
Unlaid Eggs: Ovarian Damage after Low-Dose Radiation

 , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
3. The Human Ovary and Folliculogenesis
Folliculogenesis
4. Application of Low-Dose Radiation in Clinical Practice
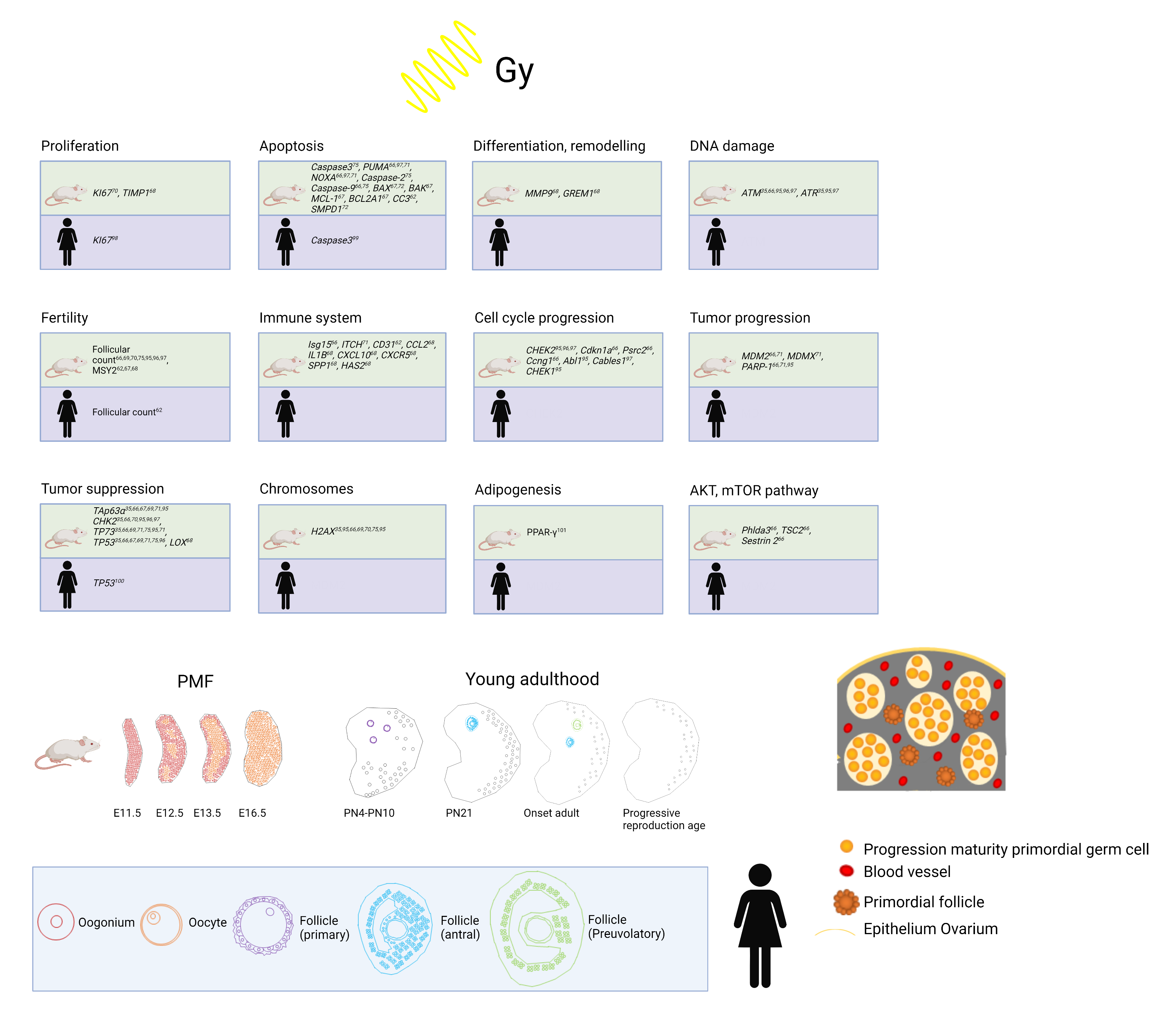
5. Animal Models and LDR: Effects on the Ovary
5.1. Selection of Model
5.2. Mouse Model: Current Knowledge and Further Approaches
5.3. Effects of Low-Dose Radiation on Ovarian Stromal and Immune Cells
6. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Miller, K.D.; Siegel, R.L.; Lin, C.C.; Mariotto, A.B.; Kramer, J.L.; Rowland, J.H.; Stein, K.D.; Alteri, R.; Jemal, A. Cancer treatment and survivorship statistics, 2016. CA A Cancer J. Clin. 2016, 66, 271–289. [Google Scholar] [CrossRef] [PubMed]
- Von der Weid, N.X. Adult life after surviving lymphoma in childhood. Support. Care Cancer 2008, 16, 339–345. [Google Scholar] [CrossRef] [PubMed]
- Anderson, R.A.; Brewster, D.H.; Wood, R.; Nowell, S.; Fischbacher, C.; Kelsey, T.W.; Wallace, W.H.B. The impact of cancer on subsequent chance of pregnancy: A population-based analysis. Hum. Reprod. 2018, 33, 1281–1290. [Google Scholar] [CrossRef] [PubMed]
- Bedoschi, G.M.; Navarro, P.A.; Oktay, K.H. Novel insights into the pathophysiology of chemotherapy-induced damage to the ovary. Panminerva Med. 2019, 61, 68–75. [Google Scholar] [CrossRef]
- Ben-Aharon, I.; Bar-Joseph, H.; Tzarfaty, G.; Kuchinsky, L.; Rizel, S.; Stemmer, S.M.; Shalgi, R. Doxorubicin-induced ovarian toxicity. Reprod. Biol. Endocrinol. 2010, 8, 20. [Google Scholar] [CrossRef]
- Bildik, G.; Akin, N.; Senbabaoglu, F.; Sahin, G.N.; Karahuseyinoglu, S.; Ince, U.; Taskiran, C.; Selek, U.; Yakin, K.; Guzel, Y.; et al. GnRH agonist leuprolide acetate does not confer any protection against ovarian damage induced by chemotherapy and radiation in vitro. Hum. Reprod. 2015, 30, 2912–2925. [Google Scholar]
- Yuksel, A.; Bildik, G.; Senbabaoglu, F.; Akin, N.; Arvas, M.; Unal, F.; Kilic, Y.; Karanfil, I.; Eryılmaz, B.; Yilmaz, P.; et al. The magnitude of gonadotoxicity of chemotherapy drugs on ovarian follicles and granulosa cells varies depending upon the category of the drugs and the type of granulosa cells. Hum. Reprod. 2015, 30, 2926–2935. [Google Scholar] [CrossRef]
- Chang, E.M.; Lim, E.; Yoon, S.; Jeong, K.; Bae, S.; Lee, D.R.; Yoon, T.K.; Choi, Y.; Lee, W.S. Cisplatin Induces Overactivation of the Dormant Primordial Follicle through PTEN/AKT/FOXO3a Pathway which Leads to Loss of Ovarian Reserve in Mice. PLoS ONE 2015, 10, e0144245. [Google Scholar] [CrossRef]
- Chow, E.J.; Stratton, K.L.; Leisenring, W.M.; Oeffinger, K.C.; Sklar, C.A.; Donaldson, S.S.; Ginsberg, J.P.; Kenney, L.B.; Levine, J.M.; Robison, L.L.; et al. Pregnancy after chemotherapy in male and female survivors of childhood cancer treated between 1970 and 1999: A report from the Childhood Cancer Survivor Study cohort. Lancet Oncol. 2016, 17, 567–576. [Google Scholar] [CrossRef]
- Nguyen, Q.N.; Zerafa, N.; Liew, S.H.; Morgan, F.H.; Strasser, A.; Scott, C.L.; Findlay, J.K.; Hickey, M.; Hutt, K.J. Loss of PUMA protects the ovarian reserve during DNA-damaging chemotherapy and preserves fertility. Cell Death Dis. 2018, 9, 1–12. [Google Scholar] [CrossRef]
- Özcan, P.; Fıçıcıoğlu, C.; Yıldırım, Ö.K.; Özkan, F.; Akkaya, H.; Aslan, İ. Protective effect of resveratrol against oxidative damage to ovarian reserve in female Sprague-Dawley rats. Reprod. Biomed. Online 2015, 31, 404–410. [Google Scholar] [CrossRef]
- Liu, Z.-Q.; Shen, M.; Wu, W.-J.; Li, B.-J.; Weng, Q.-N.; Li, M.; Liu, H.-L. Expression of PUMA in Follicular Granulosa Cells Regulated by FoxO1 Activation During Oxidative Stress. Reprod. Sci. 2015, 22, 696–705. [Google Scholar] [CrossRef]
