
The Phenotype of the Adipocytes Derived from Subcutaneous and Visceral ADMSCs Is Altered When They Originate from Morbidly Obese Women: Is There a Memory Effect?

 ,
,  , , , , and
, , , , and
Abstract

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. The Origin of Human Adipose-Derived Mesenchymal Stem Cells (ADMSCs)
2.2. ADMSCs Differentiation towards Adipocytes
2.2.1. Adipogenesis of ADMSCs
2.2.2. Assessment of the Accumulated Lipids
2.3. Flow Cytometry Characterization
2.4. RNA Isolation and Quantitative Real Time RT-PCR
2.5. Immunoblotting
2.6. 9,10-[3H]-Palmitic Acid Uptake
2.7. Lipid Content Quantification (Gas Liquid Chromatography)
2.8. Statistical Analysis
3. Results
3.1. Characterization of Human ADMSCs
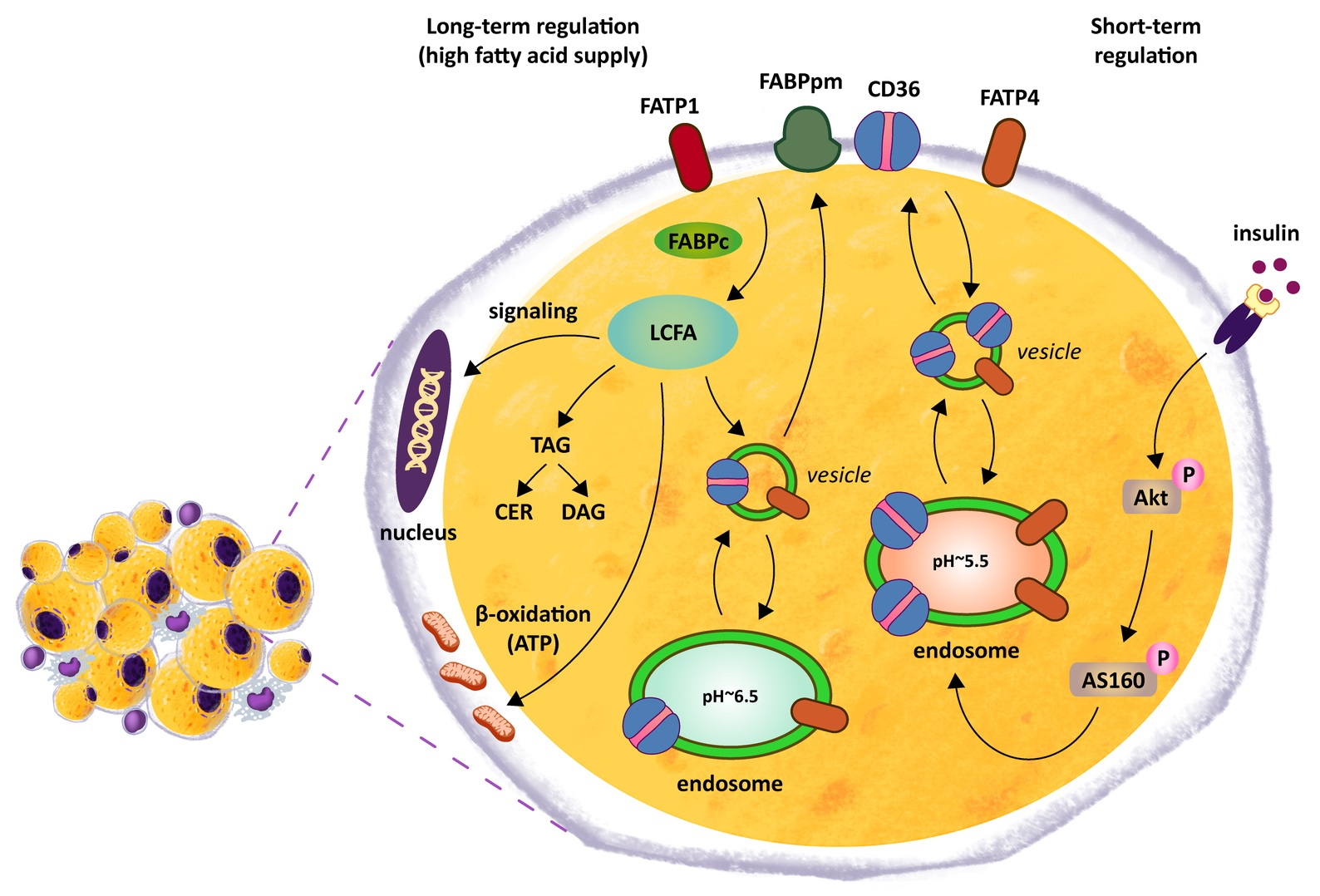
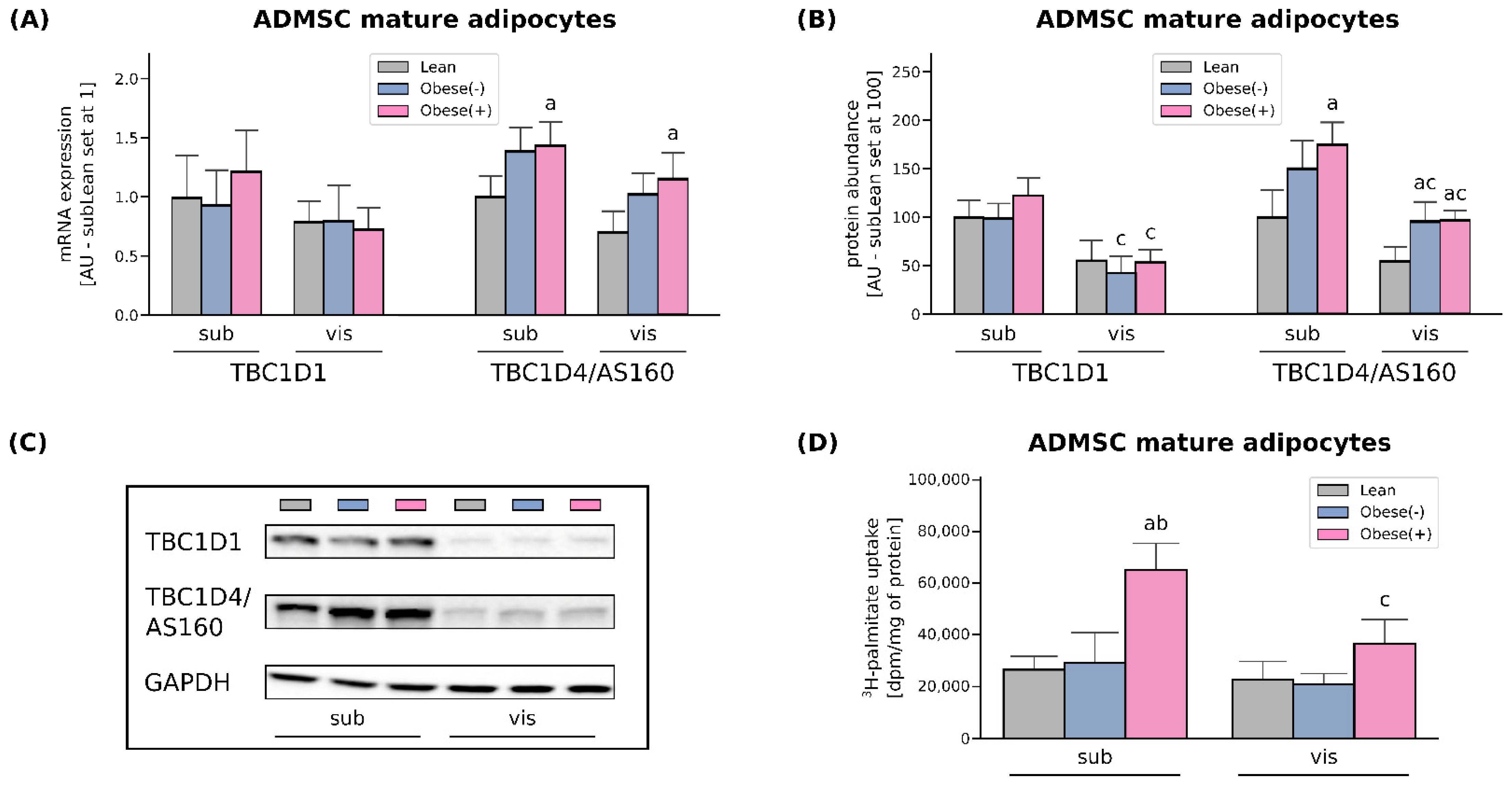
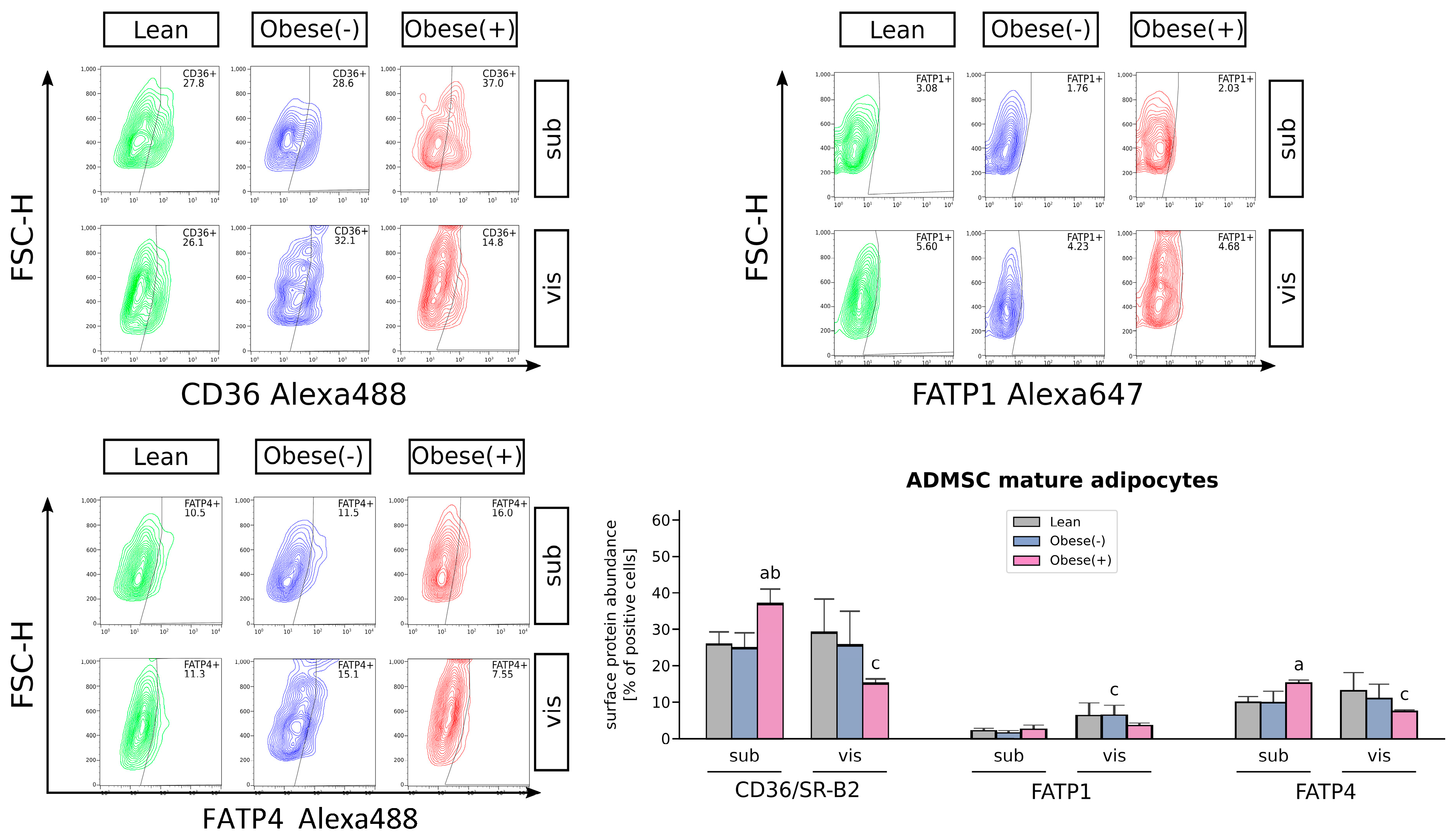
3.2. SLC27A4/FATP4 and CD36/SR-B2, but Not FATP1 or FABPpm, Contribute to Increased Fatty Acid Uptake in Obese Adipocytes Derived from subADMSCs
3.3. Effect of ADMSCs Tissue Origin and the Metabolic Status of the Patient Donor on Adipocyte Size and Granularity
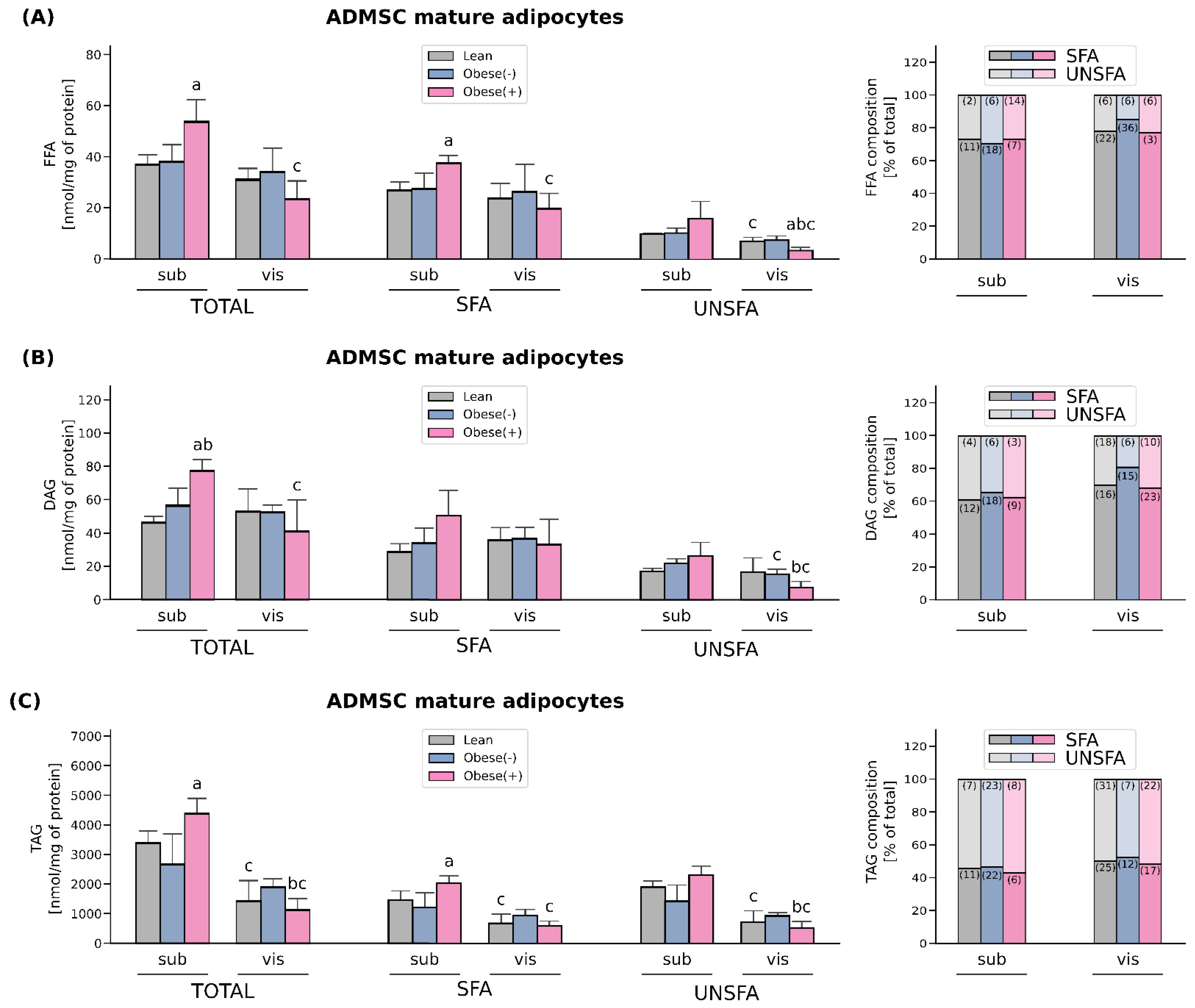
3.4. Enhanced FA Uptake Promotes Lipid Accumulation Together with Changes in the Composition of Different FA Species Mainly in the Adipocytes Derived from subADMSCs of Morbidly Obese Women with Metabolic Syndrome
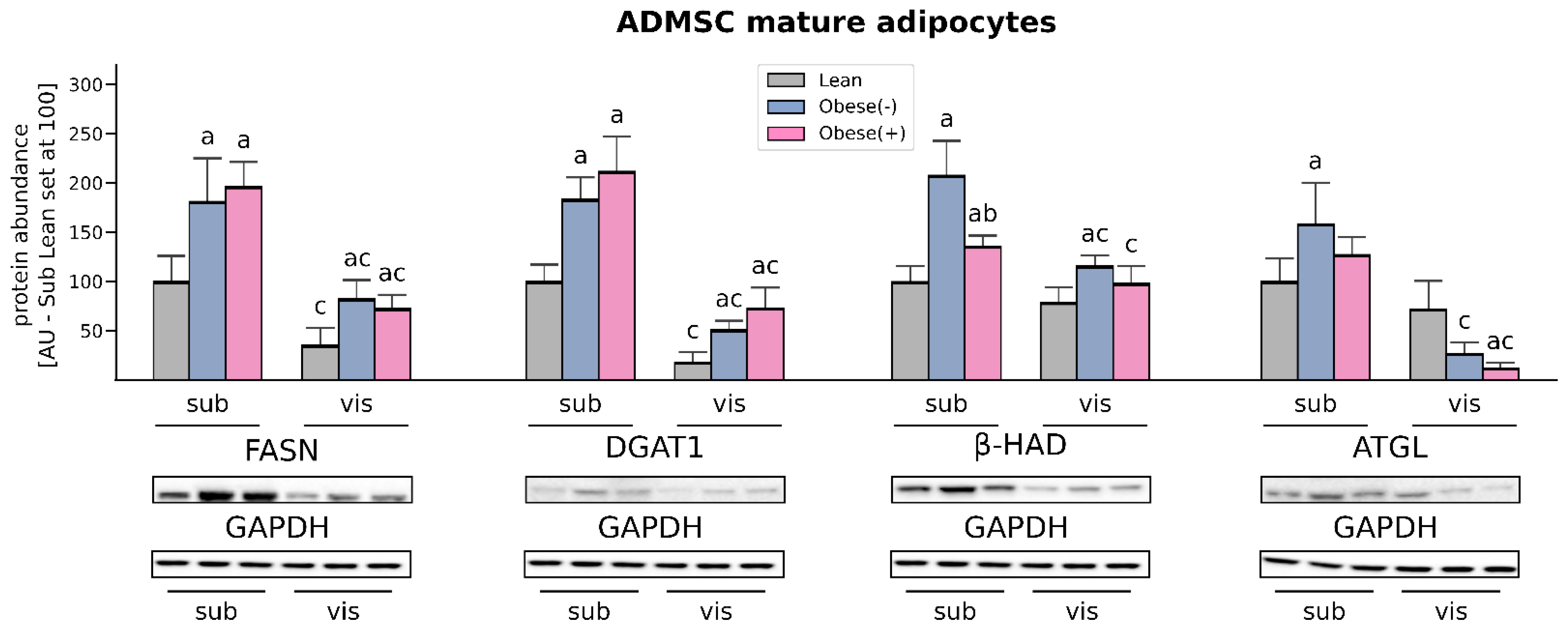
3.5. LCFAs Synthesis and Utilization Are Enhanced in the Adipocytes Derived from ADMSCs of Morbidly Obese Women
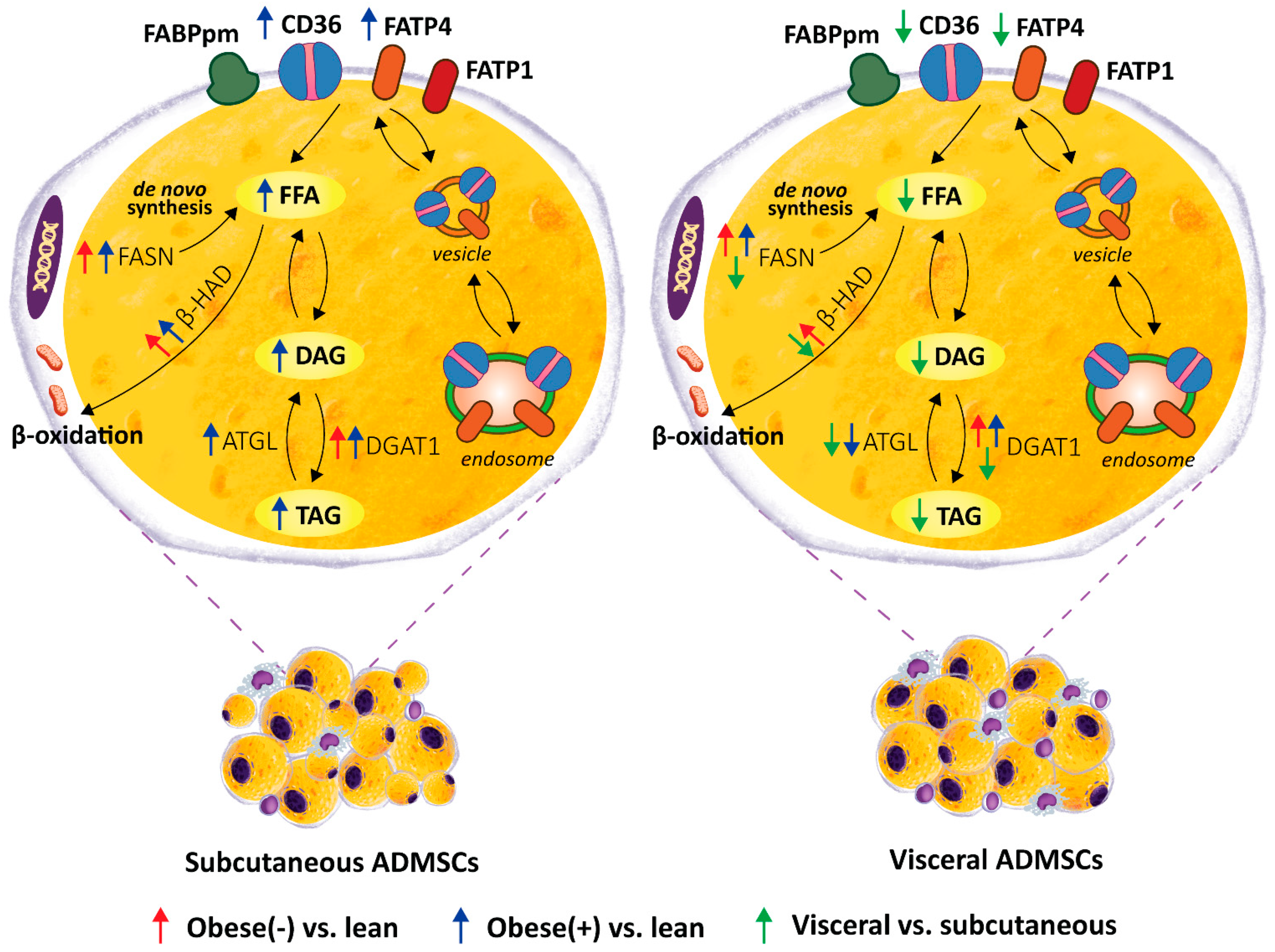
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Mikłosz, A.; Nikitiuk, B.E.; Chabowski, A. Using adipose-derived mesenchymal stem cells to fight the metabolic complications of obesity: Where do we stand? Obes. Rev. 2022, 23, e13413. [Google Scholar] [CrossRef] [PubMed]
