
Comparative Proteomic Profiling of Ectosomes Derived from Thyroid Carcinoma and Normal Thyroid Cells Uncovers Multiple Proteins with Functional Implications in Cancer

 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Cell Lines and Cell Culture Conditions
2.3. Isolation of Ectosomes
2.4. Characterization of Ectosome Samples
2.5. LC–MS/MS Proteomic Analysis
2.5.1. Sample Preparation for Mass Spectrometric Analysis
2.5.2. Liquid Chromatography and Tandem Mass Spectrometry (LC–MS/MS)
2.5.3. Analysis of Proteomic Data
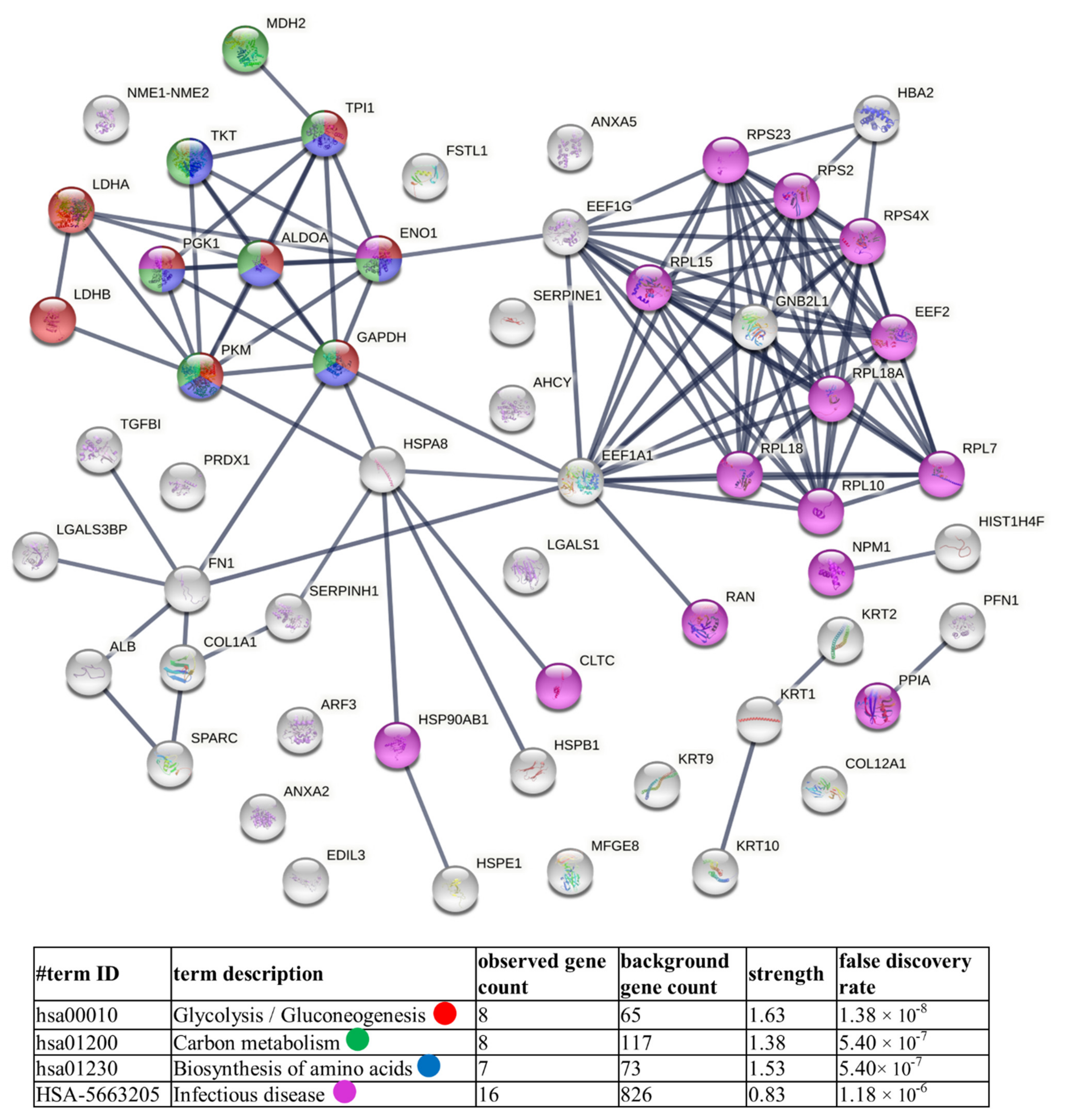
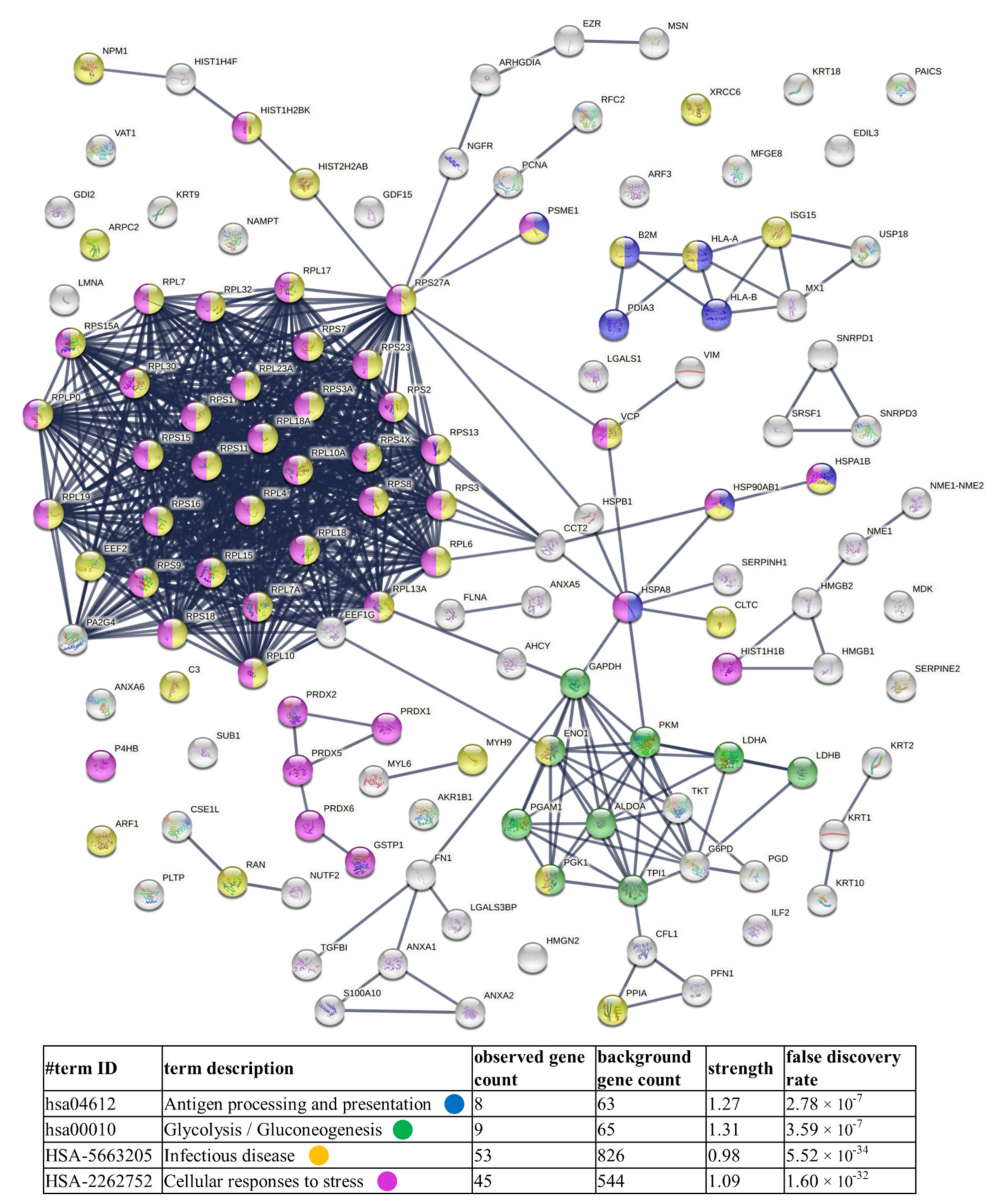
2.6. Bioinformatic Analysis
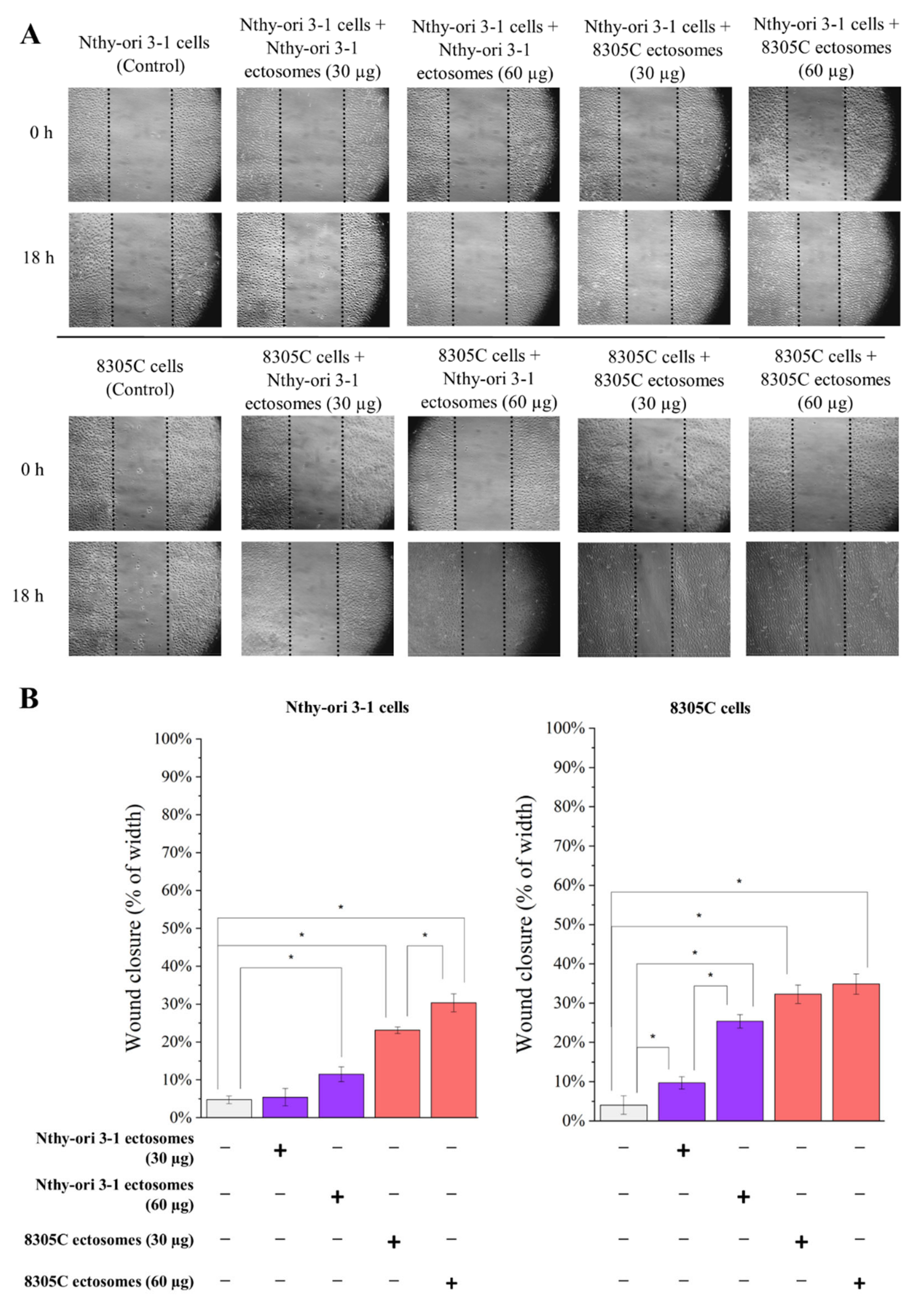
2.7. Wound Closure Assay
2.8. Alamar Blue Cell Viability Assay
2.9. Statistical Analysis
3. Results
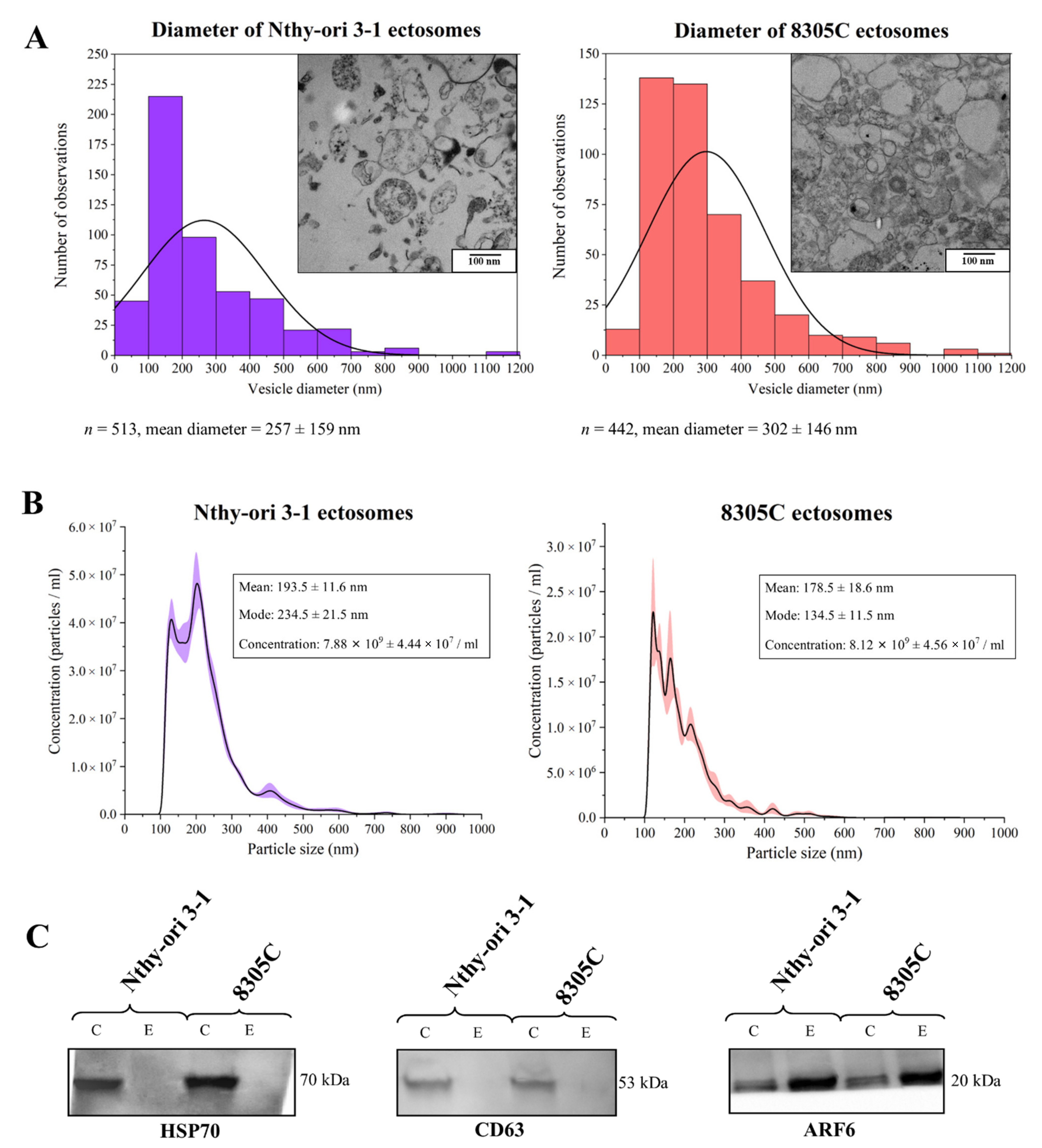
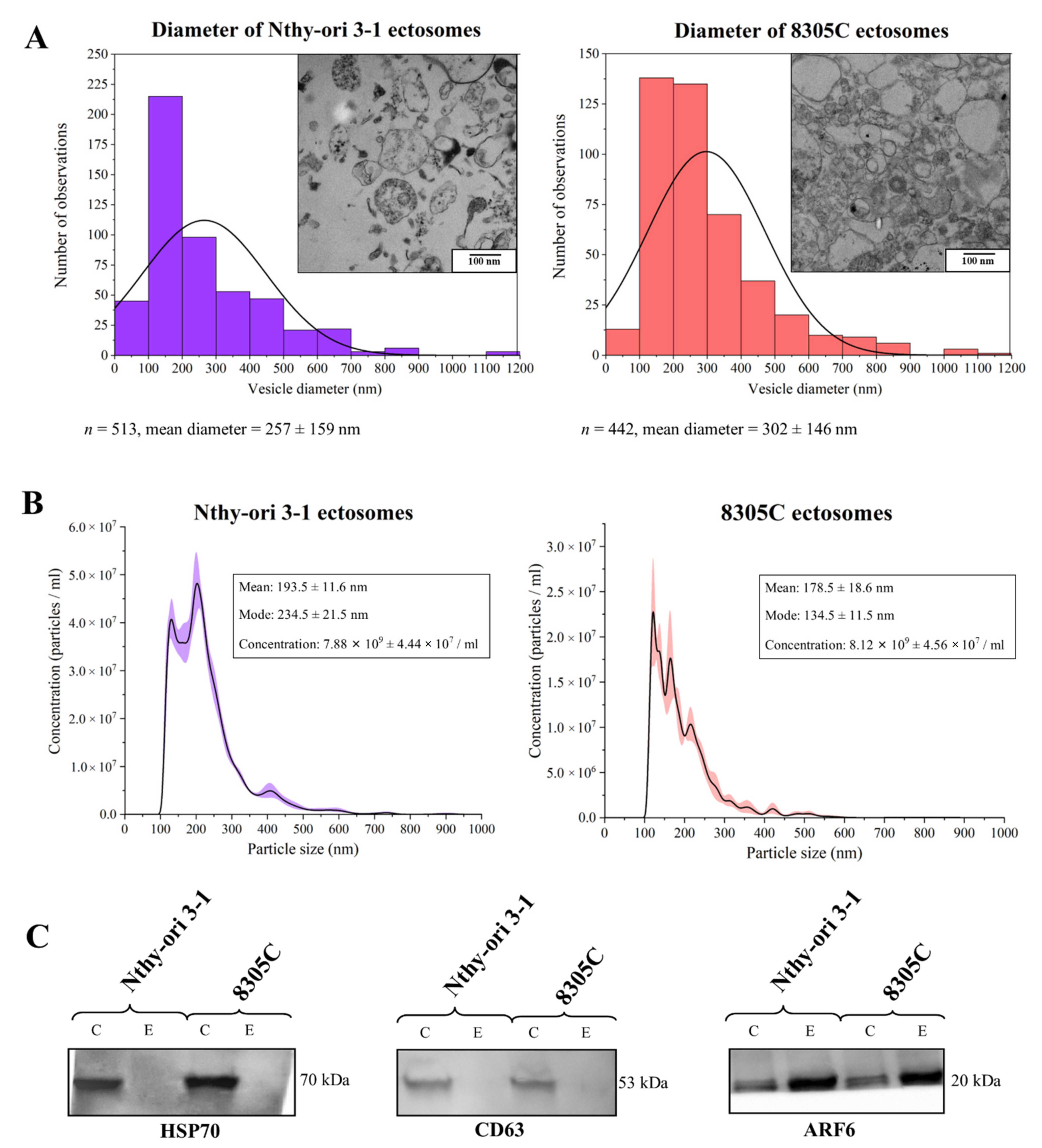
3.1. Purity of Ectosome Samples
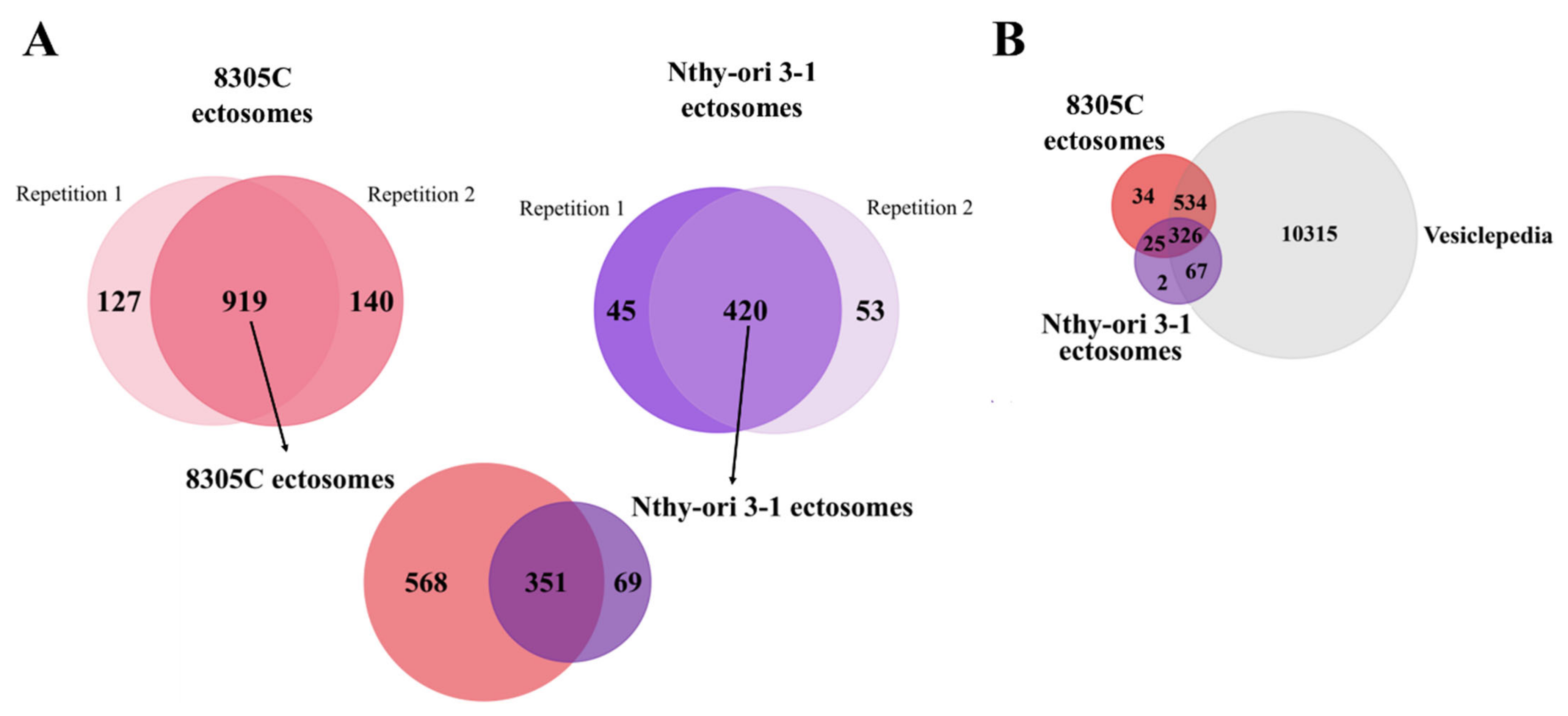
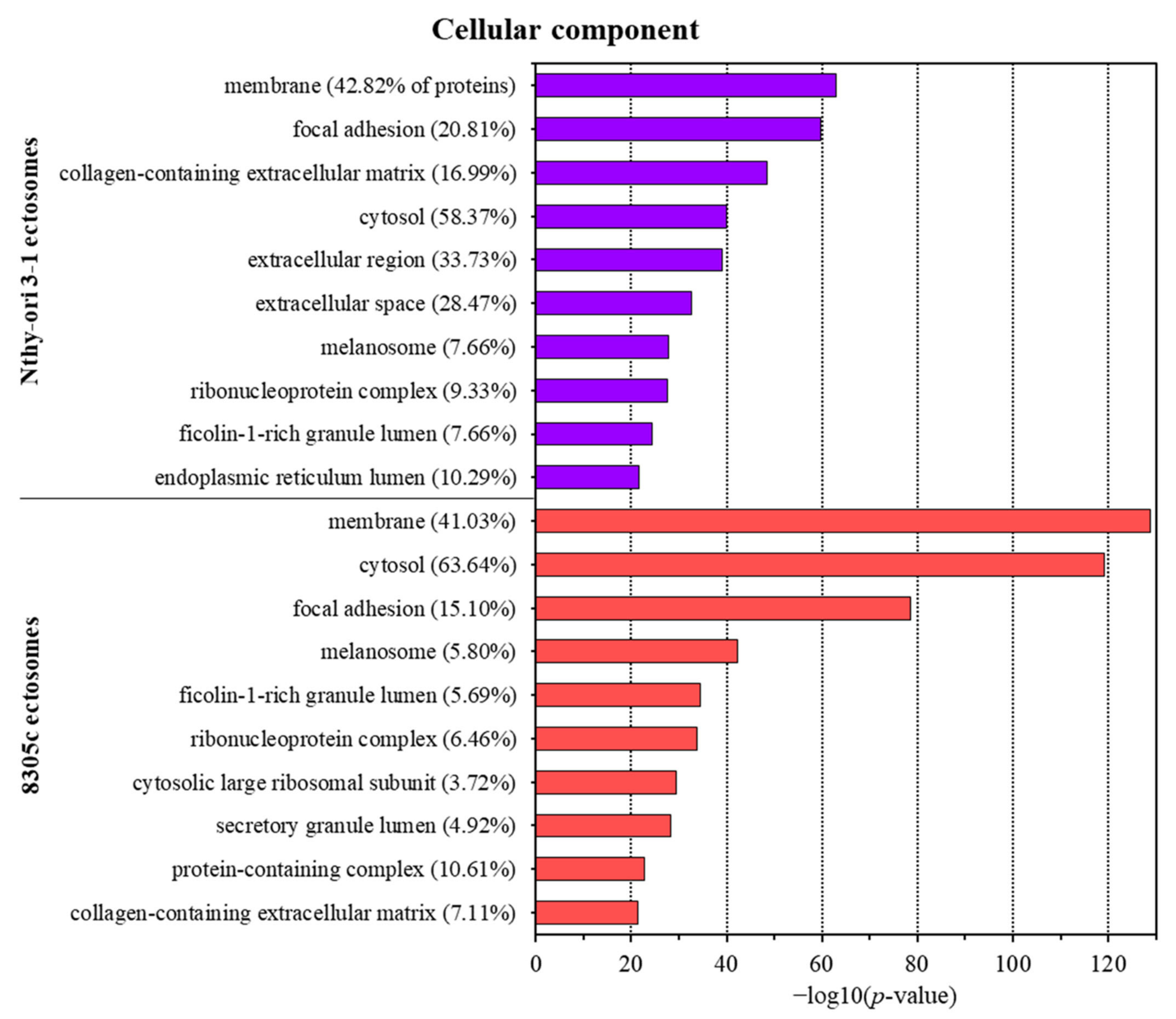
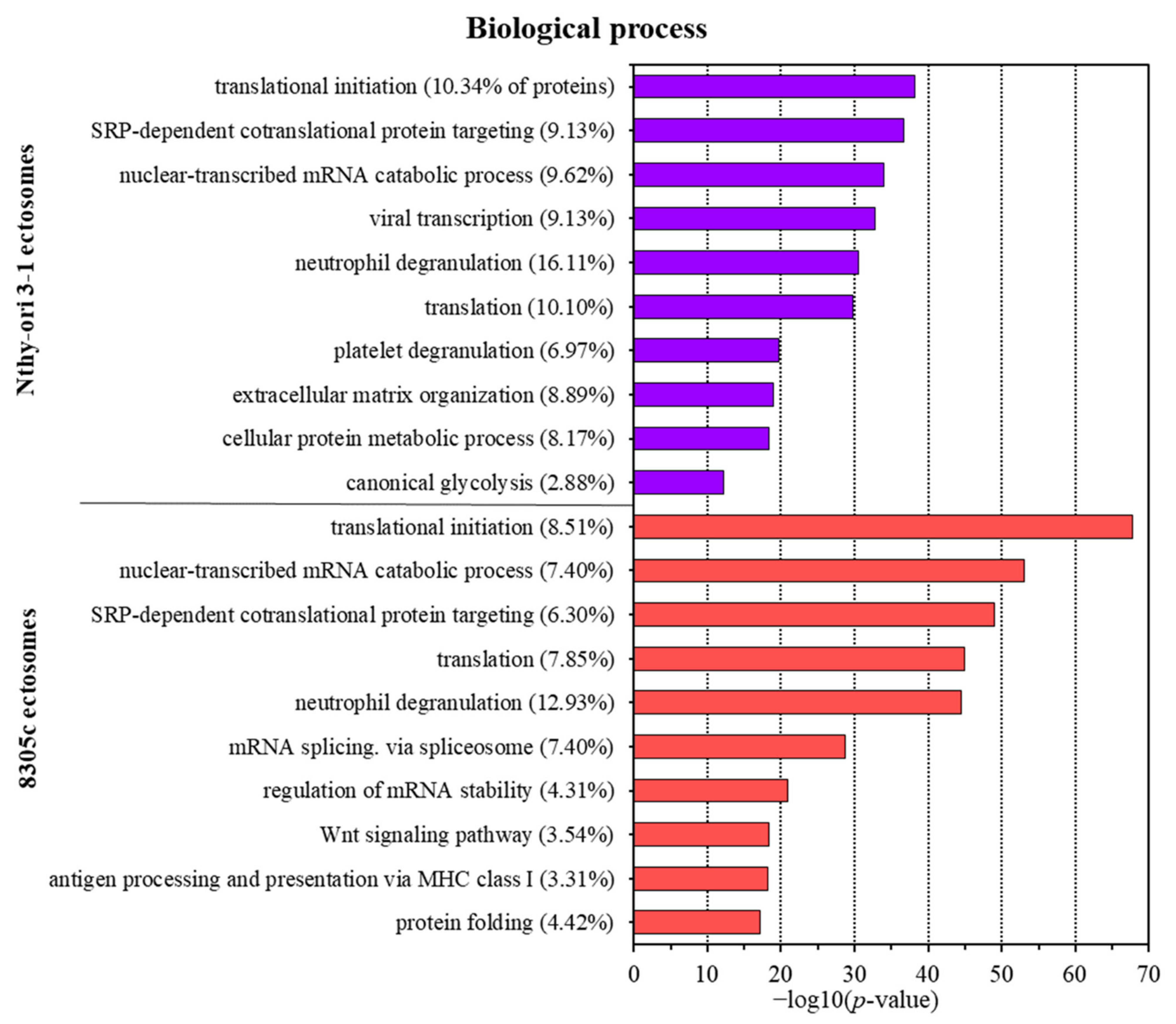
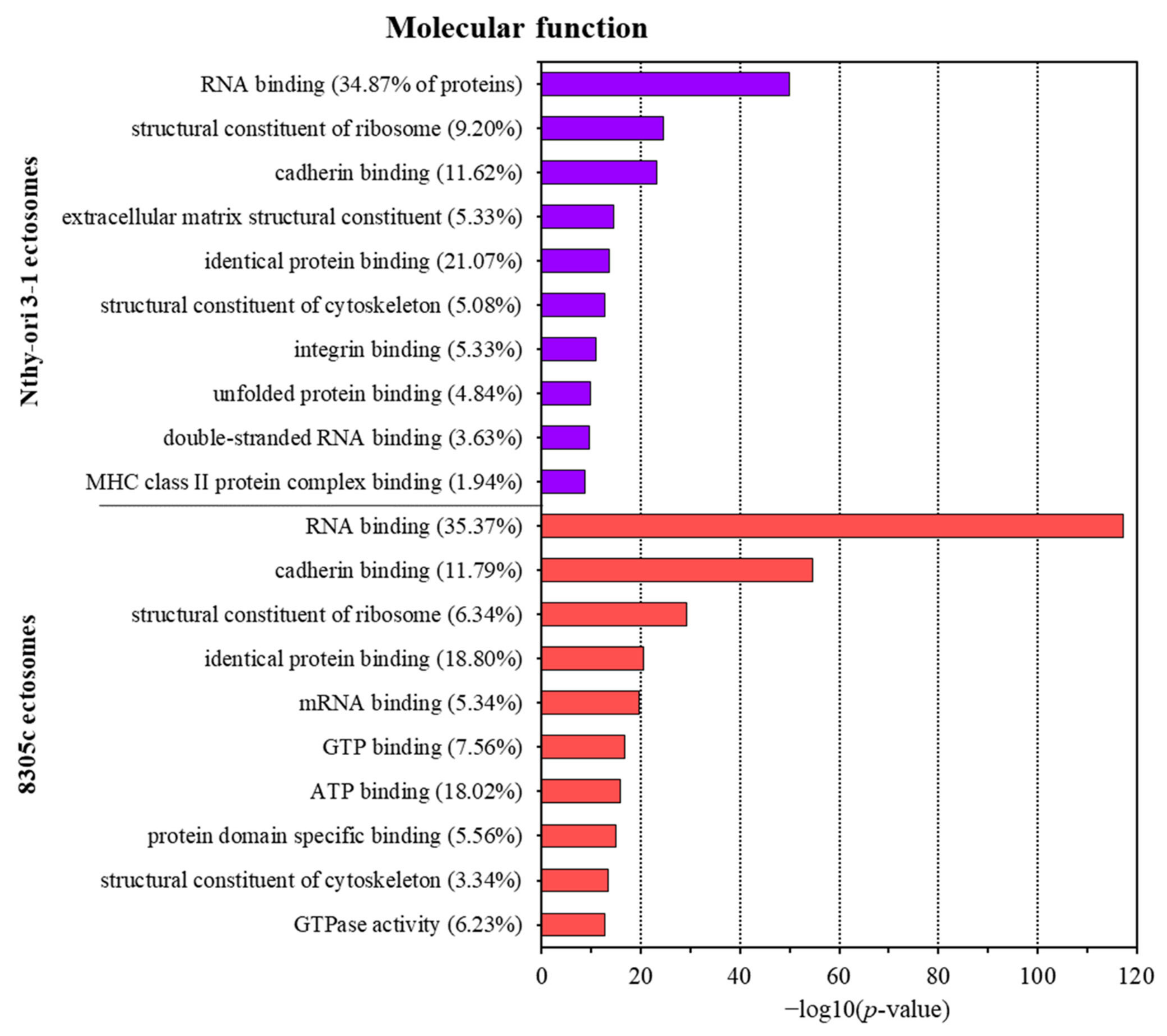
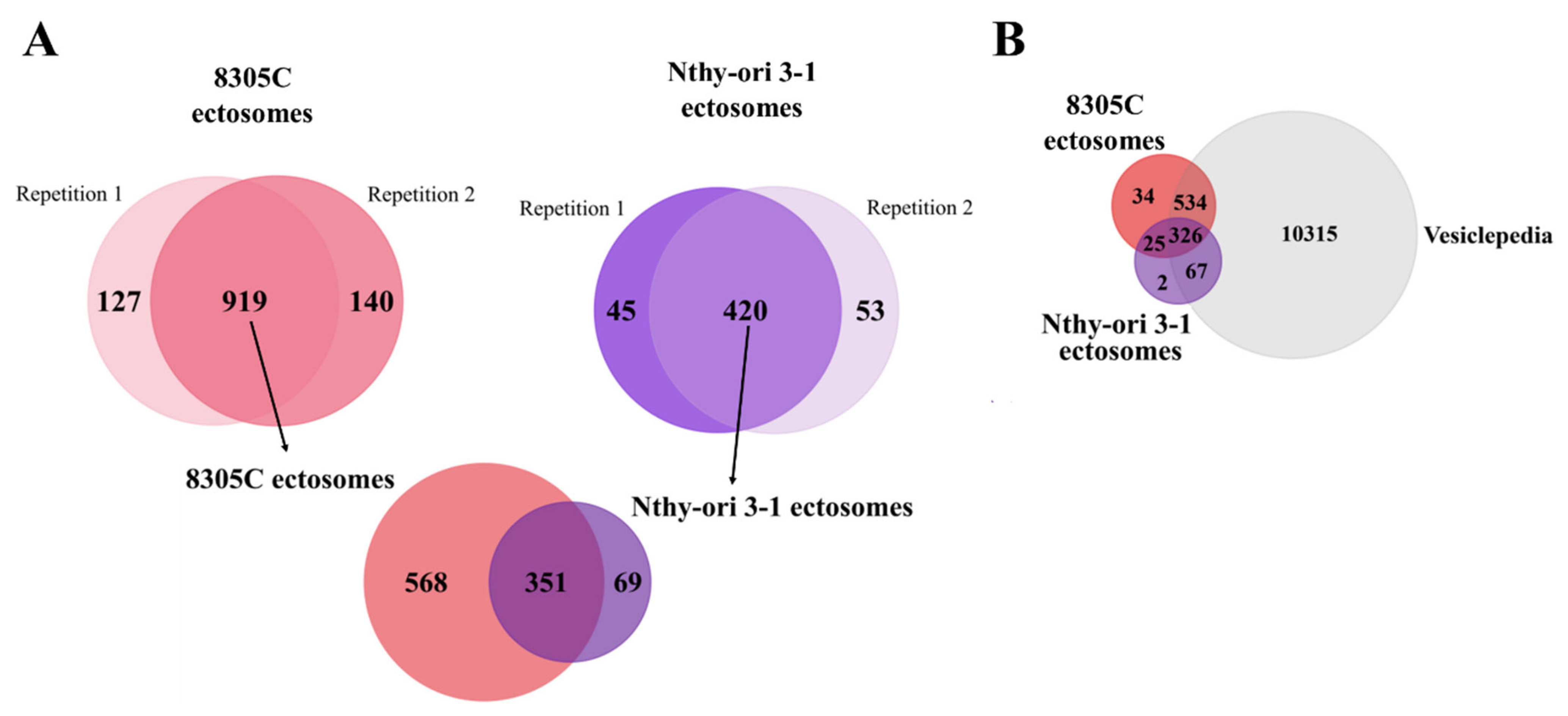
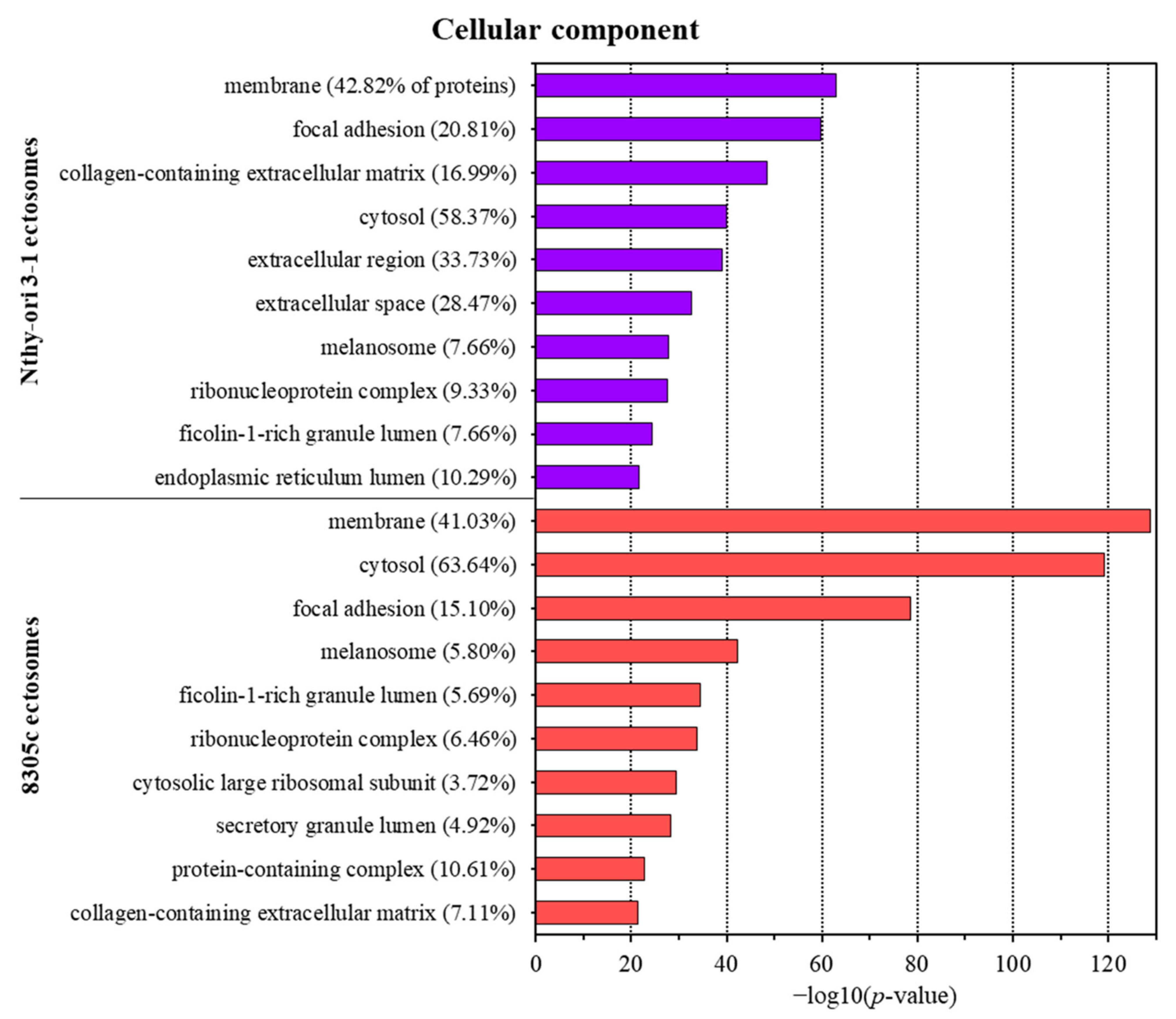
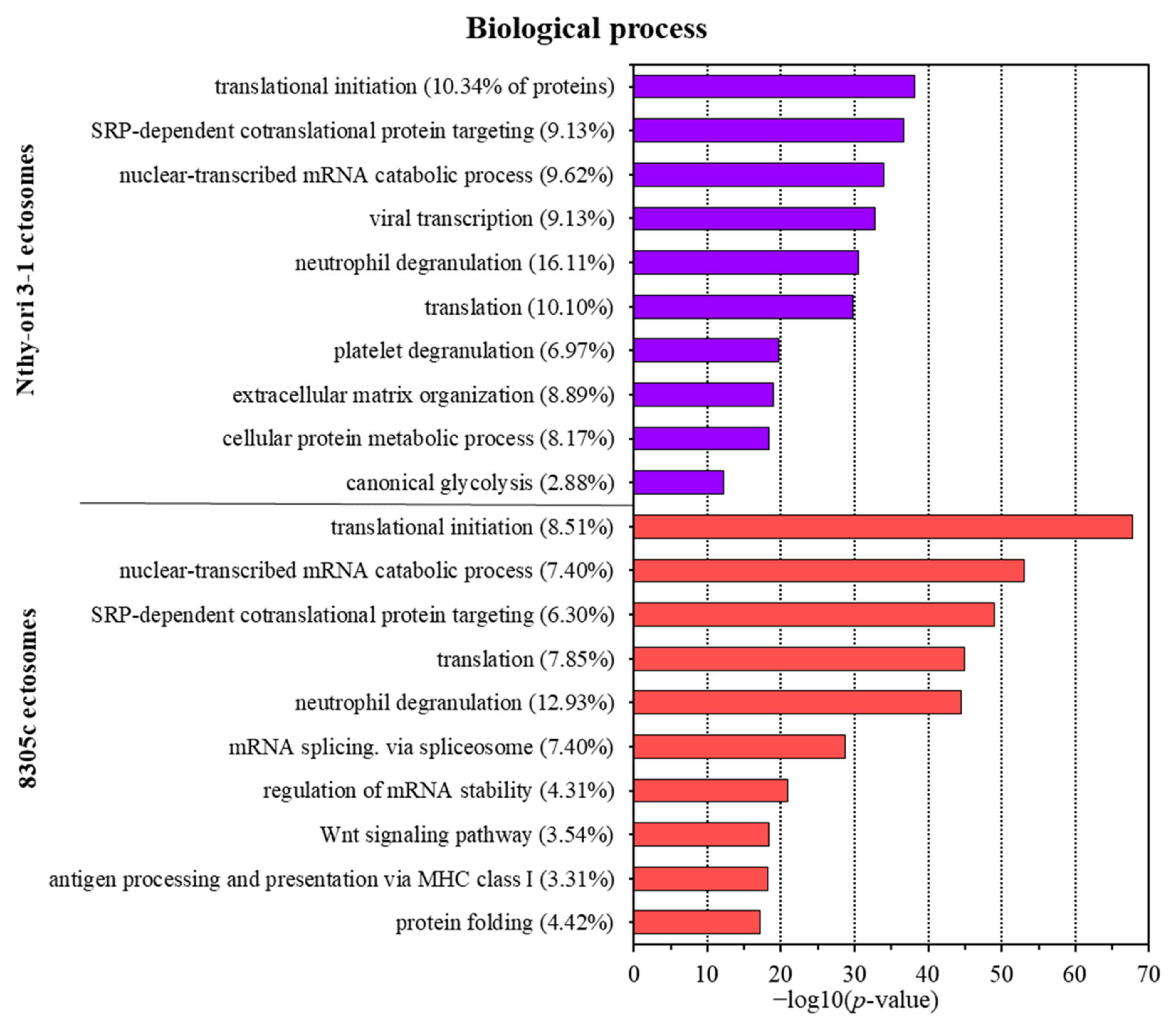
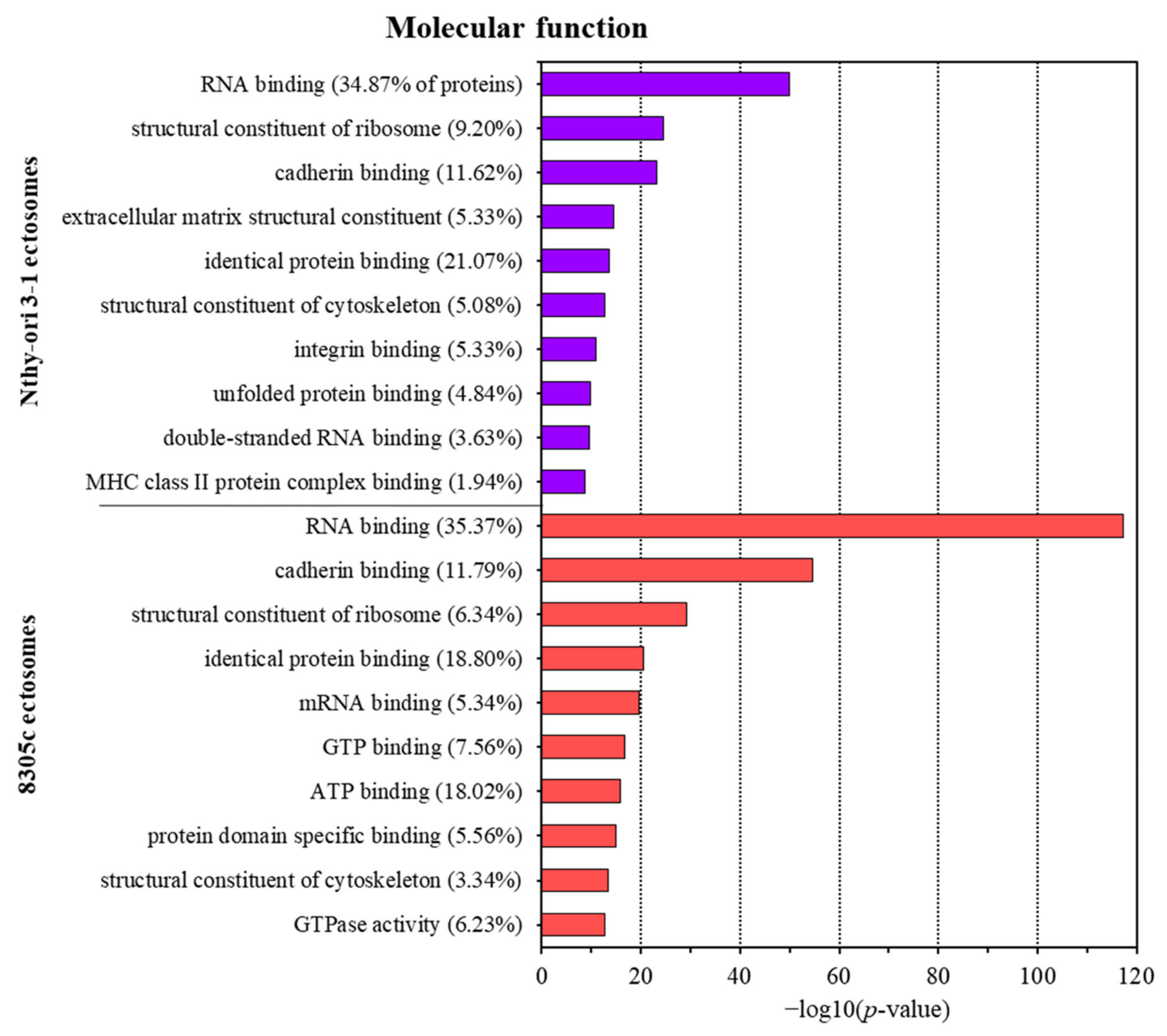
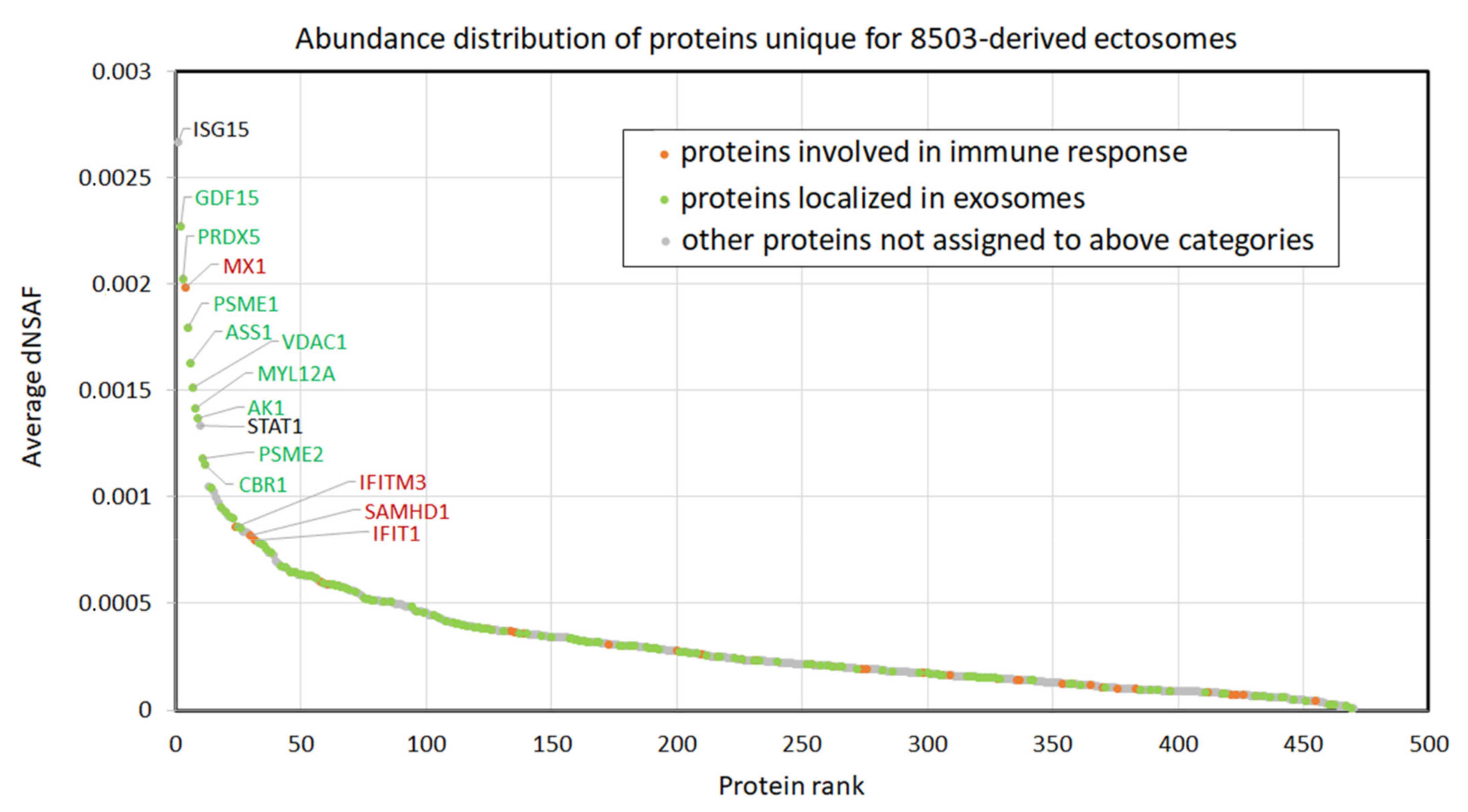
3.2. Proteins Identified in Nthy-ori 3-1- and 8305C-Derived Ectosomes
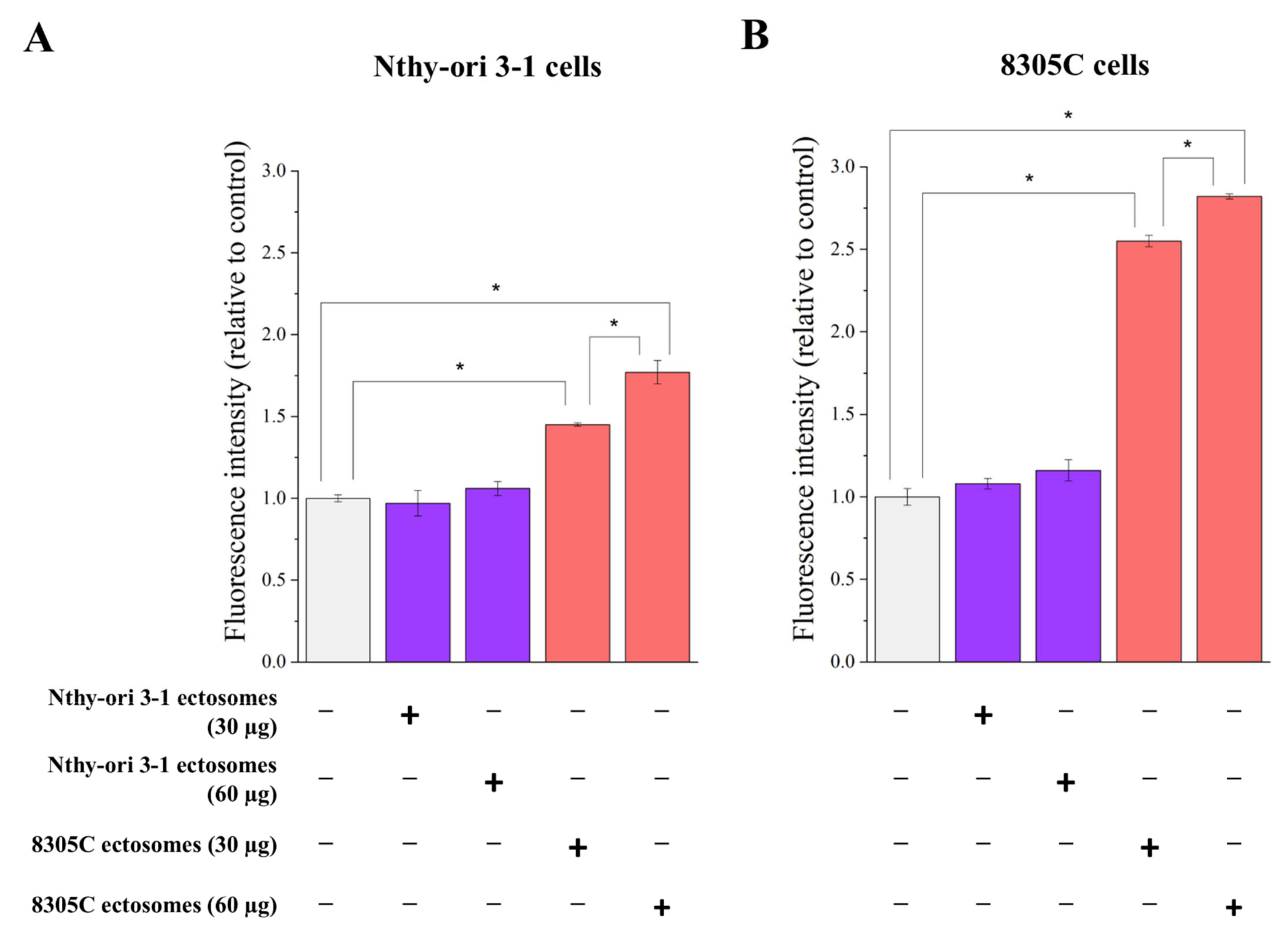
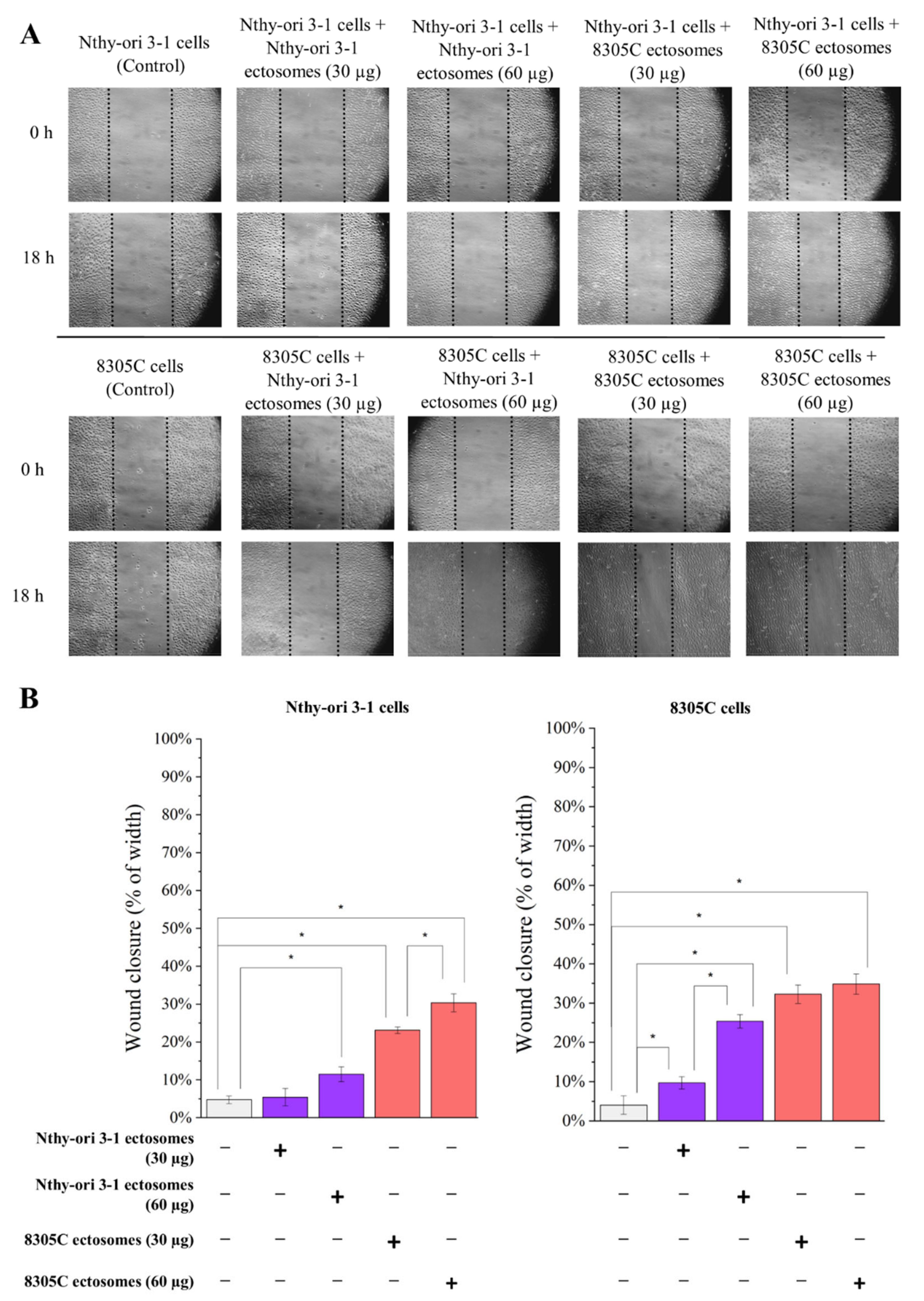
3.3. Functional Effect of Nthy-ori 3-1- and 8305C-Derived Ectosomes on Recipient Cells
4. Discussion
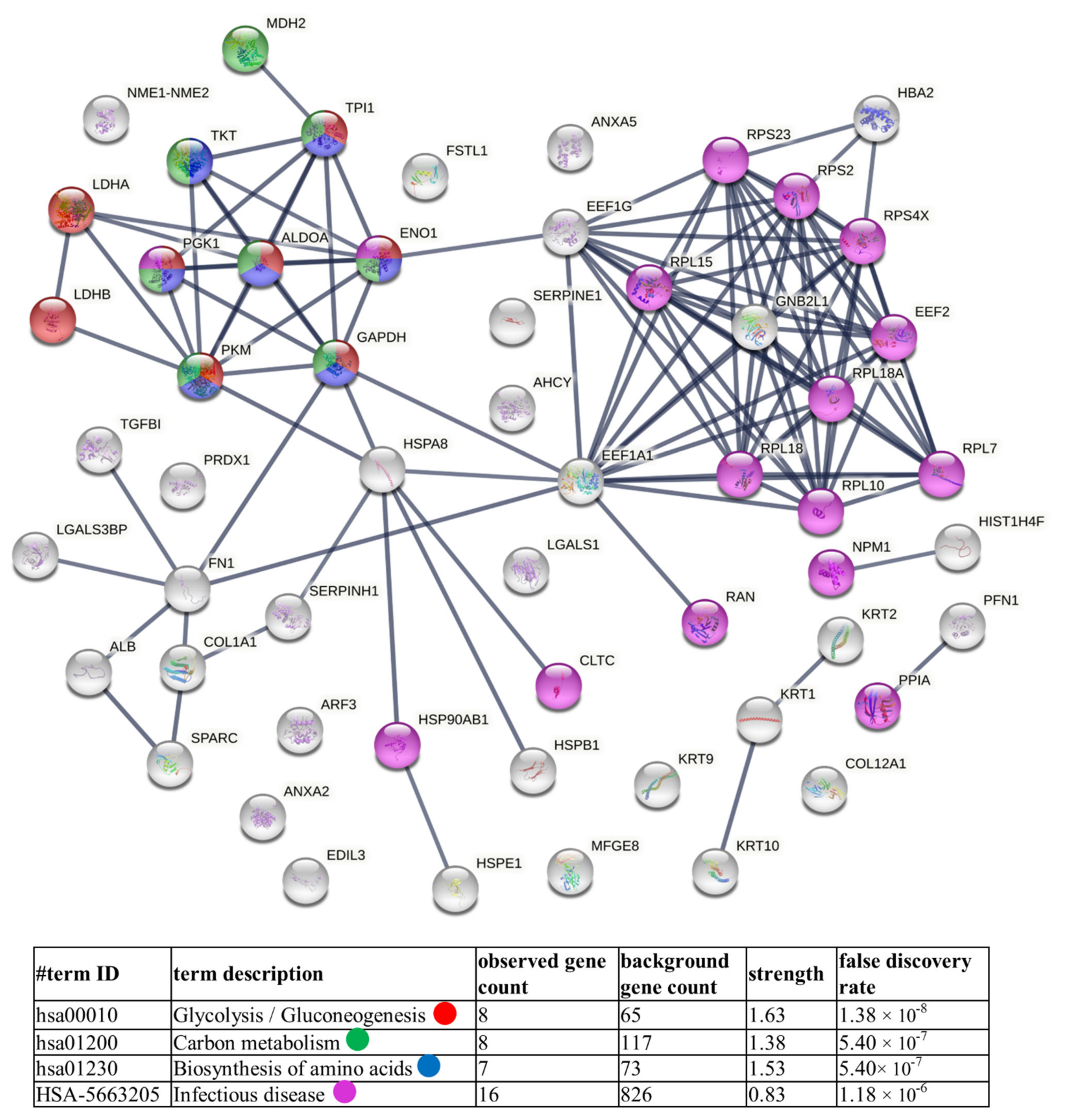
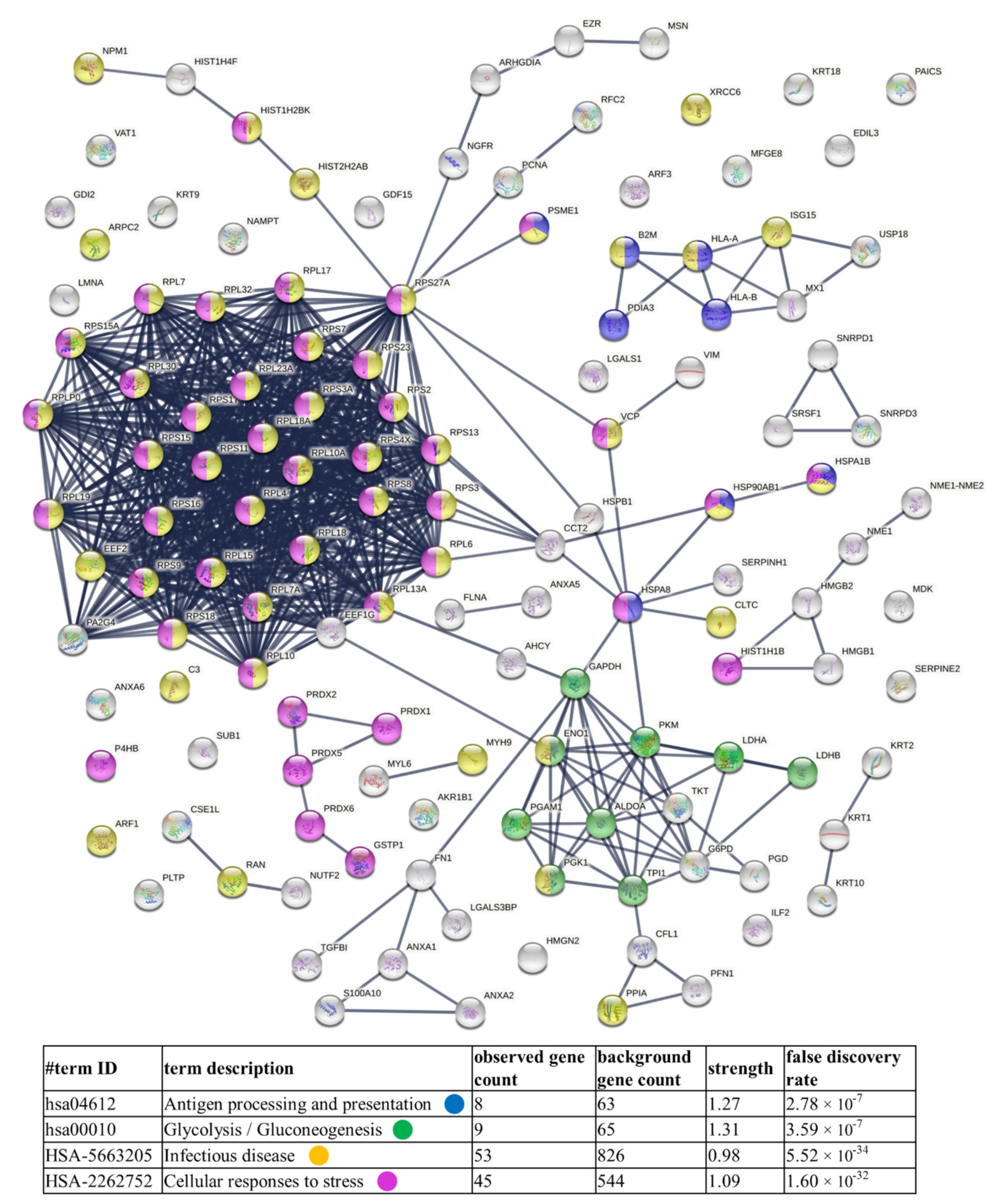
4.1. TC-Derived Ectosomes Are Enriched in Proteins Involved in Cancer Development and Progression
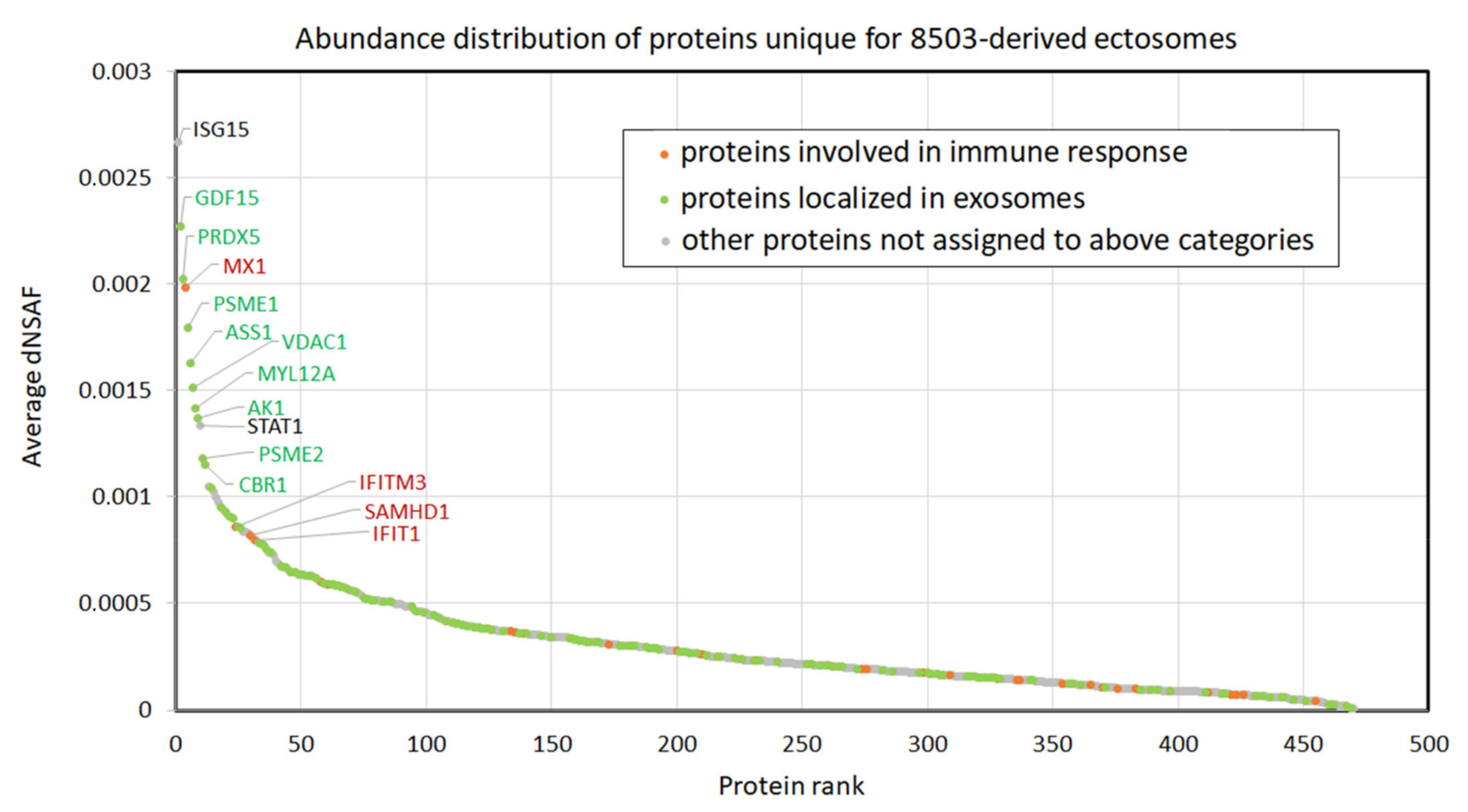
4.2. Clinical Relevance of Proteins Identified in TC-Derived Ectosomes
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ANXA1 | annexin A1 |
| dNSAF | normalized spectral counting-based abundance factor |
| EVs | extracellular vesicles |
| GDF15 | growth differentiation factor 15 |
| GO | gene ontology |
| IFIT1/3 | interferon-induced protein with tetratricopeptide repeats 1/3 |
| IFITM3 | interferon-induced transmembrane protein 3 |
| ISG15 | ubiquitin cross-reactive protein |
| LC–MS/MS | liquid chromatography coupled to tandem mass spectrometry |
| MX1 | GTP-binding protein Mx1 |
| NTA | nanoparticle tracking analysis |
| OAS3 | 2′-5′-oligoadenylate synthetase 3 |
| PD-1 | programmed death protein 1 |
| PD-L1 | programmed death ligand 1 |
| SAMHD1 | deoxynucleoside triphosphate triphosphohydrolase |
| STAT1 | signal transducer and activator of transcription 1-alpha/beta |
| TC | thyroid carcinoma |
| TEM | transmission electron microscopy |
