
Assessment of the Preventive Effect of L-carnitine on Post-statin Muscle Damage in a Zebrafish Model

 , , and
, , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethical Statement
2.2. Animal Maintenance and Handling
2.3. Experiment Design and Lovastatin Treatment
2.4. Birefringence Assay
2.5. Transmission Electron Microscopy
2.6. RNA Isolation, Reverse Transcription, and Real-Time Quantitative PCR (RT-qPCR)
2.7. Behaviour Tests
2.7.1. Spontaneous Displacement Assay
2.7.2. Touch-Evoked Response Assay
2.8. Heartbeat Analysis
2.9. Analysis of Zebrafish Larva Heart Contraction on the Basis of Time-Lapse Images
Statistical Analysis
3. Results
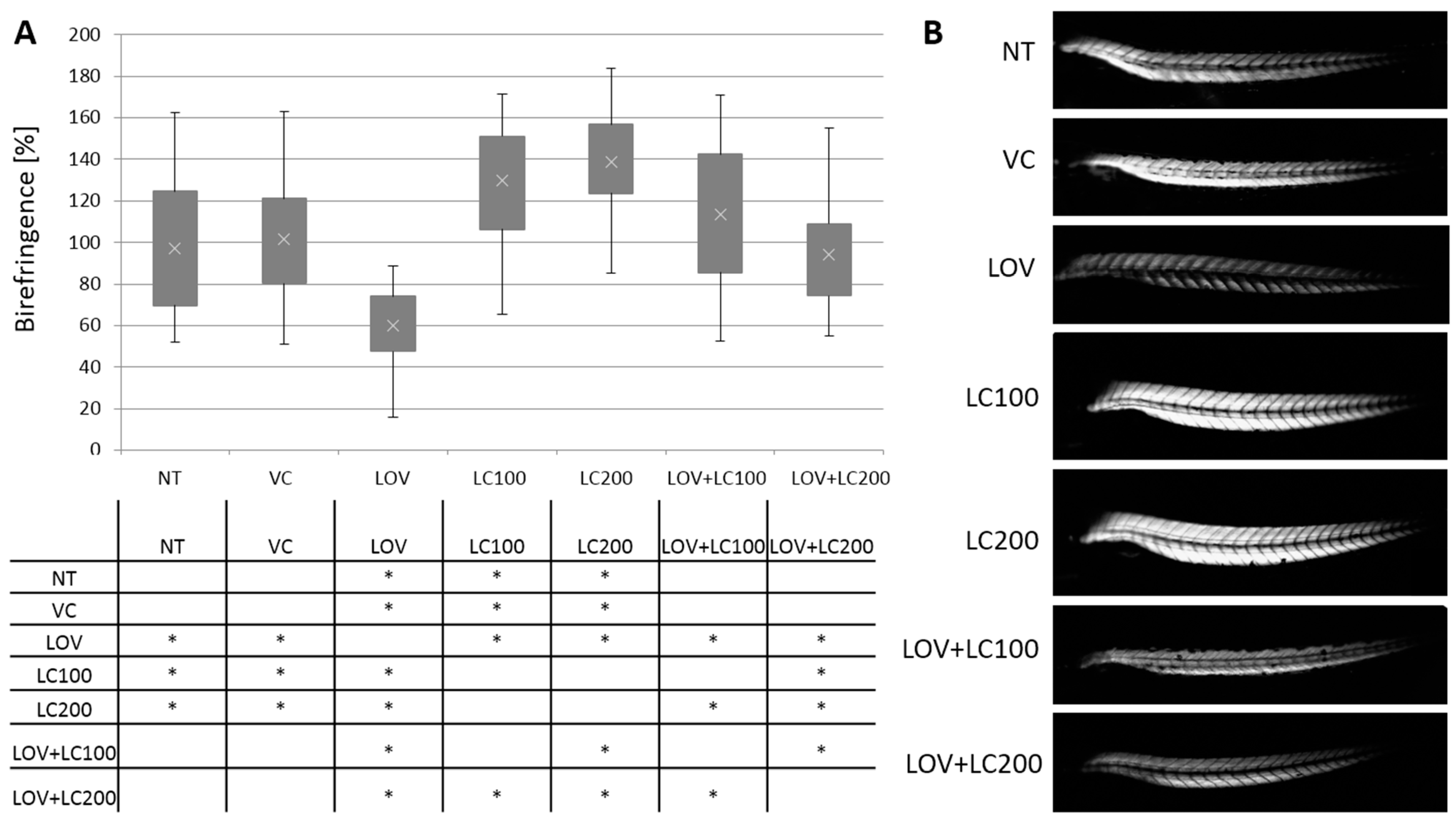
3.1. Lovastatin Treatment Disrupts Zebrafish Larva Skeletal Muscle While L-carnitine Has a Protective Effect
3.2. Lovastatin Treatment Leads to Sarcomere Malformations in Zebrafish Larva Skeletal Muscle while L-carnitine Has a Protective Effect
3.3. Lovastatin Treatment Stimulates Atrogin-1 Expression in Zebrafish Larvae While L-carnitine Has a Protective Effect
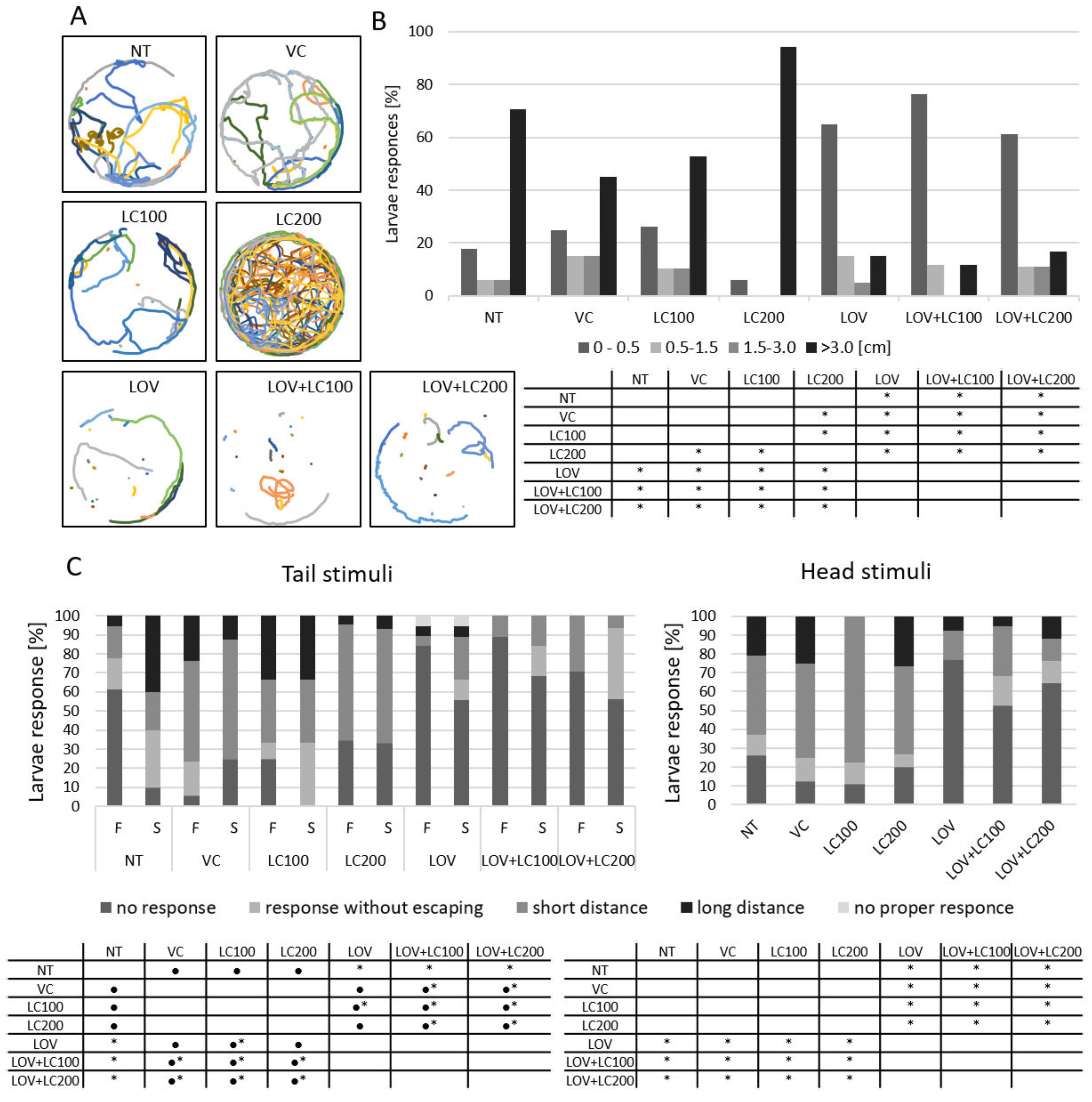
3.4. Lovastatin Treatment Alters Swimming Behaviour of Zebrafish Larvae While L-carnitine has no Protective Effect
3.5. Lovastatin Treatment Alters Heartbeat Rate of Zebrafish Larvae While L-carnitine Has no Protective Effect
3.6. Lovastatin Treatment Affects Heart Contraction in Zebrafish Larvae While L-carnitine Has a Protective Effect
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dalakas, M.C. Toxic and drug-induced myopathies. J. Neurol. Neurosurg. Psychiatry 2009, 80, 832–838. [Google Scholar] [CrossRef] [PubMed]
- Gupta, R.; Alcantara, R.; Popli, T.; Mahajan, S.; Tariq, U.; Dusaj, R.S.; Malik, A.H. Myopathy Associated With Statins and SGLT2—A Review of Literature. Curr. Probl. Cardiol. 2020, 46, 100765. [Google Scholar] [CrossRef] [PubMed]
- Zodda, D.; Giammona, R.; Schifilliti, S. Treatment Strategy for Dyslipidemia in Cardiovascular Disease Prevention: Focus on Old and New Drugs. Pharmacy 2018, 6, 10. [Google Scholar] [CrossRef] [PubMed]
- Bouitbir, J.; Sanvee, G.M.; Panajatovic, M.V.; Singh, F.; Krähenbühl, S. Mechanisms of statin-associated skeletal muscle-associated symptoms. Pharmacol. Res. 2019, 154, 104201. [Google Scholar] [CrossRef] [PubMed]
