
The Relevance of the Bacterial Microbiome, Archaeome and Mycobiome in Pediatric Asthma and Respiratory Disorders

 ,
, 
 , and
, and
Abstract
:1. Introduction
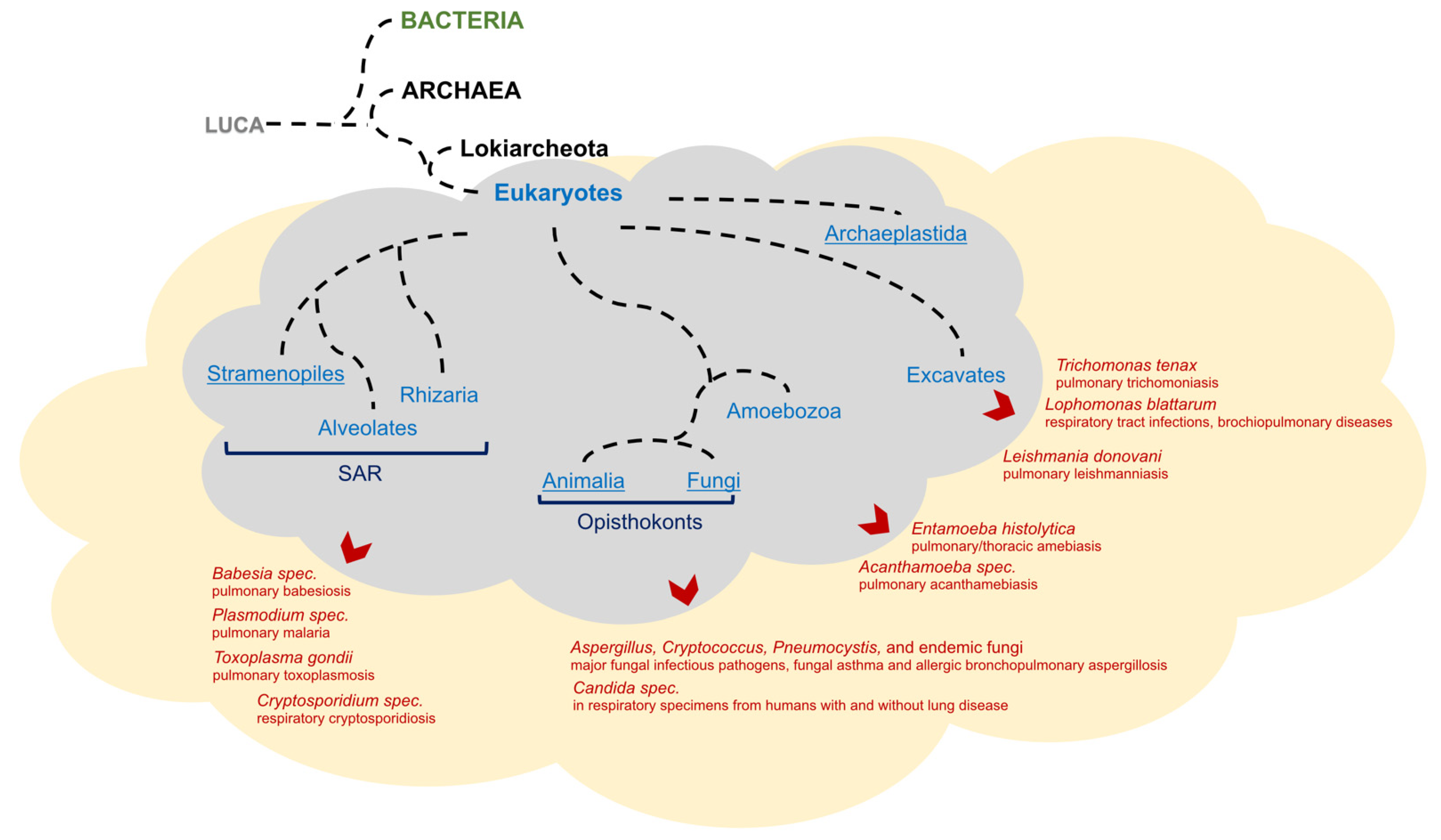
2. A Brief Evolutionary History of the Domains of Cellular Life
- THE MICROBE is so very small
- You cannot make him out at all,
- But many sanguine people hope
- To see him through a microscope.
- His jointed tongue that lies beneath
- A hundred curious rows of teeth;
- His seven tufted tails with lots
- Of lovely pink and purple spots,
- On each of which a pattern stands,
- Composed of forty separate bands;
- His eyebrows of a tender green;
- All these have never yet been seen-
- But Scientists, who ought to know,
- Assure us that is must be so...
- Oh! let us never, never doubt
- What nobody is sure about!
- ‘The Microbe’ from More Beasts for Worse Children.
- Hilaire Belloc. Duckworth, 1897 [10].
