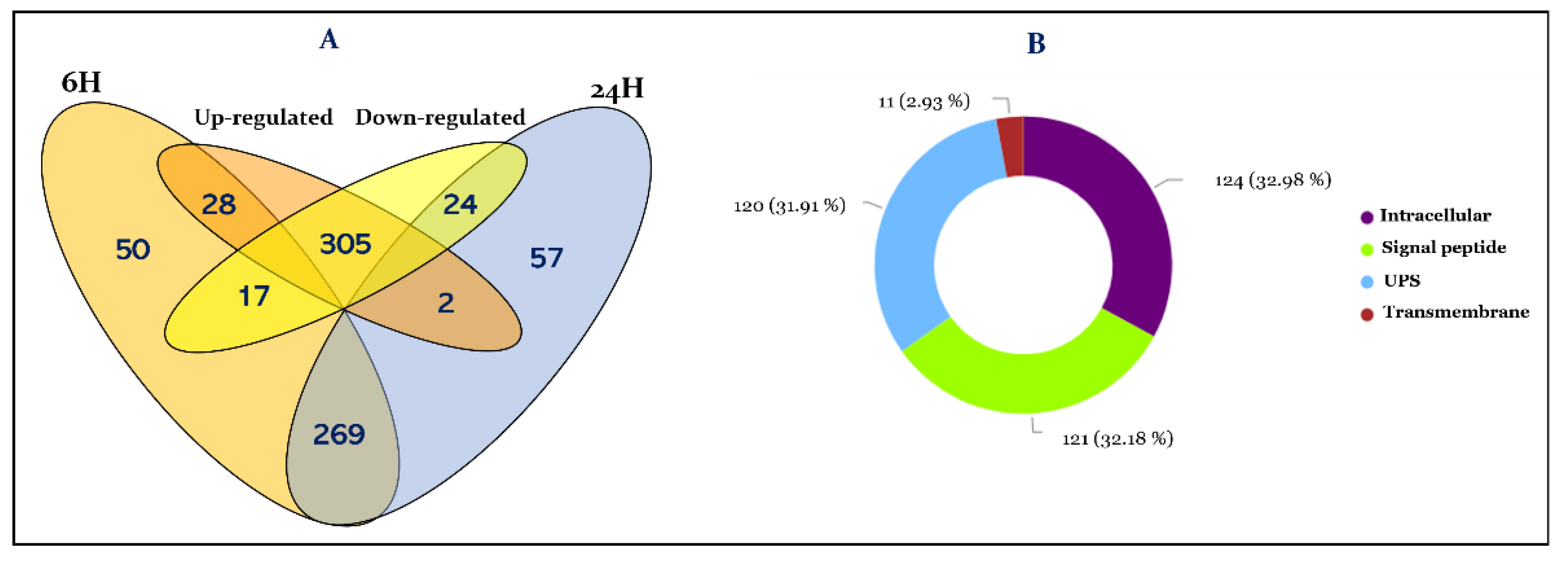
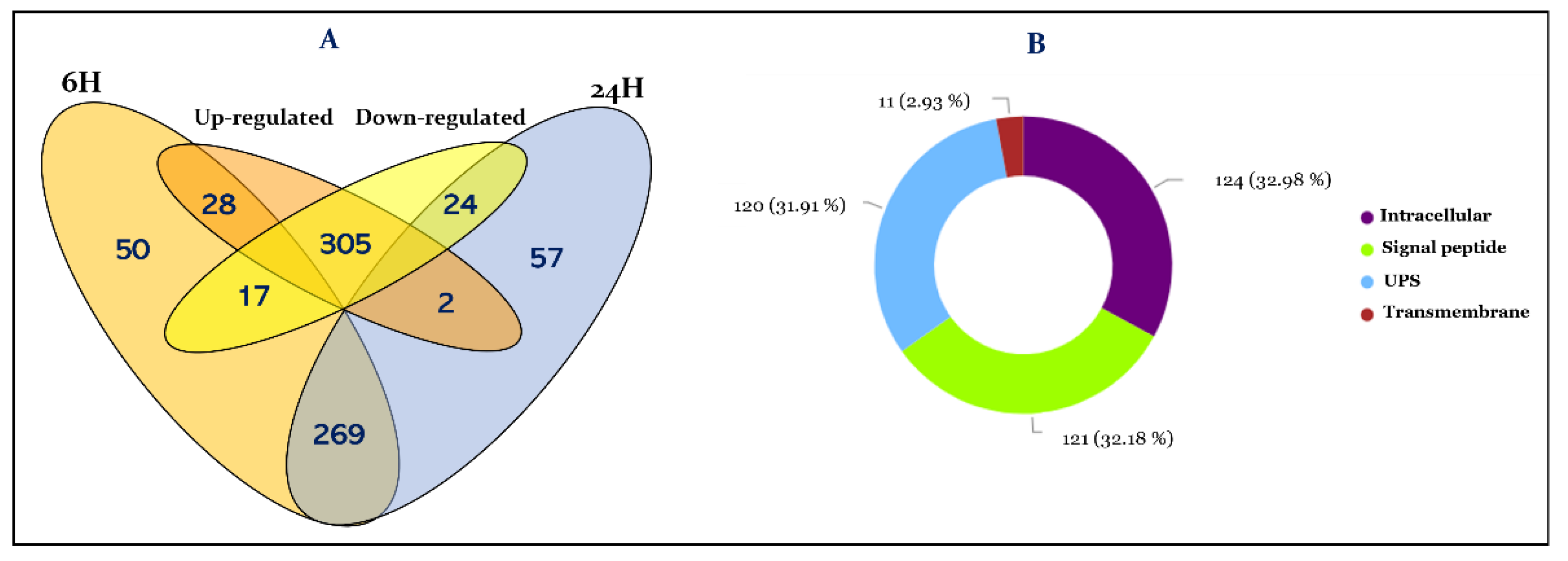
3.1. Overview of R. prolixus Hemolymph Proteome
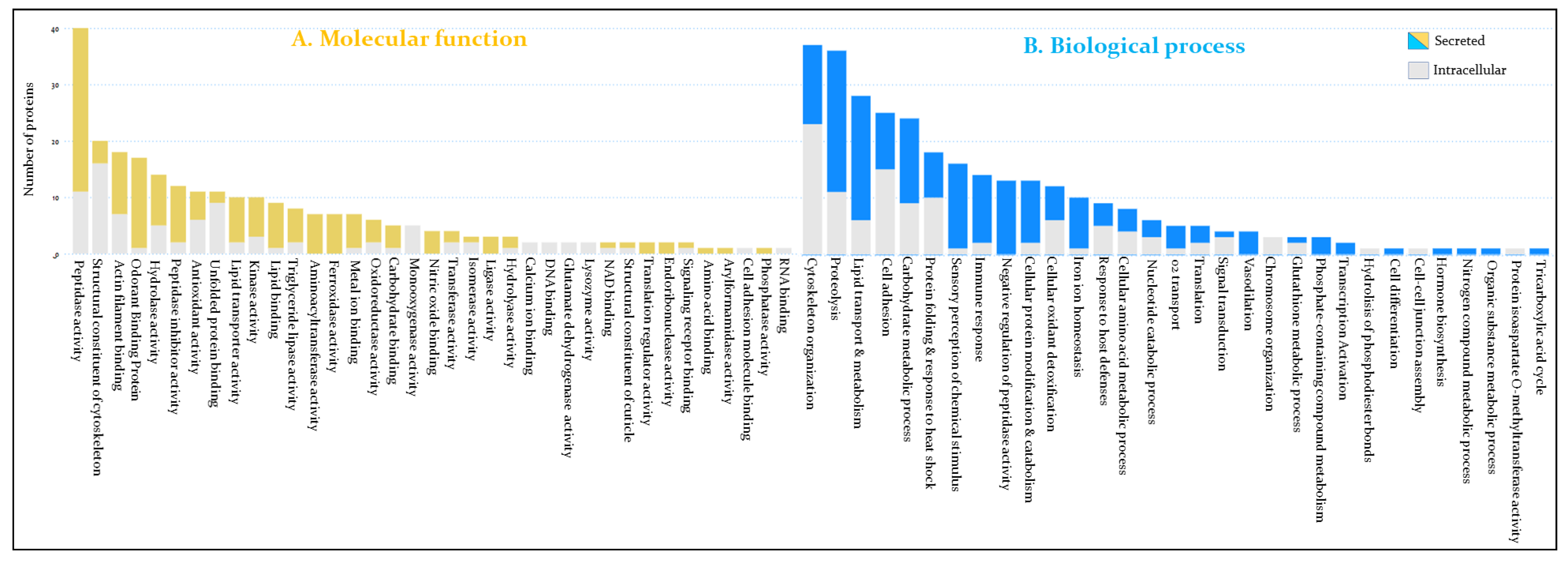
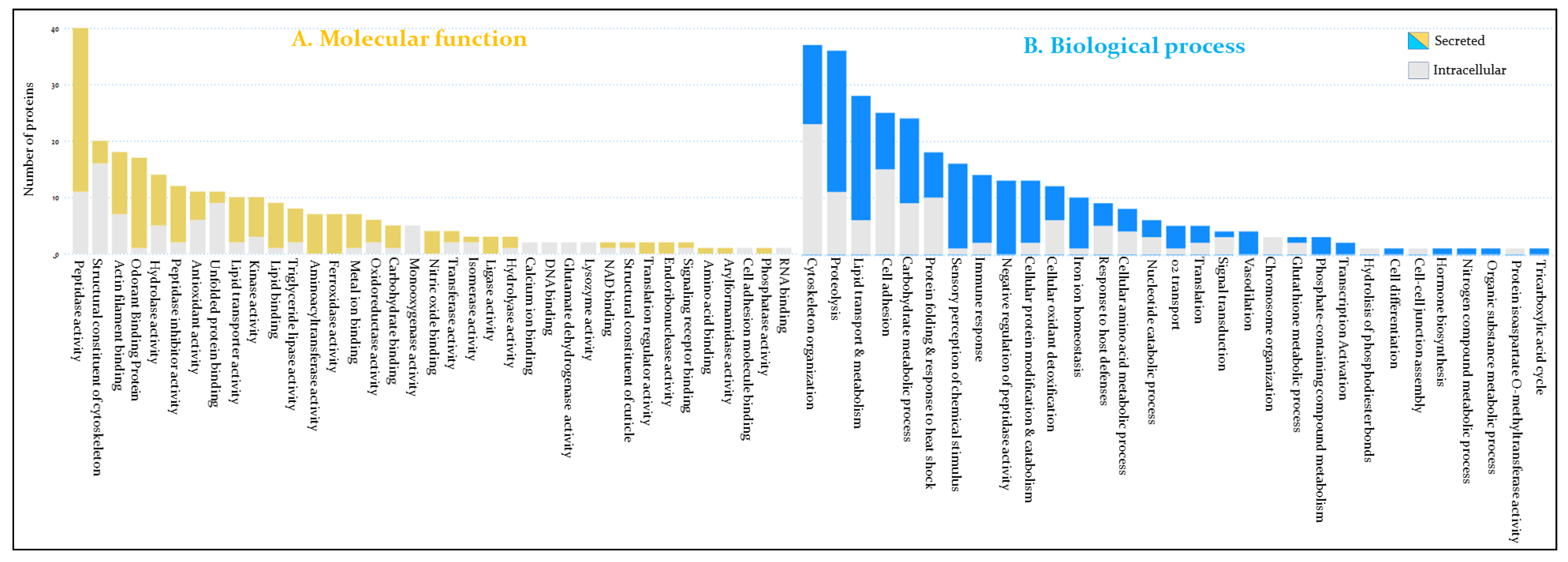
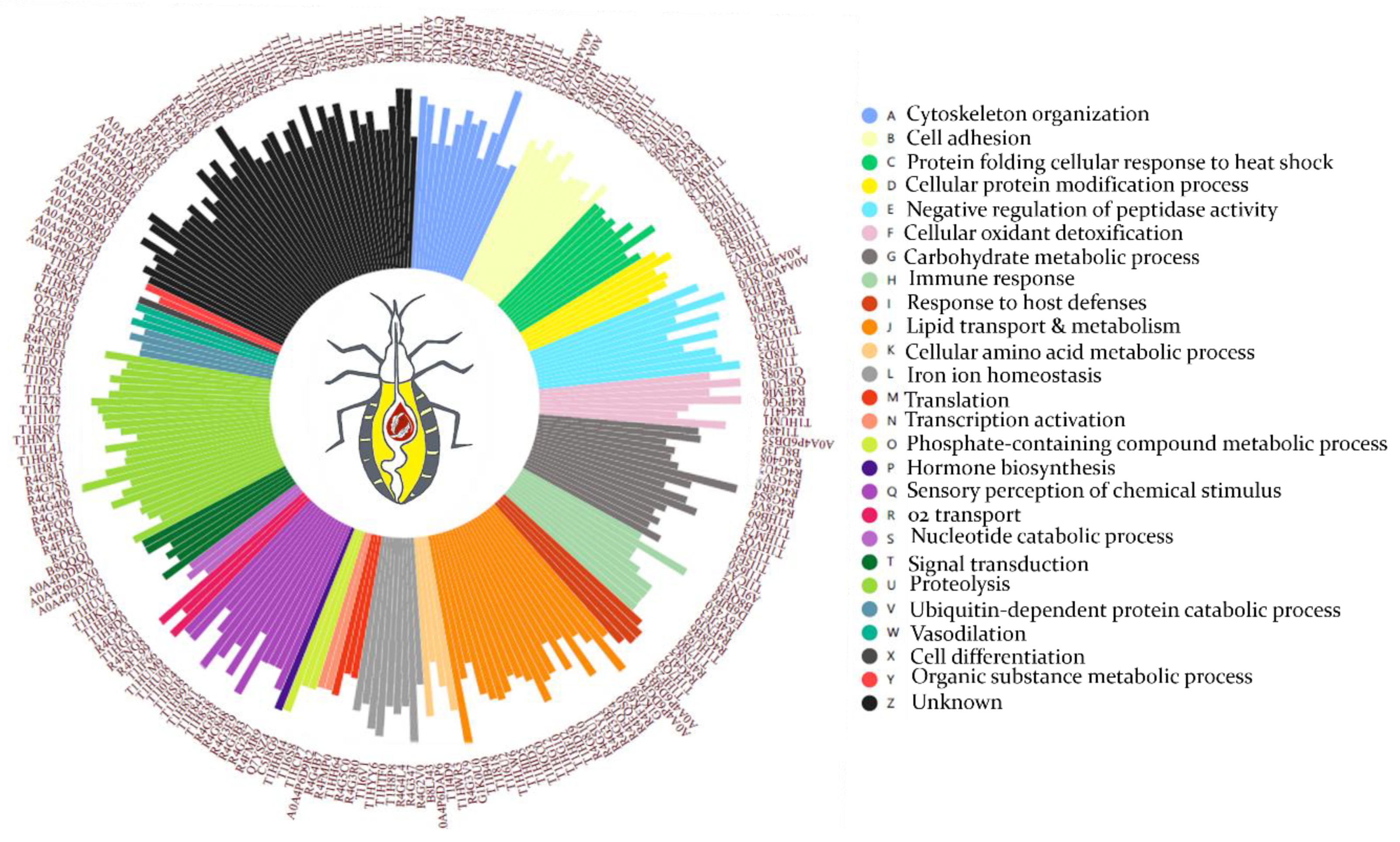
3.2. Functional Annotation of Hemolymph Proteins
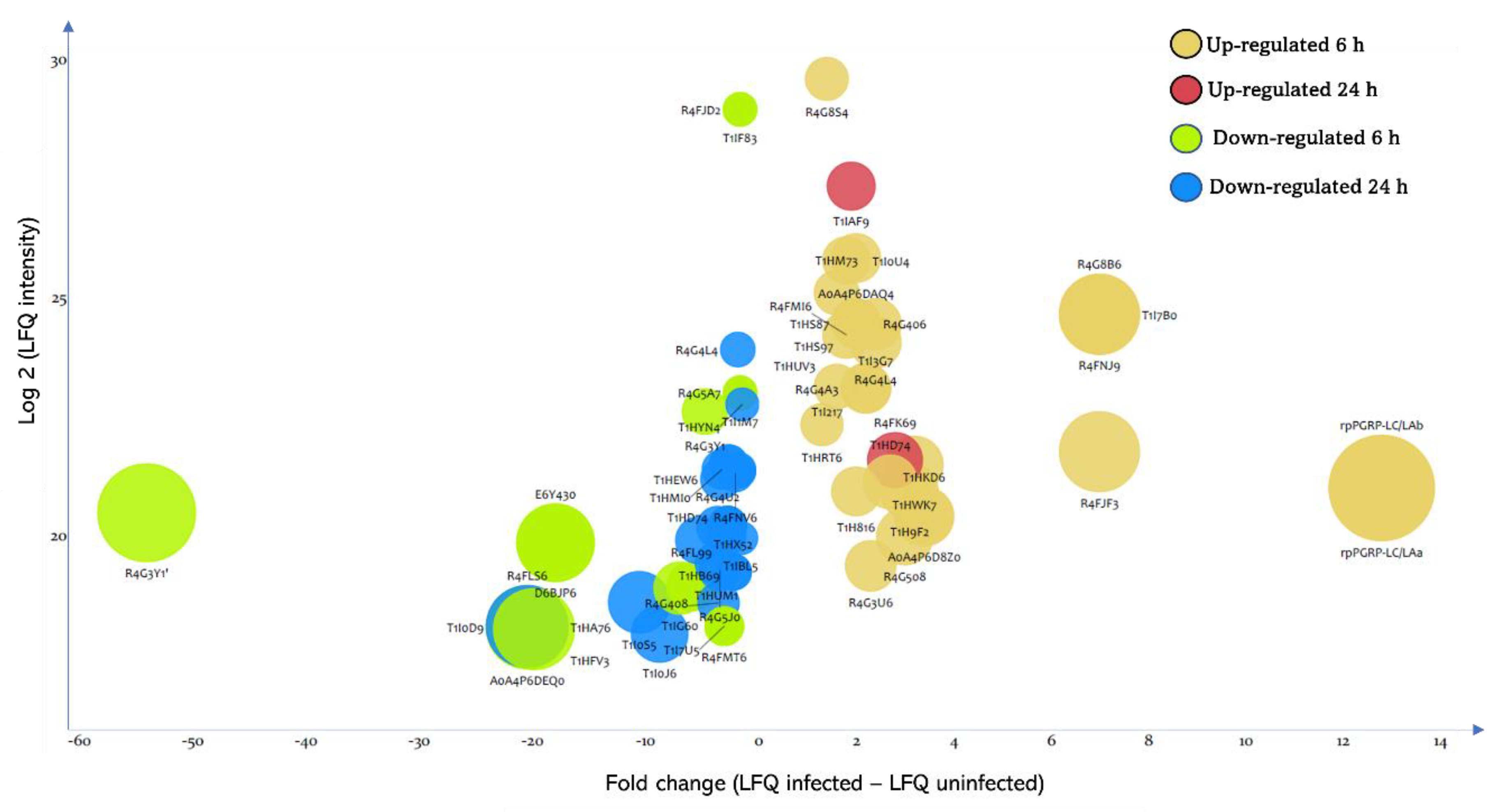