- Apostolopoulou, M.; Corsini, A.; Roden, M. The role of mitochondria in statin-induced myopathy. Eur. J. Clin. Investig. 2015, 45, 745–754. [Google Scholar] [CrossRef] [PubMed]
- Mallinson, J.E.; Constantin-Teodosiu, D.; Sidaway, J.; Westwood, F.R.; Greenhaff, P. Blunted Akt/FOXO signalling and activation of genes controlling atrophy and fuel use in statin myopathy. J. Physiol. 2009, 587, 219–230. [Google Scholar] [CrossRef] [PubMed]
- Hanai, J.-I.; Cao, P.; Tanksale, P.; Imamura, S.; Koshimizu, E.; Zhao, J.; Kishi, S.; Yamashita, M.; Phillips, P.S.; Sukhatme, V.P.; et al. The muscle-specific ubiquitin ligase atrogin-1/MAFbx mediates statin-induced muscle toxicity. J. Clin. Investig. 2007, 117, 3940–3951. [Google Scholar] [CrossRef]
- Jaśkiewicz, A.; Pająk, B.; Łabieniec-Watała, M.; De Palma, C.; Orzechowski, A. Diverse Action of Selected Statins on Skeletal Muscle Cells—An Attempt to Explain the Protective Effect of Geranylgeraniol (GGOH) in Statin-Associated Myopathy (SAM). J. Clin. Med. 2019, 8, 694. [Google Scholar] [CrossRef]
- Reijneveld, J.C.; Koot, R.W.; Bredman, J.J.; A Joles, J.; Bär, P.R. Differential Effects of 3-Hydroxy-3-methylglutaryl-Coenzyme A Reductase Inhibitors on the Development of Myopathy in Young Rats. Pediatr. Res. 1996, 39, 1028–1035. [Google Scholar] [CrossRef]
- DiNicolantonio, J.J. CoQ10 and L-carnitine for statin myalgia? Expert Rev. Cardiovasc. Ther. 2012, 10, 1329–1333. [Google Scholar] [CrossRef]
- Pasha, R.; Moon, T.W. Coenzyme Q10 protects against statin-induced myotoxicity in zebrafish larvae ( Danio rerio ). Environ. Toxicol. Pharmacol. 2017, 52, 150–160. [Google Scholar] [CrossRef] [PubMed]
- Vladutiu, G.D.; Isackson, P.J.; Kaufman, K.; Harley, J.B.; Cobb, B.; Christopher-Stine, L.; Wortmann, R.L. Genetic risk for malignant hyperthermia in non-anesthesia-induced myopathies. Mol. Genet. Metab. 2011, 104, 167–173. [Google Scholar] [CrossRef] [PubMed]
- La Guardia, P.G.; Alberici, L.C.; Ravagnani, F.G.; Catharino, R.R.; Vercesi, A.E. Protection of rat skeletal muscle fibers by either L-carnitine or coenzyme Q10 against statins toxicity mediated by mitochondrial reactive oxygen generation. Front. Physiol. 2013, 4, 103. [Google Scholar] [CrossRef] [PubMed]