- Korac, A.; Srdic-Galic, B.; Kalezic, A.; Stancic, A.; Otasevic, V.; Korac, B.; Jankovic, A. Adipokine signatures of subcutaneous and visceral abdominal fat in normal-weight and obese women with different metabolic profiles. Arch. Med. Sci. 2021, 17, 323–336. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Q.; Fu, Z.; Gong, Y.; Seshachalam, V.P.; Li, J.; Ma, Y.; Liang, H.; Guan, W.; Lin, S.; Ghosh, S.; et al. Metabolic Health Status Contributes to Transcriptome Alternation in Human Visceral Adipose Tissue During Obesity. Obesity 2020, 28, 2153–2162. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, M.M. Subcutaneous and visceral adipose tissue: Structural and functional differences. Obes. Rev. 2010, 11, 11–18. [Google Scholar] [CrossRef]
- Ferrara, D.; Montecucco, F.; Dallegri, F.; Carbone, F. Impact of different ectopic fat depots on cardiovascular and metabolic diseases. J. Cell. Physiol. 2019, 234, 21630–21641. [Google Scholar] [CrossRef]
- Barrea, L.; Vetrani, C.; Caprio, M.; El Ghoch, M.; Frias-Toral, E.; Mehta, R.J.; Mendez, V.; Moriconi, E.; Paschou, S.A.; Pazderska, A.; et al. Nutritional management of type 2 diabetes in subjects with obesity: An international guideline for clinical practice. Crit. Rev. Food Sci. Nutr. 2021, 1–13. [Google Scholar] [CrossRef]
- Abildgaard, J.; Ploug, T.; Al-Saoudi, E.; Wagner, T.; Thomsen, C.; Ewertsen, C.; Bzorek, M.; Pedersen, B.K.; Pedersen, A.T.; Lindegaard, B. Changes in abdominal subcutaneous adipose tissue phenotype following menopause is associated with increased visceral fat mass. Sci. Rep. 2021, 11, 14750. [Google Scholar] [CrossRef]