References
- Clancy, J.; D’Souza-Schorey, C. Extracellular Vesicles in Cancer: Purpose and Promise. Cancer J. 2018, 24, 65–69. [Google Scholar] [CrossRef] [PubMed]
- Bebelman, M.P.; Smit, M.J.; Pegtel, D.M.; Baglio, S.R. Biogenesis and function of extracellular vesicles in cancer. Pharm. 2018, 188, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Maacha, S.; Bhat, A.A.; Jimenez, L.; Raza, A.; Haris, M.; Uddin, S.; Grivel, J.C. Extracellular vesicles-mediated intercellular communication: Roles in the tumor microenvironment and anti-cancer drug resistance. Mol. Cancer 2019, 18, 55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalluri, R.; LeBleu, V.S. The biology, function, and biomedical applications of exosomes. Science 2020, 367, eaau6977. [Google Scholar] [CrossRef] [PubMed]
- Urabe, F.; Kosaka, N.; Ito, K.; Kimura, T.; Egawa, S.; Ochiya, T. Extracellular vesicles as biomarkers and therapeutic targets for cancer. Am. J. Physiol Cell Physiol 2020, 318, C29–C39. [Google Scholar] [CrossRef] [PubMed]
- Möller, A.; Lobb, R.J. The evolving translational potential of small extracellular vesicles in cancer. Nat. Rev. Cancer 2020, 20, 697–709. [Google Scholar] [CrossRef]
- Hu, T.; Wolfram, J.; Srivastava, S. Extracellular Vesicles in Cancer Detection: Hopes and Hypes. Trends Cancer 2021, 7, 122–133. [Google Scholar] [CrossRef]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef]
- Rappa, G.; Puglisi, C.; Santos, M.F.; Forte, S.; Memeo, L.; Lorico, A. Extracellular Vesicles from Thyroid Carcinoma: The New Frontier of Liquid Biopsy. Int J. Mol. Sci 2019, 20, 1114. [Google Scholar] [CrossRef] [Green Version]
- Lu, Y.; Lan, N.; Zhang, Y.; Nie, X.; Pu, S.; Yuan, W. Role of extracellular vesicles in the progression.; diagnosis and treatment of thyroid cancer (Review). Int J. Oncol 2020, 57, 881–889. [Google Scholar] [CrossRef]
- Luo, D.; Zhan, S.; Xia, W.; Huang, L.; Ge, W.; Wang, T. Proteomics study of serum exosomes from papillary thyroid cancer patients. Endocr. Relat. Cancer 2018, 25, 879–891. [Google Scholar] [CrossRef] [PubMed]
- Huang, T.Y.; Wang, C.Y.; Chen, K.Y.; Huang, L.T. Urinary Exosomal Thyroglobulin in Thyroid Cancer Patients With Post-ablative Therapy: A New Biomarker in Thyroid Cancer. Front. Endocrinol. 2020, 11, 382. [Google Scholar] [CrossRef]
- Caruso Bavisotto, C.; Cipolla, C.; Graceffa, G.; Barone, R.; Bucchieri, F.; Bulone, D.; Cabibi, D.; Campanella, C.; Marino Gammazza, A.; Pitruzzella, A.; et al. Immunomorphological Pattern of Molecular Chaperones in Normal and Pathological Thyroid Tissues and Circulating Exosomes: Potential Use in Clinics. Int. J. Mol. Sci. 2019, 20, 4496. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, G.; He, L.; Wang, S.; Zhang, M.; Li, Y.; Liu, Q.; Sun, N.; Zhang, X.; Liu, Y.; Zhang, J.; et al. EV PD-L1 is Correlated With Clinical Features and Contributes to T Cell Suppression in Pediatric Thyroid Cancer. J. Clin. Endocrinol. Metab. 2020, 105, dgaa309. [Google Scholar] [CrossRef] [PubMed]
