
Effects of Sigma-1 Receptor Ligands on Peripheral Nerve Regeneration


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
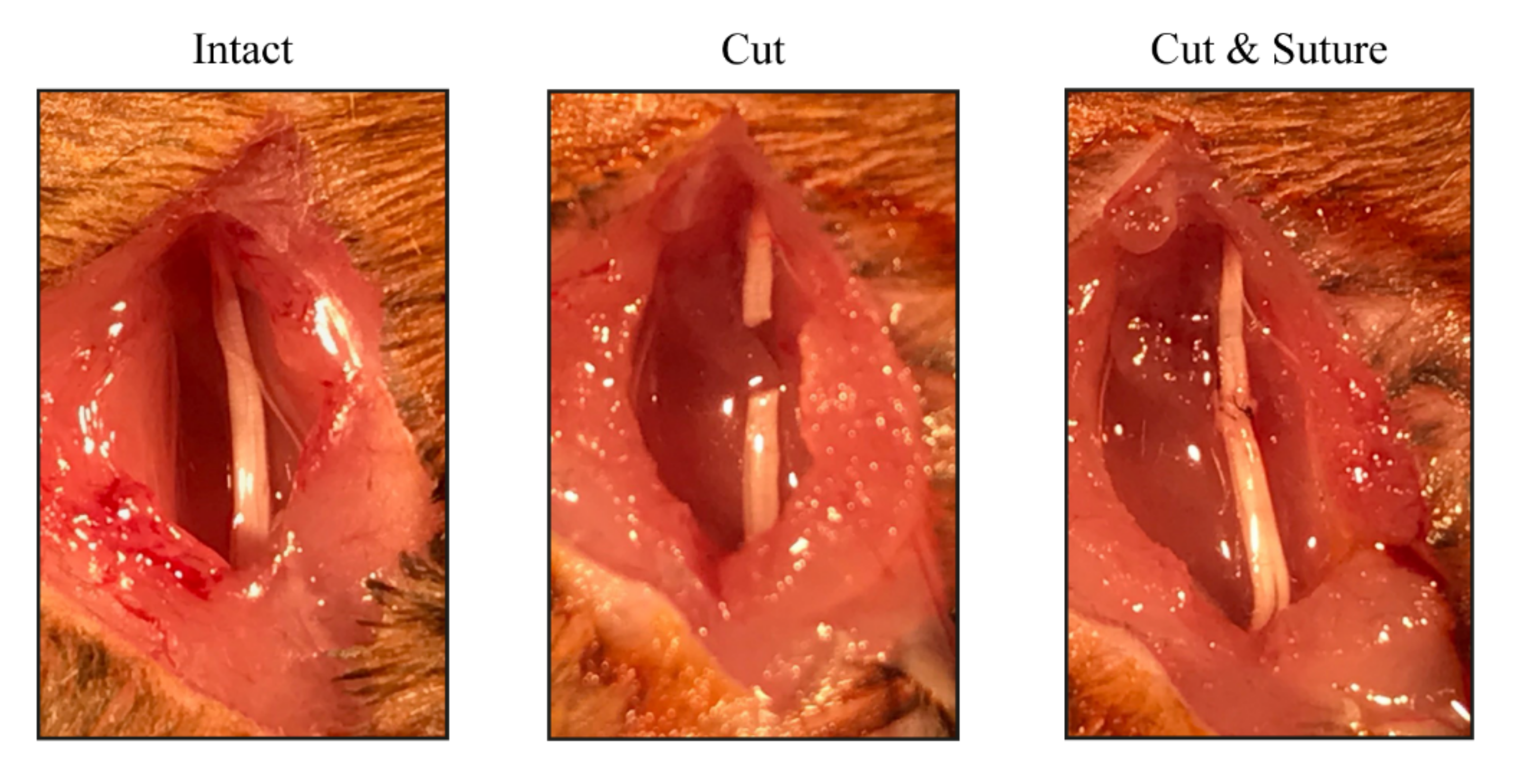
2.1. Animals and Surgical Procedures
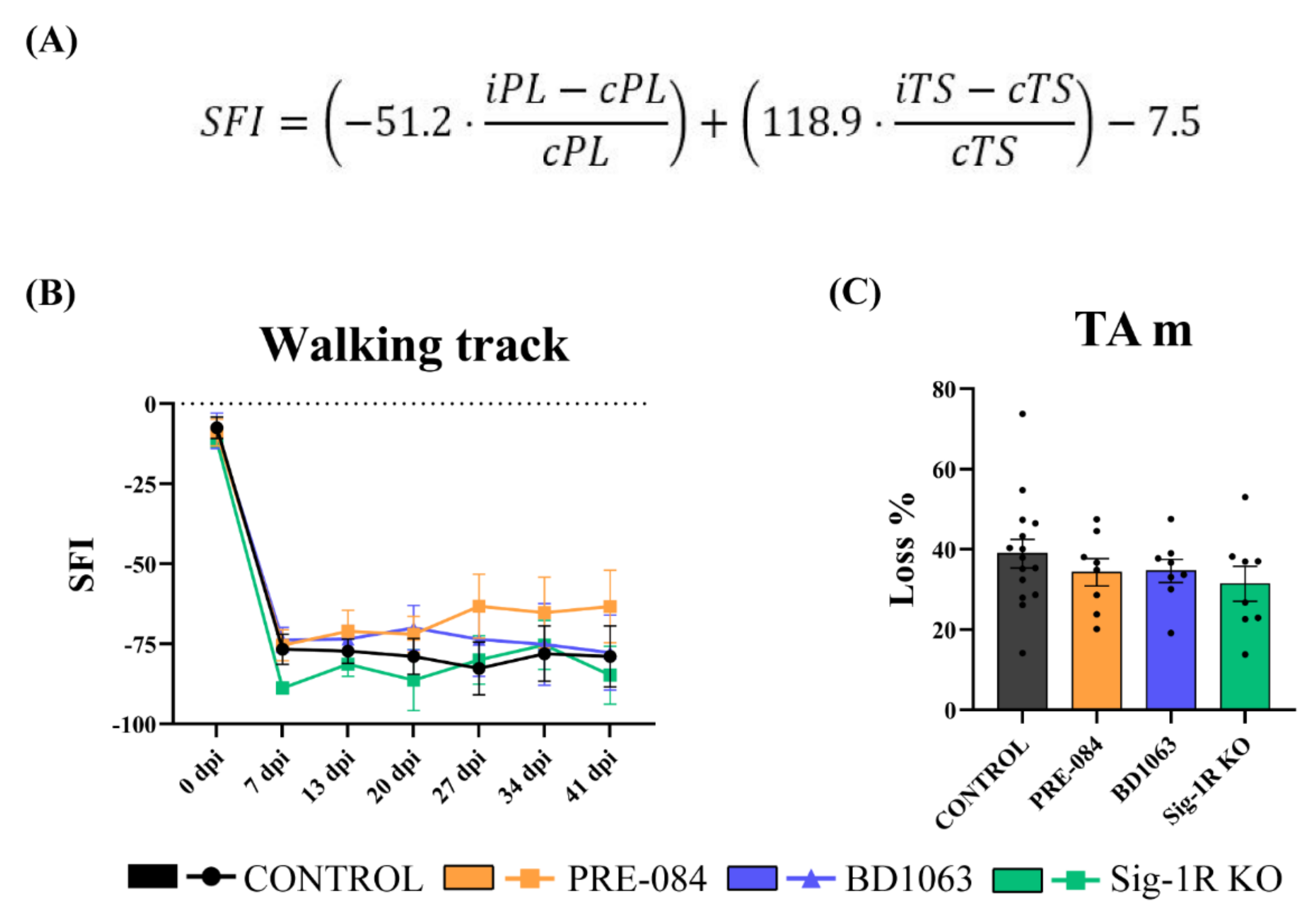
2.2. Walking Track
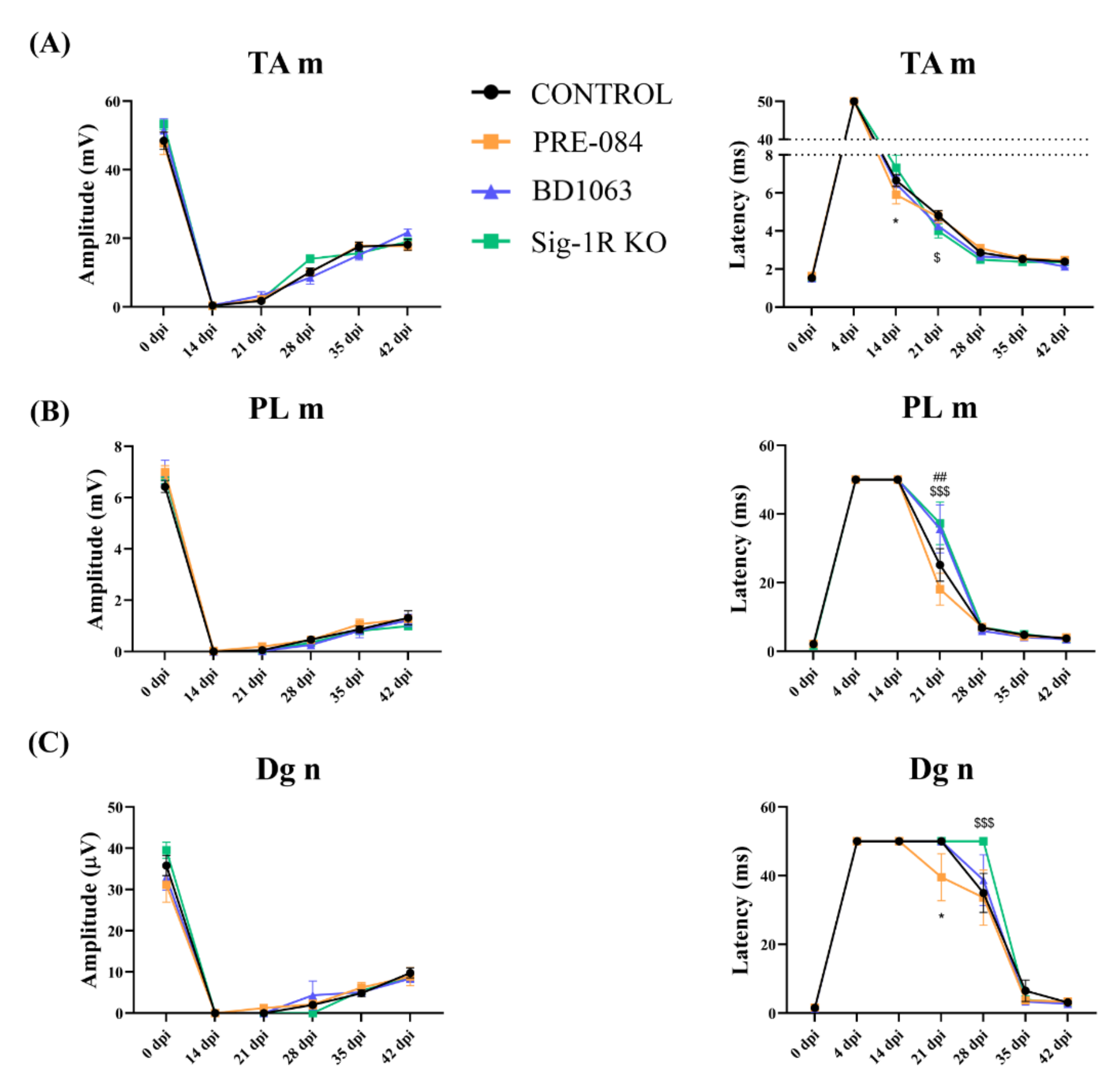
2.3. Electrophysiological Tests
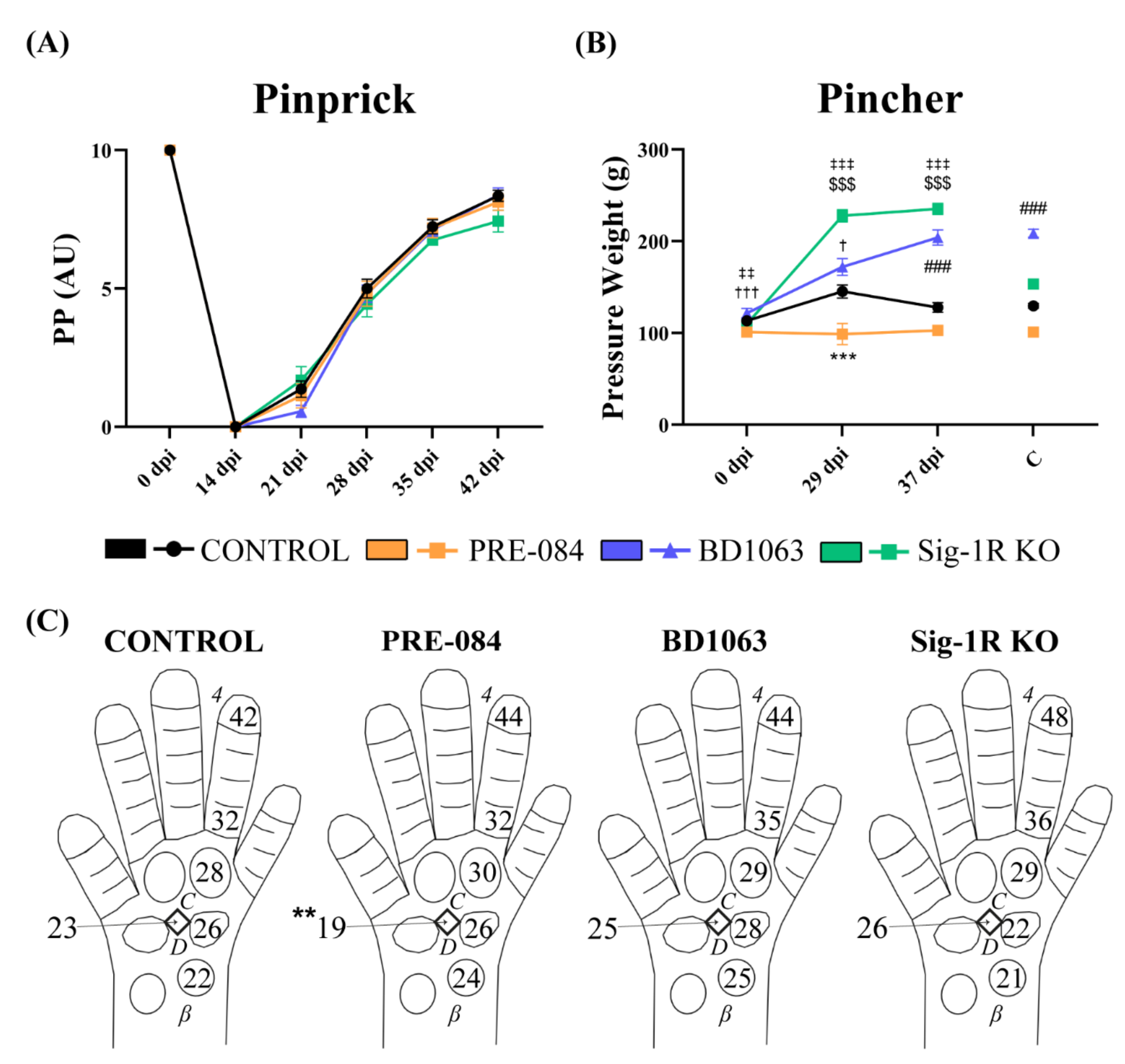
2.4. Pinprick Test
2.5. Pincher Test
2.6. Immunohistochemical Analyses
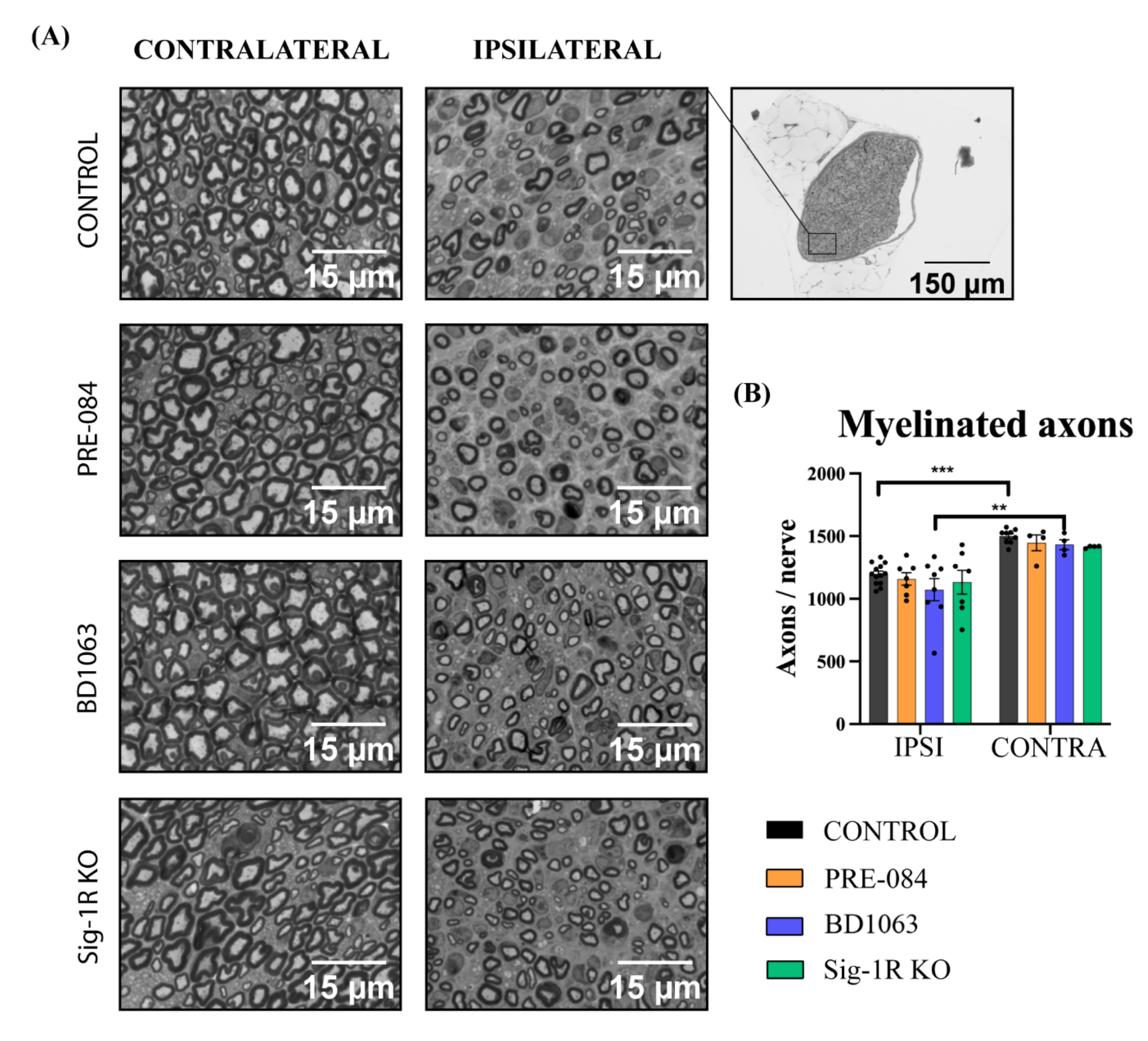
2.7. Histological Assessment of Regenerated Myelinated Axons
2.8. Statistical Analyses
3. Results
3.1. Functional Motor Recovery
3.2. Electrophysiological Results of Nerve Regeneration after Sciatic Nerve Section and Repair
3.3. Treatment with Sig-1R Antagonist BD1063 Produces Hyposensitivity after Nerve Injury
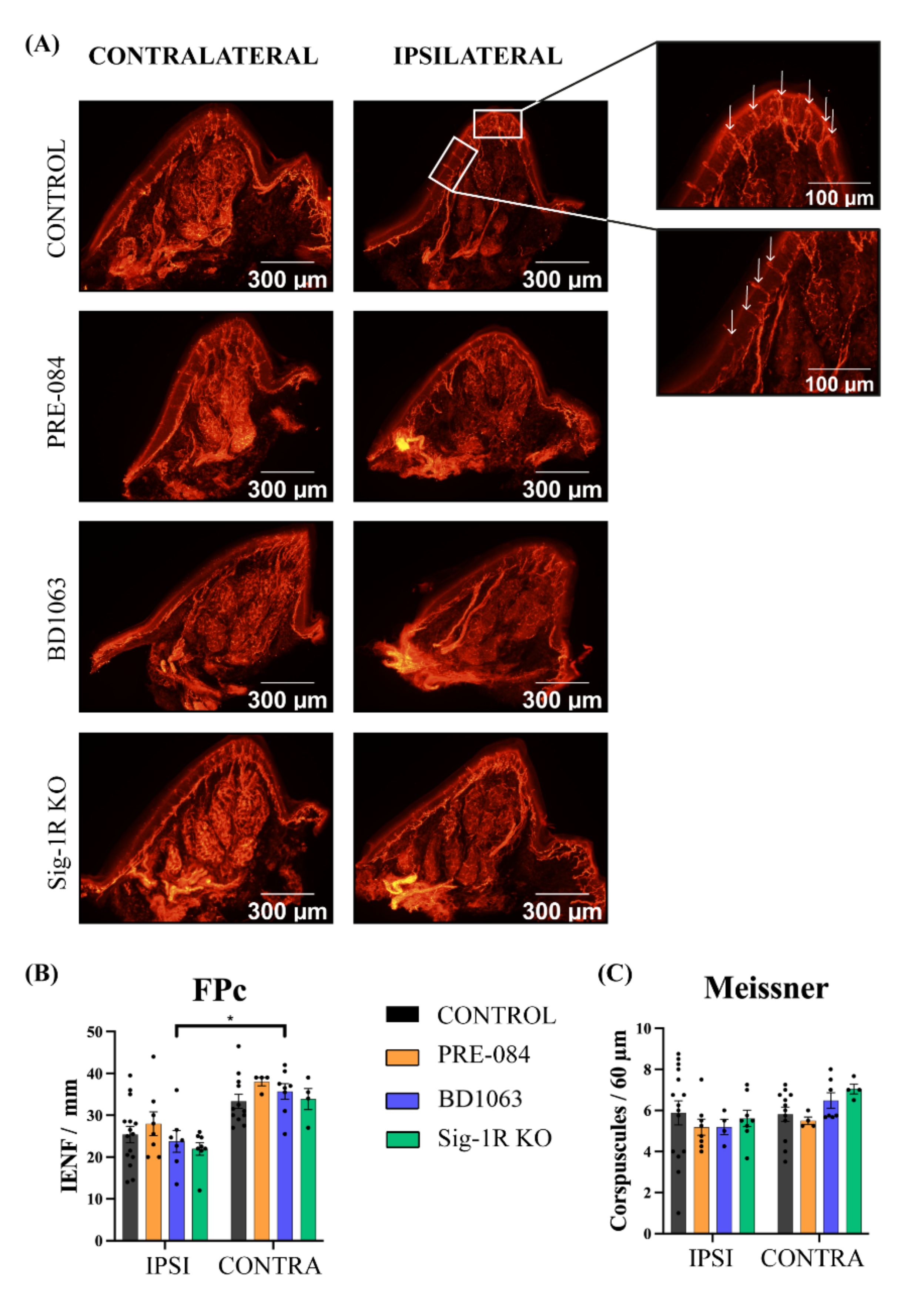
3.4. Skin Reinnervation and Number of Regenerated Axons after Nerve Injury
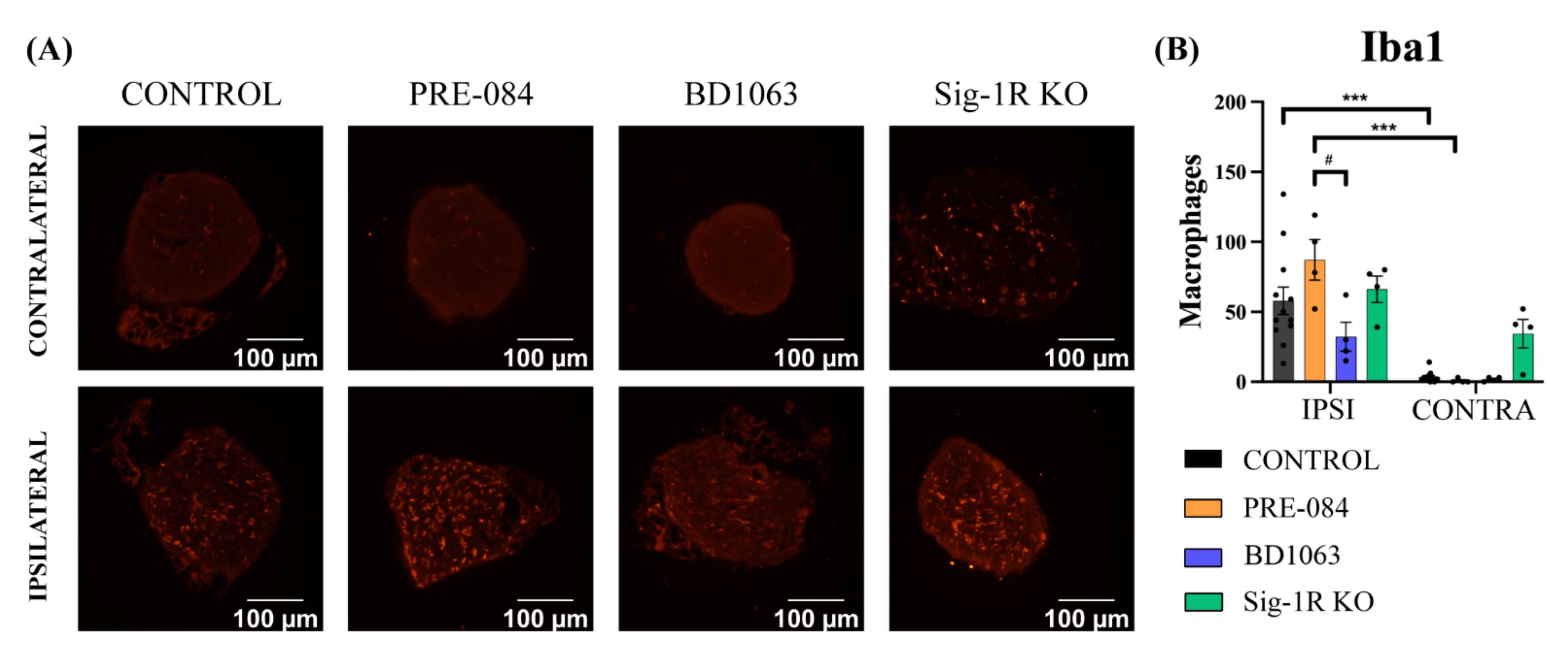
3.5. BD1063 Treatment Reduces Infiltrating Macrophages in the Injured Nerve
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pfister, B.; Gordon, T.; Loverde, J.; Kochar, A.; Mackinnon, S.; Cullen, D. Biomedical engineering strategies for peripheral nerve repair: Surgical applications, state of the art, and future challenges. Crit. Rev. Biomed. Eng. 2011, 39, 81–124. [Google Scholar] [CrossRef] [PubMed]