- Durazzo, A.; Lucarini, M.; Nazhand, A.; Souto, S.B.; Silva, A.M.; Severino, P.; Souto, E.B.; Santini, A. The Nutraceutical Value of Carnitine and Its Use in Dietary Supplements. Molecules 2020, 25, 2127. [Google Scholar] [CrossRef]
- Sharma, S.; Black, S.M. Carnitine homeostasis, mitochondrial function and cardiovascular disease. Drug Discov. Today Dis. Mech. 2009, 6, e31–e39. [Google Scholar] [CrossRef]
- Longo, N.; di San Filippo, C.A.; Pasquali, M. Disorders of carnitine transport and the carnitine cycle. Am. J. Med. Genet. Part C Semin. Med. Genet. 2006, 142C, 77–85. [Google Scholar] [CrossRef]
- Guzel, N.A.; Orer, G.E.; Bircan, F.S.; Cevher, C. Effects of acute L-carnitine supplementation on nitric oxide production and oxidative stress after exhaustive exercise in young soccer players. J. Sports Med. Phys. Fit. 2014, 55, 9–15. [Google Scholar]
- Kim, J.H.; Pan, J.H.; Lee, E.S.; Kim, Y.J. l -Carnitine enhances exercise endurance capacity by promoting muscle oxidative metabolism in mice. Biochem. Biophys. Res. Commun. 2015, 464, 568–573. [Google Scholar] [CrossRef]
- Koozehchian, M.S.; Daneshfar, A.; Fallah, E.; Agha-Alinejad, H.; Samadi, M.; Kaviani, M.; Kaveh, B.M.; Jung, Y.P.; Sablouei, M.H.; Moradi, N.; et al. Effects of nine weeks L-Carnitine supplementation on exercise performance, anaerobic power, and exercise-induced oxidative stress in resistance-trained males. J. Exerc. Nutr. Biochem. 2018, 22, 7–19. [Google Scholar] [CrossRef]
- Al-Eisa, R.A.; Al-Salmi, F.A.; Hamza, R.Z.; El-Shenawy, N.S. Role of L-carnitine in protection against the cardiac oxidative stress induced by aspartame in Wistar albino rats. PLoS ONE 2018, 13, e0204913. [Google Scholar] [CrossRef]
- Jang, J.; Park, J.; Chang, H.; Lim, K. l-Carnitine supplement reduces skeletal muscle atrophy induced by prolonged hindlimb suspension in rats. Appl. Physiol. Nutr. Metab. 2016, 41, 1240–1247. [Google Scholar] [CrossRef] [PubMed]
- Dubińska-Magiera, M.; Daczewska, M.; Lewicka, A.; Migocka-Patrzałek, M.; Niedbalska-Tarnowska, J.; Jagla, K. Zebrafish: A Model for the Study of Toxicants Affecting Muscle Development and Function. Int. J. Mol. Sci. 2016, 17, 1941. [Google Scholar] [CrossRef] [PubMed]