- Bravo-Miana, R.D.C.; Della Vedova, A.B.; De Paul, A.L.; Remedi, M.M.; Guantay, M.L.; Gilardoni, M.B.; Pellizas, C.G.; Donadio, A.C. Thyroid tumor cells-fibroblasts crosstalk: Role of extracellular vesicles. Endocr. Connect. 2020, 9, 506–518. [Google Scholar] [CrossRef] [PubMed]
- Giusti, I.; D’Ascenzo, S.; Dolo, V. Microvesicles as potential ovarian cancer biomarkers. Biomed. Res. Int. 2013, 2013, 703048. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohan, A.; Agarwal, S.; Clauss, M.; Britt, N.S.; Dhillon, N.K. Extracellular vesicles: Novel communicators in lung diseases. Respir. Res. 2020, 21, 175. [Google Scholar] [CrossRef]
- Menck, K.; Sivaloganathan, S.; Bleckmann, A.; Binder, C. Microvesicles in Cancer: Small Size.; Large Potential. Int. J. Mol. Sci. 2020, 21, 5373. [Google Scholar] [CrossRef]
- Surman, M.; Hoja-Łukowicz, D.; Szwed, S.; Kędracka-Krok, S.; Jankowska, U.; Kurtyka, M.; Drożdż, A.; Lityńska, A.; Stępień, E.; Przybyło, M. An Insight into the Proteome of Uveal Melanoma-Derived Ectosomes Reveals the Presence of Potentially Useful Biomarkers. Int. J. Mol. Sci. 2019, 20, 3789. [Google Scholar] [CrossRef] [Green Version]
- Surman, M.; Kędracka-Krok, S.; Hoja-Łukowicz, D.; Jankowska, U.; Drożdż, A.; Stępień, E.Ł.; Przybyło, M. Mass Spectrometry-Based Proteomic Characterization of Cutaneous Melanoma Ectosomes Reveals the Presence of Cancer-Related Molecules. Int. J. Mol. Sci. 2020, 21, 2934. [Google Scholar] [CrossRef] [Green Version]
- Lemoine, N.R.; Mayall, E.S.; Jones, T.; Sheer, D.; McDermid, S.; Kendall-Taylor, P.; Wynford-Thomas, D. Characterisation of human thyroid epithelial cells immortalised in vitro by simian virus 40 DNA transfection. Br. J. Cancer 1989, 60, 897–903. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Landa, I.; Pozdeyev, N.; Korch, C.; Marlow, L.A.; Smallridge, R.C.; Copland, J.A.; Henderson, Y.C.; Lai, S.Y.; Clayman, G.L.; Onoda, N.; et al. Comprehensive Genetic Characterization of Human Thyroid Cancer Cell Lines: A Validated Panel for Preclinical Studies. Clin. Cancer Res. 2019, 25, 3141–3151. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.A.; Jee, H.G.; Yi, J.W.; Kim, S.J.; Chai, Y.J.; Choi, J.Y.; Lee, K.E. Expression Profiling of a Human Thyroid Cell Line Stably Expressing the BRAFV600E Mutation. Cancer Genom. Proteom. 2017, 14, 53–67. [Google Scholar] [CrossRef] [Green Version]