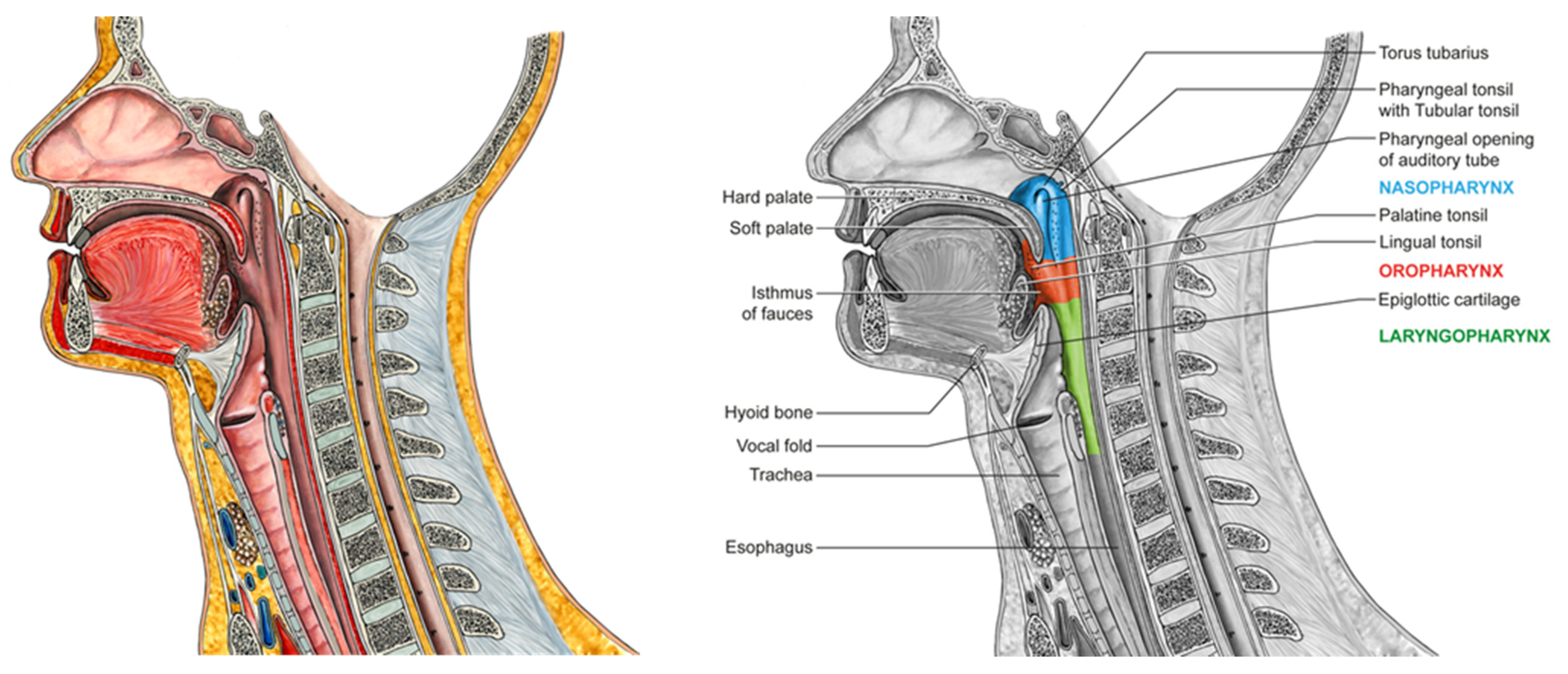
3. The Habitat: Anatomy and Physiology of the Naso- and Oropharynx
4. The Detection of Pathogens in the Naso- and Oropharynx and Considerations Regarding Respiratory Diseases during Early Childhood
4.1. The Bacterial Microbiome of the Naso- and Oropharynx in Connection with the Development of Pediatric Respiratory Diseases
4.2. The Importance of the Archaeome in the Human Body
4.3. The Variety of the Mycobiome
5. Clinical Translation of the Findings Regarding the Microbiome
5.1. The Bacterial Microbiome in Wheezing Children
5.2. Bronchiolitis in Children and the Relation to Bacterial Microbiota in the Upper Respiratory Tract
5.3. Findings in the Microbiome of Nasopharynx in Relation to the Development of Childhood Asthma
5.4. Association of Allergy in Childhood with Microbial Colonization of the Nasopharynx
5.5. Archaea as a Human Pathogenic Factor in Periodontal Disease
5.6. Chronic Rhinosinusitis in Connection with the Archaeome and Fungiome
6. Future Outlook
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bruls, T.; Weissenbach, J. The human metagenome: Our other genome? Hum. Mol. Genet. 2011, 20, R142–R148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caporaso, J.G.; Lauber, C.L.; Costello, E.K.; Berg-Lyons, D.; Gonzalez, A.; Stombaugh, J.; Knights, D.; Gajer, P.; Ravel, J.; Fierer, N.; et al. Moving pictures of the human microbiome. Genome Biol. 2011, 12, R50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qin, J.; Li, R.; Raes, J.; Arumugam, M.; Burgdorf, K.S.; Manichanh, C.; Nielsen, T.; Pons, N.; Levenez, F.; Yamada, T.; et al. A human gut microbial gene catalogue established by metagenomic sequencing. Nature 2010, 464, 59–65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wen, Z.; Xie, G.; Zhou, Q.; Qiu, C.; Li, J.; Hu, Q.; Dai, W.; Li, D.; Zheng, Y.; Wen, F. Distinct Nasopharyngeal and Oropharyngeal Microbiota of Children with Influenza A Virus Compared with Healthy Children. Biomed. Res. Int. 2018, 2018, 6362716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mac Aogain, M.; Lau, K.J.X.; Cai, Z.; Kumar Narayana, J.; Purbojati, R.W.; Drautz-Moses, D.I.; Gaultier, N.E.; Jaggi, T.K.; Tiew, P.Y.; Ong, T.H.; et al. Metagenomics Reveals a Core Macrolide Resistome Related to Microbiota in Chronic Respiratory Disease. Am. J. Respir. Crit. Care Med. 2020, 202, 433–447. [Google Scholar] [CrossRef]
- O’Dwyer, D.N.; Ashley, S.L.; Gurczynski, S.J.; Xia, M.; Wilke, C.; Falkowski, N.R.; Norman, K.C.; Arnold, K.B.; Huffnagle, G.B.; Salisbury, M.L.; et al. Lung Microbiota Contribute to Pulmonary Inflammation and Disease Progression in Pulmonary Fibrosis. Am. J. Respir. Crit. Care Med. 2019, 199, 1127–1138. [Google Scholar] [CrossRef]
- Yeoh, Y.K.; Zuo, T.; Lui, G.C.; Zhang, F.; Liu, Q.; Li, A.Y.; Chung, A.C.; Cheung, C.P.; Tso, E.Y.; Fung, K.S.; et al. Gut microbiota composition reflects disease severity and dysfunctional immune responses in patients with COVID-19. Gut 2021, 70, 698–706. [Google Scholar] [CrossRef]
- Kadioglu, A.; Weiser, J.N.; Paton, J.C.; Andrew, P.W. The role of Streptococcus pneumoniae virulence factors in host respiratory colonization and disease. Nat. Rev. Microbiol. 2008, 6, 288–301. [Google Scholar] [CrossRef]
- Young, V.B. The role of the microbiome in human health and disease: An introduction for clinicians. BMJ 2017, 356, j831. [Google Scholar] [CrossRef]
- Belloc, H. 1870–1953; B.T.B. (Basil Temple Blackwood), 1870–1917. More beasts (for worse children). Collection: Kellylibrary; Toronto, Publisher: London: E. Arnold. 1897. Available online: https://archive.org/details/morebeastsforwor00belluoft (accessed on 15 January 2022).
