
Phenotypical Screening on Neuronal Plasticity in Hippocampal-Prefrontal Cortex Connectivity Reveals an Antipsychotic with a Novel Profile

 ,
,  ,
,  , , ,
, , , 
Abstract
:1. Introduction
2. Methods and Materials
2.1. Drugs
2.2. Animals
2.3. Hippocampal Slice Preparation: Oscillations
2.4. EEG, In Vivo
2.4.1. EEG Electrode Implantation
2.4.2. Data and Statistical Analysis
2.5. Inhibition of H-PFC LTP Caused by Platform Stress
2.6. Mouse Hippocampal Slice Preparation, Influence of Stress on LTP
2.7. Reversal Learning in the Rewarded T-Maze after Sub-Chronic Treatment with Phencyclidine (PCP)
3. Results
3.1. Effects on Theta Rhythm in Hippocampal Slices
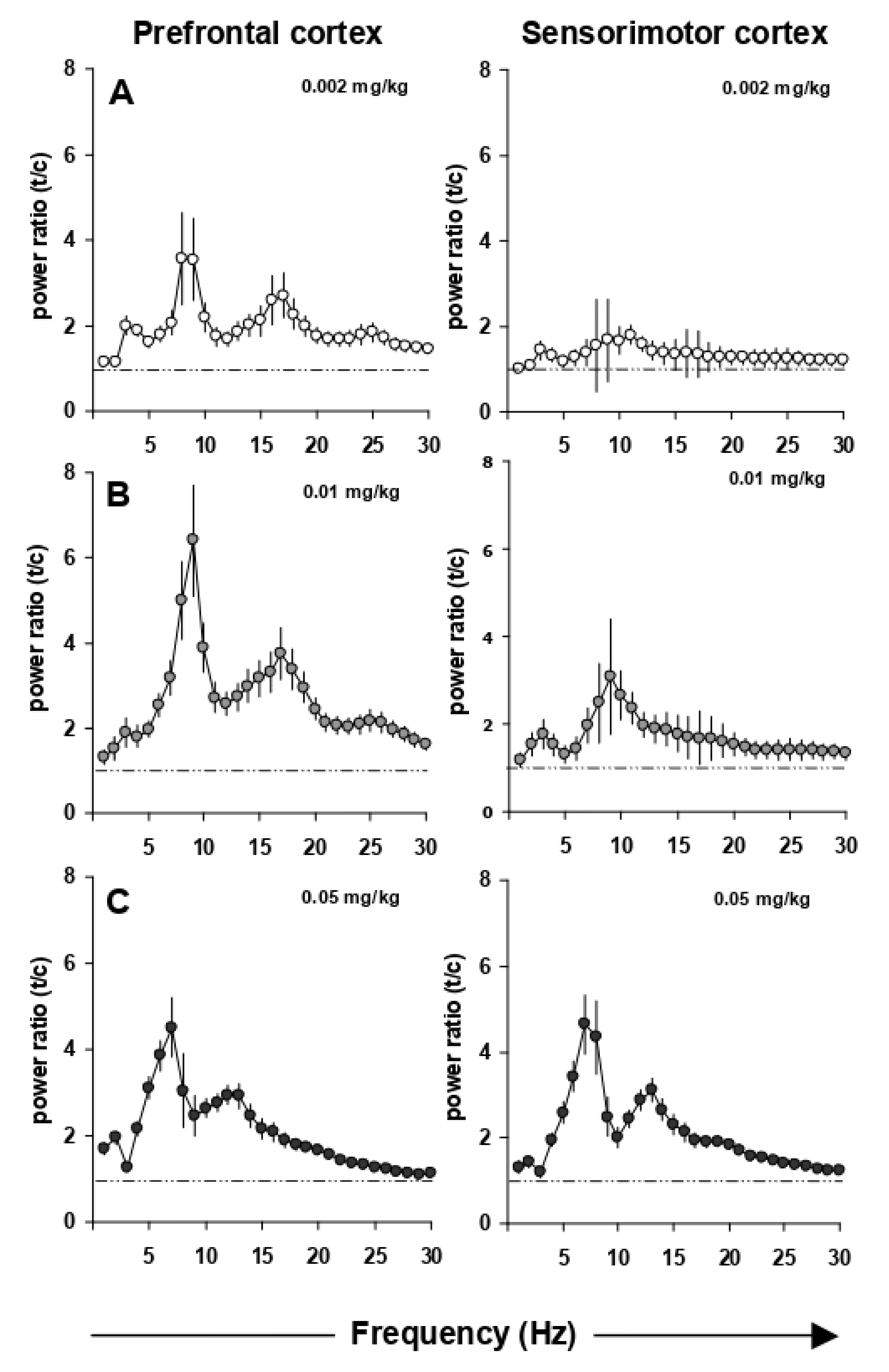
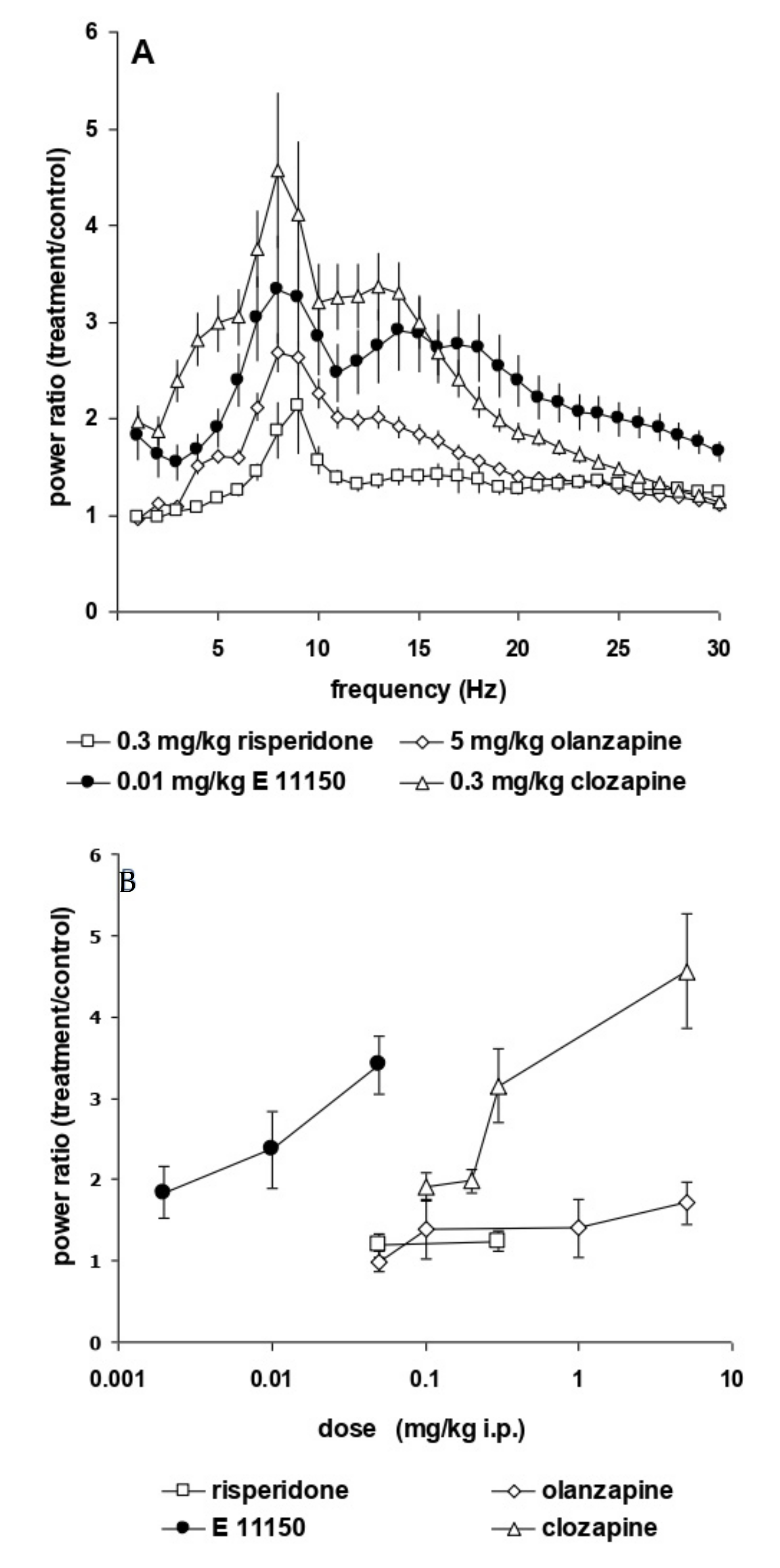
3.2. Effects of EGIS 11150 on EEG at 8–9 Hz
3.3. EGIS 11150 Rescues PCP—and Ketamine-Induced Effects on EEG
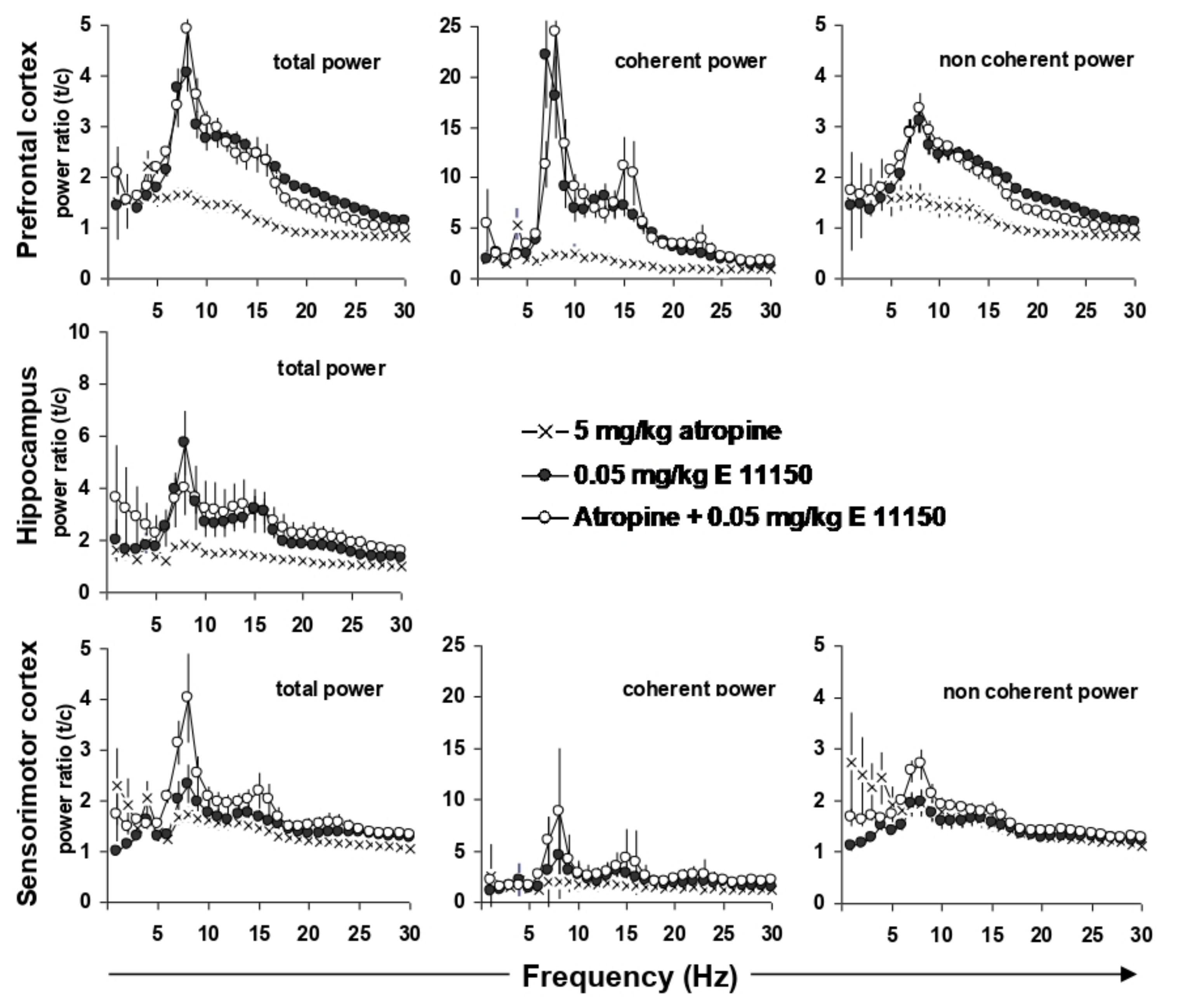
3.4. EEG Coherence
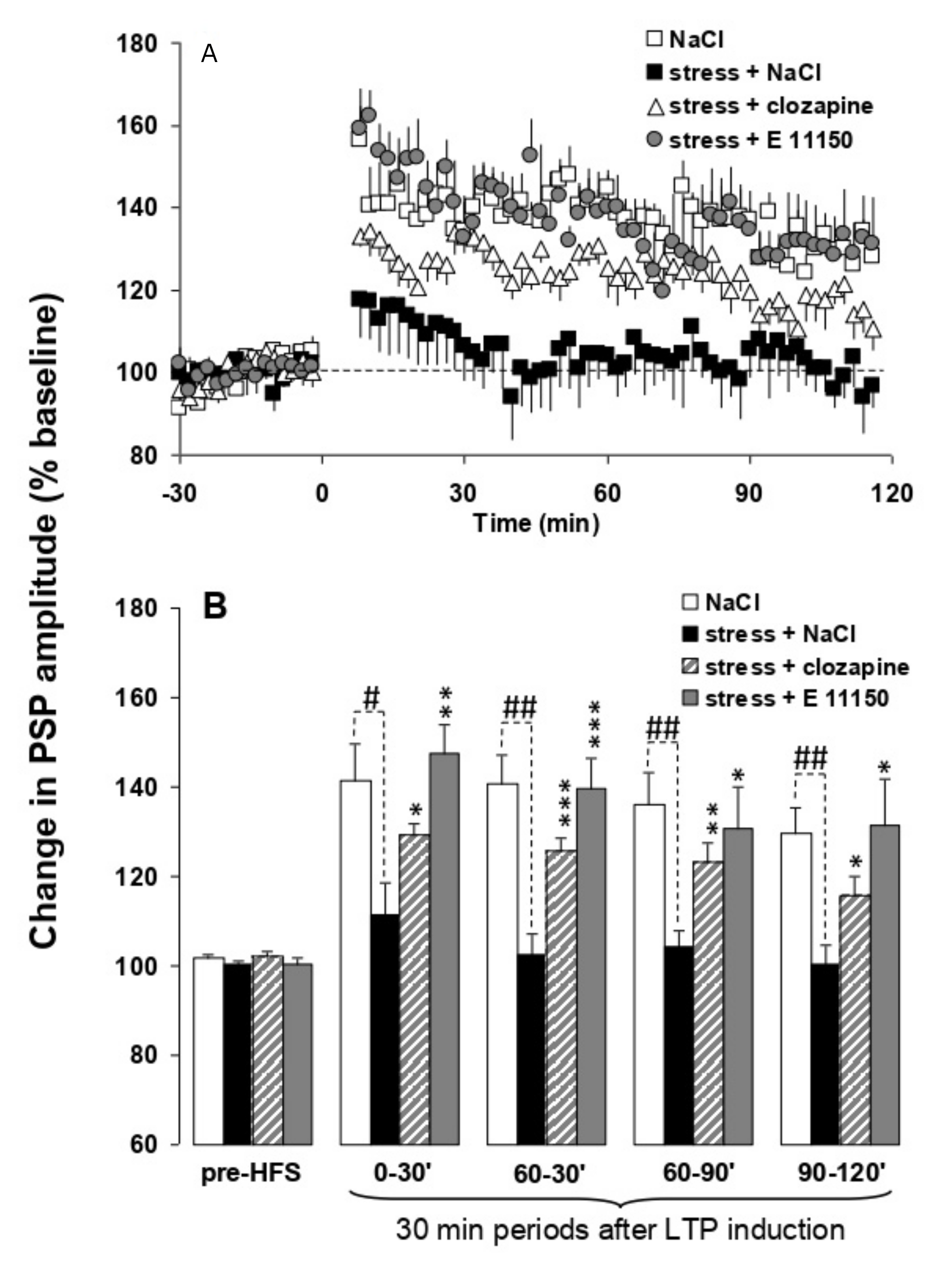
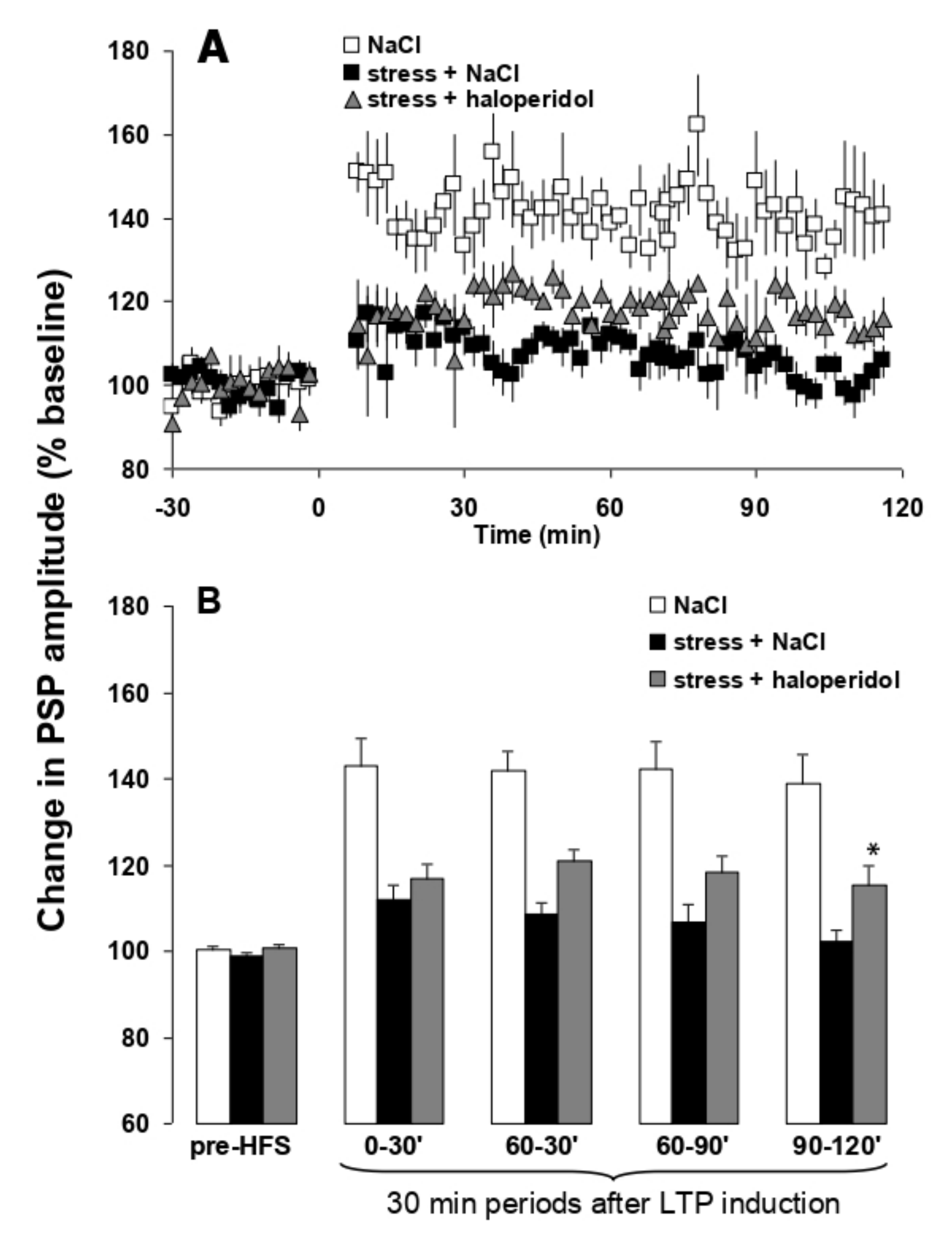
3.5. Effects of EGIS 11150 on the Inhibition of H-PFC LTP Caused by Platform Stress In Vivo
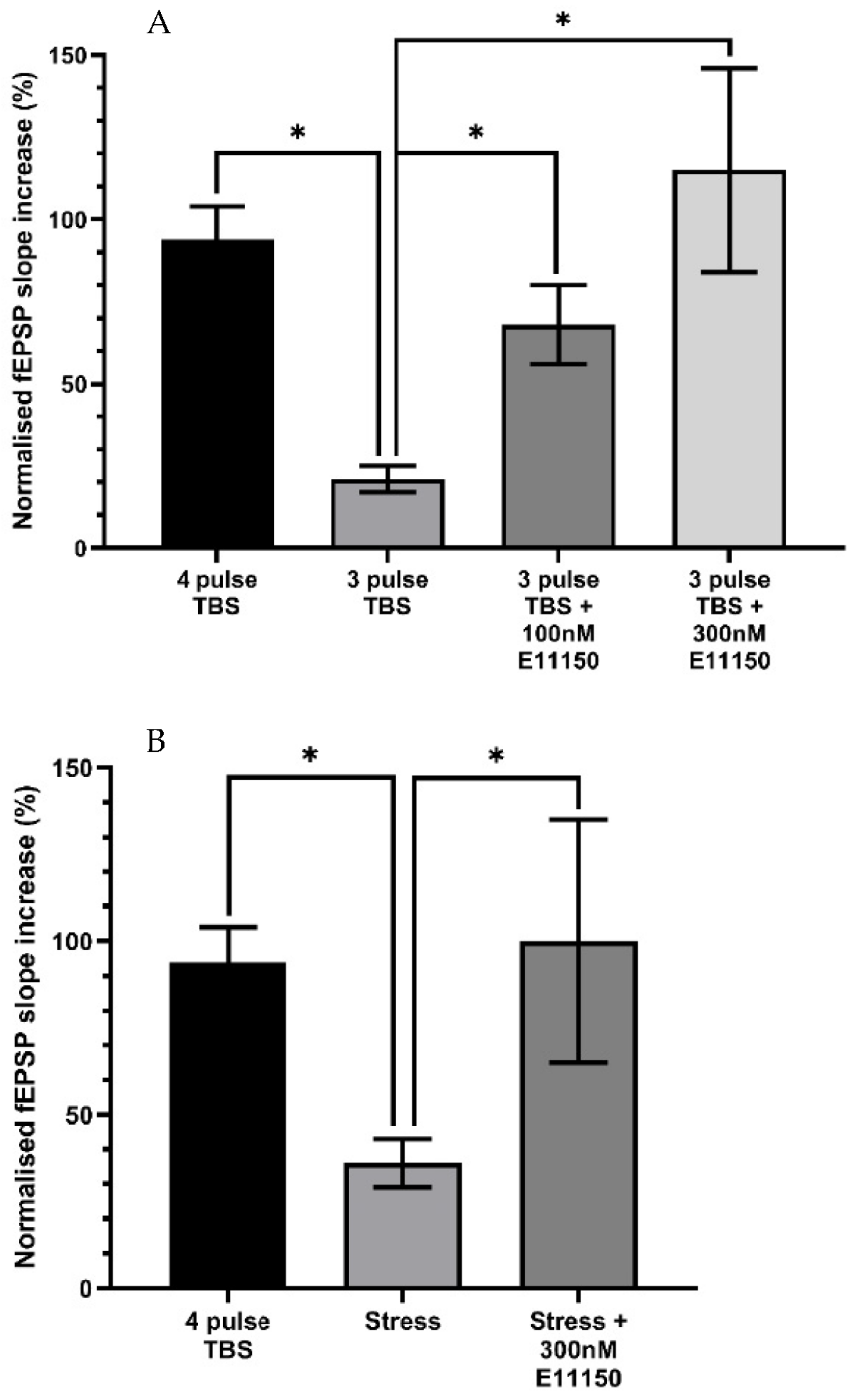
3.6. Effects of EGIS 11150 on Submaximal LTP Recorded from In Vitro Hippocampal Slices of Adult Mice
3.7. Acute Stress Suppresses Ex Vitro Hippocampal CA1 LTP In Vitro, a Deficit Reversed by EGIS 11150
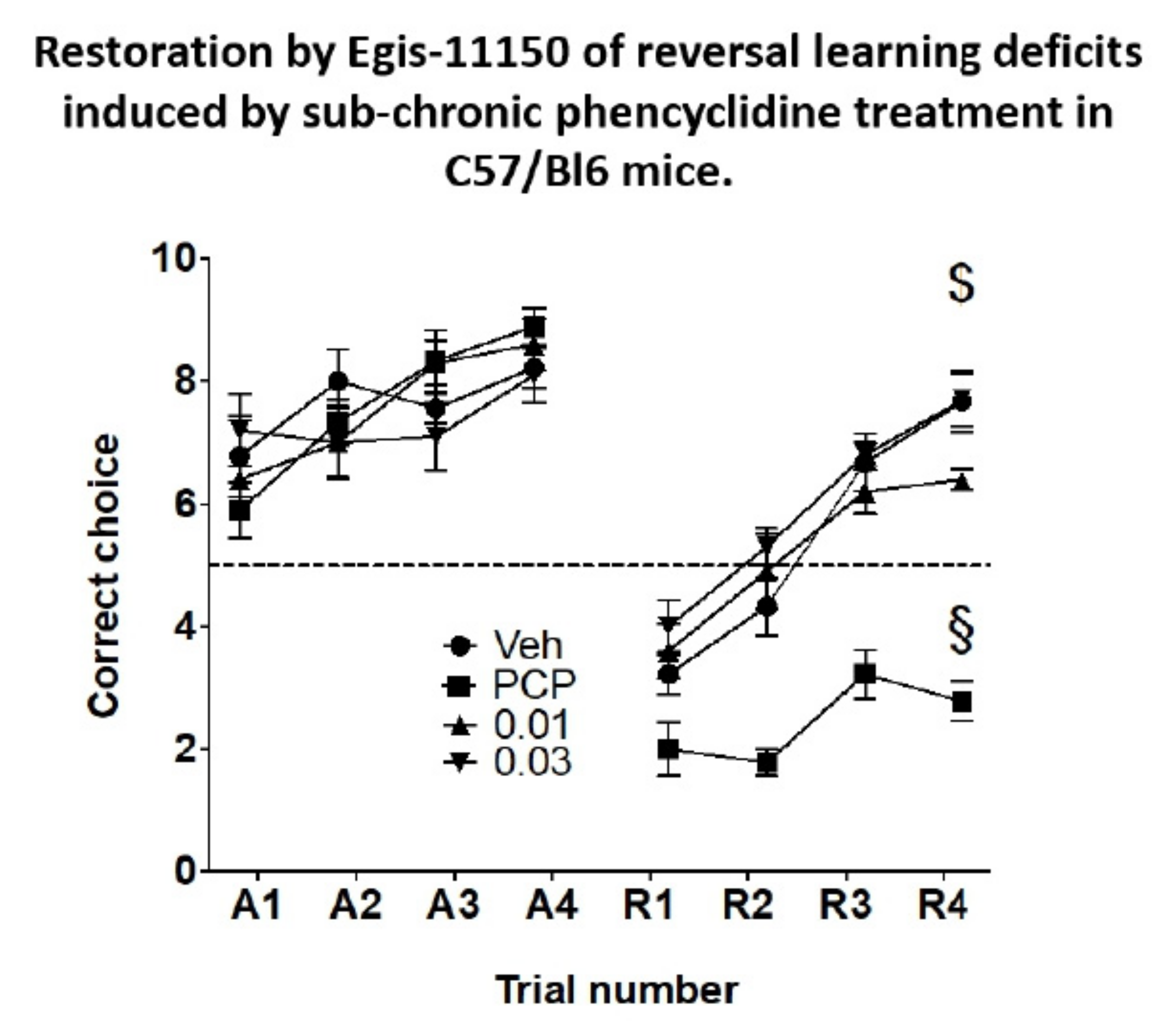
3.8. Reversal Learning in the Rewarded T-Maze
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Agid, Y.; Buzsaki, G.; Diamond, D.M.; Frackowiak, R.; Giedd, J.; Girault, J.-A.; Grace, A.; Lambert, J.J.; Manji, H.; Mayberg, H.; et al. How can drug discovery for psychiatric disorders be improved? Nat. Rev. Drug Discov. 2007, 6, 189–201. [Google Scholar] [CrossRef]
- Cohen, J.D.; Insel, T.R. Cognitive Neuroscience and Schizophrenia: Translational Research in Need of a Translator. Biol. Psychiatry 2008, 64, 2–3. [Google Scholar] [CrossRef] [PubMed]
- Carter, C.S.; Barch, D.M.; Buchanan, R.W.; Bullmore, E.; Krystal, J.H.; Cohen, J.; Geyer, M.; Green, M.; Nuechterlein, K.H.; Robbins, T.; et al. Identifying Cognitive Mechanisms Targeted for Treatment Development in Schizophrenia: An Overview of the First Meeting of the Cognitive Neuroscience Treatment Research to Improve Cognition in Schizophrenia Initiative. Biol. Psychiatry 2008, 64, 4–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kerns, J.G.; Nuechterlein, K.H.; Braver, T.; Barch, D.M. Executive Functioning Component Mechanisms and Schizophrenia. Biol. Psychiatry 2008, 64, 26–33. [Google Scholar] [CrossRef] [PubMed]