- Drożdż, A.; Kamińska, A.; Surman, M.; Gonet-Surówka, A.; Jach, R.; Huras, H.; Przybyło, M.; Stępień, E.Ł. Low-Vacuum Filtration as an Alternative Extracellular Vesicle Concentration Method: A Comparison with Ultracentrifugation and Differential Centrifugation. Pharmaceutics 2020, 12, 872. [Google Scholar] [CrossRef]
- Surman, M.; Hoja-Łukowicz, D.; Szwed, S.; Drożdż, A.; Stępień, E.; Przybyło, M. Human melanoma-derived ectosomes are enriched with specific glycan epitopes. Life Sci. 2018, 207, 395–411. [Google Scholar] [CrossRef] [PubMed]
- Surman, M.; Kędracka-Krok, S.; Jankowska, U.; Drożdż, A.; Stępień, E.; Przybyło, M. Proteomic Profiling of Ectosomes Derived from Paired Urothelial Bladder Cancer and Normal Cells Reveals the Presence of Biologically-Relevant Molecules. Int. J. Mol. Sci. 2021, 22, 6816. [Google Scholar] [CrossRef] [PubMed]
- Käll, L.; Canterbury, J.D.; Weston, J.; Noble, W.S.; MacCoss, M.J. Semi-supervised learning for peptide identification from shotgun proteomics datasets. Nat. Methods 2007, 4, 923–925. [Google Scholar] [CrossRef]
- Vizcaíno, J.A.; Deutscch, E.W.; Wang, R.; Csordas, A.; Reisinger, F.; Ríos, D.; Dianes, J.A.; Sun, Z.; Farrah, T.; Bandeira, N.; et al. ProteomeXchange provides globally coordinated proteomics data submission and dissemination. Nat. Biotechnol. 2014, 32, 223–226. [Google Scholar] [CrossRef]
- Zhang, Y.; Wen, Z.; Washburn, M.P.; Florens, L. Refinements to label free proteome quantitation: How to deal with peptides shared by multiple proteins. Anal. Chem. 2010, 82, 2272–2281. [Google Scholar] [CrossRef]
- Wang, P.; Jing, Z.; Liu, C.; Xu, M.; Wang, P.; Wang, X.; Yin, Y.; Cui, Y.; Ren, D.; Rao, X. Hepatitis C virus infection and risk of thyroid cancer: A systematic review and meta-analysis. Arab. J. Gastroenterol. 2017, 18, 1–5. [Google Scholar] [CrossRef]
- Hobbs, J.A.; Adamson-Small, L.A. Parvovirus and thyroid cancer. Semin. Oncol. 2015, 42, 304–308. [Google Scholar] [CrossRef] [PubMed]
- Stamatiou, D.; Derdas, S.P.; Symvoulakis, E.K.; Sakorafas, G.H.; Zoras, O.; Spandidos, D.A. Investigation of BK virus.; Epstein-Barr virus and human papillomavirus sequences in postoperative thyroid gland specimens. Int. J. Biol. Markers 2015, 30, e104–e110. [Google Scholar] [CrossRef] [PubMed]
- Moghoofei, M.; Mostafaei, S.; Nesaei, A.; Etemadi, A.; Sadri Nahand, J.; Mirzaei, H.; Rashidi, B.; Babaei, F.; Khodabandehlou, N. Epstein-Barr virus and thyroid cancer: The role of viral expressed proteins. J. Cell Physiol. 2019, 234, 3790–3799. [Google Scholar] [CrossRef] [PubMed]
- Almeida, J.F.M.; Peres, K.C.; Teixeira, E.S.; Teodoro, L.; Bó, I.F.D.; Ward, L.S. Epstein-Barr Virus and Thyroid Cancer. Crit. Rev. Oncog. 2019, 24, 369–377. [Google Scholar] [CrossRef]