- Mora, C.; Tittensor, D.P.; Adl, S.; Simpson, A.G.; Worm, B. How many species are there on Earth and in the ocean? PLoS Biol. 2011, 9, e1001127. [Google Scholar] [CrossRef] [Green Version]
- Louca, S.; Mazel, F.; Doebeli, M.; Parfrey, L.W. A census-based estimate of Earth’s bacterial and archaeal diversity. PLoS Biol. 2019, 17, e3000106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wiens, J.J. Vast (but avoidable) underestimation of global biodiversity. PLoS Biol. 2021, 19, e3001192. [Google Scholar] [CrossRef]
- Williams, T.A.; Foster, P.G.; Cox, C.J.; Embley, T.M. An archaeal origin of eukaryotes supports only two primary domains of life. Nature 2013, 504, 231–236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zaremba-Niedzwiedzka, K.; Caceres, E.F.; Saw, J.H.; Backstrom, D.; Juzokaite, L.; Vancaester, E.; Seitz, K.W.; Anantharaman, K.; Starnawski, P.; Kjeldsen, K.U.; et al. Asgard archaea illuminate the origin of eukaryotic cellular complexity. Nature 2017, 541, 353–358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van de Pol, J.A.; van Best, N.; Mbakwa, C.A.; Thijs, C.; Savelkoul, P.H.; Arts, I.C.; Hornef, M.W.; Mommers, M.; Penders, J. Gut Colonization by Methanogenic Archaea Is Associated with Organic Dairy Consumption in Children. Front. Microbiol. 2017, 8, 355. [Google Scholar] [CrossRef] [PubMed]
- Bowman, D.D. Georgis’ Parasitology for Veterinarians, 10th ed.; Saunders, Ed.; Elsevier: Amsterdam, The Netherlands, 2014. [Google Scholar]
- Burki, F. The eukaryotic tree of life from a global phylogenomic perspective. Cold Spring Harb. Perspect. Biol. 2014, 6, a016147. [Google Scholar] [CrossRef]
- Vijayan, V.K.; Kilani, T. Emerging and established parasitic lung infestations. Infect. Dis. Clin. N. Am. 2010, 24, 579–602. [Google Scholar] [CrossRef]
- Sponseller, J.K.; Griffiths, J.K.; Tzipori, S. The evolution of respiratory Cryptosporidiosis: Evidence for transmission by inhalation. Clin. Microbiol. Rev. 2014, 27, 575–586. [Google Scholar] [CrossRef] [Green Version]
- Moss, R.B. Treatment options in severe fungal asthma and allergic bronchopulmonary aspergillosis. Eur. Respir. J. 2014, 43, 1487–1500. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Lu, G.; Meng, G. Pathogenic Fungal Infection in the Lung. Front. Immunol. 2019, 10, 1524. [Google Scholar] [CrossRef] [Green Version]
- Pendleton, K.M.; Huffnagle, G.B.; Dickson, R.P. The significance of Candida in the human respiratory tract: Our evolving understanding. Pathog. Dis. 2017, 75, ftx029. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shamsuzzaman, S.M.; Hashiguchi, Y. Thoracic amebiasis. Clin. Chest Med. 2002, 23, 479–492. [Google Scholar] [CrossRef]
- Visvesvara, G.S.; Moura, H.; Schuster, F.L. Pathogenic and opportunistic free-living amoebae: Acanthamoeba spp., Balamuthia mandrillaris, Naegleria fowleri, and Sappinia diploidea. FEMS Immunol. Med. Microbiol. 2007, 50, 1–26. [Google Scholar] [CrossRef] [Green Version]
- Chaudhury, A.; Parija, S.C. Lophomonas blattarum: A new flagellate causing respiratory tract infections. Trop. Parasitol. 2020, 10, 7–11. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Giron, R.; Cornelis van Woerden, H. Lophomonas blattarum and bronchopulmonary disease. J. Med. Microbiol. 2013, 62, 1641–1648. [Google Scholar] [CrossRef]
- Hersh, S.M. Pulmonary trichomoniasis and Trichomonas tenax. J. Med. Microbiol. 1985, 20, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Merati, A.L.; Rieder, A.A. Normal endoscopic anatomy of the pharynx and larynx. Am. J. Med. 2003, 115 (Suppl. 3A), 10S–14S. [Google Scholar] [CrossRef]
- Paulsen, F.; Waschke, J. Sobotta-Atlas der Anatomie des Menschen, Allgemeine Anatomie, Bewegungsapparat, Innere Organe, Kopf, Hals, Neuroanatomie, 3 Volumes and Table Booklet in a Slipcase, 23rd ed. Publisher Elsevier: Munich, Germany, 2010.