- Cronin, K.A.; Lake, A.J.; Scott, S.; Sherman, R.L.; Noone, A.M.; Howlader, N.; Henley, S.J.; Anderson, R.N.; Firth, A.U.; Ma, J.; et al. Annual Report to the Nation on the Status of Cancer, part I: National cancer statistics. Cancer 2018, 124, 2785–2800. [Google Scholar] [CrossRef]
- Motta, P.M.; Makabe, S.; Nottola, S.A. The ultrastructure of human reproduction. I. The natural history of the female germ cell: Origin, migration and differentiation inside the developing ovary. Hum. Reprod. Update 1997, 3, 281–297. [Google Scholar] [CrossRef]
- Grive, K.J.; Freiman, R.N. The developmental origins of the mammalian ovarian reserve. Development 2015, 142, 2554–2563. [Google Scholar] [CrossRef]
- Mork, L.; Maatouk, D.M.; McMahon, J.A.; Guo, J.J.; Zhang, P.; McMahon, A.P.; Capel, B. Temporal differences in granulosa cell specification in the ovary reflect distinct follicle fates in mice. Biol. Reprod. 2012, 86, 37. [Google Scholar] [CrossRef]
- Liu, C.; Peng, J.; Matzuk, M.M.; Yao, H.H. Lineage specification of ovarian theca cells requires multicellular interactions via oocyte and granulosa cells. Nat. Commun. 2015, 6, 6934. [Google Scholar] [CrossRef]
- Rotgers, E.; Jørgensen, A.; Yao, H.H. At the Crossroads of Fate-Somatic Cell Lineage Specification in the Fetal Gonad. Endocr. Rev. 2018, 39, 739–759. [Google Scholar] [CrossRef]
- Wagner, M.; Yoshihara, M.; Douagi, I.; Damdimopoulos, A.; Panula, S.; Petropoulos, S.; Lu, H.; Pettersson, K.; Palm, K.; Katayama, S.; et al. Single-cell analysis of human ovarian cortex identifies distinct cell populations but no oogonial stem cells. Nat. Commun. 2020, 11, 1147. [Google Scholar] [CrossRef]
- Fan, X.; Bialecka, M.; Moustakas, I.; Lam, E.; Torrens-Juaneda, V.; Borggreven, N.V.; Trouw, L.; Louwe, L.A.; Pilgram, G.S.K.; Mei, H.; et al. Single-cell reconstruction of follicular remodeling in the human adult ovary. Nat. Commun. 2019, 10, 3164. [Google Scholar] [CrossRef]
- Thomas, P.R.; Winstanly, D.; Peckham, M.J.; Austin, D.E.; Murray, M.A.; Jacobs, H.S. Reproductive and endocrine function in patients with Hodgkin’s disease: Effects of oophoropexy and irradiation. Br. J. Cancer 1976, 33, 226–231. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Yamamoto, R.; Okamoto, K.; Yukiharu, T.; Kaneuchi, M.; Negishi, H.; Sakuragi, N.; Fujimoto, S. A study of risk factors for ovarian metastases in stage Ib-IIIb cervical carcinoma and analysis of ovarian function after a transposition. Gynecol. Oncol. 2001, 82, 312–316. [Google Scholar] [CrossRef] [PubMed]
- van Beurden, M.; Schuster-Uitterhoeve, A.; Lammes, F. Feasibility of transposition of the ovaries in the surgical and radiotherapeutical treatment of cervical cancer. Eur. J. Surg. Oncol. J. Eur. Soc. Surg. Oncol. Br. Assoc. Surg. Oncol. 1990, 16, 141–146. [Google Scholar]
- Hoekman, E.J.; Broeders, E.; Louwe, L.A.; Nout, R.A.; Jansen, F.W.; de Kroon, C.D. Ovarian function after ovarian transposition and additional pelvic radiotherapy: A systematic review. Eur J. Surg Oncol 2019, 45, 1328–1340. [Google Scholar] [CrossRef]
- Pahisa, J.; MartÍNez-RomÁN, S.; MartÍNez-Zamora, M.A.; TornÉ, A.; CaparrÓS, X.; SanjuÁN, A.; LejÁRcegui, J.A. Laparoscopic ovarian transposition in patients with early cervical cancer. Int. J. Gynecol. Cancer 2008, 18, 584–589. [Google Scholar] [CrossRef]
- Husseinzadeh, N.; Van Aken, M.L.; Aron, B. Ovarian transposition in young patients with invasive cervical cancer receiving radiation therapy. Int. J. Gynecol. Cancer Off. J. Int. Gynecol. Cancer Soc. 1994, 4, 61–65. [Google Scholar] [CrossRef]
- Anderson, R.A.; Amant, F.; Braat, D.; D’Angelo, A.; Chuva de Sousa Lopes, S.M.; Demeestere, I.; Dwek, S.; Frith, L.; Lambertini, M.; Maslin, C.; et al. ESHRE guideline: Female fertility preservation. Hum. Reprod. Open 2020, 2020, hoaa052. [Google Scholar] [CrossRef]