- Li, Q.; Liu, W.; Wang, Z.; Wang, C.; Ai, Z. Exosomal ANXA1 derived from thyroid cancer cells is associated with malignant transformation of human thyroid follicular epithelial cells by promoting cell proliferation. Int. J. Oncol. 2021, 59, 104. [Google Scholar] [CrossRef]
- Foo, S.L.; Yap, G.; Cui, J.; Lim, L.H.K. Annexin-A1 – A Blessing or a Curse in Cancer? Trends Mol. Med. 2019, 25, 315–327. [Google Scholar] [CrossRef]
- Kang, Y.E.; Kim, J.M.; Lim, M.A.; Lee, S.E.; Yi, S.; Kim, J.T.; Oh, C.; Liu, L.; Jin, Y.; Jung, S.N.; et al. Growth Differentiation Factor 15 is a Cancer Cell-Induced Mitokine That Primes Thyroid Cancer Cells for Invasiveness. Thyroid 2021, 31, 772–786. [Google Scholar] [CrossRef]
- Wu, F.; Li, F.; Lin, X.; Xu, F.; Cui, R.R.; Zhong, J.Y.; Zhu, T.; Shan, S.K.; Liao, X.B.; Yuan, L.Q.; et al. Exosomes increased angiogenesis in papillary thyroid cancer microenvironment. Endocr. Relat. Cancer 2019, 26, 525–538. [Google Scholar] [CrossRef] [Green Version]
- Marech, I.; Leporini, C.; Ammendola, M.; Porcelli, M.; Gadaleta, C.D.; Russo, E.; De Sarro, G.; Ranieri, G. Classical and non-classical proangiogenic factors as a target of antiangiogenic therapy in tumor microenvironment. Cancer Lett. 2016, 380, 216–226. [Google Scholar] [CrossRef]
- Davis, P.J.; Mousa, S.A.; Schechter, G.P.; Lin, H.Y. Platelet ATP, Thyroid Hormone Receptor on Integrin alphavbeta3 and Cancer Metastasis. Horm. Cancer 2020, 11, 13–16. [Google Scholar] [CrossRef]
- Xi, C.; Zhang, G.Q.; Sun, Z.K.; Song, H.J.; Shen, C.T.; Chen, X.Y.; Sun, J.W.; Qiu, Z.L.; Luo, Q.Y. Interleukins in Thyroid Cancer: From Basic Researches to Applications in Clinical Practice. Front. Immunol. 2020, 11, 1124. [Google Scholar] [CrossRef] [PubMed]
- Gan, C.P.; Sam, K.K.; Yee, P.S.; Zainal, N.S.; Lee, B.K.B.; Abdul Rahman, Z.A.; Patel, V.; Tan, A.C.; Zain, R.B.; Cheong, S.C. IFITM3 knockdown reduces the expression of CCND1 and CDK4 and suppresses the growth of oral squamous cell carcinoma cells. Cell Oncol. 2019, 42, 477–490. [Google Scholar] [CrossRef]
- Zhang, D.; Wang, H.; He, H.; Niu, H.; Li, Y. Interferon induced transmembrane protein 3 regulates the growth and invasion of human lung adenocarcinoma. Thorac Cancer 2017, 8, 337–343. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Yu, C. Prognostic characterization of OAS1/OAS2/OAS3/OASL in breast cancer. BMC Cancer 2020, 20, 575. [Google Scholar] [CrossRef] [PubMed]
- Li, H.P.; Yang, T.Z.; Wei, L.; Ge, Y.F.; Meng, Q.S. STAT1-induced upregulation of lncRNA LINP1 promotes cell proliferation and inhibits apoptosis via AMPK signaling pathway in papillary thyroid cancer. Eur. Rev. Med. Pharm. Sci. 2020, 24, 8911–8917. [Google Scholar] [CrossRef]