- Mankowski, N.L.; Bordoni, B. Anatomy, Head and Neck, Nasopharynx. 2021 Aug 11; StatPearls: Treasure Island, FL, USA, 2022. [Google Scholar]
- Fossum, C.C.; Chintakuntlawar, A.V.; Price, D.L.; Garcia, J.J. Characterization of the oropharynx: Anatomy, histology, immunology, squamous cell carcinoma and surgical resection. Histopathology 2017, 70, 1021–1029. [Google Scholar] [CrossRef] [PubMed]
- Schünke, M.; Schulte, E.; Schumacher, U.; Voll, M.; Wesker, K. Prometheus LernAtlas der Anatomie—Kopf, Hals und Neuroanatomie; Hrsg. 4. Auflage; Thieme: Stuttgart, Germany, 2015. [Google Scholar] [CrossRef]
- Lüllmann-Rauch, R.; Asan, E. Taschenlehrbuch Histologie, 5th ed.; Thieme: Stuttgart, Germany, 2015. [Google Scholar]
- Beule, A.G. The Microbiome—The Unscheduled Parameter for Future Therapies. Laryngorhinootologie 2018, 97, S279–S311. [Google Scholar] [CrossRef] [Green Version]
- Boutin, S.; Depner, M.; Stahl, M.; Graeber, S.Y.; Dittrich, S.A.; Legatzki, A.; von Mutius, E.; Mall, M.; Dalpke, A.H. Comparison of Oropharyngeal Microbiota from Children with Asthma and Cystic Fibrosis. Mediat. Inflamm. 2017, 2017, 5047403. [Google Scholar] [CrossRef] [Green Version]
- Cardenas, P.A.; Cooper, P.J.; Cox, M.J.; Chico, M.; Arias, C.; Moffatt, M.F.; Cookson, W.O. Upper airways microbiota in antibiotic-naive wheezing and healthy infants from the tropics of rural Ecuador. PLoS ONE 2012, 7, e46803. [Google Scholar] [CrossRef] [PubMed]
- Cuthbertson, L.; Oo, S.W.C.; Cox, M.J.; Khoo, S.K.; Cox, D.W.; Chidlow, G.; Franks, K.; Prastanti, F.; Borland, M.L.; Gern, J.E.; et al. Viral respiratory infections and the oropharyngeal bacterial microbiota in acutely wheezing children. PLoS ONE 2019, 14, e0223990. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, Q.; Dai, W.; Zhou, Q.; Fu, D.; Zheng, Y.; Wang, W.; Liu, Y.; Yang, Q.; Dai, D.; Liu, S.; et al. Dynamic oropharyngeal and faecal microbiota during treatment in infants hospitalized for bronchiolitis compared with age-matched healthy subjects. Sci. Rep. 2017, 7, 11266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Powell, E.A.; Fontanella, S.; Boakes, E.; Belgrave, D.; Shaw, A.G.; Cornwell, E.; Fernandez-Crespo, R.; Fink, C.G.; Custovic, A.; Kroll, J.S. Temporal association of the development of oropharyngeal microbiota with early life wheeze in a population-based birth cohort. EBioMedicine 2019, 46, 486–498. [Google Scholar] [CrossRef] [Green Version]
- Kelly, M.S.; Surette, M.G.; Smieja, M.; Pernica, J.M.; Rossi, L.; Luinstra, K.; Steenhoff, A.P.; Feemster, K.A.; Goldfarb, D.M.; Arscott-Mills, T.; et al. The Nasopharyngeal Microbiota of Children With Respiratory Infections in Botswana. Pediatr. Infect. Dis. J. 2017, 36, e211–e218. [Google Scholar] [CrossRef]
- Perez, G.F.; Perez-Losada, M.; Isaza, N.; Rose, M.C.; Colberg-Poley, A.M.; Nino, G. Nasopharyngeal microbiome in premature infants and stability during rhinovirus infection. J. Investig. Med. 2017, 65, 984–990. [Google Scholar] [CrossRef] [Green Version]
- Teo, S.M.; Mok, D.; Pham, K.; Kusel, M.; Serralha, M.; Troy, N.; Holt, B.J.; Hales, B.J.; Walker, M.L.; Hollams, E.; et al. The infant nasopharyngeal microbiome impacts severity of lower respiratory infection and risk of asthma development. Cell Host Microbe 2015, 17, 704–715. [Google Scholar] [CrossRef] [Green Version]
- Verhagen, L.M.; Rivera-Olivero, I.A.; Clerc, M.; Chu, M.; van Engelsdorp Gastelaars, J.; Kristensen, M.I.; Berbers, G.A.M.; Hermans, P.W.M.; de Jonge, M.I.; de Waard, J.H.; et al. Nasopharyngeal Microbiota Profiles in Rural Venezuelan Children Are Associated With Respiratory and Gastrointestinal Infections. Clin. Infect. Dis. 2021, 72, 212–221. [Google Scholar] [CrossRef] [Green Version]
- Dumas, O.; Hasegawa, K.; Mansbach, J.M.; Sullivan, A.F.; Piedra, P.A.; Camargo, C.A., Jr. Severe bronchiolitis profiles and risk of recurrent wheeze by age 3 years. J. Allergy Clin. Immunol. 2019, 143, 1371–1379 e1377. [Google Scholar] [CrossRef] [Green Version]
- Luna, P.N.; Hasegawa, K.; Ajami, N.J.; Espinola, J.A.; Henke, D.M.; Petrosino, J.F.; Piedra, P.A.; Sullivan, A.F.; Camargo, C.A., Jr.; Shaw, C.A.; et al. The association between anterior nares and nasopharyngeal microbiota in infants hospitalized for bronchiolitis. Microbiome 2018, 6, 2. [Google Scholar] [CrossRef]
- Rosas-Salazar, C.; Shilts, M.H.; Tovchigrechko, A.; Schobel, S.; Chappell, J.D.; Larkin, E.K.; Gebretsadik, T.; Halpin, R.A.; Nelson, K.E.; Moore, M.L.; et al. Nasopharyngeal Lactobacillus is associated with a reduced risk of childhood wheezing illnesses following acute respiratory syncytial virus infection in infancy. J. Allergy Clin. Immunol. 2018, 142, 1447–1456 e1449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Man, W.H.; Clerc, M.; de Steenhuijsen Piters, W.A.A.; van Houten, M.A.; Chu, M.; Kool, J.; Keijser, B.J.F.; Sanders, E.A.M.; Bogaert, D. Loss of Microbial Topography between Oral and Nasopharyngeal Microbiota and Development of Respiratory Infections Early in Life. Am. J. Respir. Crit. Care Med. 2019, 200, 760–770. [Google Scholar] [CrossRef] [PubMed]