- Koh, W.J.; Abu-Rustum, N.R.; Bean, S.; Bradley, K.; Campos, S.M.; Cho, K.R.; Chon, H.S.; Chu, C.; Clark, R.; Cohn, D.; et al. Cervical Cancer, Version 3.2019, NCCN Clinical Practice Guidelines in Oncology. J. Natl. Compr. Cancer Netw. 2019, 17, 64–84. [Google Scholar] [CrossRef]
- Lee, S.J.; Schover, L.R.; Partridge, A.H.; Patrizio, P.; Wallace, W.H.; Hagerty, K.; Beck, L.N.; Brennan, L.V.; Oktay, K. American Society of Clinical Oncology recommendations on fertility preservation in cancer patients. J. Clin. Oncol. 2006, 24, 2917–2931. [Google Scholar] [CrossRef]
- Oktay, K.; Harvey, B.E.; Partridge, A.H.; Quinn, G.P.; Reinecke, J.; Taylor, H.S.; Wallace, W.H.; Wang, E.T.; Loren, A.W. Fertility Preservation in Patients With Cancer: ASCO Clinical Practice Guideline Update. J. Clin. Oncol. 2018, 36, 1994–2001. [Google Scholar] [CrossRef]
- Nagy, Z.P.; Anderson, R.E.; Feinberg, E.C.; Hayward, B.; Mahony, M.C. The Human Oocyte Preservation Experience (HOPE) Registry: Evaluation of cryopreservation techniques and oocyte source on outcomes. Reprod. Biol. Endocrinol. 2017, 15, 1–10. [Google Scholar] [CrossRef]
- Balcerek, M.; Wolff, M.; Nawroth, F. Indikation Und Durchführung Fertilitätsprotektiver Massnahmen Bei Onkologischen Und Nichtonkologischen Erkrankungen. 2017. Available online: https://www.sggg.ch/fr/formation/formation-continue/evenements/detail/1/indikation-und-durchfuehrung-fertilitaetsprospektiver-massnahmen-bei-onkologischen-und-nicht-onkologischen-erkrankungen/ (accessed on 12 December 2021).
- Winkler-Crepaz, K.; Bottcher, B.; Toth, B.; Wildt, L.; Hofer-Tollinger, S. What is new in 2017? Update on fertility preservation in cancer patients. Minerva Endocrinol. 2017, 42, 331–339. [Google Scholar] [CrossRef]
- Adriaens, I.; Smitz, J.; Jacquet, P. The current knowledge on radiosensitivity of ovarian follicle development stages. Hum. Reprod. Update 2009, 15, 359–377. [Google Scholar] [CrossRef]
- Gebel, J.; Tuppi, M.; Sänger, N.; Schumacher, B.; Dötsch, V. DNA Damaged Induced Cell Death in Oocytes. Molecules 2020, 25, 5714. [Google Scholar] [CrossRef]
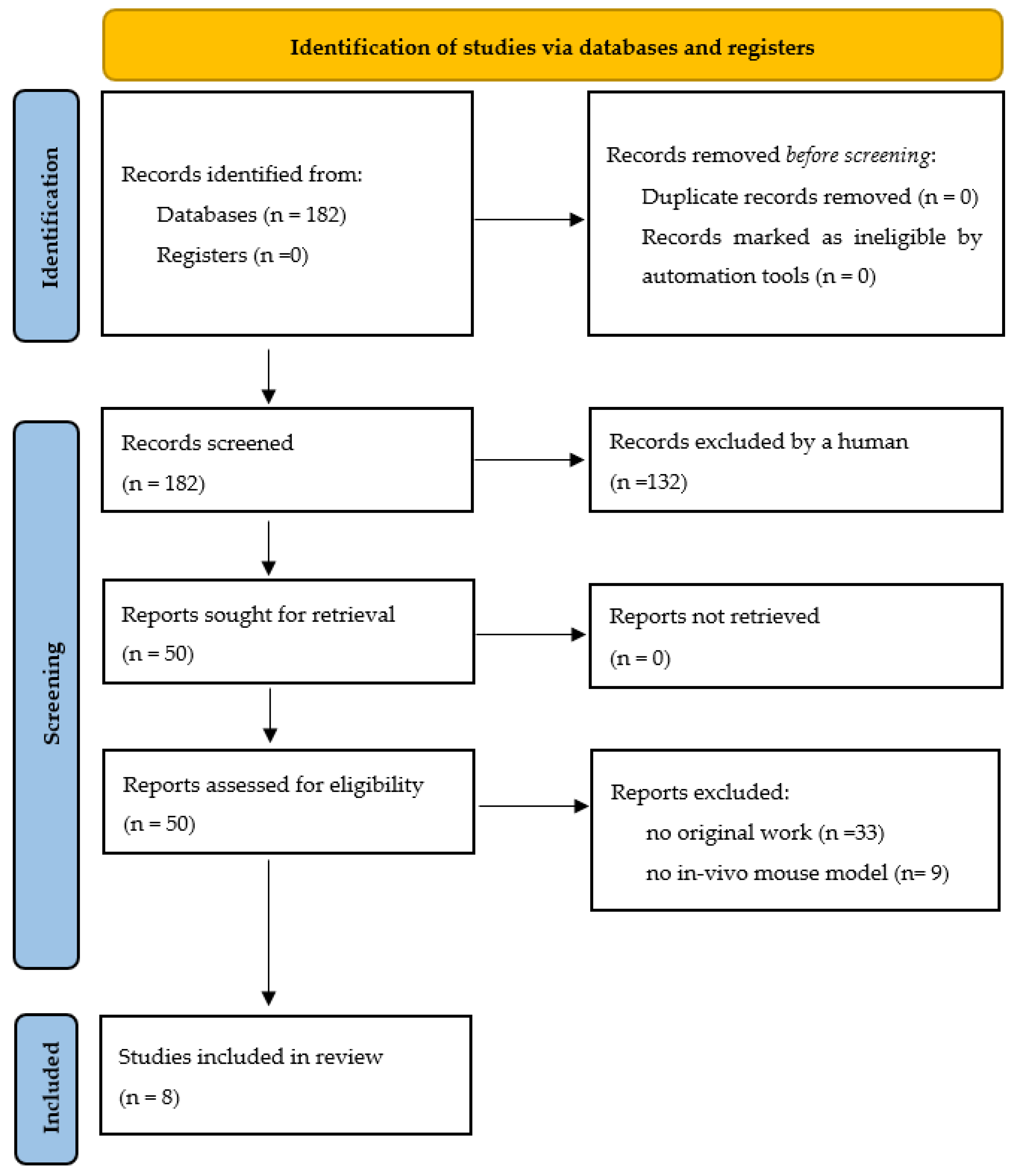
- Page, M.J.; McKenzie, J.E.; Bossuyt, P.M.; Boutron, I.; Hoffmann, T.C.; Mulrow, C.D.; Shamseer, L.; Tetzlaff, J.M.; Akl, E.A.; Brennan, S.E.; et al. The PRISMA 2020 statement: An updated guideline for reporting systematic reviews. Int. J. Surg. 2021, 372, 105906. [Google Scholar] [CrossRef]
- Suh, E.K.; Yang, A.; Kettenbach, A.; Bamberger, C.; Michaelis, A.H.; Zhu, Z.; Elvin, J.A.; Bronson, R.T.; Crum, C.P.; McKeon, F. p63 protects the female germ line during meiotic arrest. Nature 2006, 444, 624–628. [Google Scholar] [CrossRef]
- Kerr, J.B.; Hutt, K.J.; Michalak, E.M.; Cook, M.; Vandenberg, C.J.; Liew, S.H.; Bouillet, P.; Mills, A.; Scott, C.L.; Findlay, J.K.; et al. DNA damage-induced primordial follicle oocyte apoptosis and loss of fertility require TAp63-mediated induction of Puma and Noxa. Mol. Cell 2012, 48, 343–352. [Google Scholar] [CrossRef]
- Deutsch, G.B.; Zielonka, E.M.; Coutandin, D.; Weber, T.A.; Schäfer, B.; Hannewald, J.; Luh, L.M.; Durst, F.G.; Ibrahim, M.; Hoffmann, J.; et al. DNA damage in oocytes induces a switch of the quality control factor TAp63α from dimer to tetramer. Cell 2011, 144, 566–576. [Google Scholar] [CrossRef]
- Puy, V.; Barroca, V.; Messiaen, S.; Ménard, V.; Torres, C.; Devanand, C.; Moison, D.; Lewandowski, D.; Guerquin, M.J.; Martini, E.; et al. Mouse model of radiation-induced premature ovarian insufficiency reveals compromised oocyte quality: Implications for fertility preservation. Reprod. Biomed. Online 2021, 43, 799–809. [Google Scholar] [CrossRef]
- Quan, N.; Harris, L.R.; Halder, R.; Trinidad, C.V.; Johnson, B.W.; Horton, S.; Kimler, B.F.; Pritchard, M.T.; Duncan, F.E. Differential sensitivity of inbred mouse strains to ovarian damage in response to low-dose total body irradiation. Biol. Reprod. 2020, 102, 133–144. [Google Scholar] [CrossRef]