- Langa, F.; Codony, X.; Tovar, V.; Lavado, A.; Giménez, E.; Cozar, P.; Cantero, M.; Dordal, A.; Hernández, E.; Pérez, R.; et al. Generation and phenotypic analysis of sigma receptor type I (σ1) knockout mice. Eur. J. Neurosci. 2003, 18, 2188–2196. [Google Scholar] [CrossRef] [PubMed]
- Weng, T.Y.; Tsai, S.A.; Su, T.P. Roles of sigma-1 receptors on mitochondrial functions relevant to neurodegenerative diseases. J. Biomed. Sci. 2017, 24, 74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herrando-Grabulosa, M.; Gaja-Capdevila, N.; Vela, J.M.; Navarro, X. Sigma 1 receptor as a therapeutic target for amyotrophic lateral sclerosis. Br. J. Pharmacol. 2021, 178, 1336–1352. [Google Scholar] [CrossRef] [PubMed]
- Tarasov, A.I.; Griffiths, E.J.; Rutter, G.A. Regulation of ATP production by mitochondrial Ca2+. Cell Calcium 2012, 52, 28–35. [Google Scholar] [CrossRef] [Green Version]
- Hayashi, T.; Su, T.P. Sigma-1 receptor chaperones at the ER-mitochondrion interface regulate Ca2+ signaling and cell survival. Cell 2007, 131, 596–610. [Google Scholar] [CrossRef] [Green Version]
- Li, D.; Zhang, S.Z.; Yao, Y.H.; Xiang, Y.; Ma, X.Y.; Wei, X.L.; Yan, H.T.; Liu, X.Y. Sigma-1 receptor agonist increases axon outgrowth of hippocampal neurons via voltage-gated calcium ions channels. CNS Neurosci. Ther. 2017, 23, 930–939. [Google Scholar] [CrossRef] [Green Version]
- Guzmán-Lenis, M.S.; Navarro, X.; Casas, C. Selective sigma receptor agonist 2-(4-morpholinethyl)1-phenylcyclohexanecarboxylate (PRE084) promotes neuroprotection and neurite elongation through protein kinase C (PKC) signaling on motoneurons. Neuroscience 2009, 162, 31–38. [Google Scholar] [CrossRef]