- Yokokawa, H.; Fukuda, H.; Saita, M.; Goto, K.; Kaku, T.; Miyagami, T.; Takahashi, Y.; Hamada, C.; Hisaoka, T.; Naito, T. An association between visceral or subcutaneous fat accumulation and diabetes mellitus among Japanese subjects. Diabetol. Metab. Syndr. 2021, 13, 44. [Google Scholar] [CrossRef]
- Ritter, A.; Friemel, A.; Roth, S.; Kreis, N.N.; Hoock, S.C.; Safdar, B.K.; Fischer, K.; Möllmann, C.; Solbach, C.; Louwen, F.; et al. Subcutaneous and Visceral Adipose-Derived Mesenchymal Stem Cells: Commonality and Diversity. Cells 2019, 8, 1288. [Google Scholar] [CrossRef]
- Fang, J.; Chen, F.; Liu, D.; Gu, F.; Wang, Y. Adipose tissue-derived stem cells in breast reconstruction: A brief review on biology and translation. Stem Cell Res. Ther. 2021, 12, 8. [Google Scholar] [CrossRef]
- Baglioni, S.; Cantini, G.; Poli, G.; Francalanci, M.; Squecco, R.; Franco, A.; Borgogni, E.; Frontera, S.; Nesi, G.; Liotta, F.; et al. Functional differences in visceral and subcutaneous fat pads originate from differences in the adipose stem cell. PLoS ONE 2012, 7, e36569. [Google Scholar] [CrossRef] [PubMed]
- Abkenar, I.K.; Rahmani-Nia, F.; Lombardi, G. The effects of acute and chronic aerobic activity on the signaling pathway of the inflammasome NLRP3 complex in young men. Medicina 2019, 55, 105. [Google Scholar] [CrossRef] [PubMed]
- Alviz, L.; Tebar-García, D.; Lopez-Rosa, R.; Galan-Moya, E.M.; Moratalla-López, N.; Alonso, G.L.; Nava, E.; Llorens, S. Pathogenic microenvironment from diabetic–obese visceral and subcutaneous adipocytes activating differentiation of human healthy preadipocytes increases intracellular fat, effect of the apocarotenoid crocetin. Nutrients 2021, 13, 1032. [Google Scholar] [CrossRef] [PubMed]
- Tang, Q.Q.; Lane, M.D. Adipogenesis: From stem cell to adipocyte. Annu. Rev. Biochem. 2012, 81, 715–736. [Google Scholar] [CrossRef]