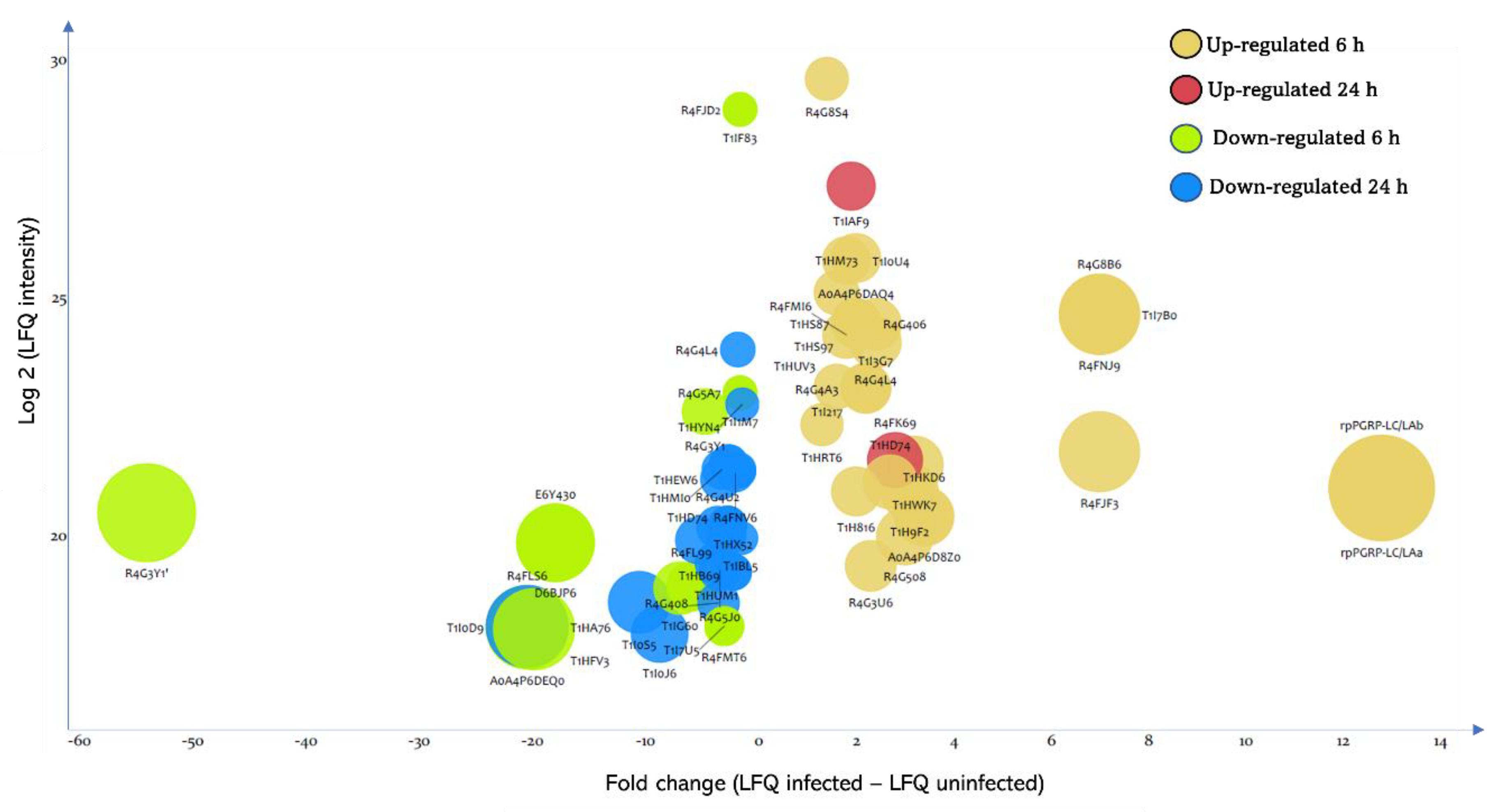
3.3. Effect of T. cruzi on the Dynamic of Hemolymph Protein Expression
4. Discussion
4.1. R. prolixus Hemolymph Proteome Homeostasis under Blood Feeding Condition
4.2. Exploring the Hemolymph Immunoproteins
4.2.1. Nonself Perception and Recognition
4.2.2. Humoral Response through Immune Effectors