- Ochsner, K.N. The Social-Emotional Processing Stream: Five Core Constructs and Their Translational Potential for Schizophrenia and Beyond. Biol. Psychiatry 2008, 64, 48–61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ranganath, C.; Minzenberg, M.J.; Ragland, J.D. The Cognitive Neuroscience of Memory Function and Dysfunction in Schizophrenia. Biol. Psychiatry 2008, 64, 18–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Artigas, F.; Schenker, E.; Celada, P.; Spedding, M.; Lladó-Pelfort, L.; Jurado, N.; Núñez, M.; Santana, N.; Troyano-Rodriguez, E.; Riga, M.S.; et al. Defining the brain circuits involved in psychiatric disorders: IMI-NEWMEDS. Nat. Rev. Drug Discov. 2016, 16, 1–2. [Google Scholar] [CrossRef] [Green Version]
- Friston, K.; Frith, C.D. Schizophrenia: A disconnection syndrome? Clin. Neurosci. 1995, 3, 89–97. [Google Scholar]
- Stephan, K.E.; Friston, K.J.; Frith, C.D. Dysconnection in schizophrenia: From abnormal synaptic plasticity to failures of self-monitoring. Schizophr. Bull. 2009, 35, 509–527. [Google Scholar] [CrossRef] [Green Version]
- Pocklington, A.J.; Rees, E.; Walters, J.; Han, J.; Kavanagh, D.H.; Chambert, K.D.; Holmans, P.; Moran, J.; McCarroll, S.A.; Kirov, G.; et al. Novel Findings from CNVs Implicate Inhibitory and Excitatory Signaling Complexes in Schizophrenia. Neuron 2015, 86, 1203–1214. [Google Scholar] [CrossRef] [Green Version]
- Young, K.A.; Manaye, K.F.; Liang, C.-L.; Hicks, P.B.; German, D.C. Reduced number of mediodorsal and anterior thalamic neurons in schizophrenia. Biol. Psychiatry 2000, 47, 944–953. [Google Scholar] [CrossRef]
- Godsil, B.; Bontempi, B.; Mailliet, F.; Delagrange, P.; Spedding, M.; Jay, T.M. Acute tianeptine treatment selectively modulates neuronal activation in the central nucleus of the amygdala and attenuates fear extinction. Mol. Psychiatry 2015, 20, 1420–1427. [Google Scholar] [CrossRef]
- Spedding, M.; Chattarji, S.; Spedding, C.; Jay, T.M. Brain circuits at risk in psychiatric diseases and pharmacological pathways. Therapies 2021, 76, 75–86. [Google Scholar] [CrossRef] [PubMed]
- Park, A.J.; Harris, A.Z.; Martyniuk, K.M.; Chang, C.-Y.; Abbas, A.I.; Lowes, D.C.; Kellendonk, C.; Gogos, J.A.; Gordon, J.A. Reset of hippocampal–prefrontal circuitry facilitates learning. Nature 2021, 591, 615–619. [Google Scholar] [CrossRef] [PubMed]
- Takashim, A.; Petersson, K.M.; Rutters, F.; Tendolkar, I.; Jensen, O.; Zwarts, M.J.; McNaughton, B.L.; Fernández, G. Declarative memory consolidation in humans: A prospective functional magnetic resonance imaging study. Proc. Natl. Acad. Sci. USA 2006, 103, 756–761. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bähner, F.; Meyer-Lindenberg, A. Hippocampal–prefrontal connectivity as a translational phenotype for schizophrenia. Eur. Neuropsychopharmacol. 2017, 27, 93–106. [Google Scholar] [CrossRef]
- Tamminga, C.A. The neurobiology of cognition in schizophrenia. J. Clin. Psychiatry 2006, 9, 9. [Google Scholar] [CrossRef]
- Kupferschmidt, D.A.; Gordon, J.A. The dynamics of disordered dialogue: Prefrontal, hippocampal and thalamic miscommunication underlying working memory deficits in schizophrenia. Brain Neurosci. Adv. 2018, 2. [Google Scholar] [CrossRef] [PubMed]