- Plantié, E.; Migocka-Patrzałek, M.; Daczewska, M.; Jagla, K. Model Organisms in the Fight against Muscular Dystrophy: Lessons from Drosophila and Zebrafish. Molecules 2015, 20, 6237–6253. [Google Scholar] [CrossRef] [PubMed]
- Bandaru, M.K.; Emmanouilidou, A.; Ranefall, P.; von der Heyde, B.; Mazzaferro, E.; Klingström, T.; Masiero, M.; Dethlefsen, O.; Ledin, J.; Larsson, A. Zebrafish larvae as a model system for systematic characterization of drugs and genes in dyslipidemia and atherosclerosis. bioRxiv 2019, 502674. [Google Scholar] [CrossRef]
- Campos, L.M.; Rios, E.A.; Midlej, V.D.V.P.; Atella, G.C.; Herculano-Houzel, S.; Benchimol, M.; Mermelstein, C.; Costa, M.L. Structural Analysis of Alterations in Zebrafish Muscle Differentiation Induced by Simvastatin and Their Recovery with Cholesterol. J. Histochem. Cytochem. 2015, 63, 427–437. [Google Scholar] [CrossRef]
- Campos, L.M.; A Rios, E.; Guapyassu, L.; Midlej, V.D.V.P.; Atella, G.C.; Herculano-Houzel, S.; Benchimol, M.; Mermelstein, C.; Costa, M.L. Alterations in zebrafish development induced by simvastatin: Comprehensive morphological and physiological study, focusing on muscle. Exp. Biol. Med. 2016, 241, 1950–1960. [Google Scholar] [CrossRef] [PubMed]
- Cao, P.; Hanai, J.-I.; Tanksale, P.; Imamura, S.; Sukhatme, V.P.; Lecker, S.H. Statin-induced muscle damage and atrogin-1 induction is the result of a geranylgeranylation defect. FASEB J. 2009, 23, 2844–2854. [Google Scholar] [CrossRef]
- Hoppstädter, J.; Perez, J.V.V.; Linnenberger, R.; Dahlem, C.; Legroux, T.M.; Hecksteden, A.; Tse, K.F.W.; Flamini, S.; Andreas, A.; Herrmann, J.; et al. The glucocorticoid-induced leucine zipper mediates statin-induced muscle damage. FASEB J. 2020, 34, 4684–4701. [Google Scholar] [CrossRef]
- Ribeiro, S.; Torres, T.; Martins, R.; Santos, M.M. Toxicity screening of Diclofenac, Propranolol, Sertraline and Simvastatin using Danio rerio and Paracentrotus lividus embryo bioassays. Ecotoxicol. Environ. Saf. 2015, 114, 67–74. [Google Scholar] [CrossRef]
- Dubińska-Magiera, M.; Migocka-Patrzałek, M.; Lewandowski, D.; Daczewska, M.; Jagla, K. Zebrafish as a Model for the Study of Lipid-Lowering Drug-Induced Myopathies. Int. J. Mol. Sci. 2021, 22, 5654. [Google Scholar] [CrossRef]