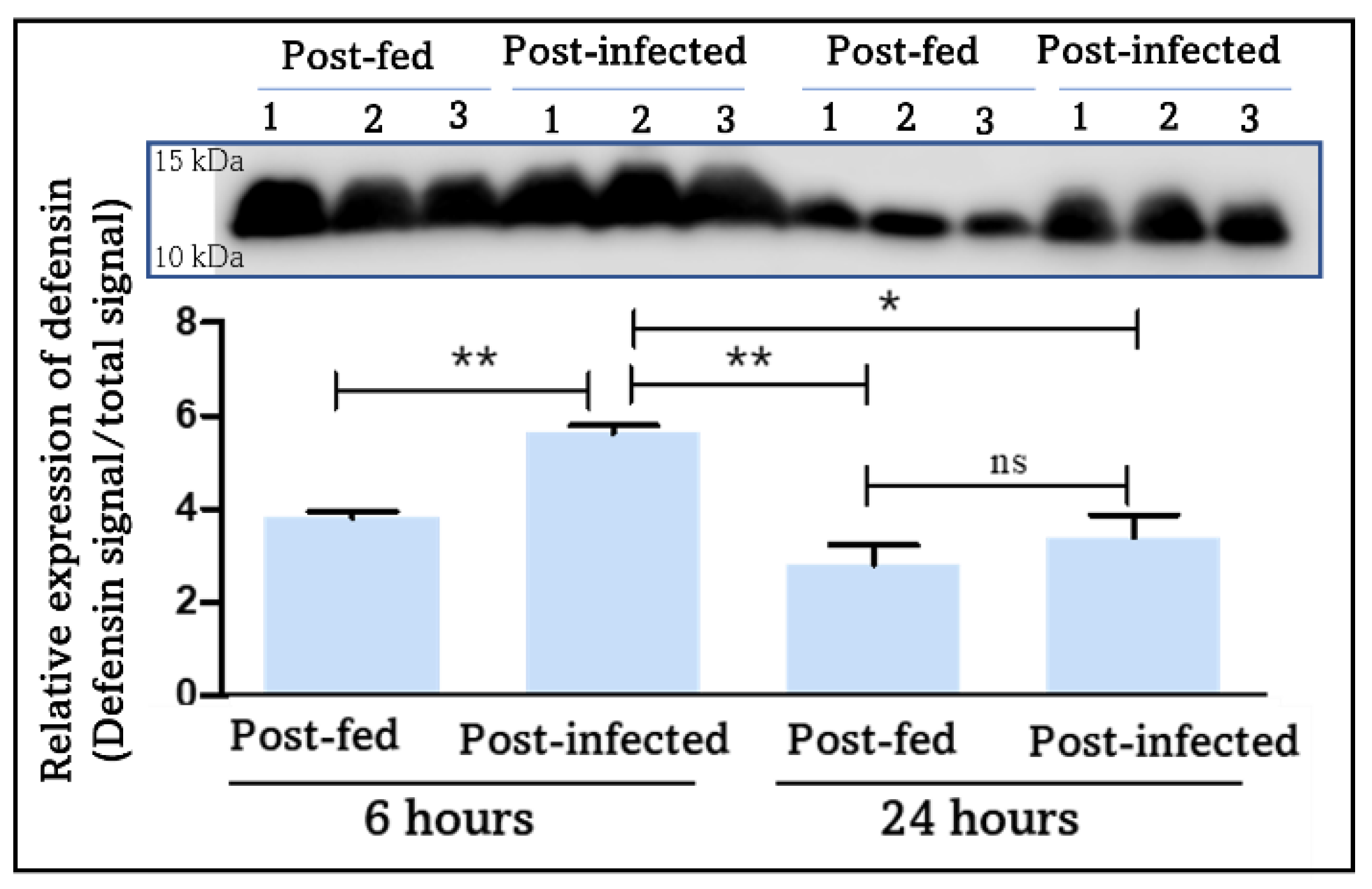
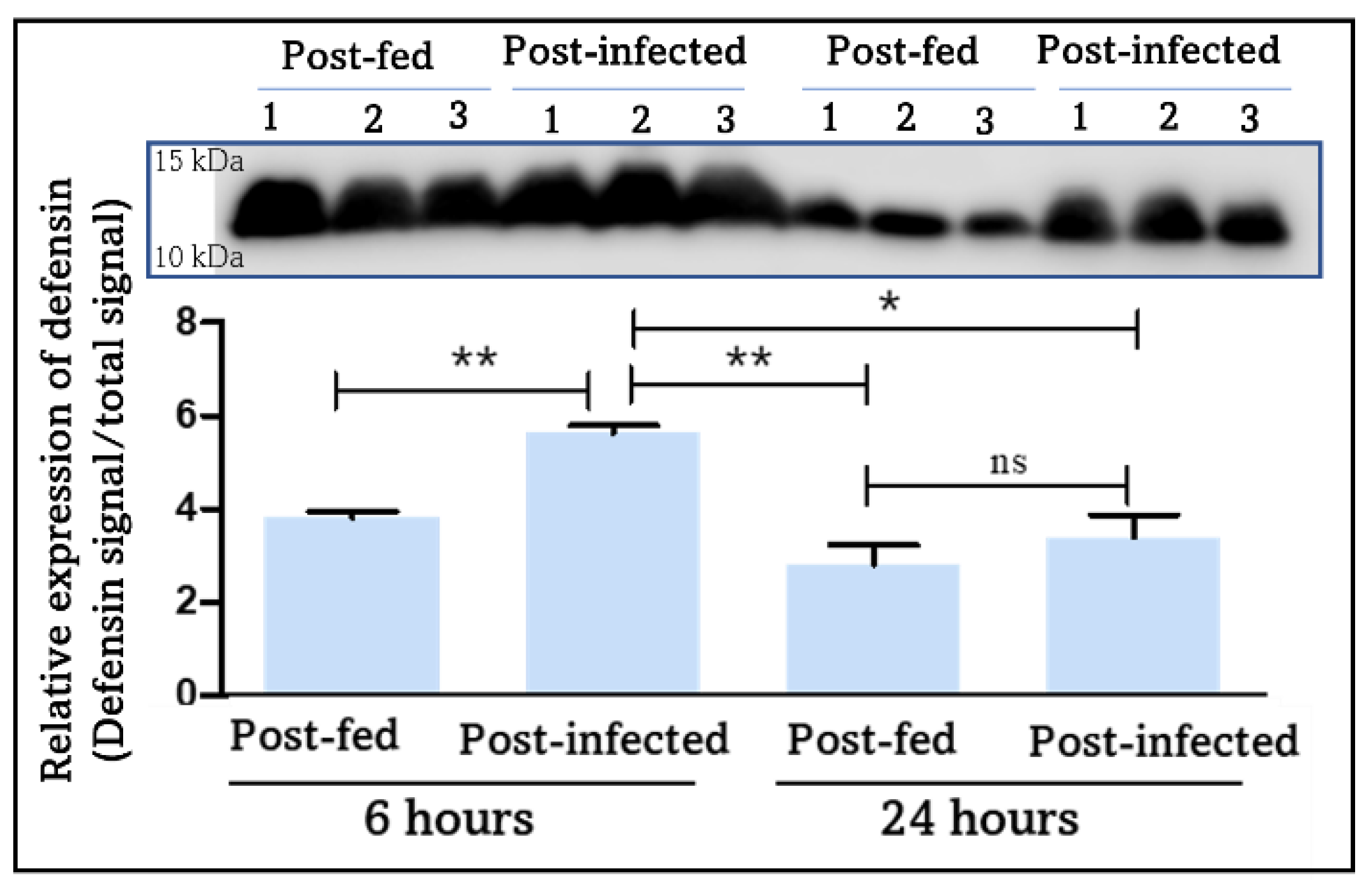
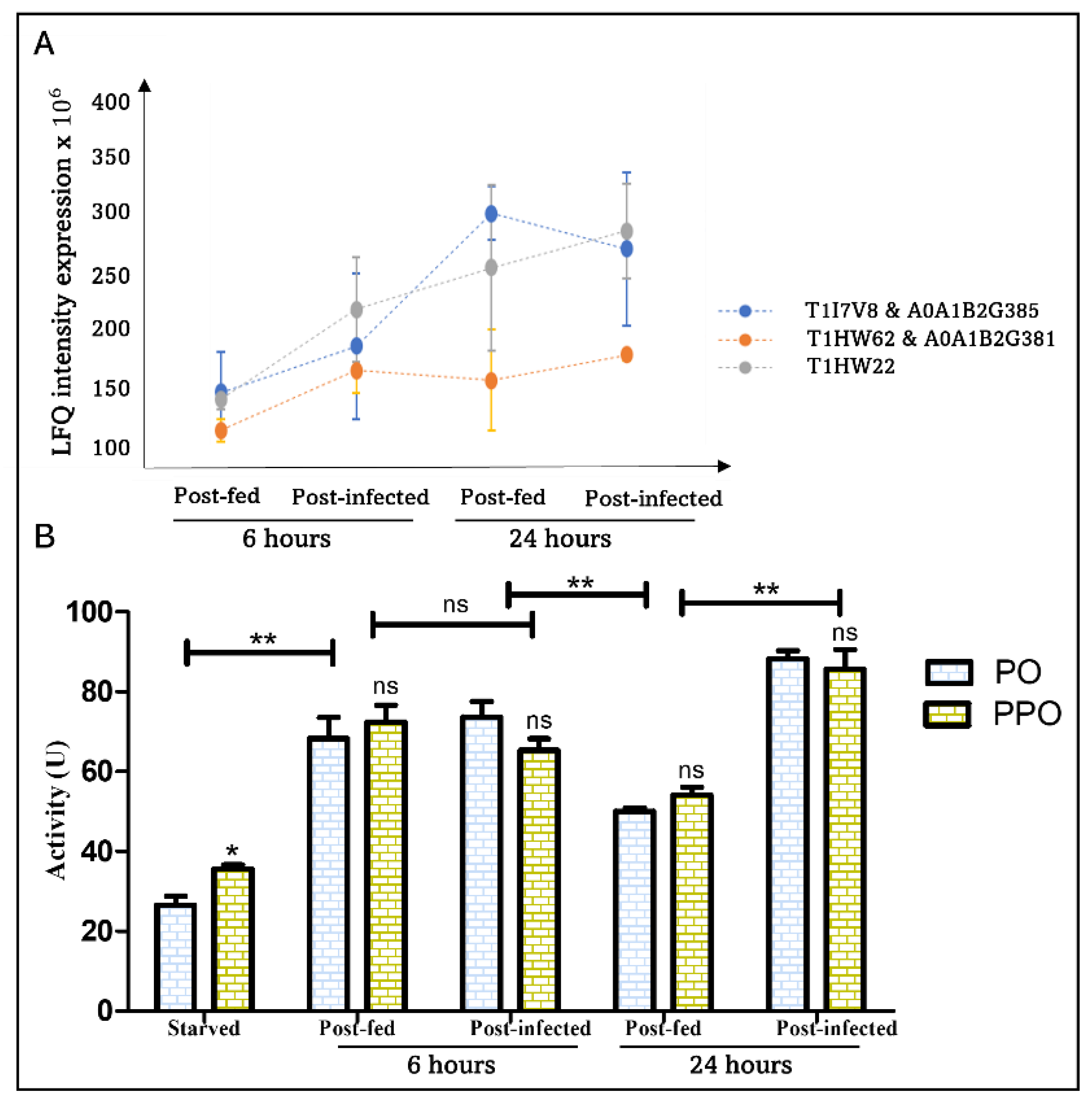
4.2.3. Melanization and Clotting: Blood-Feeding Induces an Early Immune Response Mediated by PO and Extended by T. cruzi
4.2.4. Oxidative Stress Related to Immunity
4.2.5. Pathogen’s Opsonization
4.2.6. T. cruzi Modulates JHBP Expression
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- World Health Organisation (WHO). Chagas Disease (Also Known as American Trypanosomiasis). 2020. Available online: https://www.who.int/news-room/fact-sheets/detail/chagas-disease-(american-trypanosomiasis) (accessed on 15 November 2021).