- Tang, H.H.F.; Lang, A.; Teo, S.M.; Judd, L.M.; Gangnon, R.; Evans, M.D.; Lee, K.E.; Vrtis, R.; Holt, P.G.; Lemanske, R.F., Jr.; et al. Developmental patterns in the nasopharyngeal microbiome during infancy are associated with asthma risk. J. Allergy Clin. Immunol. 2021, 147, 1683–1691. [Google Scholar] [CrossRef] [PubMed]
- Teo, S.M.; Tang, H.H.F.; Mok, D.; Judd, L.M.; Watts, S.C.; Pham, K.; Holt, B.J.; Kusel, M.; Serralha, M.; Troy, N.; et al. Airway Microbiota Dynamics Uncover a Critical Window for Interplay of Pathogenic Bacteria and Allergy in Childhood Respiratory Disease. Cell Host Microbe 2018, 24, 341–352 e345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rinke, C.; Schwientek, P.; Sczyrba, A.; Ivanova, N.N.; Anderson, I.J.; Cheng, J.F.; Darling, A.; Malfatti, S.; Swan, B.K.; Gies, E.A.; et al. Insights into the phylogeny and coding potential of microbial dark matter. Nature 2013, 499, 431–437. [Google Scholar] [CrossRef] [Green Version]
- Woese, C.R.; Kandler, O.; Wheelis, M.L. Towards a natural system of organisms: Proposal for the domains Archaea, Bacteria, and Eucarya. Proc. Natl. Acad. Sci. USA 1990, 87, 4576–4579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schleifer, K.H. Classification of Bacteria and Archaea: Past, present and future. Syst. Appl. Microbiol. 2009, 32, 533–542. [Google Scholar] [CrossRef]
- Woese, C.R.; Fox, G.E. Phylogenetic structure of the prokaryotic domain: The primary kingdoms. Proc. Natl. Acad. Sci. USA 1977, 74, 5088–5090. [Google Scholar] [CrossRef] [Green Version]
- Eme, L.; Spang, A.; Lombard, J.; Stairs, C.W.; Ettema, T.J.G. Archaea and the origin of eukaryotes. Nat. Rev. Microbiol. 2017, 15, 711–723. [Google Scholar] [CrossRef]
- Cavicchioli, R. Archaea--timeline of the third domain. Nat. Rev. Microbiol. 2011, 9, 51–61. [Google Scholar] [CrossRef]
- Baker, B.J.; De Anda, V.; Seitz, K.W.; Dombrowski, N.; Santoro, A.E.; Lloyd, K.G. Diversity, ecology and evolution of Archaea. Nat. Microbiol. 2020, 5, 887–900. [Google Scholar] [CrossRef] [PubMed]
- Krzmarzick, M.J.; Taylor, D.K.; Fu, X.; McCutchan, A.L. Diversity and Niche of Archaea in Bioremediation. Archaea 2018, 2018, 3194108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cavicchioli, R.; Curmi, P.M.; Saunders, N.; Thomas, T. Pathogenic archaea: Do they exist? Bioessays 2003, 25, 1119–1128. [Google Scholar] [CrossRef] [PubMed]
- Bang, C.; Schmitz, R.A. Archaea associated with human surfaces: Not to be underestimated. FEMS Microbiol Rev. 2015, 39, 631–648. [Google Scholar] [CrossRef] [Green Version]
- de Cena, J.A.; Zhang, J.; Deng, D.; Dame-Teixeira, N.; Do, T. Low-Abundant Microorganisms: The Human Microbiome’s Dark Matter, a Scoping Review. Front. Cell. Infect. Microbiol. 2021, 11, 689197. [Google Scholar] [CrossRef]
- Pausan, M.R.; Csorba, C.; Singer, G.; Till, H.; Schopf, V.; Santigli, E.; Klug, B.; Hogenauer, C.; Blohs, M.; Moissl-Eichinger, C. Exploring the Archaeome: Detection of Archaeal Signatures in the Human Body. Front. Microbiol. 2019, 10, 2796. [Google Scholar] [CrossRef] [Green Version]
- Koskinen, K.; Pausan, M.R.; Perras, A.K.; Beck, M.; Bang, C.; Mora, M.; Schilhabel, A.; Schmitz, R.; Moissl-Eichinger, C. First Insights into the Diverse Human Archaeome: Specific Detection of Archaea in the Gastrointestinal Tract, Lung, and Nose and on Skin. mBio 2017, 8, e00824-17. [Google Scholar] [CrossRef] [Green Version]
- Horz, H.P.; Conrads, G. Methanogenic Archaea and oral infections—Ways to unravel the black box. J. Oral Microbiol. 2011, 3, 5940. [Google Scholar] [CrossRef] [Green Version]
- Miragoli, F.; Federici, S.; Ferrari, S.; Minuti, A.; Rebecchi, A.; Bruzzese, E.; Buccigrossi, V.; Guarino, A.; Callegari, M.L. Impact of cystic fibrosis disease on archaea and bacteria composition of gut microbiota. FEMS Microbiol. Ecol. 2017, 93, fiw230. [Google Scholar] [CrossRef] [Green Version]
- Wampach, L.; Heintz-Buschart, A.; Hogan, A.; Muller, E.E.L.; Narayanasamy, S.; Laczny, C.C.; Hugerth, L.W.; Bindl, L.; Bottu, J.; Andersson, A.F.; et al. Colonization and Succession within the Human Gut Microbiome by Archaea, Bacteria, and Microeukaryotes during the First Year of Life. Front. Microbiol. 2017, 8, 738. [Google Scholar] [CrossRef]
- Probst, A.J.; Auerbach, A.K.; Moissl-Eichinger, C. Archaea on human skin. PLoS ONE 2013, 8, e65388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Houshyar, Y.; Massimino, L.; Lamparelli, L.A.; Danese, S.; Ungaro, F. Going Beyond Bacteria: Uncovering the Role of Archaeome and Mycobiome in Inflammatory Bowel Disease. Front. Physiol. 2021, 12, 783295. [Google Scholar] [CrossRef] [PubMed]
- Diaz, P.I. Subgingival fungi, Archaea, and viruses under the omics loupe. Periodontol. 2000 2021, 85, 82–89. [Google Scholar] [CrossRef] [PubMed]
- Dridi, B.; Raoult, D.; Drancourt, M. Archaea as emerging organisms in complex human microbiomes. Anaerobe 2011, 17, 56–63. [Google Scholar] [CrossRef]