- Matsushima, Y.; Terada, K.; Takata, J.; Karube, Y.; Kamei, C.; Sugimoto, Y. Effects of fluvoxamine on nerve growth factor-induced neurite outgrowth inhibition by dexamethasone in PC12 cells. Biosci. Biotechnol. Biochem. 2019, 83, 659–665. [Google Scholar] [CrossRef]
- Matsushima, Y.; Terada, K.; Kamei, C.; Sugimoto, Y. Sertraline inhibits nerve growth factor-induced neurite outgrowth in PC12 cells via a mechanism involving the sigma-1 receptor. Eur. J. Pharmacol. 2019, 853, 129–135. [Google Scholar] [CrossRef]
- Tsai, S.A.; Su, T.P. Sigma-1 receptors fine-tune the neuronal networks. Adv. Exp. Med. Biol. 2017, 964, 79–83. [Google Scholar] [CrossRef] [PubMed]
- Entrena, J.M.; Cobos, E.J.; Nieto, F.R.; Cendán, C.M.; Gris, G.; Del Pozo, E.; Zamanillo, D.; Baeyens, J.M. Sigma-1 receptors are essential for capsaicin-induced mechanical hypersensitivity: Studies with selective sigma-1 ligands and sigma-1 knockout mice. Pain 2009, 143, 252–261. [Google Scholar] [CrossRef] [PubMed]
- Nieto, F.R.; Cendán, C.M.; Sánchez-Fernández, C.; Cobos, E.J.; Entrena, J.M.; Tejada, M.A.; Zamanillo, D.; Vela, J.M.; Baeyens, J.M. Role of sigma-1 receptors in paclitaxel-induced neuropathic pain in mice. J. Pain 2012, 13, 1107–1121. [Google Scholar] [CrossRef]