- Ren, R.; Du, Y.; Niu, X.; Zang, R. ZFPM2-AS1 transcriptionally mediated by STAT1 regulates thyroid cancer cell growth.; migration and invasion via miR-515-5p/TUSC3. J. Cancer 2021, 12, 3393–3406. [Google Scholar] [CrossRef] [PubMed]
- Lin, P.; Yao, Z.; Sun, Y.; Li, W.; Liu, Y.; Liang, K.; Liu, Y.; Qin, J.; Hou, X.; Chen, L. Deciphering novel biomarkers of lymph node metastasis of thyroid papillary microcarcinoma using proteomic analysis of ultrasound-guided fine-needle aspiration biopsy samples. J. Proteom. 2019, 204, 103414. [Google Scholar] [CrossRef]
- Sun, H.; Chen, Q.; Liu, W.; Liu, Y.; Ruan, S.; Zhu, C.; Ruan, Y.; Ying, S.; Lin, P. TROP2 modulates the progression in papillary thyroid carcinoma. J. Cancer 2021, 12, 6883–6893. [Google Scholar] [CrossRef]
- Van der Spek, A.H.; Fliers, E.; Boelen, A. Thyroid hormone metabolism in innate immune cells. J. Endocrinol. 2017, 232, R67–R81. [Google Scholar] [CrossRef]
- Galdiero, M.R.; Varricchi, G.; Loffredo, S.; Bellevicine, C.; Lansione, T.; Ferrara, A.L.; Iannone, R.; di Somma, S.; Borriello, F.; Clery, E.; et al. Potential involvement of neutrophils in human thyroid cancer. PLoS ONE 2018, 13, e019974. [Google Scholar] [CrossRef]
- Caruso Bavisotto, C.; Graziano, F.; Rappa, F.; Marino Gammazza, A.; Logozzi, M.; Fais, S.; Maugeri, R.; Bucchieri, F.; Conway de Macario, E.; Macario, A.J.L.; et al. Exosomal Chaperones and miRNAs in Gliomagenesis: State-of-Art and Theranostics Perspectives. Int. J. Mol. Sci. 2018, 19, 2626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lettini, G.; Pietrafesa, M.; Lepore, S.; Maddalena, F.; Crispo, F.; Sgambato, A.; Esposito, F.; Landriscina, M. Heat shock proteins in thyroid malignancies: Potential therapeutic targets for poorly-differentiated and anaplastic tumours? Mol. Cell Endocrinol. 2020, 502, 10676. [Google Scholar] [CrossRef] [PubMed]
- Gangadaran, P.; Li, X.J.; Kalimuthu, S.K.; Min, O.J.; Hong, C.M.; Rajendran, R.L.; Lee, H.W.; Zhu, L.; Baek, S.H.; Jeong, S.Y.; et al. New Optical Imaging Reporter-labeled Anaplastic Thyroid Cancer-Derived Extracellular Vesicles as a Platform for In Vivo Tumor Targeting in a Mouse Model. Sci. Rep. 2018, 8, 13509. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Kalimuthu, S.; Oh, J.M.; Gangadaran, P.; Baek, S.H.; Jeong, S.Y.; Lee, S.W.; Lee, J.; Ahn, B.C. Enhancement of antitumor potency of extracellular vesicles derived from natural killer cells by IL-15 priming. Biomaterials 2019, 190–191, 38–50. [Google Scholar] [CrossRef]
- Rajendran, R.L.; Paudel, S.; Gangadaran, P.; Oh, J.M.; Oh, E.J.; Hong, C.M.; Lee, S.; Chung, H.Y.; Lee, J.; Ahn, B.C. Extracellular Vesicles Act as Nano-Transporters of Tyrosine Kinase Inhibitors to Revert Iodine Avidity in Thyroid Cancer. Pharmaceutics 2021, 13, 248. [Google Scholar] [CrossRef]