- Kimler, B.F.; Briley, S.M.; Johnson, B.W.; Armstrong, A.G.; Jasti, S.; Duncan, F.E. Radiation-induced ovarian follicle loss occurs without overt stromal changes. Reproduction 2018, 155, 553–562. [Google Scholar] [CrossRef] [PubMed]
- Morita, Y.; Perez, G.I.; Paris, F.; Miranda, S.R.; Ehleiter, D.; Haimovitz-Friedman, A.; Fuks, Z.; Xie, Z.; Reed, J.C.; Schuchman, E.H.; et al. Oocyte apoptosis is suppressed by disruption of the acid sphingomyelinase gene or by sphingosine-1-phosphate therapy. Nat. Med. 2000, 6, 1109–1114. [Google Scholar] [CrossRef] [PubMed]
- Grover, A.R.; Kimler, B.F.; Duncan, F.E. Use of a Small Animal Radiation Research Platform (SARRP) facilitates analysis of systemic versus targeted radiation effects in the mouse ovary. J. Ovarian Res. 2018, 11, 72. [Google Scholar] [CrossRef] [PubMed]
- Hirshfield, A.N. Development of follicles in the mammalian ovary. Int. Rev. Cytol. 1991, 124, 43–101. [Google Scholar] [CrossRef]
- Gougeon, A.; Ecochard, R.; Thalabard, J.C. Age-related changes of the population of human ovarian follicles: Increase in the disappearance rate of non-growing and early-growing follicles in aging women. Biol. Reprod. 1994, 50, 653–663. [Google Scholar] [CrossRef]
- Coulam, C.B.; Adamson, S.C.; Annegers, J.F. Incidence of premature ovarian failure. Obstet. Gynecol. 1986, 67, 604–606. [Google Scholar] [CrossRef]
- Ernst, E.H.; Grøndahl, M.L.; Grund, S.; Hardy, K.; Heuck, A.; Sunde, L.; Franks, S.; Andersen, C.Y.; Villesen, P.; Lykke-Hartmann, K. Dormancy and activation of human oocytes from primordial and primary follicles: Molecular clues to oocyte regulation. Hum. Reprod. 2017, 32, 1684–1700. [Google Scholar] [CrossRef]
- Demeestere, I.; Brice, P.; Peccatori, F.A.; Kentos, A.; Dupuis, J.; Zachee, P.; Casasnovas, O.; Van Den Neste, E.; Dechene, J.; De Maertelaer, V.; et al. No Evidence for the Benefit of Gonadotropin-Releasing Hormone Agonist in Preserving Ovarian Function and Fertility in Lymphoma Survivors Treated With Chemotherapy: Final Long-Term Report of a Prospective Randomized Trial. J. Clin. Oncol. 2016, 34, 2568–2574. [Google Scholar] [CrossRef]
- Grosbois, J.; Demeestere, I. Dynamics of PI3K and Hippo signaling pathways during in vitro human follicle activation. Hum. Reprod. 2018, 33, 1705–1714. [Google Scholar] [CrossRef]
- Kawamura, K.; Cheng, Y.; Suzuki, N.; Deguchi, M.; Sato, Y.; Takae, S.; Ho, C.H.; Kawamura, N.; Tamura, M.; Hashimoto, S.; et al. Hippo signaling disruption and Akt stimulation of ovarian follicles for infertility treatment. Proc. Natl. Acad. Sci. USA 2013, 110, 17474–17479. [Google Scholar] [CrossRef]
- Delgado-Rosas, F.; Gaytán, M.; Morales, C.; Gómez, R.; Gaytán, F. Superficial ovarian cortex vascularization is inversely related to the follicle reserve in normal cycling ovaries and is increased in polycystic ovary syndrome. Hum. Reprod. 2009, 24, 1142–1151. [Google Scholar] [CrossRef]
- Peters, H.; Byskov, A.G.; Grinsted, J. Follicular growth in fetal and prepubertal ovaries of humans and other primates. Clin. Endocrinol. Metab. 1978, 7, 469–485. [Google Scholar] [CrossRef]
- Rinaldi, V.D.; Hsieh, K.; Munroe, R.; Bolcun-Filas, E.; Schimenti, J.C. Pharmacological Inhibition of the DNA Damage Checkpoint Prevents Radiation-Induced Oocyte Death. Genetics 2017, 206, 1823–1828. [Google Scholar] [CrossRef]
- Kim, S.Y.; Nair, D.M.; Romero, M.; Serna, V.A.; Koleske, A.J.; Woodruff, T.K.; Kurita, T. Transient inhibition of p53 homologs protects ovarian function from two distinct apoptotic pathways triggered by anticancer therapies. Cell Death Differ. 2019, 26, 502–515. [Google Scholar] [CrossRef]