- Dabdoub, S.M.; Ganesan, S.M.; Kumar, P.S. Comparative metagenomics reveals taxonomically idiosyncratic yet functionally congruent communities in periodontitis. Sci. Rep. 2016, 6, 38993. [Google Scholar] [CrossRef]
- Matarazzo, F.; Ribeiro, A.C.; Faveri, M.; Taddei, C.; Martinez, M.B.; Mayer, M.P. The domain Archaea in human mucosal surfaces. Clin. Microbiol. Infect. 2012, 18, 834–840. [Google Scholar] [CrossRef] [Green Version]
- Vanessa Demonfort Nkamgaa, B.H. Michel Drancourta. Archaea: Essential inhabitants of the human digestive microbiota. Hum. Microbiome J. 2017, 3, 1–8. [Google Scholar] [CrossRef]
- Koskinen, K.; Reichert, J.L.; Hoier, S.; Schachenreiter, J.; Duller, S.; Moissl-Eichinger, C.; Schopf, V. The nasal microbiome mirrors and potentially shapes olfactory function. Sci. Rep. 2018, 8, 1296. [Google Scholar] [CrossRef] [Green Version]
- Choi, J.; Kim, S.H. A genome Tree of Life for the Fungi kingdom. Proc. Natl. Acad. Sci. USA 2017, 114, 9391–9396. [Google Scholar] [CrossRef] [Green Version]
- Spatafora, J.W.; Aime, M.C.; Grigoriev, I.V.; Martin, F.; Stajich, J.E.; Blackwell, M. The Fungal Tree of Life: From Molecular Systematics to Genome-Scale Phylogenies. Microbiol. Spectr. 2017, 5, 3–34. [Google Scholar] [CrossRef] [Green Version]
- Kohler, J.R.; Hube, B.; Puccia, R.; Casadevall, A.; Perfect, J.R. Fungi that Infect Humans. Microbiol. Spectr. 2017, 5, 813–843. [Google Scholar] [CrossRef] [PubMed]
- Schmitt, H.J.; Blevins, A.; Sobeck, K.; Armstrong, D. Aspergillus species from hospital air and from patients. Mycoses 1990, 33, 539–541. [Google Scholar] [CrossRef] [PubMed]
- Cui, L.; Morris, A.; Ghedin, E. The human mycobiome in health and disease. Genome Med. 2013, 5, 63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arrieta, M.C.; Arevalo, A.; Stiemsma, L.; Dimitriu, P.; Chico, M.E.; Loor, S.; Vaca, M.; Boutin, R.C.T.; Morien, E.; Jin, M.; et al. Associations between infant fungal and bacterial dysbiosis and childhood atopic wheeze in a nonindustrialized setting. J. Allergy Clin. Immunol. 2018, 142, 424–434 e410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujimura, K.E.; Sitarik, A.R.; Havstad, S.; Lin, D.L.; Levan, S.; Fadrosh, D.; Panzer, A.R.; LaMere, B.; Rackaityte, E.; Lukacs, N.W.; et al. Neonatal gut microbiota associates with childhood multisensitized atopy and T cell differentiation. Nat. Med. 2016, 22, 1187–1191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ward, T.L.; Dominguez-Bello, M.G.; Heisel, T.; Al-Ghalith, G.; Knights, D.; Gale, C.A. Development of the Human Mycobiome over the First Month of Life and across Body Sites. mSystems 2018, 3, e00140-17. [Google Scholar] [CrossRef] [Green Version]
- Ghannoum, M.A.; Jurevic, R.J.; Mukherjee, P.K.; Cui, F.; Sikaroodi, M.; Naqvi, A.; Gillevet, P.M. Characterization of the oral fungal microbiome (mycobiome) in healthy individuals. PLoS Pathog. 2010, 6, e1000713. [Google Scholar] [CrossRef] [Green Version]
- Dupuy, A.K.; David, M.S.; Li, L.; Heider, T.N.; Peterson, J.D.; Montano, E.A.; Dongari-Bagtzoglou, A.; Diaz, P.I.; Strausbaugh, L.D. Redefining the human oral mycobiome with improved practices in amplicon-based taxonomy: Discovery of Malassezia as a prominent commensal. PLoS ONE 2014, 9, e90899. [Google Scholar] [CrossRef] [Green Version]
- Tipton, L.; Ghedin, E.; Morris, A. The lung mycobiome in the next-generation sequencing era. Virulence 2017, 8, 334–341. [Google Scholar] [CrossRef]
- Porter, P.C.; Lim, D.J.; Maskatia, Z.K.; Mak, G.; Tsai, C.L.; Citardi, M.J.; Fakhri, S.; Shaw, J.L.; Fothergil, A.; Kheradmand, F.; et al. Airway surface mycosis in chronic TH2-associated airway disease. J. Allergy Clin. Immunol. 2014, 134, 325–331. [Google Scholar] [CrossRef] [Green Version]
- van Woerden, H.C.; Gregory, C.; Brown, R.; Marchesi, J.R.; Hoogendoorn, B.; Matthews, I.P. Differences in fungi present in induced sputum samples from asthma patients and non-atopic controls: A community based case control study. BMC Infect. Dis. 2013, 13, 69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cleland, E.J.; Bassiouni, A.; Boase, S.; Dowd, S.; Vreugde, S.; Wormald, P.J. The fungal microbiome in chronic rhinosinusitis: Richness, diversity, postoperative changes and patient outcomes. Int. Forum. Allergy Rhinol. 2014, 4, 259–265. [Google Scholar] [CrossRef] [PubMed]
- Hoggard, M.; Zoing, M.; Biswas, K.; Taylor, M.W.; Douglas, R.G. The sinonasal mycobiota in chronic rhinosinusitis and control patients. Rhinology 2019, 57, 190–199. [Google Scholar] [CrossRef] [PubMed]
- Guilbert, T.W.; Mauger, D.T.; Lemanske, R.F., Jr. Childhood asthma-predictive phenotype. J. Allergy Clin. Immunol. Pract. 2014, 2, 664–670. [Google Scholar] [CrossRef] [PubMed]
- Fretzayas, A.; Moustaki, M. Etiology and clinical features of viral bronchiolitis in infancy. World J. Pediatr. 2017, 13, 293–299. [Google Scholar] [CrossRef]
- Mauer, Y.; Taliercio, R.M. Managing adult asthma: The 2019 GINA guidelines. Clevel. Clin. J. Med. 2020, 87, 569–575. [Google Scholar] [CrossRef]
- GINA Report, Global Strategy for Asthma Management and Prevention. 2021. Available online: www.ginasthma.org (accessed on 1 February 2022).