- Silva, K.R.; Liechocki, S.; Carneiro, J.R.; Claudio-Da-Silva, C.; Maya-Monteiro, C.M.; Borojevic, R.; Baptista, L.S. Stromal-vascular fraction content and adipose stem cell behavior are altered in morbid obese and post bariatric surgery ex-obese women. Stem Cell Res. Ther. 2015, 6, 72. [Google Scholar] [CrossRef]
- Silva, K.R.; Côrtes, I.; Liechocki, S.; Carneiro, J.R.I.; Souza, A.A.P.; Borojevic, R.; Maya-Monteiro, C.M.; Baptista, L.S. Characterization of stromal vascular fraction and adipose stem cells from subcutaneous, preperitoneal and visceral morbidly obese human adipose tissue depots. PLoS ONE 2017, 12, e0174115. [Google Scholar] [CrossRef]
- Posada-González, M.; Villagrasa, A.; García-Arranz, M.; Vorwald, P.; Olivera, R.; Olmedillas-López, S.; Vega-Clemente, L.; Salcedo, G.; García-Olmo, D. Comparative Analysis Between Mesenchymal Stem Cells From Subcutaneous Adipose Tissue and Omentum in Three Types of Patients: Cancer, Morbid Obese and Healthy Control. Surg. Innov. 2021, 29, 9–21. [Google Scholar] [CrossRef]
- Suárez-Cuenca, J.A.; De La Peña-Sosa, G.; De La Vega-Moreno, K.; Banderas-Lares, D.Z.; Salamanca-García, M.; Martínez-Hernández, J.E.; Vera-Gómez, E.; Hernández-Patricio, A.; Zamora-Alemán, C.R.; Domínguez-Pérez, G.A.; et al. Enlarged adipocytes from subcutaneous vs. visceral adipose tissue differentially contribute to metabolic dysfunction and atherogenic risk of patients with obesity. Sci. Rep. 2021, 11, 1831. [Google Scholar] [CrossRef]
- Lefevre, C.; Chartoire, D.; Ferraz, J.C.; Verdier, T.; Pinteur, C.; Chanon, S.; Pesenti, S.; Vieille-Marchiset, A.; Genestier, L.; Vidal, H.; et al. Obesity activates immunomodulating properties of mesenchymal stem cells in adipose tissue with differences between localizations. FASEB J. 2021, 35, e21650. [Google Scholar] [CrossRef]