- Woodard, T.L.; Bolcun-Filas, E. Prolonging Reproductive Life after Cancer: The Need for Fertoprotective Therapies. Trends Cancer 2016, 2, 222–233. [Google Scholar] [CrossRef]
- Rahmanzadeh, R.; Rai, P.; Celli, J.P.; Rizvi, I.; Baron-Lühr, B.; Gerdes, J.; Hasan, T. Ki-67 as a molecular target for therapy in an in vitro three-dimensional model for ovarian cancer. Cancer Res. 2010, 70, 9234–9242. [Google Scholar] [CrossRef]
- Wiweko, B.; Soebijanto, S.; Boediono, A.; Mansyur, M.; Siregar, N.C.; Suryandari, D.A.; Aulia, A.; Djuwantono, T.; Affandi, B. Survival of isolated human preantral follicles after vitrification: Analyses of morphology and Fas ligand and caspase-3 mRNA expression. Clin. Exp. Reprod. Med. 2019, 46, 152–165. [Google Scholar] [CrossRef]
- Zhang, Y.; Cao, L.; Nguyen, D.; Lu, H. TP53 mutations in epithelial ovarian cancer. Transl. Cancer Res. 2016, 5, 650–663. [Google Scholar] [CrossRef]
- Chi, T.; Wang, M.; Wang, X.; Yang, K.; Xie, F.; Liao, Z.; Wei, P. PPAR-γ Modulators as Current and Potential Cancer Treatments. Front. Oncol. 2021, 11, 737776. [Google Scholar] [CrossRef]
- Heeren, A.M.; van Iperen, L.; Klootwijk, D.B.; de Melo Bernardo, A.; Roost, M.S.; Gomes Fernandes, M.M.; Louwe, L.A.; Hilders, C.G.; Helmerhorst, F.M.; van der Westerlaken, L.A.; et al. Development of the follicular basement membrane during human gametogenesis and early folliculogenesis. BMC Dev. Biol. 2015, 15, 4. [Google Scholar] [CrossRef]
- Soygur, B.; Laird, D.J. Ovary Development: Insights From a Three-Dimensional Imaging Revolution. Front. Cell Dev. Biol. 2021, 9, 698315. [Google Scholar] [CrossRef]
- Palmer, J.D.; Tsang, D.S.; Tinkle, C.L.; Olch, A.J.; Kremer, L.C.M.; Ronckers, C.M.; Gibbs, I.C.; Constine, L.S. Late effects of radiation therapy in pediatric patients and survivorship. Pediatr. Blood Cancer 2021, 68 (Suppl. 2), e28349. [Google Scholar] [CrossRef]
- Barton, S.E.; Najita, J.S.; Ginsburg, E.S.; Leisenring, W.M.; Stovall, M.; Weathers, R.E.; Sklar, C.A.; Robison, L.L.; Diller, L. Infertility, infertility treatment, and achievement of pregnancy in female survivors of childhood cancer: A report from the Childhood Cancer Survivor Study cohort. Lancet Oncol. 2013, 14, 873–881. [Google Scholar] [CrossRef]
- Oonsiri, P.; Vannavijit, C.; Wimolnoch, M.; Suriyapee, S.; Saksornchai, K. Estimated radiation doses to ovarian and uterine organs in breast cancer irradiation using radio-photoluminescent glass dosimeters (RPLDs). J. Med. Radiat. Sci. 2021, 68, 167–174. [Google Scholar] [CrossRef]
- Kovtun, K.A.; Yeo, W.-P.; Phillips, C.H.; Viswanathan, A.; Baldini, E.H. Ovary-Sparing Radiation Planning Techniques Can Achieve Ovarian Dose Reduction for Soft Tissue Sarcoma of the Buttock and Thigh. Sarcoma 2017, 2017, 2796925. [Google Scholar] [CrossRef]
- Doss, M. Are We Approaching the End of the Linear No-Threshold Era? J. Nucl. Med. 2018, 59, 1786–1793. [Google Scholar] [CrossRef]
- Tang, F.R.; Loke, W.K. Molecular mechanisms of low dose ionizing radiation-induced hormesis, adaptive responses, radioresistance, bystander effects, and genomic instability. Int. J. Radiat. Biol. 2015, 91, 13–27. [Google Scholar] [CrossRef]
- Jaworowski, Z. Observations on the Chernobyl Disaster and LNT. Dose Response 2010, 8, 148–171. [Google Scholar] [CrossRef]
- Sutou, S. Low-dose radiation from A-bombs elongated lifespan and reduced cancer mortality relative to un-irradiated individuals. Genes Environ. 2018, 40, 26. [Google Scholar] [CrossRef]
- Shore, R.E.; Beck, H.L.; Boice, J.D.; Caffrey, E.A.; Davis, S.; Grogan, H.A.; Mettler, F.A.; Preston, R.J.; Till, J.E.; Wakeford, R.; et al. Implications of recent epidemiologic studies for the linear nonthreshold model and radiation protection. J. Radiol. Prot. 2018, 38, 1217–1233. [Google Scholar] [CrossRef]
- Rodgers, B.E.; Holmes, K.M. Radio-adaptive response to environmental exposures at Chernobyl. Dose Response 2008, 6, 209–221. [Google Scholar] [CrossRef] [PubMed]