- Piotrowska, U.; Adler, G.; Mackiewicz, U. TSH signal transduction in thyroid cells of Nthy-ori 3-1 line. Postępy Nauk Med. 2011, 24, 924–928. [Google Scholar]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell Proliferation (GO:0008283) | Cell Migration (GO:0016477) | Angiogenesis (GO:0001525) | Immune Response (GO:0006955) | Cellular Response to Drug (GO:0035690) |
|---|---|---|---|---|
| Cyclin-dependent kinase 1 (CDK1) Guanine nucleotide-binding protein G(i) subunit alpha-2 (GNAI2) Guanine nucleotide-binding protein G(I)/G(S)/G(T) subunit beta-1 (GNB1) H3 histone, family 3B (H3F3B ) Lysosomal acid lipase/cholesteryl ester hydrolase (LIPA) Minichromosome maintenance complex component 7 (MCM7) Myosin-10 (MYH10) Peroxiredoxin-1 (PRDX1) Protein scribble homolog (SCRIB) Proto-oncogene tyrosine protein kinase Src (SRC) Ras-related C3 botulinum toxin substrate 1 (RAC1) Ras-related protein Rap-1b (RAP1B) Thioredoxin reductase 1, cytoplasmic (TXNRD1) Transforming growth factor-beta-induced protein ig-h3 (TGFBI) Ubiquitin carboxyl-terminal hydrolase isozyme L1 (UCHL1) Ubiquitin-conjugating enzyme E2L 3 (UBE2L3) X-ray repair cross-complementing protein 5 (XRCC5) | ADP-ribosylation factor 4 (ARF4) Asparagine-tRNA ligase, cytoplasmic (NARS1) CD151 antigen (CD151) CD44 antigen (CD44) CD63 antigen (CD63) Cell division control protein 42 homolog (CDC42) Collagen alpha-1(V) chain (CO5A1) Coronin-1B (CORO1B) Coronin-1C (CORO1C) Cullin-3 (CUL3) Cyclin-dependent kinase 1 (CKD1) Ephrin type-A receptor 2 (EPHA2) Fascin (FSCN1) Glypican-1 (GPC1) Guanine nucleotide exchange factor VAV2 (VAV2) Inactive tyrosine protein kinase 7 (PTK7) Integrin beta-1 (ITB1) Junction plakoglobin (PLAK) Laminin subunit alpha-5 (LAMA5) Laminin subunit beta-1 (LAMB1) Laminin subunit beta-2 (LAMB2) Laminin subunit gamma-1 (LAMC1) Microtubule-associated protein RP/EB family member 1 (MAPRE1) Nck-associated protein 1 (NCKP1) Neural cell adhesion molecule L1 (L1CAM) Protein scribble homolog (SCRIB) Ras GTPase-activating-like protein IQGAP1 (IQGAP1) Ras-related protein Rab-1A (RAB1A) Rho-related GTP-binding protein RhoA (RHOA) Thrombospondin-1 (TSP1) | 14-3-3 protein zeta/delta (YWHAZ) 72 kDa type IV collagenase (MMP2) Actin, cytoplasmic 2 (ACTG1) Aminoacyl tRNA synthase complex-interacting multifunctional protein 1 (AIMP1) Annexin A2 (ANXA2) ATP synthase subunit beta, mitochondrial (ATP5F1B) Basement membrane-specific heparan sulfate proteoglycan core protein (HSPG2) Caveolin-1 (CAV1) Chloride intracellular channel protein 4 (CLIC4) Collagen alpha-1(XVIII) chain (COL18A1) Collagen