- Shaw, W.R.; Catteruccia, F. Vector biology meets disease control: Using basic research to fight vector-borne diseases. Nat. Microbiol. 2019, 4, 20–34. [Google Scholar] [CrossRef]
- Baxter, R.H.G.; Contet, A.; Krueger, K. Arthropod Innate Immune Systems and Vector-Borne Diseases. Biochemistry 2017, 56, 907–918. [Google Scholar] [CrossRef] [Green Version]
- Schaub, G.A. An Update on the Knowledge of Parasite–Vector Interactions of Chagas Disease. Res. Rep. Trop. Med. 2021, 12, 63–76. [Google Scholar] [CrossRef]
- Garcia, E.S.; Genta, F.A.; De Azambuja, P.; Schaub, G.A. Interactions between intestinal compounds of triatomines and Trypanosoma cruzi. Trends Parasitol. 2010, 26, 499–505. [Google Scholar] [CrossRef]
- Salcedo-Porras, N.; Lowenberger, C. The innate immune system of kissing bugs, vectors of chagas disease. Dev. Comp. Immunol. 2019, 98, 119–128. [Google Scholar] [CrossRef]
- Garcia, E.S.; Castro, D.P.; Figueiredo, M.B.; Genta, F.A.; Azambuja, P. Trypanosoma rangeli: A new perspective for studying the modulation of immune reactions of Rhodnius prolixus. Parasites Vectors 2009, 2, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Mello, C.B.; Garcia, E.S.; Ratcliffe, N.A.; Azambuja, P. Trypanosoma cruzi and Trypanosoma rangeli: Interplay with hemolymph components of Rhodnius prolixus. J. Invertebr. Pathol. 1995, 65, 261–268. [Google Scholar] [CrossRef]
- Gregório, E.A.; A.Ratcliffe, N. The prophenoloxidase system and in vitro interaction of Trypanosoma rangeli with Rhodnius prolixus and Triatoma infestans haemolymph. Parasite Immunol. 1991, 13, 551–564. [Google Scholar] [CrossRef]
- Garcia, E.S.; Castro, D.P.; Figueiredo, M.B.; Azambuja, P. Parasite-mediated interactions within the insect vector: Trypanosoma rangeli strategies. Parasites Vectors 2012, 5, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Flores-Villegas, A.L.; Salazar-Schettino, P.M.; Córdoba-Aguilar, A.; Gutiérrez-Cabrera, A.E.; Rojas-Wastavino, G.E.; Bucio-Torres, M.I.; Cabrera-Bravo, M. Immune defence mechanisms of triatomines against bacteria, viruses, fungi and parasites. Bull. Entomol. Res. 2015, 105, 523–532. [Google Scholar] [CrossRef]
- Salcedo-Porras, N.; Noor, S.; Cai, C.; Oliveira, P.L.; Lowenberger, C. Rhodnius prolixus uses the peptidoglycan recognition receptor rpPGRP-LC/LA to detect Gram-negative bacteria and activate the IMD pathway. Curr. Res. Insect Sci. 2021, 1, 100006. [Google Scholar] [CrossRef]
- Hillyer, J.F.; Pass, G. The insect circulatory system: Structure, function, and evolution. Annu. Rev. Entomol. 2020, 65, 121–143. [Google Scholar] [CrossRef] [Green Version]
- Kanost, M.R. Hemolymph. Encycl. Insects 2009, 446–449. [Google Scholar] [CrossRef]
- Söderhäll, K.; Cerenius, L.; Jiravanichpaisal, P.; Liu, H.; Söderhäll, I. Invertebrate Inmunity; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2010; ISBN 9781441980588. [Google Scholar]
- Imler, J.L. Overview of Drosophila immunity: A historical perspective. Dev. Comp. Immunol. 2014, 42, 3–15. [Google Scholar] [CrossRef]
- Kumar, A.; Srivastava, P.; Sirisena, P.D.N.N.; Dubey, S.K.; Kumar, R.; Shrinet, J.; Sunil, S. Mosquito innate immunity. Insects 2018, 9, 95. [Google Scholar] [CrossRef] [Green Version]
- Levashina, E.A. Immune responses in Anopheles gambiae. Insect Biochem. Mol. Biol. 2004, 34, 673–678. [Google Scholar] [CrossRef]
- Masson, F.; Rommelaere, S.; Marra, A.; Schüpfer, F.; Lemaitre, B. Dual proteomics of Drosophila melanogaster hemolymph infected with the heritable endosymbiont Spiroplasma poulsonii. PLoS ONE 2021, 16, 1–15. [Google Scholar] [CrossRef]
- Vierstraete, E.; Verleyen, P.; Sas, F.; Van Den Bergh, G.; De Loof, A.; Arckens, L.; Schoofs, L. The instantly released Drosophila immune proteome is infection-specific. Biochem. Biophys. Res. Commun. 2004, 317, 1052–1060. [Google Scholar] [CrossRef]
- Handke, B.; Poernbacher, I.; Goetze, S.; Ahrens, C.H.; Omasits, U.; Marty, F.; Simigdala, N.; Meyer, I.; Wollscheid, B.; Brunner, E.; et al. The Hemolymph Proteome of Fed and Starved Drosophila Larvae. PLoS ONE 2013, 8, e67208. [Google Scholar] [CrossRef] [Green Version]
- Xu, P.Z.; Zhang, M.R.; Gao, L.; Wu, Y.C.; Qian, H.Y.; Li, G.; Xu, A.Y. Comparative proteomic analysis reveals immune competence in hemolymph of bombyx mori pupa parasitized by silkworm maggot exorista sorbillans. Insects 2019, 10, 413. [Google Scholar] [CrossRef] [Green Version]
- Hou, Y.; Zou, Y.; Wang, F.; Gong, J.; Zhong, X.; Xia, Q.; Zhao, P. Comparative analysis of proteome maps of silkworm hemolymph during different developmental stages. Proteome Sci. 2010, 8, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Araujo, R.V.; Maciel, C.; Hartfelder, K.; Capurro, M.L. Effects of Plasmodium gallinaceum on hemolymph physiology of Aedes aegypti during parasite development. J. Insect Physiol. 2011, 57, 265–273. [Google Scholar] [CrossRef] [PubMed]
- Paskewitz, S.M.; Shi, L. The hemolymph proteome of Anopheles gambiae. Insect Biochem. Mol. Biol. 2005, 35, 815–824. [Google Scholar] [CrossRef] [PubMed]
- Hu, H.; Bienefeld, K.; Wegener, J.; Zautke, F.; Hao, Y.; Feng, M.; Han, B.; Fang, Y.; Wubie, A.J.; Li, J. Proteome analysis of the hemolymph, mushroom body, and antenna provides novel insight into honeybee resistance against varroa infestation. J. Proteome Res. 2016, 15, 2841–2854. [Google Scholar] [CrossRef] [PubMed]
- Neuhauser, N.; Michalski, A.; Scheltema, R.A.; Olsen, J.V.; Mann, M. Andromeda: A Peptide Search Engine Integrated into the MaxQuant Environment. J. Proteome Res. 2011, 10, 1794–1805. [Google Scholar] [CrossRef]
- Mi, H.; Muruganujan, A.; Casagrande, J.T.; Thomas, P.D. Large-scale gene function analysis with PANTHER classification system. Nat. Protoc. 2013, 8, 1551–1566. [Google Scholar] [CrossRef]
- Kolberg, L.; Raudvere, U.; Kuzmin, I.; Vilo, J.; Peterson, H.; Mathe, E. Gprofiler2—An R package for gene list functional enrichment analysis and namespace conversion toolset g:Profiler. F1000Research 2020, 9, 1–27. [Google Scholar] [CrossRef]
- Zhao, L.; Poschmann, G.; Waldera-Lupa, D.; Rafiee, N.; Kollmann, M.; Stühler, K. OutCyte: A novel tool for predicting unconventional protein secretion. Sci. Rep. 2019, 9, 1–9. [Google Scholar] [CrossRef]
- Ferreira, R.C.; Kessler, R.L.; Lorenzo, M.G.; Paim, R.M.M.; Ferreira, L.D.L.; Probst, C.M.; Alves-Silva, J.; Guarneri, A.A. Colonization of Rhodnius prolixus gut by Trypanosoma cruzi involves an extensive parasite killing. Parasitology 2016, 143, 434–443. [Google Scholar] [CrossRef]
- Capilla, A.; Karachentsev, D.; Patterson, R.A.; Hermann, A.; Juarez, M.T.; McGinnis, W. Toll pathway is required for wound-induced expression of barrier repair genes in the Drosophila epidermis. Proc. Natl. Acad. Sci. USA 2017, 114, E2682–E2688. [Google Scholar] [CrossRef] [Green Version]
- Leprêtre, M.; Almunia, C.; Armengaud, J.; Salvador, A.; Geffard, A.; Palos-Ladeiro, M. The immune system of the freshwater zebra mussel, Dreissena polymorpha, decrypted by proteogenomics of hemocytes and plasma compartments. J. Proteomics 2019, 202, 103366. [Google Scholar] [CrossRef] [PubMed]
- Campos, A.; Apraiz, I.; da Fonseca, R.R.; Cristobal, S. Shotgun analysis of the marine mussel Mytilus edulis hemolymph proteome and mapping the innate immunity elements. Proteomics 2015, 15, 4021–4029. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lemaitre, B.; Hoffmann, J. The host defense of Drosophila melanogaster. Annu. Rev. Immunol. 2007, 25, 697–743. [Google Scholar] [CrossRef] [Green Version]