- Thorpe, J.L.; Doitsidou, M.; Ho, S.-Y.; Raz, E.; A Farber, S. Germ Cell Migration in Zebrafish Is Dependent on HMGCoA Reductase Activity and Prenylation. Dev. Cell 2004, 6, 295–302. [Google Scholar] [CrossRef]
- Bruscoli, S.; Donato, V.; Velardi, E.; Di Sante, M.; Migliorati, G.; Donato, R.; Riccardi, C. Glucocorticoid-induced Leucine Zipper (GILZ) and Long GILZ Inhibit Myogenic Differentiation and Mediate Anti-myogenic Effects of Glucocorticoids. J. Biol. Chem. 2010, 285, 10385–10396. [Google Scholar] [CrossRef] [PubMed]
- Maerz, L.D.; Burkhalter, M.; Schilpp, C.; Wittekindt, O.H.; Frick, M.; Philipp, M. Pharmacological cholesterol depletion disturbs ciliogenesis and ciliary function in developing zebrafish. Commun. Biol. 2019, 2, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Jao, L.-E.; Appel, B.; Wente, S.R. A zebrafish model of lethal congenital contracture syndrome 1 reveals Gle1 function in spinal neural precursor survival and motor axon arborization. Development 2012, 139, 1316–1326. [Google Scholar] [CrossRef] [PubMed]
- Aleström, P.; D’Angelo, L.; Midtlyng, P.J.; Schorderet, D.F.; Schulte-Merker, S.; Sohm, F.; Warner, S. Zebrafish: Housing and husbandry recommendations. Lab. Anim. 2019, 54, 213–224. [Google Scholar] [CrossRef]
- Kimmel, C.B.; Ballard, W.W.; Kimmel, S.R.; Ullmann, B.; Schilling, T.F. Stages of embryonic development of the zebrafish. Dev. Dyn. 1995, 203, 253–310. [Google Scholar] [CrossRef]
- Robinson, B.L.; Dumas, M.; Paule, M.G.; Ali, S.F.; Kanungo, J. Opposing effects of ketamine and acetyl l-carnitine on the serotonergic system of zebrafish. Neurosci. Lett. 2015, 607, 17–22. [Google Scholar] [CrossRef][Green Version]
- Dubińska-Magiera, M.; Niedbalska-Tarnowska, J.; Migocka-Patrzałek, M.; Posyniak, E.; Daczewska, M. Characterization of Hspb8 in Zebrafish. Cells 2020, 9, 1562. [Google Scholar] [CrossRef]
- Xi, Y.; Ryan, J.; Noble, S.; Yu, M.; Yilbas, A.E.; Ekker, M. Impaired dopaminergic neuron development and locomotor function in zebrafish with loss of pink1 function. Eur. J. Neurosci. 2010, 31, 623–633. [Google Scholar] [CrossRef]