- Espinosa-Juárez, J.V.; Jaramillo-Morales, O.A.; Déciga-Campos, M.; Moreno-Rocha, L.A.; López-Muñoz, F.J. Sigma-1 receptor antagonist (BD-1063) potentiates the antinociceptive effect of quercetin in neuropathic pain induced by chronic constriction injury. Drug Dev. Res. 2021, 82, 267–277. [Google Scholar] [CrossRef]
- Peng, Y.; Zhang, Q.; Welsh, W.J. Novel Sigma 1 Receptor Antagonists as Potential Therapeutics for Pain Management. J. Med. Chem. 2021, 64, 890–904. [Google Scholar] [CrossRef] [PubMed]
- Bravo-Caparrós, I.; Perazzoli, G.; Yeste, S.; Cikes, D.; Baeyens, J.M.; Cobos, E.J.; Nieto, F.R. Sigma-1 receptor inhibition reduces neuropathic pain induced by partial sciatic nerve transection in mice by opioid-dependent and -independent mechanisms. Front. Pharmacol. 2019, 10, 613. [Google Scholar] [CrossRef] [PubMed]
- Paniagua, N.; Girón, R.; Goicoechea, C.; López-Miranda, V.; Vela, J.M.; Merlos, M.; Martín Fontelles, M.I. Blockade of sigma 1 receptors alleviates sensory signs of diabetic neuropathy in rats. Eur. J. Pain 2017, 21, 61–72. [Google Scholar] [CrossRef] [PubMed]
- Bruna, J.; Videla, S.; Argyriou, A.A.; Velasco, R.; Villoria, J.; Santos, C.; Nadal, C.; Cavaletti, G.; Alberti, P.; Briani, C.; et al. Efficacy of a Novel Sigma-1 Receptor Antagonist for Oxaliplatin-Induced Neuropathy: A Randomized, Double-Blind, Placebo-Controlled Phase IIa Clinical Trial. Neurotherapeutics 2018, 15, 178–189. [Google Scholar] [CrossRef]