- Rocher, C.; Spedding, M.; Munoz, C.; Jay, T.M. Acute Stress-induced Changes in Hippocampal/Prefrontal Circuits in Rats: Effects of Antidepressants. Cereb. Cortex 2004, 14, 224–229. [Google Scholar] [CrossRef] [PubMed]
- Spedding, M.; Neau, I.; Harsing, L. Brain plasticity and pathology in psychiatric disease: Sites of action for potential therapy. Curr. Opin. Pharmacol. 2002, 3, 33–40. [Google Scholar] [CrossRef]
- Sigurdsson, T.; Stark, K.L.; Karayiorgou, M.; Gogos, J.A.; Gordon, J.A. Impaired hippocampal–prefrontal synchrony in a genetic mouse model of schizophrenia. Nature 2010, 464, 763–767. [Google Scholar] [CrossRef]
- Karayiorgou, M.; Simon, T.J.; Gogos, J.A. 22q11.2 microdeletions: Linking DNA structural variation to brain dysfunction and schizophrenia. Nat. Rev. Neurosci. 2010, 11, 402–416. [Google Scholar] [CrossRef]
- Spedding, M. Resolution of controversies in drug/receptor interactions by protein structure. Limitations and pharmacological solutions. Neuropharmacology 2011, 60, 3–6. [Google Scholar] [CrossRef]
- Urban, J.; Clarke, W.; Von Zastrow, M.; Nichols, D.E.; Kobilka, B.; Weinstein, H.; Javitch, J.; Roth, B.L.; Christopoulos, A.; Sexton, P.; et al. Functional Selectivity and Classical Concepts of Quantitative Pharmacology. J. Pharmacol. Exp. Ther. 2006, 320, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Gacsályi, I.; Nagy, K.; Pallagi, K.; Lévay, G.; Hársing, L.G.; Móricz, K.; Kertész, S.; Varga, P.; Haller, J.; Gigler, G.; et al. Egis-11150: A candidate antipsychotic compound with procognitive efficacy in rodents. Neuropharmacology 2012, 64, 254–263. [Google Scholar] [CrossRef] [PubMed]
- Spedding, M.; Jay, T.; Silva, J.C.; Perret, L. A pathophysiological paradigm for the therapy of psychiatric disease. Nat. Rev. Drug Discov. 2005, 4, 467–476. [Google Scholar] [CrossRef] [PubMed]
- Buzsáki, G. Theta rhythm of navigation: Link between path integration and landmark navigation, episodic and semantic memory. Hippocampus 2005, 15, 827–840. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamaguchi, Y.; Sato, N.; Wagatsuma, H.; Wu, Z.; Molter, C.; Aota, Y. A unified view of theta-phase coding in the entorhinal–hippocampal system. Curr. Opin. Neurobiol. 2007, 17, 197–204. [Google Scholar] [CrossRef]
- Seager, M.A.; Johnson, L.D.; Chabot, E.S.; Asaka, Y.; Berry, S.D. Oscillatory brain states and learning: Impact of hippocampal theta-contingent training. Proc. Natl. Acad. Sci. USA 2002, 99, 1616–1620. [Google Scholar] [CrossRef] [Green Version]
- Sebban, C.; Zhang, X.Q.; Tesolin-Decros, B.; Millan, M.J.; Spedding, M. Changes in EEG spectral power in the prefrontal cortex of conscious rats elicited by drugs interacting with dopaminergic and noradrenergic transmission. J. Cereb. Blood Flow Metab. 1999, 128, 1045–1054. [Google Scholar] [CrossRef] [PubMed]