- Granato, M.; van Eeden, F.; Schach, U.; Trowe, T.; Brand, M.; Furutani-Seiki, M.; Haffter, P.; Hammerschmidt, M.; Heisenberg, C.; Jiang, Y.; et al. Genes controlling and mediating locomotion behavior of the zebrafish embryo and larva. Development 1996, 123, 399–413. [Google Scholar] [CrossRef]
- V3 Plugin. Available online: https://imagej.nih.gov/ij/plugins/time-series.html (accessed on 1 February 2022).
- Smith, L.L.; Beggs, A.H.; Gupta, V.A. Analysis of skeletal muscle defects in larval zebrafish by birefringence and touch-evoke escape response assays. J. Vis. Exp. 2013, e50925. [Google Scholar] [CrossRef] [PubMed]
- Haschek, W.M.; Rousseaux, C.G.; Wallig, M.A.; Bolon, B.; Ochoa, R. Haschek and Rousseaux’s Handbook of Toxicologic Pathology; Academic Press: Cambridge, MA, USA, 2013. [Google Scholar]
- Kristofco, L.A.; Cruz, L.C.; Haddad, S.P.; Behra, M.L.; Chambliss, C.K.; Brooks, B.W. Age matters: Developmental stage of Danio rerio larvae influences photomotor response thresholds to diazinion or diphenhydramine. Aquat. Toxicol. 2015, 170, 344–354. [Google Scholar] [CrossRef] [PubMed]
- Martinez, R.; Vera-Chang, M.; Haddad, M.; Zon, J.; Navarro-Martin, L.; Trudeau, V.L.; Mennigen, J.A. Developmental fluoxetine exposure in zebrafish reduces offspring basal cortisol concentration via life stage-dependent maternal transmission. PLoS ONE 2019, 14, e0212577. [Google Scholar] [CrossRef] [PubMed]
- Brown, D.R.; Samsa, L.A.; Qian, L.; Liu, J. Advances in the Study of Heart Development and Disease Using Zebrafish. J. Cardiovasc. Dev. Dis. 2016, 3, 13. [Google Scholar] [CrossRef]
- Volek, J.S.; Kraemer, W.J.; Rubin, M.R.; Gómez, A.L.; Ratamess, N.A.; Gaynor, P. l-Carnitinel-tartrate supplementation favorably affects markers of recovery from exercise stress. Am. J. Physiol. Metab. 2002, 282, E474–E482. [Google Scholar] [CrossRef]
- A Spiering, B.; Kraemer, W.J.; Hatfield, D.L.; Vingren, J.L.; Fragala, M.S.; Ho, J.-Y.; A Thomas, G.; Häkkinen, K.; Volek, J.S. Effects of L-Carnitine L-Tartrate Supplementation on Muscle Oxygenation Responses to Resistance Exercise. J. Strength Cond. Res. 2008, 22, 1130–1135. [Google Scholar] [CrossRef]
- Fielding, R.; Riede, L.; Lugo, J.P.; Bellamine, A. l-Carnitine Supplementation in Recovery after Exercise. Nutrients 2018, 10, 349. [Google Scholar] [CrossRef]