- Mikłosz, A.; Łukaszuk, B.; Supruniuk, E.; Grubczak, K.; Moniuszko, M.; Choromańska, B.; Myśliwiec, P.; Chabowski, A. Does TBC1D4 (AS160) or TBC1D1 Deficiency Affect the Expression of Fatty Acid Handling Proteins in the Adipocytes Differentiated from Human Adipose-Derived Mesenchymal Stem Cells (ADMSCs) Obtained from Subcutaneous and Visceral Fat Depots? Cells 2021, 10, 1515. [Google Scholar] [CrossRef]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef] [PubMed]
- Glatz, J.F.C.; Nabben, M.; Luiken, J.J.F.P. CD36 (SR-B2) as master regulator of cellular fatty acid homeostasis. Curr. Opin. Lipidol. 2022, 33, 103–111. [Google Scholar] [CrossRef] [PubMed]
- Badimon, L.; Cubedo, J. Adipose tissue depots and inflammation: Effects on plasticity and resident mesenchymal stem cell function. Cardiovasc. Res. 2017, 113, 1064–1073. [Google Scholar] [CrossRef] [PubMed]
- Arner, P.; Rydén, M. Human white adipose tissue: A highly dynamic metabolic organ. J. Intern. Med. 2021, 291, 611–621. [Google Scholar] [CrossRef]
- Wang, T.; Sharma, A.K.; Wolfrum, C. Novel insights into adipose tissue heterogeneity. Rev. Endocr. Metab. Disord. 2021, 23, 5–12. [Google Scholar] [CrossRef]
- Thompson, B.R.; Lobo, S.; Bernlohr, D.A. Fatty acid flux in adipocytes: The in’s and out’s of fat cell lipid trafficking. Mol. Cell. Endocrinol. 2010, 318, 24–33. [Google Scholar] [CrossRef]
- Nickerson, J.G.; Alkhateeb, H.; Benton, C.R.; Lally, J.; Nickerson, J.; Han, X.X.; Wilson, M.H.; Jain, S.S.; Snook, L.A.; Glatz, J.F.C.; et al. Greater transport efficiencies of the membrane fatty acid transporters FAT/CD36 and FATP4 compared with FABPpm and FATP1 and differential effects on fatty acid esterification and oxidation in rat skeletal muscle. J. Biol. Chem. 2009, 284, 16522–16530. [Google Scholar] [CrossRef]