- Mei, J.; Pasternak, G.W. Sigma1 receptor modulation of opioid analgesia in the mouse. J. Pharmacol. Exp. Ther. 2002, 300, 1070–1074. [Google Scholar] [CrossRef] [Green Version]
- Zamanillo, D.; Romero, L.; Merlos, M.; Vela, J.M. Sigma 1 receptor: A new therapeutic target for pain. Eur. J. Pharmacol. 2013, 716, 78–93. [Google Scholar] [CrossRef]
- Bravo-Caparrós, I.; Ruiz-Cantero, M.C.; Perazzoli, G.; Cronin, S.J.F.; Vela, J.M.; Hamed, M.F.; Penninger, J.M.; Baeyens, J.M.; Cobos, E.J.; Nieto, F.R. Sigma-1 receptors control neuropathic pain and macrophage infiltration into the dorsal root ganglion after peripheral nerve injury. FASEB J. 2020, 34, 5951–5966. [Google Scholar] [CrossRef] [PubMed]
- Penas, C.; Pascual-Font, A.; Mancuso, R.; Forés, J.; Casas, C.; Navarro, X. Sigma receptor agonist 2-(4-morpholinethyl)1 phenylcyclohexanecarboxylate (Pre084) increases GDNF and BiP expression and promotes neuroprotection after root avulsion injury. J. Neurotrauma 2011, 28, 831–840. [Google Scholar] [CrossRef] [PubMed]
- Gaja-Capdevila, N.; Hernández, N.; Zamanillo, D.; Vela, J.M.; Merlos, M.; Navarro, X.; Herrando-Grabulosa, M. Neuroprotective effects of sigma 1 receptor ligands on motoneuron death after spinal root injury in mice. Int. J. Mol. Sci. 2021, 22, 6956. [Google Scholar] [CrossRef] [PubMed]
- Tejada, M.A.; Montilla-García, A.; Sánchez-Fernández, C.; Entrena, J.M.; Perazzoli, G.; Baeyens, J.M.; Cobos, E.J. Sigma-1 receptor inhibition reverses acute inflammatory hyperalgesia in mice: Role of peripheral sigma-1 receptors. Psychopharmacology 2014, 231, 3855–3869. [Google Scholar] [CrossRef]