- Sebban, C.; Tesolin-Decros, B.; Millan, M.J.; Spedding, M. Contrasting EEG profiles elicited by antipsychotic agents in the prefrontal cortex of the conscious rat: Antagonism of the effects of clozapine by modafinil. J. Cereb. Blood Flow Metab. 1999, 128, 1055–1063. [Google Scholar] [CrossRef] [PubMed]
- Sebban, C.; Tesolin-Decros, B.; Ciprian-Ollivier, J.; Perret, L.; Spedding, M. Effects of phencyclidine (PCP) and MK 801 on the EEGq in the prefrontal cortex of conscious rats; antagonism by clozapine, and antagonists of AMPA-, α1- and 5-HT2A -receptors. J. Cereb. Blood Flow Metab. 2002, 135, 65–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gross, A.; Joutsiniemi, S.-L.; Rimon, R.; Appelberg, B. Clozapine-Induced QEEG Changes Correlate with Clinical Response in Schizophrenic Patients: A Prospective, Longitudinal Study. Pharmacopsychiatry 2004, 37, 119–122. [Google Scholar] [CrossRef]
- Tislerova, B.; Brunovsky, M.; Horáček, J.; Novak, T.; Kopecek, M.; Mohr, P.; Krajca, V. LORETA Functional Imaging in Antipsychotic-Naive and Olanzapine-, Clozapine- and Risperidone-Treated Patients with Schizophrenia. Neuropsychobiology 2008, 58, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Lisman, J.; Buzsaki, G. A Neural Coding Scheme Formed by the Combined Function of Gamma and Theta Oscillations. Schizophr. Bull. 2008, 34, 974–980. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujisawa, S.; Amarasingham, A.; Harrison, M.T.; Buzsáki, G. Behavior-dependent short-term assembly dynamics in the medial prefrontal cortex. Nat. Neurosci. 2008, 11, 823–833. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sirota, A.; Montgomery, S.; Fujisawa, S.; Isomura, Y.; Zugaro, M.; Buzsáki, G. Entrainment of Neocortical Neurons and Gamma Oscillations by the Hippocampal Theta Rhythm. Neuron 2008, 60, 683–697. [Google Scholar] [CrossRef] [Green Version]
- Krystal, J.H.; D’Souza, D.C.; Mathalon, D.; Perry, E.; Belger, A.; Hoffman, R. NMDA receptor antagonist effects, cortical glutamatergic function, and schizophrenia: Toward a paradigm shift in medication development. Psychopharmacology 2003, 169, 215–233. [Google Scholar] [CrossRef]
- Lahti, A.C.; Weiler, M.A.; Tamara, M.; Parwani, A.; Tamminga, C.A. Effects of Ketamine in Normal and Schizophrenic Volunteers. Neuropsychopharmacology 2001, 25, 455–467. [Google Scholar] [CrossRef] [Green Version]
- Jentsch, J.D.; Roth, R.H. The neuropsychopharmacology of phencyclidine: From NMDA receptor hypofunction to the dopamine hypothesis of schizophrenia. Neuropsychopharmacology 1999, 20, 201–225. [Google Scholar] [CrossRef] [Green Version]
- Büttner-Ennever, J. The Rat Brain in Stereotaxic Coordinates, 3rd edn. J. Anat. 1997, 191, 315–317. [Google Scholar] [CrossRef]