- Stahl, A. A current review of fatty acid transport proteins (SLC27). Pflug. Arch. Eur. J. Physiol. 2004, 447, 722–727. [Google Scholar] [CrossRef]
- Bonen, A.; Tandon, N.N.; Glatz, J.F.C.; Luiken, J.J.F.P.; Heigenhauser, G.J.F. The fatty acid transporter FAT/CD36 is upregulated in subcutaneous and visceral adipose tissues in human obesity and type 2 diabetes. Int. J. Obes. 2006, 30, 877–883. [Google Scholar] [CrossRef]
- Gertow, K.; Pietiläinen, K.H.; Yki-Järvinen, H.; Kaprio, J.; Rissanen, A.; Eriksson, P.; Hamsten, A.; Fisher, R.M. Expression of fatty-acid-handling proteins in human adipose tissue in relation to obesity and insulin resistance. Diabetologia 2004, 47, 1118–1125. [Google Scholar] [CrossRef][Green Version]
- Mikłosz, A.; Łukaszuk, B.; Żendzian-Piotrowska, M.; Brańska-Januszewska, J.; Ostrowska, H.; Chabowski, A. Challenging of AS160/TBC1D4 Alters Intracellular Lipid milieu in L6 Myotubes Incubated With Palmitate. J. Cell. Physiol. 2017, 232, 2373–2386. [Google Scholar] [CrossRef] [PubMed]
- Glatz, J.F.C.; Luiken, J. From fat to FAT (CD36/SR-B2): Understanding the regulation of cellular fatty acid uptake. Biochimie 2017, 136, 21–26. [Google Scholar] [CrossRef] [PubMed]
- Binnert, C.; Koistinen, H.A.; Martin, G.; Andreelli, F.; Ebeling, P.; Koivisto, V.A.; Laville, M.; Auwerx, J.; Vidal, H. Fatty acid transport protein-1 mRNA expression in skeletal muscle and in adipose tissue in humans. Am. J. Physiol.-Endocrinol. Metab. 2000, 279, 1072–1079. [Google Scholar] [CrossRef] [PubMed]
- Wajchenberg, B.L.; Lé, B.; Wajchenberg, O. Subcutaneous and Visceral Adipose Tissue: Their Relation to the Metabolic Syndrome. Endocr. Rev. 2000, 21, 697–738. [Google Scholar]