- Gaja-Capdevila, N.; Hernández, N.; Navarro, X.; Herrando-Grabulosa, M. Sigma-1 Receptor is a Pharmacological Target to Promote Neuroprotection in the SOD1G93A ALS Mice. Front. Pharmacol. 2021, 12, 780588. [Google Scholar] [CrossRef]
- Navarro, X. Functional evaluation of peripheral nerve regeneration and target reinnervation in animal models: A critical overview. Eur. J. Neurosci. 2016, 43, 274–286. [Google Scholar] [CrossRef]
- Cobianchi, S.; de Cruz, J.; Navarro, X. Assessment of sensory thresholds and nociceptive fiber growth after sciatic nerve injury reveal the differential contribution of collateral reinnervation and nerve regeneration to neuropathic pain. Exp. Neurol. 2014, 255, 1–11. [Google Scholar] [CrossRef]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [Green Version]
- Saulite, L.; Vavers, E.; Zvejniece, L.; Dambrova, M.; Riekstina, U. The differentiation of skin mesenchymal stem cells towards a Schwann cell phenotype: Impact of sigma-1 receptor activation. Mol. Neurobiol. 2018, 55, 2840–2850. [Google Scholar] [CrossRef]
- Mancuso, R.; Oliván, S.; Rando, A.; Casas, C.; Osta, R.; Navarro, X. Sigma-1R agonist improves motor function and motoneuron survival in ALS mice. Neurotherapeutics 2012, 9, 814–826. [Google Scholar] [CrossRef] [Green Version]