- Zhang, H.; Etherington, L.-A.; Hafner, A.-S.; Belelli, D.; Coussen, F.; Delagrange, P.; Chaouloff, F.; Spedding, M.; Lambert, J.J.; Choquet, D.; et al. Regulation of AMPA receptor surface trafficking and synaptic plasticity by a cognitive enhancer and antidepressant molecule. Mol. Psychiatry 2012, 18, 471–484. [Google Scholar] [CrossRef] [PubMed]
- Deacon, R.M.J.; Rawlins, J.N.P. T-maze alternation in the rodent. Nat. Protoc. 2006, 1, 7–12. [Google Scholar] [CrossRef] [PubMed]
- Gacsályi, I.; Móricz, K.; Gigler, G.; Megyeri, K.; Machado, P.; Antoni, F.A. Persistent therapeutic effect of a novel α5-GABAA receptor antagonist in rodent preclinical models of vascular cognitive impairment. Eur. J. Pharmacol. 2018, 834, 118–125. [Google Scholar] [CrossRef]
- Singer, P.; Boison, D.; Möhler, H.; Feldon, J.; Yee, B.K. Deletion of glycine transporter 1 (GlyT1) in forebrain neurons facilitates reversal learning: Enhanced cognitive adaptability? Behav. Neurosci. 2009, 123, 1012–1027. [Google Scholar] [CrossRef] [Green Version]
- Gacsalyi, I.; Gigler, G.; Kompagne, H.; Haller, J.; Azar, M.R.; Moricz, K.; Levay, G. Pharmacology of a new atypical antipsychotic agent, EGIS-11150/S36549. Schizophr. Bull. 2007, 33, 472. [Google Scholar]
- Gillies, M.J.; Traub, R.D.; LeBeau, F.E.N.; Davies, C.H.; Gloveli, T.; Buhl, E.H.; Whittington, M.A. A Model of Atropine—Resistant Theta Oscillations in Rat Hippocampal Area CA1. J. Physiol. 2002, 543, 779–793. [Google Scholar] [CrossRef]
- Chrobak, J.J.; Buzsáki, G. Operational dynamics in the hippocampal-entorhinal axis. Neurosci. Biobehav. Rev. 1998, 22, 303–310. [Google Scholar] [CrossRef]
- Jensen, O.; Tesche, C.D. Frontal theta activity in humans increases with memory load in a working memory task. Eur. J. Neurosci. 2002, 15, 1395–1399. [Google Scholar] [CrossRef]
- Sederberg, P.B.; Kahana, M.J.; Howard, M.W.; Donner, E.J.; Madsen, J.R. Theta and Gamma Oscillations during Encoding Predict Subsequent Recall. J. Neurosci. 2003, 23, 10809–10814. [Google Scholar] [CrossRef] [Green Version]
- Jay, T.M.; Witter, M.P. Distribution of hippocampal CA1 and subicular efferents in the prefrontal cortex of the rat studied by means of anterograde transport of Phaseolus vulgaris-leucoagglutinin. J. Comp. Neurol. 1991, 313, 574–586. [Google Scholar] [CrossRef] [PubMed]
- Tierney, P.L.; Dégenètais, E.; Thierry, A.; Glowinski, J.; Gioanni, Y. Influence of the Hippocampus on Interneurons of the Rat Prefrontal Cortex. The European Journal of Neuroscience. Available online: https://pubmed.ncbi.nlm.nih.gov/15233760/ (accessed on 20 April 2021).
- Colgin, L.L. Oscillations and hippocampal–prefrontal synchrony. Curr. Opin. Neurobiol. 2011, 21, 467–474. [Google Scholar] [CrossRef] [Green Version]
- Benchenane, K.; Peyrache, A.; Khamassi, M.; Tierney, P.L.; Gioanni, Y.; Battaglia, F.P.; Wiener, S.I. Coherent Theta Oscillations and Reorganization of Spike Timing in the Hippocampal- Prefrontal Network upon Learning. Neuron 2010, 66, 921–936. [Google Scholar] [CrossRef] [PubMed]