- Shore, R.; Walsh, L.; Azizova, T.; Rühm, W. Risk of solid cancer in low dose-rate radiation epidemiological studies and the dose-rate effectiveness factor. Int. J. Radiat. Biol. 2017, 93, 1064–1078. [Google Scholar] [CrossRef] [PubMed]
- ICRP. The 2007 Recommendations of the International Commission on Radiological Protection. Ann. ICRP 2007, 37, 1–332. [Google Scholar] [CrossRef]
- Stewart, F.A.; Akleyev, A.V.; Hauer-Jensen, M.; Hendry, J.H.; Kleiman, N.J.; Macvittie, T.J.; Aleman, B.M.; Edgar, A.B.; Mabuchi, K.; Muirhead, C.R.; et al. ICRP publication 118: ICRP statement on tissue reactions and early and late effects of radiation in normal tissues and organs--threshold doses for tissue reactions in a radiation protection context. Ann. ICRP 2012, 41, 1–322. [Google Scholar] [CrossRef]
- Harrison, J.D.; Balonov, M.; Bochud, F.; Martin, C.; Menzel, H.G.; Ortiz-Lopez, P.; Smith-Bindman, R.; Simmonds, J.R.; Wakeford, R. ICRP Publication 147: Use of Dose Quantities in Radiological Protection. Ann. ICRP 2021, 50, 9–82. [Google Scholar] [CrossRef]
- Barnett, G.C.; West, C.M.; Dunning, A.M.; Elliott, R.M.; Coles, C.E.; Pharoah, P.D.; Burnet, N.G. Normal tissue reactions to radiotherapy: Towards tailoring treatment dose by genotype. Nat. Rev. Cancer 2009, 9, 134–142. [Google Scholar] [CrossRef]
- Baker, T.G. Effects of ionizing radiations on mammalian oogenesis: A model for chemical effects. Environ. Health Perspect. 1978, 24, 31–37. [Google Scholar] [CrossRef]
- Onder, G.O.; Balcioglu, E.; Baran, M.; Ceyhan, A.; Cengiz, O.; Suna, P.A.; Yıldız, O.G.; Yay, A. The different doses of radiation therapy-induced damage to the ovarian environment in rats. Int. J. Radiat. Biol. 2021, 97, 367–375. [Google Scholar] [CrossRef]
- Dubois, C.; Pophillat, M.; Audebert, S.; Fourquet, P.; Lecomte, C.; Dubourg, N.; Galas, S.; Camoin, L.; Frelon, S. Differential modification of the C. elegans proteome in response to acute and chronic gamma radiation: Link with reproduction decline. Sci. Total Environ. 2019, 676, 767–781. [Google Scholar] [CrossRef]
- Zelinski, M.B.; Murphy, M.K.; Lawson, M.S.; Jurisicova, A.; Pau, K.Y.; Toscano, N.P.; Jacob, D.S.; Fanton, J.K.; Casper, R.F.; Dertinger, S.D.; et al. In vivo delivery of FTY720 prevents radiation-induced ovarian failure and infertility in adult female nonhuman primates. Fertil. Steril. 2011, 95, 1440–1445. [Google Scholar] [CrossRef]
- Fester, N.; Zielonka, E.; Goldmann, J.; Frombach, A.S.; Müller-Kuller, U.; Gutfreund, N.; Riegel, K.; Smits, J.G.A.; Schleiff, E.; Rajalingam, K.; et al. Enhanced pro-apoptosis gene signature following the activation of TAp63α in oocytes upon γ irradiation. Cell Death Dis. 2022, 13, 204. [Google Scholar] [CrossRef]
- Hanoux, V.; Pairault, C.; Bakalska, M.; Habert, R.; Livera, G. Caspase-2 involvement during ionizing radiation-induced oocyte death in the mouse ovary. Cell Death Differ. 2007, 14, 671–681. [Google Scholar] [CrossRef]
- Livera, G.; Petre-Lazar, B.; Guerquin, M.J.; Trautmann, E.; Coffigny, H.; Habert, R. p63 null mutation protects mouse oocytes from radio-induced apoptosis. Reproduction 2008, 135, 3–12. [Google Scholar] [CrossRef]
- Duncan, F.E.; Kimler, B.F.; Briley, S.M. Combating radiation therapy-induced damage to the ovarian environment. Future Oncol. 2016, 12, 1687–1690. [Google Scholar] [CrossRef]
- Straub, J.M.; New, J.; Hamilton, C.D.; Lominska, C.; Shnayder, Y.; Thomas, S.M. Radiation-induced fibrosis: Mechanisms and implications for therapy. J. Cancer Res. Clin. Oncol. 2015, 141, 1985–1994. [Google Scholar] [CrossRef]
- Meirow, D.; Biederman, H.; Anderson, R.A.; Wallace, W.H.B. Toxicity of chemotherapy and radiation on female reproduction. Clin. Obstet. Gynecol. 2010, 53, 727–739. [Google Scholar] [CrossRef]