- Entrena, J.M.; Sánchez-Fernández, C.; Nieto, F.R.; González-Cano, R.; Yeste, S.; Cobos, E.J.; Baeyens, J.M. Sigma-1 receptor agonism promotes mechanical allodynia after priming the nociceptive system with capsaicin. Sci. Rep. 2016, 6, 37835. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruscher, K.; Inácio, A.R.; Valind, K.; Rowshan, R.A.; Kuric, E.; Wieloch, T. Effects of the sigma-1 receptor agonist 1-(3,4-dimethoxyphenethyl)-4-(3-phenylpropyl)-piperazine dihydro-chloride on inflammation after stroke. PLoS ONE 2012, 7, e45118. [Google Scholar] [CrossRef] [PubMed]
- Cerveró, C.; Blasco, A.; Tarabal, O.; Casanovas, A.; Piedrafita, L.; Navarro, X.; Esquerda, J.E.; Calderó, J. Glial activation and central synapse loss, but not motoneuron degeneration, are prevented by the sigma-1 receptor agonist PRE-084 in the Smn2B/- mouse model of spinal muscular atrophy. J. Neuropathol. Exp. Neurol. 2018, 77, 577–597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jia, J.; Cheng, J.; Wang, C.; Zhen, X. Sigma-1 receptor-modulated neuroinflammation in neurological diseases. Front. Cell Neurosci. 2018, 12, 314. [Google Scholar] [CrossRef] [Green Version]
- Jeong, Y.C.; Son, J.S.; Kwon, Y.B. The spinal antinociceptive mechanism determined by systemic administration of BD1047 in zymosan-induced hyperalgesia in rats. Brain Res. Bull. 2015, 119, 93–100. [Google Scholar] [CrossRef]
- Zhu, S.; Wang, C.; Han, Y.; Song, C.; Hu, X.; Liu, Y. Sigma-1 receptor antagonist BD1047 reduces mechanical allodynia in a rat model of bone cancer pain through the inhibition of spinal nr1 phosphorylation and microglia activation. Mediat. Inflamm. 2015, 2015, 265056. [Google Scholar] [CrossRef] [Green Version]