- Fraczek, M.G.; Chishimba, L.; Niven, R.M.; Bromley, M.; Simpson, A.; Smyth, L.; Denning, D.W.; Bowyer, P. Corticosteroid treatment is associated with increased filamentous fungal burden in allergic fungal disease. J. Allergy Clin. Immunol. 2018, 142, 407–414. [Google Scholar] [CrossRef] [Green Version]
- Bachus, H.; Kaur, K.; Papillion, A.M.; Marquez-Lago, T.T.; Yu, Z.; Ballesteros-Tato, A.; Matalon, S.; Leon, B. Impaired Tumor-Necrosis-Factor-alpha-driven Dendritic Cell Activation Limits Lipopolysaccharide-Induced Protection from Allergic Inflammation in Infants. Immunity 2019, 50, 225–240.e224. [Google Scholar] [CrossRef] [Green Version]
- Castanhinha, S.; Sherburn, R.; Walker, S.; Gupta, A.; Bossley, C.J.; Buckley, J.; Ullmann, N.; Grychtol, R.; Campbell, G.; Maglione, M.; et al. Pediatric severe asthma with fungal sensitization is mediated by steroid-resistant IL-33. J. Allergy Clin. Immunol. 2015, 136, 312–322.e317. [Google Scholar] [CrossRef] [Green Version]
- Stein, M.M.; Hrusch, C.L.; Gozdz, J.; Igartua, C.; Pivniouk, V.; Murray, S.E.; Ledford, J.G.; Marques Dos Santos, M.; Anderson, R.L.; Metwali, N.; et al. Innate Immunity and Asthma Risk in Amish and Hutterite Farm Children. N. Engl. J. Med. 2016, 375, 411–421. [Google Scholar] [CrossRef] [Green Version]
- Del Giacco, S.R.; Bakirtas, A.; Bel, E.; Custovic, A.; Diamant, Z.; Hamelmann, E.; Heffler, E.; Kalayci, O.; Saglani, S.; Sergejeva, S.; et al. Allergy in severe asthma. Allergy 2017, 72, 207–220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaitanis, G.; Magiatis, P.; Hantschke, M.; Bassukas, I.D.; Velegraki, A. The Malassezia genus in skin and systemic diseases. Clin. Microbiol. Rev. 2012, 25, 106–141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moissl-Eichinger, C.; Pausan, M.; Taffner, J.; Berg, G.; Bang, C.; Schmitz, R.A. Archaea Are Interactive Components of Complex Microbiomes. Trends Microbiol. 2018, 26, 70–85. [Google Scholar] [CrossRef] [PubMed]
- Belmok, A.; de Cena, J.A.; Kyaw, C.M.; Dame-Teixeira, N. The Oral Archaeome: A Scoping Review. J. Dent. Res. 2020, 99, 630–643. [Google Scholar] [CrossRef] [PubMed]
- Huynh, H.T.; Pignoly, M.; Nkamga, V.D.; Drancourt, M.; Aboudharam, G. The repertoire of archaea cultivated from severe periodontitis. PLoS ONE 2015, 10, e0121565. [Google Scholar] [CrossRef]
- Lepp, P.W.; Brinig, M.M.; Ouverney, C.C.; Palm, K.; Armitage, G.C.; Relman, D.A. Methanogenic Archaea and human periodontal disease. Proc. Natl. Acad. Sci. USA 2004, 101, 6176–6181. [Google Scholar] [CrossRef] [Green Version]
- Li, C.L.; Liu, D.L.; Jiang, Y.T.; Zhou, Y.B.; Zhang, M.Z.; Jiang, W.; Liu, B.; Liang, J.P. Prevalence and molecular diversity of Archaea in subgingival pockets of periodontitis patients. Oral Microbiol. Immunol. 2009, 24, 343–346. [Google Scholar] [CrossRef]
- Wagner Mackenzie, B.; West, A.G.; Waite, D.W.; Lux, C.A.; Douglas, R.G.; Taylor, M.W.; Biswas, K. A Novel Description of the Human Sinus Archaeome During Health and Chronic Rhinosinusitis. Front. Cell. Infect. Microbiol. 2020, 10, 398. [Google Scholar] [CrossRef]
- Dykewicz, M.S.; Rodrigues, J.M.; Slavin, R.G. Allergic fungal rhinosinusitis. J. Allergy Clin. Immunol. 2018, 142, 341–351. [Google Scholar] [CrossRef] [Green Version]
- Hadebe, S.; Brombacher, F. Environment and Host-Genetic Determinants in Early Development of Allergic Asthma: Contribution of Fungi. Front. Immunol. 2019, 10, 2696. [Google Scholar] [CrossRef] [Green Version]
- Zhang, I.; Pletcher, S.D.; Goldberg, A.N.; Barker, B.M.; Cope, E.K. Fungal Microbiota in Chronic Airway Inflammatory Disease and Emerging Relationships with the Host Immune Response. Front. Microbiol. 2017, 8, 2477. [Google Scholar] [CrossRef] [PubMed] [Green Version]

{kind=link}
{kind=link}
| Research Article | Disease | Key Points |
|---|---|---|
| Boutin and colleagues (2017) Comparison of oropharyngeal microbiota from children with asthma and cystic fibrosis [36] | Asthma, cystic fibrosis |
|
| Cardenas and colleagues (2012) Upper Airways Microbiota in Antibiotic-Naïve Wheezing and Healthy Infants from Tropics of Rural Ecuador [37] | Wheezing |
|