- Reeves, G. Specific stroma in the cortex and medulla of the ovary. Cell types and vascular supply in relation to follicular apparatus and ovulation. Obstet. Gynecol. 1971, 37, 832–844. [Google Scholar] [CrossRef]
- Pepe, G.; Locati, M.; Della Torre, S.; Mornata, F.; Cignarella, A.; Maggi, A.; Vegeto, E. The estrogen-macrophage interplay in the homeostasis of the female reproductive tract. Hum. Reprod. Update 2018, 24, 652–672. [Google Scholar] [CrossRef]
- Carlock, C.; Wu, J.; Zhou, C.; Ross, A.; Adams, H.; Lou, Y. Ovarian phagocyte subsets and their distinct tissue distribution patterns. Reproduction 2013, 146, 491–500. [Google Scholar] [CrossRef]
- DeLoia, J.A.; Stewart-Akers, A.M.; Brekosky, J.; Kubik, C.J. Effects of exogenous estrogen on uterine leukocyte recruitment. Fertil. Steril. 2002, 77, 548–554. [Google Scholar] [CrossRef]
- Ljunggren, H.G.; Kärre, K. In search of the ’missing self’: MHC molecules and NK cell recognition. Immunol. Today 1990, 11, 237–244. [Google Scholar] [CrossRef]
- Strobel, L.; Vomstein, K.; Kyvelidou, C.; Hofer-Tollinger, S.; Feil, K.; Kuon, R.-J.; Ebner, S.; Troppmair, J.; Toth, B. Different Background: Natural Killer Cell Profiles in Secondary versus Primary Recurrent Pregnancy Loss. J. Clin. Med. 2021, 10, 194. [Google Scholar] [CrossRef] [PubMed]
- Kuon, R.J.; Vomstein, K.; Weber, M.; Müller, F.; Seitz, C.; Wallwiener, S.; Strowitzki, T.; Schleussner, E.; Markert, U.R.; Daniel, V.; et al. The "killer cell story" in recurrent miscarriage: Association between activated peripheral lymphocytes and uterine natural killer cells. J. Reprod. Immunol. 2017, 119, 9–14. [Google Scholar] [CrossRef] [PubMed]
- Toth, B.; Vomstein, K.; Togawa, R.; Böttcher, B.; Hudalla, H.; Strowitzki, T.; Daniel, V.; Kuon, R.J. The impact of previous live births on peripheral and uterine natural killer cells in patients with recurrent miscarriage. Reprod. Biol. Endocrinol. 2019, 17, 72. [Google Scholar] [CrossRef]
- Amargant, F.; Manuel, S.L.; Larmore, M.J.; Johnson, B.W.; Lawson, M.; Pritchard, M.T.; Zelinski, M.B.; Duncan, F.E. Sphingosine-1-phosphate and its mimetic FTY720 do not protect against radiation-induced ovarian fibrosis in the nonhuman primate†. Biol. Reprod. 2021, 104, 1058–1070. [Google Scholar] [CrossRef]
- Du, Y.; Carranza, Z.; Luan, Y.; Busman-Sahay, K.; Wolf, S.; Campbell, S.P.; Kim, S.Y.; Pejovic, T.; Estes, J.D.; Zelinski, M.; et al. Evidence of cancer therapy-induced chronic inflammation in the ovary across multiple species: A potential cause of persistent tissue damage and follicle depletion. J. Reprod. Immunol. 2022, 150, 103491. [Google Scholar] [CrossRef]
- Redon, C.E.; Dickey, J.S.; Bonner, W.M.; Sedelnikova, O.A. γ-H2AX as a biomarker of DNA damage induced by ionizing radiation in human peripheral blood lymphocytes and artificial skin. Adv. Space Res. 2009, 43, 1171–1178. [Google Scholar] [CrossRef]
- Lee, Y.; Wang, Q.; Shuryak, I.; Brenner, D.J.; Turner, H.C. Development of a high-throughput γ-H2AX assay based on imaging flow cytometry. Radiat. Oncol. 2019, 14, 150. [Google Scholar] [CrossRef]
- Gross, J.P.; Kim, S.Y.; Gondi, V.; Pankuch, M.; Wagner, S.; Grover, A.; Luan, Y.; Woodruff, T.K. Proton Radiotherapy to Preserve Fertility and Endocrine Function: A Translational Investigation. Int. J. Radiat. Oncol. Biol. Phys. 2021, 109, 84–94. [Google Scholar] [CrossRef]
- Rothkamm, K.; Löbrich, M. Evidence for a lack of DNA double-strand break repair in human cells exposed to very low x-ray doses. Proc. Natl. Acad. Sci. USA 2003, 100, 5057–5062. [Google Scholar] [CrossRef]
- Grudzenski, S.; Raths, A.; Conrad, S.; Rübe, C.E.; Löbrich, M. Inducible response required for repair of low-dose radiation damage in human fibroblasts. Proc. Natl. Acad. Sci. USA 2010, 107, 14205–14210. [Google Scholar] [CrossRef]