- Jones, M.; Wilson, M.A. Theta Rhythms Coordinate Hippocampal–Prefrontal Interactions in a Spatial Memory Task. PLoS Biol. 2005, 3, e402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hyman, J.M.; Zilli, E.A.; Paley, A.M.; Hasselmo, M.E. Medial prefrontal cortex cells show dynamic modulation with the hippocampal theta rhythm dependent on behavior. Hippocampus 2005, 15, 739–749. [Google Scholar] [CrossRef] [PubMed]
- Siapas, A.G.; Lubenov, E.V.; Wilson, M.A. Prefrontal Phase Locking to Hippocampal Theta Oscillations. Neuron 2005, 46, 141–151. [Google Scholar] [CrossRef] [Green Version]
- Fein, G.; Raz, J.; Brown, F.F.; Merrin, E.L. Common reference coherence data are confounded by power and phase effects. Electroencephalogr. Clin. Neurophysiol. 1988, 69, 581–584. [Google Scholar] [CrossRef]
- Nunez, P.L.; Srinivasan, R.; Westdorp, A.F.; Wijesinghe, R.S.; Tucker, D.M.; Silberstein, R.B.; Cadusch, P.J. EEG coherency: I: Statistics, reference electrode, volume conduction, Laplacians, cortical imaging, and interpretation at multiple scales. Electroencephalogr. Clin. Neurophysiol. 1997, 103, 499–515. [Google Scholar] [CrossRef]
- Roopun, A.K.; Kramer, M.A.; Carracedo, L.M.; Kaiser, M.; Davies, C.H.; Traub, R.D.; Kopell, N.J.; Whittington, M.A. Temporal interactions between cortical rhythms. Front. Behav. Neurosci. 2008, 2, 145–154. [Google Scholar] [CrossRef] [Green Version]
- Aghajanian, G.K.; Marek, G.J. Serotonin model of schizophrenia: Emerging role of glutamate mechanisms. Brain Res. Brain Res. Rev. 2000, 31, 302–312. [Google Scholar] [CrossRef]
- Aghajanian, G.K.; Marek, G.J. Serotonin, via 5-HT2A receptors, increases EPSCs in layer V pyramidal cells of prefrontal cortex by an asynchronous mode of glutamate release. Brain Res. 1999, 825, 161–171. [Google Scholar] [CrossRef]
- Hársing, L.G.; Gacsalyi, I.; Szabo, G.; Schmidt, E.; Sziray, N.; Sebban, C.; Tesolin-Decros, B.; Matyus, P.; Egyed, A.; Spedding, M.; et al. The glycine transporter-1 inhibitors NFPS and Org 24461: A pharmacological study. Pharmacol. Biochem. Behav. 2003, 74, 811–825. [Google Scholar] [CrossRef]
- Nagy, K.; Marko, B.; Zsilla, G.; Matyus, P.; Pallagi, K.; Szabo, G.; Juranyi, Z.; Barkoczy, J.; Levay, G.; Harsing, L.G. Alterations in Brain Extracellular Dopamine and Glycine Levels Following Combined Administration of the Glycine Transporter Type-1 Inhibitor Org-24461 and Risperidone. Neurochem. Res. 2010, 35, 2096–2106. [Google Scholar] [CrossRef] [PubMed]
- Lim, E.P.; Tan, C.H.; Jay, T.M.; Dawe, G.S. Locus coeruleus stimulation and noradrenergic modulation of hippocampo-prefrontal cortex long-term potentiation. Int. J. Neuropsychopharmacol. 2010, 13, 1219–1231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kobayashi, M.; Sasabe, T.; Shiohama, Y.; Koshikawa, N. Activation of α1-adrenoceptors increases firing frequency through protein kinase C in pyramidal neurons of rat visual cortex. Neurosci. Lett. 2008, 430, 175–180. [Google Scholar] [CrossRef]