- Ruiz-Cantero, M.C.; González-Cano, R.; Tejada, M.; Santos-Caballero, M.; Perazzoli, G.; Nieto, F.R.; Cobos, E.J. Sigma-1 receptor: A drug target for the modulation of neuroimmune and neuroglial interactions during chronic pain. Pharmacol. Res. 2021, 163, 105339. [Google Scholar] [CrossRef]
- Maurice, T. Bi-phasic dose response in the preclinical and clinical developments of sigma-1 receptor ligands for the treatment of neurodegenerative disorders. Expert Opin. Drug Discov. 2021, 16, 373–389. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cottilli, P.; Gaja-Capdevila, N.; Navarro, X. Effects of Sigma-1 Receptor Ligands on Peripheral Nerve Regeneration. Cells 2022, 11, 1083. https://doi.org/10.3390/cells11071083
Cottilli P, Gaja-Capdevila N, Navarro X. Effects of Sigma-1 Receptor Ligands on Peripheral Nerve Regeneration. Cells. 2022; 11(7):1083. https://doi.org/10.3390/cells11071083
Chicago/Turabian StyleCottilli, Patrick, Núria Gaja-Capdevila, and Xavier Navarro. 2022. "Effects of Sigma-1 Receptor Ligands on Peripheral Nerve Regeneration" Cells 11, no. 7: 1083. https://doi.org/10.3390/cells11071083
APA StyleCottilli, P., Gaja-Capdevila, N., & Navarro, X. (2022). Effects of Sigma-1 Receptor Ligands on Peripheral Nerve Regeneration. Cells, 11(7), 1083. https://doi.org/10.3390/cells11071083