- Nagle, P.W.; Hosper, N.A.; Barazzuol, L.; Jellema, A.L.; Baanstra, M.; van Goethem, M.J.; Brandenburg, S.; Giesen, U.; Langendijk, J.A.; van Luijk, P.; et al. Lack of DNA Damage Response at Low Radiation Doses in Adult Stem Cells Contributes to Organ Dysfunction. Clin. Cancer Res. 2018, 24, 6583–6593. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Area | Mouse Strain | Age at Radiation | Time Post-Radiation | Radiation | Ovarian Reserve | Ovarian Follicular Development and Atresia | Stroma | Reference | ||
|---|---|---|---|---|---|---|---|---|---|---|
| Source | Rate Dose | Total Dose | ||||||||
| Total Body Irradiation (TBI) | BALB/c | 5 d | 5 d | γ-ray | 141 rad/min | 0.1 Gy | Similar primordial follicles | Similar follicular development | ND | [37] |
| 0.45 Gy | Near to depletion of primordial follicles | Near to depletion of small primary; presence of secondary follicles | ||||||||
| C57BL/6 | 5 d | 5 d | NI | 0.45 Gy | Depletion of primordial follicles; no follicular renewal | Near to depletion of primary follicles; presence of secondary follicles | [38] | |||
| CD1 | 5 d | 6 h | 2.387 Gy/min | 0.52 Gy | ND | ND | [39] | |||
| NMRI | 8 d | 2 d | X- or γ-ray | γ-ray: 35.57 mGy/min X-ray: 0.6 Gy/min | 0.02 Gy | = Or < primordial follicles | = Growing follicles | [40] | ||
| 0.1 Gy | < Primordial follicles | |||||||||
| 0.5 Gy | Depletion of primordial follicles | |||||||||
| BALB/c | 5 w | 2 w | γ-ray | 2.1 Gy/min for 0.48 min | 1 Gy | Near to depletion of primordial follicles | < Primary, secondary, and antral follicles persistence of secondary and antral in 129 > in BALB/c mice | No fibrosis < Spp1 | [41] | |
| 129 | 2 w | No fibrosis = Spp1 | ||||||||
| CD1 | 6 w | 2 w | 2.1 Gy/min for 48 s | 0.1 Gy | = Primordial follicles | = Follicular development; = Proliferating and apoptotic cells in follicles and corpora lutea | No fibrosis; = ovarian vasculature; = apoptotic and proliferating cells | [42] | ||
| 1 Gy | Depletion of primordial follicles | Near to depletion of primary follicles; < secondary and antral follicles; = early antral follicles; > proliferating and = apoptotic cells in follicles and corpora lutea | ||||||||
| 5 w | 0.1 Gy | = Primordial follicles | = Follicular development; = proliferating and apoptotic cells in follicles and corpora lutea | |||||||
| 1 Gy | Depletion of primordial follicles | Near to depletion of total follicles; no proliferating and = apoptotic cells in follicles and corpora lutea | ||||||||
| NI | 6 w | 2 w | NI | NI | 0.1 Gy | Near to depletion of primordial follicles | < Primary follicle; = pre-antral and antral follicles | ND | [43] | |
| Targeted Irradiation | CD1 | 6 w | 2 w | X-ray | 0.037 Gy/s for 27 s | < Primordial follicles | < growing follicles; = antral follicles | ND | [44] | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Reiser, E.; Bazzano, M.V.; Solano, M.E.; Haybaeck, J.; Schatz, C.; Mangesius, J.; Ganswindt, U.; Toth, B. Unlaid Eggs: Ovarian Damage after Low-Dose Radiation. Cells 2022, 11, 1219. https://doi.org/10.3390/cells11071219
Reiser E, Bazzano MV, Solano ME, Haybaeck J, Schatz C, Mangesius J, Ganswindt U, Toth B. Unlaid Eggs: Ovarian Damage after Low-Dose Radiation. Cells. 2022; 11(7):1219. https://doi.org/10.3390/cells11071219
Chicago/Turabian StyleReiser, Elisabeth, Maria Victoria Bazzano, Maria Emilia Solano, Johannes Haybaeck, Christoph Schatz, Julian Mangesius, Ute Ganswindt, and Bettina Toth. 2022. "Unlaid Eggs: Ovarian Damage after Low-Dose Radiation" Cells 11, no. 7: 1219. https://doi.org/10.3390/cells11071219
APA StyleReiser, E., Bazzano, M. V., Solano, M. E., Haybaeck, J., Schatz, C., Mangesius, J., Ganswindt, U., & Toth, B. (2022). Unlaid Eggs: Ovarian Damage after Low-Dose Radiation. Cells, 11(7), 1219. https://doi.org/10.3390/cells11071219