- McAlonan, K.; Brown, V.J. Orbital prefrontal cortex mediates reversal learning and not attentional set shifting in the rat. Behav. Brain Res. 2003, 146, 97–103. [Google Scholar] [CrossRef]
- Bartolo, R.; Averbeck, B.B. Prefrontal Cortex Predicts State Switches during Reversal Learning. Neuron 2020, 106, 1044–1054.e4. [Google Scholar] [CrossRef]
- Tripathi, A.; Spedding, M.; Schenker, E.; Didriksen, M.; Cressant, A.; Jay, T.M. Cognition- and circuit-based dysfunction in a mouse model of 22q11.2 microdeletion syndrome: Effects of stress. Transl. Psychiatry 2020, 10, 41. [Google Scholar] [CrossRef] [Green Version]
- Qi, H.; Mailliet, F.; Spedding, M.; Rocher, C.; Zhang, X.; Delagrange, P.; McEwen, B.; Jay, T.M.; Svenningsson, P. Antidepressants reverse the attenuation of the neurotrophic MEK/MAPK cascade in frontal cortex by elevated platform stress; reversal of effects on LTP is associated with GluA1 phosphorylation. Neuropharmacology 2009, 56, 37–46. [Google Scholar] [CrossRef]
- Huang, L.; Abuhamdah, S.; Howes, M.-J.R.; Dixon, C.L.; Elliot, M.S.J.; Ballard, C.; Holmes, C.; Burns, A.; Perry, E.K.; Francis, P.T.; et al. Pharmacological profile of essential oils derived from Lavandula angustifolia and Melissa officinalis with anti-agitation properties: Focus on ligand-gated channels. J. Pharm. Pharmacol. 2008, 60, 1515–1522. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Preparation, Location, Lead Author | Animals, Weight Age, n | Ethical Approval | Drug, Dose, Concentration |
|---|---|---|---|
| 2.3 Hippocampal slice preparation: oscillations, in vitro Newcastle, UK; Cunningham M. O. | Adult male Wistar rats (~150 g) and aged male (8 months) n = 6 (EGIS; CA1) n = 6 (EGIS; CA3) n = 5 (ZD7288) n = 5 (gabazine) n = 3 (atropine) | UK Animals Act 1986 & European Union Directive2010/63/EU (No. 25308) | Isoflurane (6–8 mls), Xylazine (~10 mg/kg i.m.), Ketamine (~100 mg/kg i.m.) EGIS 11150 (200 nM), ZD7288 (10 µM), gabazine (2 µM), atropine (100 nM) |
| 2.4 EEG Electrode implantation, in vivo Paris, France; Sebban C. | Conscious rats Adult male Wistar rats (~150 g) and aged male (8 months) n = 6/group, 1 animal/cage One or Two-way ANOVA (see text) | The institutional guidelines (Centre de recherche scientifique) and the prerogatives from the French Agriculture and Forestry Ministry (decree 874848, licence A91429) | Chloral hydrate (350 mg/kg i.p.) EGIS 11150 (0.02–0.01–0.05 mg/kg) Clozapine 0.3 mg/kg (s.c.) Risperidone 0.3 mg/kg (i.p.) Olanzapine 5 mg/kg (i.p.) PCP 1 mg/kg (i.p.) Drugs dissolved in 0.9% NaCl |