- Salcedo-Porras, N.; Lowenberger, C. The Immune System of Triatomines; Springer: Cham, Switzerland, 2021; ISBN 9783030645472. [Google Scholar]
- Nishide, Y.; Kageyama, D.; Yokoi, K.; Jouraku, A.; Tanaka, H.; Futahashi, R.; Fukatsu, T. Functional crosstalk across IMD and Toll pathways: Insight into the evolution of incomplete immune cascades. Proc. R. Soc. B Biol. Sci. 2019, 286, 20182207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mesquita, R.D.; Vionette-Amaral, R.J.; Lowenberger, C.; Rivera-Pomar, R.; Monteiro, F.A.; Minx, P.; Spieth, J.; Carvalho, A.B.; Panzera, F.; Lawson, D.; et al. Genome of Rhodnius prolixus, an insect vector of Chagas disease, reveals unique adaptations to hematophagy and parasite infection. Proc. Natl. Acad. Sci. USA 2015, 112, 14936–14941. [Google Scholar] [CrossRef] [Green Version]
- Zumaya-Estrada, F.A.; Martínez-Barnetche, J.; Lavore, A.; Rivera-Pomar, R.; Rodríguez, M.H. Comparative genomics analysis of triatomines reveals common first line and inducible immunity-related genes and the absence of IMD canonical components among hemimetabolous arthropods. Parasites Vectors 2018, 11, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Ursic-Bedoya, R.J.; Lowenberger, C.A. Rhodnius prolixus: Identification of immune-related genes up-regulated in response to pathogens and parasites using suppressive subtractive hybridization. Dev. Comp. Immunol. 2007, 31, 109–120. [Google Scholar] [CrossRef]
- Kim, Y.S.; Ryu, J.H.; Han, S.J.; Choi, K.H.; Nam, K.B.; Jang, I.H.; Lemaitre, B.; Brey, P.T.; Lee, W.J. Gram-negative bacteria-binding protein, a pattern recognition receptor for lipopolysaccharide and β-1,3-glucan that mediates the signaling for the induction of innate immune genes in Drosophila melanogaster cells. J. Biol. Chem. 2000, 275, 32721–32727. [Google Scholar] [CrossRef] [Green Version]
- Diaz-albiter, H.M. Biochemical and Functional Characterization of Glycoside Hydrolase Family 16 Genes in Aedes aegypti Larvae: Identification of the Major Digestive β-1,3-Glucanase. Front. Physiol. 2019, 10, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Ouali, R.; Vieira, L.R.; Salmon, D.; Bousbata, S. Early Post-Prandial Regulation of Protein Expression in the Midgut of Chagas Disease Vector Rhodnius prolixus Highlights New Potential Targets for Vector Control Strategy. Microorganisms 2021, 9, 804. [Google Scholar] [CrossRef]
- Ouali, R.; de Brito, K.C.V.; Salmon, D.; Bousbata, S. High-Throughput Identification of the Rhodnius prolixus Midgut Proteome Unravels a Sophisticated Hematophagic Machinery. Proteomes 2020, 8, 16. [Google Scholar] [CrossRef] [PubMed]
- Inohara, N.; Nuez, G. ML—A conserved domain involved in innate immunity and lipid metabolism. Trends Biochem. Sci. 2002, 27, 219–221. [Google Scholar] [CrossRef]
- Merle, N.S.; Church, S.E.; Fremeaux-Bacchi, V.; Roumenina, L.T. Complement system part I—molecular mechanisms of activation and regulation. Front. Immunol. 2015, 6, 1–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rangarajan, N.; Bakshi, S.; Weisshaar, J.C. Localized Permeabilization of E. coli Membranes by the Antimicrobial Peptide Cecropin A. Biochemistry 2013, 52, 6584–6594. [Google Scholar] [CrossRef] [Green Version]
- Oppedijk, S.F.; Martin, N.I.; Breukink, E. Hit ’em where it hurts: The growing and structurally diverse family of peptides that target lipid-II. BBA-Biomembr. 2015, 1858, 947–957. [Google Scholar] [CrossRef]
- Wenzel, M.; Chiriac, A.I.; Otto, A.; Zweytick, D.; May, C.; Schumacher, C.; Gust, R.; Albada, H.B.; Penkova, M.; Krämer, U.; et al. Small cationic antimicrobial peptides delocalize peripheral membrane proteins. Proc. Natl. Acad. Sci. USA 2014, 111, E1409–E1418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boulanger, N.; Bulet, P.; Lowenberger, C. Antimicrobial peptides in the interactions between insects and flagellate parasites. Trends Parasitol. 2006, 22, 262–268. [Google Scholar] [CrossRef]
- Imler, J.L.; Bulet, P. Antimicrobial peptides in Drosophila: Structures, activities and gene regulation. Chem. Immunol. Allergy 2005, 86, 1–21. [Google Scholar] [CrossRef]
- Hanson, M.A.; Dostálová, A.; Ceroni, C.; Poidevin, M.; Kondo, S.; Lemaître, B. Correction: Synergy and remarkable specificity of antimicrobial peptides in vivo using a systematic knockout approach. Elife 2019, 8, e48778. [Google Scholar] [CrossRef]
- Ghosh, D.; Porter, E.; Shen, B.; Lee, S.K.; Wilk, D.; Drazba, J.; Yadav, S.P.; Crabb, J.W.; Ganz, T.; Bevins, C.L. Paneth cell trypsin is the processing enzyme for human defensin-5. Nat. Immunol. 2002, 3, 583–590. [Google Scholar] [CrossRef]
- Lopez, L.; Morales, G.; Ursic, R.; Wolff, M.; Lowenberger, C. Isolation and characterization of a novel insect defensin from Rhodnius prolixus, a vector of Chagas disease. Insect Biochem. Mol. Biol. 2003, 33, 439–447. [Google Scholar] [CrossRef]
- Vieira, C.S.; Waniek, P.J.; Castro, D.P.; Mattos, D.P.; Moreira, O.C.; Azambuja, P. Impact of Trypanosoma cruzi on antimicrobial peptide gene expression and activity in the fat body and midgut of Rhodnius prolixus. Parasit. Vectors 2016, 9, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boulanger, N.; Lowenberger, C.; Volf, P.; Ursic, R.; Sigutova, L.; Sabatier, L.; Svobodova, M.; Beverley, S.M.; Spa, G.; Brun, R.; et al. Characterization of a Defensin from the Sand Fly Phlebotomus duboscqi Induced by Challenge with Bacteria or the Protozoan Parasite Leishmania major. Infect. Immun. 2004, 72, 7140–7146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boulanger, N. Immunopeptides in the defense reactions of Glossina morsitans to bacterial and Trypanosoma brucei brucei infections. Insect Biochem. Mol. Biol. 2002, 32, 369–375. [Google Scholar] [CrossRef]
- Boulanger, N.; Bulet, P.; Imler, J. Immune response of Drosophila melanogaster to infection with the flagellate parasite Crithidia spp. Insect Biochem. Mol. Biol. 2001, 31, 129–137. [Google Scholar] [CrossRef]
- Castro, D.P.; Moraes, C.S.; Gonzalez, M.S.; Ratcliffe, N.A.; Azambuja, P.; Garcia, E.S. Trypanosoma cruzi immune response modulation decreases microbiota in rhodnius prolixus gut and is crucial for parasite survival and development. PLoS ONE 2012, 7, 3–10. [Google Scholar] [CrossRef] [Green Version]
- Dias, F.d.A.; Guerra, B.; Vieira, L.R.; Perdomo, H.D.; Gandara, A.C.P.; do Amaral, R.J.V.; Vollú, R.E.; Gomes, S.A.O.; Lara, F.A.; Sorgine, M.H.F.; et al. Monitoring of the Parasite Load in the Digestive Tract of Rhodnius prolixus by Combined qPCR Analysis and Imaging Techniques Provides New Insights into the Trypanosome Life Cycle. PLoS Negl. Trop. Dis. 2015, 9, e0004186. [Google Scholar] [CrossRef] [Green Version]
- Nash, J.A.; Ballard, T.N.S.; Weaver, T.E.; Akinbi, H.T. The Peptidoglycan-Degrading Property of Lysozyme Is Not Required for Bactericidal Activity In Vivo. J. Immunol. 2006, 177, 519–526. [Google Scholar] [CrossRef] [Green Version]
- Ursic-Bedoya, R.J.; Nazzari, H.; Cooper, D.; Triana, O.; Wolff, M.; Lowenberger, C. Identification and characterization of two novel lysozymes from Rhodnius prolixus, a vector of Chagas disease. J. Insect Physiol. 2008, 54, 593–603. [Google Scholar] [CrossRef]