| Cuthbertson and colleagues (2019) Viral respiratory infections and oropharyngeal microbiota in acutely wheezing children [38] | Wheezing |
|
| Hu and colleagues (2017) Dynamic oropharyngeal and faecal microbiota during treatment in infants hospitalized for bronchiolitis compared with age-matched healthy subjects [39] | Bronchiolitis |
|
| Powell and colleagues (2019) Temporal association of development of oropharyngeal microbiota with early life wheeze in population-based birth cohort [40] | Wheezing |
|
| Research Article | Disease | Key Points |
|---|---|---|
| Kelly et al. (2017) The nasopharyngeal microbiota of children with respiratory infections in Botswana [41] | Respiratory infections |
|
| Luna et al. (2018) The association between anterior nares and nasopharyngeal microbiota in infants hospitalized for bronchiolitis [46] | Bronchiolitis |
|
| Man et al. (2019) Loss of Microbial Topography between oral and nasopharyngeal microbiota and development of respiratory infections early in life [48] | Respiratory infections |
|
| Perez et al. (2017) Nasopharyngeal microbiome in premature infants and stability during rhinovirus-infection [42] | Rhinovirus infection |
|
| Rosas-Salazar et al. (2018) Nasopharyngeal Lactobacillus is associated with reduced risk of childhood wheezing illnesses following acute respiratory syncytial virus infection in infancy [47] | Wheezing, RSV infection |
|
| Tang et al. (2021) Developmental patterns in the nasopharyngeal microbiome during infancy are associated with asthma risk [49] | Asthma |
|
| Teo et al. (2015) The infant nasopharyngeal microbiome impacts severity of lower respiratory infection and risk of asthma development [43] | Asthma, respiratory infections |
|
| Teo et al. (2018) Airway microbiota dynamics uncover a critical window for interplay of pathogenic bacteria and allergy in childhood respiratory disease [50] | Allergy, respiratory disease, wheezing |
|
| Research Article | Disease | Key Points |
|---|---|---|
| Ward et al. (2018) Development of the human mycobiome over the first month of life and across body sites [82] | - |
|
| Ghannoum et al. (2010) Characterization of the oral fungal microbiome (mycobiome) in healthy individuals [83] | - |
|
| Porter et al. (2014) Airway Surface Mycosis in Chronic Th2-Associated Airway Disease [86] | CRS CRSsNP CRSwNP AFRS control cohort |
|
| Van Woerden (2013) Differences in fungi present in induced sputum samples from asthma patients and non-atopic controls: a community-based case–control study [87] | Asthma control cohort |
|
| Cleland et al. (2014) The fungal microbiome in chronic rhinosinusitis: richness, diversity, postoperative changes and patient outcomes [88] | CRS control cohort |
|
| Hoggard et al. (2019) The sinonasal mycobiota in chronic rhinosinusitis and control patients [89] | CRS -CRSsNP -CRSwNP -CRSwCF Control cohort |
|
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Baehren, C.; Buedding, E.; Bellm, A.; Schult, F.; Pembaur, A.; Wirth, S.; Ehrhardt, A.; Paulsen, F.; Postberg, J.; Aydin, M. The Relevance of the Bacterial Microbiome, Archaeome and Mycobiome in Pediatric Asthma and Respiratory Disorders. Cells 2022, 11, 1287. https://doi.org/10.3390/cells11081287
Baehren C, Buedding E, Bellm A, Schult F, Pembaur A, Wirth S, Ehrhardt A, Paulsen F, Postberg J, Aydin M. The Relevance of the Bacterial Microbiome, Archaeome and Mycobiome in Pediatric Asthma and Respiratory Disorders. Cells. 2022; 11(8):1287. https://doi.org/10.3390/cells11081287
Chicago/Turabian StyleBaehren, Carolin, Eleni Buedding, Aliyah Bellm, Frank Schult, Anton Pembaur, Stefan Wirth, Anja Ehrhardt, Friedrich Paulsen, Jan Postberg, and Malik Aydin. 2022. "The Relevance of the Bacterial Microbiome, Archaeome and Mycobiome in Pediatric Asthma and Respiratory Disorders" Cells 11, no. 8: 1287. https://doi.org/10.3390/cells11081287
APA StyleBaehren, C., Buedding, E., Bellm, A., Schult, F., Pembaur, A., Wirth, S., Ehrhardt, A., Paulsen, F., Postberg, J., & Aydin, M. (2022). The Relevance of the Bacterial Microbiome, Archaeome and Mycobiome in Pediatric Asthma and Respiratory Disorders. Cells, 11(8), 1287. https://doi.org/10.3390/cells11081287