- Joe, A.W.B.; Lin, Y.; Even, Y.; Vogl, A.W.; Rossi, F.M.V. Depot-specific differences in adipogenic progenitor abundance and proliferative response to high-fat diet. Stem Cells 2009, 27, 2563–2570. [Google Scholar] [CrossRef]
- Kim, B.; Lee, B.; Kim, M.K.; Gong, S.P.; Park, N.H.; Chung, H.H.; Kim, H.S.; No, J.H.; Park, W.Y.; Park, A.K.; et al. Gene expression profiles of human subcutaneous and visceral adipose-derived stem cells. Cell Biochem. Funct. 2016, 34, 563–571. [Google Scholar] [CrossRef]
- Zacharia, A.; Saidemberg, D.; Mannully, C.T.; Kogan, N.M.; Shehadeh, A.; Sinai, R.; Zucker, A.; Bruck-Haimson, R.; Goldstein, N.; Haim, Y.; et al. Distinct infrastructure of lipid networks in visceral and subcutaneous adipose tissues in overweight humans. Am. J. Clin. Nutr. 2020, 112, 979–990. [Google Scholar] [CrossRef]
- Małodobra-Mazur, M.; Cierzniak, A.; Pawełka, D.; Kaliszewski, K.; Rudnicki, J.; Dobosz, T. Metabolic Differences between Subcutaneous and Visceral Adipocytes Differentiated with an Excess of Saturated and Monounsaturated Fatty Acids. Genes 2020, 11, 1092. [Google Scholar] [CrossRef]
- Berndt, J.; Kralisch, S.; Klöting, N.; Ruschke, K.; Kern, M.; Fasshauer, M.; Schön, M.R.; Stumvoll, M.; Blüher, M. Adipose triglyceride lipase gene expression in human visceral obesity. Exp. Clin. Endocrinol. Diabetes 2008, 116, 203–210. [Google Scholar] [CrossRef]
- Gaidhu, M.P.; Anthony, N.M.; Patel, P.; Hawke, T.J.; Ceddia, R.B. Dysregulation of lipolysis and lipid metabolism in visceral and subcutaneous adipocytes by high-fat diet: Role of ATGL, HSL, and AMPK. Am. J. Physiol.-Cell Physiol. 2010, 298, 961–971. [Google Scholar] [CrossRef]