- Santiago, P.B.; Assumpção, T.C.F.; de Araújo, C.N.; Bastos, I.M.D.; Neves, D.; da Silva, I.G.; Charneau, S.; Queiroz, R.M.L.; Raiol, T.; de Araújo Oliveira, J.V.; et al. A Deep Insight into the Sialome of Rhodnius neglectus, a Vector of Chagas Disease. PLoS Negl. Trop. Dis. 2016, 10, 1–25. [Google Scholar] [CrossRef] [Green Version]
- Bachali, S.; Jager, M.; Hassanin, A.; Schoentgen, F.; Jollès, P.; Fiala-Medioni, A.; Deutsch, J.S. Phylogenetic analysis of invertebrate lysozymes and the evolution of lysozyme function. J. Mol. Evol. 2002, 54, 652–664. [Google Scholar] [CrossRef] [PubMed]
- Ursic-Bedoya, R.; Buchhop, J.; Joy, J.B.; Durvasula, R.; Lowenberger, C. Prolixicin: A novel antimicrobial peptide isolated from Rhodnius prolixus with differential activity against bacteria and Trypanosoma cruzi. Insect Mol. Biol. 2011, 20, 775–786. [Google Scholar] [CrossRef] [PubMed]
- Hedengren, M.; Borge, K.; Hultmark, D. Expression and evolution of the Drosophila Attacin/Diptericin gene family. Biochem. Biophys. Res. Commun. 2000, 279, 574–581. [Google Scholar] [CrossRef] [PubMed]
- González-Santoyo, I.; Córdoba-Aguilar, A. Phenoloxidase: A key component of the insect immune system. Entomol. Exp. Appl. 2012, 142, 1–16. [Google Scholar] [CrossRef]
- Christensen, B.M.; Li, J.; Chen, C.; Nappi, A.J. Melanization immune responses in mosquito vectors. Trends Parasitol. 2005, 21, 192–199. [Google Scholar] [CrossRef] [PubMed]
- Nakhleh, J.; El Moussawi, L.; Osta, M.A. The Melanization Response in Insect Immunity, 1st ed.; Elsevier Ltd.: Amsterdam, The Netherlands, 2017; Volume 52. [Google Scholar]
- Gomes, S.A.O.; Feder, D.; Garcia, E.S.; Azambuja, P. Suppression of the prophenoloxidase system in Rhodnius prolixus orally infected with Trypanosoma rangeli. J. Insect Physiol. 2003, 49, 829–837. [Google Scholar] [CrossRef]
- Bryant, W.B.; Michel, K. Blood feeding induces hemocyte proliferation and activation in the African malaria mosquito, Anopheles gambiae Giles. J. Exp. Biol. 2014, 217, 1238–1245. [Google Scholar] [CrossRef] [Green Version]
- Eichler, S.; Schaub, G.A. Development of symbionts in triatomine bugs and the effects of infections with trypanosomatids. Exp. Parasitol. 2002, 100, 17–27. [Google Scholar] [CrossRef]
- Genta, F.A.; Souza, R.S.; Garcia, E.S.; Azambuja, P. Phenol oxidases from Rhodnius prolixus: Temporal and tissue expression pattern and regulation by ecdysone. J. Insect Physiol. 2010, 56, 1253–1259. [Google Scholar] [CrossRef]
- Kanost, M.R.; Jiang, H. Clip-domain serine proteases as immune factors in insect hemolymph. Curr. Opin. Insect Sci. 2015, 11, 47–55. [Google Scholar] [CrossRef] [Green Version]
- Jiang, H.; Kanost, M.R. The clip-domain family of serine proteinases in arthropods. Insect Biochem. Mol. Biol. 2000, 30, 95–105. [Google Scholar] [CrossRef]
- Gubb, D.; Sanz-Parra, A.; Barcena, L.; Troxler, L.; Fullaondo, A. Protease inhibitors and proteolytic signalling cascades in insects. Enferm. Infecc. Microbiol. Clin. 2010, 28, 1749–1759. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.Q.; Jiang, H.; Wang, Y.; Kanost, M.R. Nonproteolytic serine proteinase homologs are involved in prophenoloxidase activation in the tobacco hornworm, Manduca sexta. Insect Biochem. Mol. Biol. 2003, 33, 197–208. [Google Scholar] [CrossRef]
- Volz, J.; Müller, H.M.; Zdanowicz, A.; Kafatos, F.C.; Osta, M.A. A genetic module regulates the melanization response of Anopheles to Plasmodium. Cell. Microbiol. 2006, 8, 1392–1405. [Google Scholar] [CrossRef]
- Gulley, M.M.; Zhang, X.; Michel, K. The roles of serpins in mosquito immunology and physiology. J. Insect Physiol. 2013, 59, 138–147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Z.; Wilhelmsson, C.; Hyrsl, P.; Loof, T.G.; Dobes, P.; Klupp, M.; Ikle, J.; Cripps, R.M.; Herwald, H.; Theopold, U.; et al. Pathogen Entrapment by Transglutaminase—A Conserved Early Innate Immune Mechanism. PLoS Pathog. 2010, 6, e1000763. [Google Scholar] [CrossRef] [Green Version]
- Shibata, T.; Sekihara, S.; Fujikawa, T.; Miyaji, R.; Maki, K.; Ishihara, T.; Koshiba, T.; Kawabata, S. Transglutaminase-Catalyzed Protein-Protein Cross-Linking Suppresses the Activity of the NF-kB—Like Transcription Factor Relish. Sci. Signal. 2013, 6, ra61. [Google Scholar] [CrossRef] [PubMed]
- Nsango, S.E.; Pompon, J.; Xie, T.; Rademacher, A.; Fraiture, M.; Thoma, M.; Awono-ambene, P.H.; Moyou, R.S.; Morlais, I.; Levashina, E.A. AP-1/Fos-TGase2 Axis Mediates Wounding-induced Plasmodium falciparum Killing in Anopheles gambiae. J. Biol. Chem. 2013, 288, 16145–16154. [Google Scholar] [CrossRef] [Green Version]
- Felton, G.W.; Summers, C.B. Antioxidant systems in insects. Arch. Insect Biochem. Physiol. 1995, 29, 187–197. [Google Scholar] [CrossRef]
- Su, X.; Gastinel, L.N.; Vaughn, D.E. Crystal Structure of Hemolin: A Horseshoe Shape with Implications for Hemophilic Adhesion. Science 1998, 281, 991–995. [Google Scholar] [CrossRef]
- Andersson, K.; Steiner, H.K.A.N. Structure and properties of protein P4, the major bacteria-inducible protein in pupae of Hyalophora cecropia. Insect Biochem. 1987, 17, 133–140. [Google Scholar] [CrossRef]
- Ladendorff, N.E.; Kanost, M.R. Isolation and Characterization of Bacteria- Induced Protein P4 From Hemolymph of Manduca sexta. Arch. Insect Biochem. Physiol. 1990, 41, 33–41. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.; Horodyski, F.M.; Valaitis, A.P.; Denlinger, D.L. Molecular characterization of the insect immune protein hemolin and its high induction during embryonic diapause in the gypsy moth, Lymantria dispar. Insect Biochem. Mol. Biol. 2002, 32, 1457–1467. [Google Scholar] [CrossRef]
- Kanost, M.R.; Kanost, M.R. Innate immune responses of a lepidopteran insect, Manduca sexta. Immunol. Rev. 2004, 198, 97–105. [Google Scholar] [CrossRef] [PubMed]
- Ladendorff, N.E.; Kanost, M.R. Bacteria-induced protein P4 (hemolin) from Manduca sexta: A member of the immunoglobulin superfamily which can inhibit hemocyte aggregation. Arch. Insect Biochem. Physiol. 1991, 300, 285–300. [Google Scholar] [CrossRef]
- Wang, Y.; Willott, E.; Kanost, M.R. Organization and expression of the hemolin gene, a member of the immunoglobulin superfamily in an insect, Manduca sexta. Insect Mol. Biol. 1995, 4, 113–123. [Google Scholar] [CrossRef]
- Yu, X.; Kanost, M.R. Binding of hemolin to bacterial lipopolysaccharide and lipoteichoic acid receptor. Eur. J. Biochem. 2002, 1834, 1827–1834. [Google Scholar] [CrossRef]
- Nonaka, M.; Yoshizaki, F. Primitive complement system of invertebrates. Mol. Immunol. 2004, 198, 203–215. [Google Scholar] [CrossRef]
- Carland, T.M.; Gerwick, L. The C1q domain containing proteins: Where do they come from and what do they do? Dev. Comp. Immunol. 2010, 34, 785–790. [Google Scholar] [CrossRef]
- Fujita, T.; Endo, Y.; Nonaka, M. Primitive complement system—Recognition and activation. Mol. Immunol. 2004, 41, 103–111. [Google Scholar] [CrossRef]
- Adema, C.M.; Hanington, P.C.; Lun, C.; Rosenberg, G.H.; Aragon, A.D.; Stout, B.A.; Lennard, M.L.; Gross, P.S.; Loker, E.S. Differential transcriptomic responses of Biomphalaria glabrata (Gastropoda, Mollusca) to bacteria and metazoan parasites, Schistosoma mansoni and Echinostoma paraensei (Digenea, Platyhelminthes). Mol. Immunol. 2020, 47, 849–860. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hidayat, P.; Goodmant, W.G. Juvenile Hormone and Hemolymph Juvenile Hormone Binding Protein Titers and their Interaction in the Hemolymph of Fourth Stadium Manduca sexta. Insect Biochem. Mol. Biol. 1994, 24, 709–715. [Google Scholar] [CrossRef]