| 2.5 Inhibition of H-PFC LTP caused by platform stress, in vivo Paris, France; Jay T. M. | Adult male Srague Dawley rats (300–400 g) n = 6/non stressed NaCl, stressed EGIS11150 and stressed Hal n = 8/stressed NaCl and stressed clozapine | All animal experiments were performed in accordance with our institution guidelines (Centre National de la Recherche Scientifique) and the prerogatives from the French Agriculture and Forestry Ministry (decree 874848, license A91429). | Pentobarbital (60 mg/kg i.p.) EGIS11150 (0.01 mg/kg, i.p.), clozapine (0.3 mg/kg, I.p.) or Haloperidol (1 mg/kg, i.p.) |
| 2.6 Mouse hippocampal slice preparation, Influence of stress on LTP, in vitro Dundee, Scotland; Lambert J. J. | 2–4 month-old male C57BL/6 mice (22–25 g) n = 4–12 | Schedule 1 of the UK Government Animals Act 1986 & Home Office project license PPL 60/3575 | EGIS 11150 (100–300 nM) |
| 2.7 Reversal learning in the rewarded T-maze, in vivo EGIS, Budapest, Hungary; Antoni F. A. | Male C57Bl/6j Mice (20–25 g) 5 animals/cage 40 animals 10/treatment group, all included in the results | Animal Care and Use Ethical Committee of Egis Pharmaceuticals PLC and complied with the Hungarian Law of Animal Care and Use (1998. XVIII) and Directive 2010/63/EU on the protection of animals used for scientific purposes. | PCP (3 mg/kg i.p.) or saline (i.p.) derived from previous studies with Egis 11150 (0.01 or 0.03 mg/kg p.o.) (Gacsályi, I. et al., 2013) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Spedding, M.; Sebban, C.; Jay, T.M.; Rocher, C.; Tesolin-Decros, B.; Chazot, P.; Schenker, E.; Szénási, G.; Lévay, G.I.; Megyeri, K.; et al. Phenotypical Screening on Neuronal Plasticity in Hippocampal-Prefrontal Cortex Connectivity Reveals an Antipsychotic with a Novel Profile. Cells 2022, 11, 1181. https://doi.org/10.3390/cells11071181
Spedding M, Sebban C, Jay TM, Rocher C, Tesolin-Decros B, Chazot P, Schenker E, Szénási G, Lévay GI, Megyeri K, et al. Phenotypical Screening on Neuronal Plasticity in Hippocampal-Prefrontal Cortex Connectivity Reveals an Antipsychotic with a Novel Profile. Cells. 2022; 11(7):1181. https://doi.org/10.3390/cells11071181
Chicago/Turabian StyleSpedding, Michael, Claude Sebban, Thérèse M. Jay, Cyril Rocher, Brigitte Tesolin-Decros, Paul Chazot, Esther Schenker, Gabor Szénási, György I. Lévay, Katalin Megyeri, and et al. 2022. "Phenotypical Screening on Neuronal Plasticity in Hippocampal-Prefrontal Cortex Connectivity Reveals an Antipsychotic with a Novel Profile" Cells 11, no. 7: 1181. https://doi.org/10.3390/cells11071181
APA StyleSpedding, M., Sebban, C., Jay, T. M., Rocher, C., Tesolin-Decros, B., Chazot, P., Schenker, E., Szénási, G., Lévay, G. I., Megyeri, K., Barkóczy, J., Hársing, L. G., Jr., Thomson, I., Cunningham, M. O., Whittington, M. A., Etherington, L.-A., Lambert, J. J., Antoni, F. A., & Gacsályi, I. (2022). Phenotypical Screening on Neuronal Plasticity in Hippocampal-Prefrontal Cortex Connectivity Reveals an Antipsychotic with a Novel Profile. Cells, 11(7), 1181. https://doi.org/10.3390/cells11071181