- Yao-Borengasser, A.; Varma, V.; Coker, R.H.; Ranganathan, G.; Phanavanh, B.; Rasouli, N.; Kern, P.A. Adipose triglyceride lipase expression in human adipose tissue and muscle. Role in insulin resistance and response to training and pioglitazone. Metabolism 2011, 60, 1012–1020. [Google Scholar] [CrossRef] [PubMed]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mikłosz, A.; Łukaszuk, B.; Supruniuk, E.; Grubczak, K.; Starosz, A.; Kusaczuk, M.; Naumowicz, M.; Chabowski, A. The Phenotype of the Adipocytes Derived from Subcutaneous and Visceral ADMSCs Is Altered When They Originate from Morbidly Obese Women: Is There a Memory Effect? Cells 2022, 11, 1435. https://doi.org/10.3390/cells11091435
Mikłosz A, Łukaszuk B, Supruniuk E, Grubczak K, Starosz A, Kusaczuk M, Naumowicz M, Chabowski A. The Phenotype of the Adipocytes Derived from Subcutaneous and Visceral ADMSCs Is Altered When They Originate from Morbidly Obese Women: Is There a Memory Effect? Cells. 2022; 11(9):1435. https://doi.org/10.3390/cells11091435
Chicago/Turabian StyleMikłosz, Agnieszka, Bartłomiej Łukaszuk, Elżbieta Supruniuk, Kamil Grubczak, Aleksandra Starosz, Magdalena Kusaczuk, Monika Naumowicz, and Adrian Chabowski. 2022. "The Phenotype of the Adipocytes Derived from Subcutaneous and Visceral ADMSCs Is Altered When They Originate from Morbidly Obese Women: Is There a Memory Effect?" Cells 11, no. 9: 1435. https://doi.org/10.3390/cells11091435
APA StyleMikłosz, A., Łukaszuk, B., Supruniuk, E., Grubczak, K., Starosz, A., Kusaczuk, M., Naumowicz, M., & Chabowski, A. (2022). The Phenotype of the Adipocytes Derived from Subcutaneous and Visceral ADMSCs Is Altered When They Originate from Morbidly Obese Women: Is There a Memory Effect? Cells, 11(9), 1435. https://doi.org/10.3390/cells11091435