alpha-2(IV) chain (COL4A2) E3 ubiquitin-protein ligase RNF213 (RNF213) Fibronectin (FN1) Guanine nucleotide exchange factor VAV2 (VAV2) Integrin alpha-V (ITGAV) Lactadherin (MFGE8) Myosin-9 (MYH9) Nucleolin (NCL) Peptidyl-prolyl cis/trans isomerase H (PPIH) Plasminogen activator inhibitor 1 (PAI1) Programmed cell death protein 6 (PDCD6) Transforming growth factor-beta-induced protein ig-h3 (TGFBI) Tryptophan-tRNA ligase, cytoplasmic (WARS1) | Complement C1r subcomponent (C1R) Complement C3 (C3) Deoxynucleoside triphosphate triphosphohydrolase (SAMHD1) HLA class I histocompatibility antigen, A alpha chain (HLA-A) HLA class I histocompatibility antigen, B-7 alpha chain (HLA-B) HLA class I histocompatibility antigen, Cw-7 alpha chain (HLA-C) Interferon-induced transmembrane protein 3 (IFITM3) Peroxidasin homolog (PXDN) Purine nucleoside phosphorylase (PNP) Thrombospondin-1 (THBS1) | 116 kDa U5 small nuclear ribonucleoprotein component (EFTUD2) 78 kDa glucose-regulated protein (HSPA5) CAD protein (CAD) Cellular tumor antigen p53 (TP53) Growth arrest-specific protein 6 (GAS6) Heat shock protein 90 alpha family class b member 1 (HSP90AB1) Nucleoside diphosphate kinase A (NME1) Ras-related protein Rap-1b (RAP1B) Transferrin receptor protein 1 (TFRC) Ubiquitin carboxyl-terminal hydrolase isozyme L1 (UCHL1) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Surman, M.; Kędracka-Krok, S.; Wilczak, M.; Rybczyński, P.; Jankowska, U.; Przybyło, M. Comparative Proteomic Profiling of Ectosomes Derived from Thyroid Carcinoma and Normal Thyroid Cells Uncovers Multiple Proteins with Functional Implications in Cancer. Cells 2022, 11, 1184. https://doi.org/10.3390/cells11071184
Surman M, Kędracka-Krok S, Wilczak M, Rybczyński P, Jankowska U, Przybyło M. Comparative Proteomic Profiling of Ectosomes Derived from Thyroid Carcinoma and Normal Thyroid Cells Uncovers Multiple Proteins with Functional Implications in Cancer. Cells. 2022; 11(7):1184. https://doi.org/10.3390/cells11071184
Chicago/Turabian StyleSurman, Magdalena, Sylwia Kędracka-Krok, Magdalena Wilczak, Piotr Rybczyński, Urszula Jankowska, and Małgorzata Przybyło. 2022. "Comparative Proteomic Profiling of Ectosomes Derived from Thyroid Carcinoma and Normal Thyroid Cells Uncovers Multiple Proteins with Functional Implications in Cancer" Cells 11, no. 7: 1184. https://doi.org/10.3390/cells11071184
APA StyleSurman, M., Kędracka-Krok, S., Wilczak, M., Rybczyński, P., Jankowska, U., & Przybyło, M. (2022). Comparative Proteomic Profiling of Ectosomes Derived from Thyroid Carcinoma and Normal Thyroid Cells Uncovers Multiple Proteins with Functional Implications in Cancer. Cells, 11(7), 1184. https://doi.org/10.3390/cells11071184