- Kraemer, W.J.; Volek, J.S.; French, D.N.; Rubin, M.R.; Sharman, M.J.; Gómez, A.L.; Ratamess, N.A.; Newton, R.U.; Jemiolo, B.; Craig, B.W. The effects of L-carnitine L-tartrate supplementation on hormonal responses to resistance exercise and recovery. J. Strength Cond. Res. 2003, 17, 455–462. [Google Scholar]
- Nakahara, K.; Kuriyama, M.; Sonoda, Y.; Yoshidome, H.; Nakagawa, H.; Fujiyama, J.; Higuchi, I.; Osame, M. Myopathy Induced by HMG–CoA Reductase Inhibitors in Rabbits: A Pathological, Electrophysiological, and Biochemical Study. Toxicol. Appl. Pharmacol. 1998, 152, 99–106. [Google Scholar] [CrossRef]
- Dubińska-Magiera, M.; Migocka-Patrzałek, M.; Cegłowska, A. Danio adventure. Developmental biology of the zebrafish in science popularisation. J. Biol. Educ. 2020, 1–11. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target Gene | Accession Number | Seq F (Forward Primer) | Seq R (Reverse Primer) |
|---|---|---|---|
| rpl13a | NM_212784.1 | CGCTATTGTGGCCAAGCAAG | TCTTGCGGAGGAAAGCCAAA |
| atrogin-1 | NM_200917.1 | AAGCTCTGCCAGTATCACTTC | AGTGCAAGGATGGTCTGTATC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Niedbalska-Tarnowska, J.; Ochenkowska, K.; Migocka-Patrzałek, M.; Dubińska-Magiera, M. Assessment of the Preventive Effect of L-carnitine on Post-statin Muscle Damage in a Zebrafish Model. Cells 2022, 11, 1297. https://doi.org/10.3390/cells11081297
Niedbalska-Tarnowska J, Ochenkowska K, Migocka-Patrzałek M, Dubińska-Magiera M. Assessment of the Preventive Effect of L-carnitine on Post-statin Muscle Damage in a Zebrafish Model. Cells. 2022; 11(8):1297. https://doi.org/10.3390/cells11081297
Chicago/Turabian StyleNiedbalska-Tarnowska, Joanna, Katarzyna Ochenkowska, Marta Migocka-Patrzałek, and Magda Dubińska-Magiera. 2022. "Assessment of the Preventive Effect of L-carnitine on Post-statin Muscle Damage in a Zebrafish Model" Cells 11, no. 8: 1297. https://doi.org/10.3390/cells11081297
APA StyleNiedbalska-Tarnowska, J., Ochenkowska, K., Migocka-Patrzałek, M., & Dubińska-Magiera, M. (2022). Assessment of the Preventive Effect of L-carnitine on Post-statin Muscle Damage in a Zebrafish Model. Cells, 11(8), 1297. https://doi.org/10.3390/cells11081297