- Wigglesworth, B.Y.V.B. The Action of Moulting Hormone and Juvenile Hormone at the Cellular Level in Rhodnius prolixus. J. Exp. Biol. 1963, 1, 231–245. [Google Scholar] [CrossRef]
- Wang, Z.; Davey, K.G. The Role of Juvenile Hormone in Vitellogenin Production in Rhodnius prolixus. J. Insect Physiol. 1993, 39, 471–476. [Google Scholar] [CrossRef]
- Flatt, T.; Heyland, A.; Rus, F.; Porpiglia, E.; Sherlock, C.; Yamamoto, R.; Garbuzov, A.; Palli, S.R.; Tatar, M.; Silverman, N. Hormonal regulation of the humoral innate immune response in Drosophila melanogaster. J. Exp. Biol. 2008, 211, 2712–2724. [Google Scholar] [CrossRef] [Green Version]
- Id, C.; Aryan, A.; Kim, I.H.; Ce, J.; Nouzova, M.; Id, F.G.N.; Beatriz, A.; Id, F.B.; Id, E.C.; Id, M.C.R.; et al. A mosquito juvenile hormone binding protein (mJHBP) regulates the activation of innate immune defenses and hemocyte development. PLoS Pathog. 2020, 16, e1008288. [Google Scholar]
- Perez-riverol, Y.; Bai, J.; Bandla, C.; Garc, D.; Hewapathirana, S.; Kamatchinathan, S.; Kundu, D.J.; Prakash, A.; Frericks-zipper, A.; Eisenacher, M.; et al. The PRIDE database resources in 2022: A hub for mass spectrometry-based proteomics evidences. Nucleic Acids Res. 2022, 50, 543–552. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Immune Category | IDs | Protein Names | Fold Increase |
|---|---|---|---|
| Pattern recognition receptors (PRRs) |  | ||
| Peptidoglycan recognition receptor (PGRPs) | rpPGRP-LC/LAa |  | |
 | |||
| rpPGRP-LC/LAb | |||
| Gram-negative binding proteins (GNBPs) | B8LJ39 | Beta-GRP |  |
 | |||
 | |||
 | |||
| T1HGN7 | GH16 domain-containing protein |  | |
 | |||
 | |||
 | |||
| T1I650 | GH16 domain-containing protein |  | |
 | |||
 | |||
 | |||
| T1HU92 | ML domain-containing protein |  | |
 | |||
 | |||
 | |||
| Mucins | T1HEN7 | Putative mucin |  |
 | |||
 | |||
 | |||
| Antimicrobial peptides (AMPs) | |||
| Lysozymes | T1I5M5 | Lysozyme |  |
 | |||
 | |||
 | |||
| A9LN32 | Lysozyme |  | |
 | |||
| Prolixicins | B8QEI8 | Prolixin antimicrobial peptide |  |
 | |||
 | |||
 | |||
| Attacins | T1I7V7 | Attacin_C domain-containing protein |  |
 | |||
 | |||
 | |||
| Defensins | T1I7B0 | INVERT_DEFENSINS domain-containing protein |  |
 | |||
| R4G8B6 | Putative defensin |  | |
 | |||
| R4FNJ9 | Putative defensin a |  | |
 | |||
 | |||
| Diptericins | D6BJP6 E6Y430 | Diptericin |  |
 | |||
 | |||
 | |||
 | |||
| Glycine rich peptides | T1HS54 | Uncharacterized protein |  |
 | |||
 | |||
 | |||
| R4G489 | Putative glycine-rich cuticle protein | ||
 | |||
 | |||
| A0A4P6DAB8 | Putative glycine-rich cuticle protein rhodnius neglectus | ||
 | |||
 | |||
| A0A4P6D8R0 | Putative glycine-rich cuticle protein rhodnius neglectus | ||
 | |||
 | |||
| Melanization | |||
| Prophenoloxidases | T1I7V8 | Phenoloxidase |  |
 | |||
 | |||
 | |||
| A0A1B2G385 | Phenoloxidase 1 (EC 1.14.18.1) |  | |
 | |||
 | |||
 | |||
| T1HW62 | Phenoloxidase (Fragment) |  | |
 | |||
 | |||
 | |||
| A0A1B2G381 | Phenoloxidase 2 (EC 1.14.18.1) |  | |
 | |||
 | |||
 | |||
| T1HW22 | Phenoloxidase (Fragment) |  | |
 | |||
 | |||
 | |||
| Serine proteases | |||
| Clip-domain SP | T1HGB7 | CLIP domain-containing serine protease (EC 3.4.21.-) |  |
 | |||
 | |||
 | |||
| R4FQA1 | CLIP domain-containing serine protease (EC 3.4.21.-) |  | |
 | |||
 | |||
 | |||
| B8QQQ1 | CLIP domain-containing serine protease (EC 3.4.21.-) |  | |
 | |||
 | |||
 | |||
| Protease inhibitors | |||
| Serpins | T1IF83 | SERPIN domain-containing protein |  |
 | |||
 | |||
 | |||
| R4FJD2 | Putative serpin length |  | |
 | |||
 | |||
 | |||
| T1I8D5 | SERPIN domain-containing protein |  | |
 | |||
 | |||
 | |||
| R4FLP4 | Putative serpin length |  | |
 | |||
 | |||
 | |||
| Cystatins | R4FP01 | Cystatin (Putative secreted protein) |  |
 | |||
 | |||
 | |||
| T1I2F3 | Cystatin domain-containing protein |  | |
 | |||
 | |||
 | |||
| R4G3U6 | Protein with 4 pacifastin inhibitor domains lcmii |  | |
 | |||
 | |||
 | |||
| Detoxification | |||
| Superoxide dismutases | R4FMI6 | Superoxide dismutase [Cu-Zn] (EC 1.15.1.1) |  |
 | |||
 | |||
 | |||
| G1K083 | Superoxide dismutase (EC 1.15.1.1) (Fragment) | ||
 | |||
 | |||
| T1HRT6 | Superoxide dismutase [Cu-Zn] (EC 1.15.1.1) |  | |
 | |||
 | |||
 | |||
| A0A4P6D9T0 | Superoxide dismutase (EC 1.15.1.1) (Fragment) |  | |
 | |||
 | |||
 | |||
| R4FPK6 | Superoxide dismutase (EC 1.15.1.1) (Fragment) |  | |
 | |||
 | |||
 | |||
| Catalases | T1I0W4 | Catalase domain-containing protein (Fragment) |  |
 | |||
| T1HV37 | Catalase domain-containing protein |  | |
 | |||
| Peroxidases | T1I489 | Glutathione peroxidase (Fragment) |  |
 | |||
 | |||
 | |||
| Glutathione S-transferases | T1HVN9 | Glutathione S-transferase domain containing protein |  |
 | |||
 | |||
 | |||
| R4G417 | Putative glutathione S-transferase |  | |
 | |||
 | |||
 | |||
| T1HUM1 | Putative glutathione S-transferase | ||
 | |||
 | |||
| Transglutaminases | T1HFP9 | TGc domain-containing protein (Fragment) |  |
 | |||
 | |||
 | |||
| T1HFS7 | TGc domain-containing protein (Fragment) |  | |
 | |||
 | |||
 | |||
| T1HFV3 | TGc domain-containing protein |  | |
 | |||
| T1HFR5 | TGc domain-containing protein (Fragment) |  | |
 | |||
 | |||
 | |||
| T1HFR9 | TGc domain-containing protein (Fragment) |  | |
 | |||
 | |||
 | |||
| T1HFV2 | Uncharacterized protein |  | |
 | |||
| T1I362 | Transglut_C domain-containing protein |  | |
 | |||
 | |||
 | |||
| Transferrins | B8LJ43 | Transferrin |  |
 | |||
 | |||
 | |||
| T1HAU6 | Melanotransferrin |  | |
 | |||
 | |||
 | |||
| A0A4P6DAP6 | Putative transferrin isoform x4 (Fragment) |  | |
 | |||
 | |||
 | |||
| Opsonization | A0A4P6D700 | Putative transmembrane protein of the immunoglobulin family of cell adhesion molecules (Fragment) |  |
 | |||
 | |||
 | |||
| T1HCN4 | I-set domain-containing protein |  | |
 | |||
 | |||
 | |||
| R4FJF3 | Putative c1q domain protein |  | |
 | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ouali, R.; Vieira, L.R.; Salmon, D.; Bousbata, S. Rhodnius prolixus Hemolymph Immuno-Physiology: Deciphering the Systemic Immune Response Triggered by Trypanosoma cruzi Establishment in the Vector Using Quantitative Proteomics. Cells 2022, 11, 1449. https://doi.org/10.3390/cells11091449
Ouali R, Vieira LR, Salmon D, Bousbata S. Rhodnius prolixus Hemolymph Immuno-Physiology: Deciphering the Systemic Immune Response Triggered by Trypanosoma cruzi Establishment in the Vector Using Quantitative Proteomics. Cells. 2022; 11(9):1449. https://doi.org/10.3390/cells11091449
Chicago/Turabian StyleOuali, Radouane, Larissa Rezende Vieira, Didier Salmon, and Sabrina Bousbata. 2022. "Rhodnius prolixus Hemolymph Immuno-Physiology: Deciphering the Systemic Immune Response Triggered by Trypanosoma cruzi Establishment in the Vector Using Quantitative Proteomics" Cells 11, no. 9: 1449. https://doi.org/10.3390/cells11091449
APA StyleOuali, R., Vieira, L. R., Salmon, D., & Bousbata, S. (2022). Rhodnius prolixus Hemolymph Immuno-Physiology: Deciphering the Systemic Immune Response Triggered by Trypanosoma cruzi Establishment in the Vector Using Quantitative Proteomics. Cells, 11(9), 1449. https://doi.org/10.3390/cells11091449