
The Novel Competing Endogenous Long Noncoding RNA SM2 Regulates Gonadotropin Secretion in the Hu Sheep Anterior Pituitary by Targeting the Oar-miR-16b/TGF-β/SMAD2 Signaling Pathway


Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Sample Collection
2.2. Cell Culture and Treatments
2.3. Vector Construction
2.4. Cell Transfection and Treatment
2.5. lncRNA Coding Ability Prediction and miRNA Prediction
2.6. Quantitative Real-Time Polymerase Chain Reaction (qPCR)
2.7. EDU Assay
2.8. Flow Cytometry Analysis
2.9. Immunofluorescence (IF)
2.10. Western Blot
2.11. FISH Analysis
2.12. ELISA Assay
2.13. Dual-Luciferase Reporter Assays
2.14. Data Analysis
3. Results
3.1. lncRNA SM2 Was Highly Expressed in Pituitary Gland
3.2. lncRNA SM2 Knockdown Downregulated the Secretion of Gonadotropins in Pituitary Cells
3.3. lncRNA SM2 Knockdown Compromised Pituitary Cell Proliferation and Viability
3.4. lncRNA SM2 Regulates SMAD2 Via oar-miR-16b
3.5. Effect of Oar-miR-16b Level on Pituitary Cell Proliferation and Apoptosis
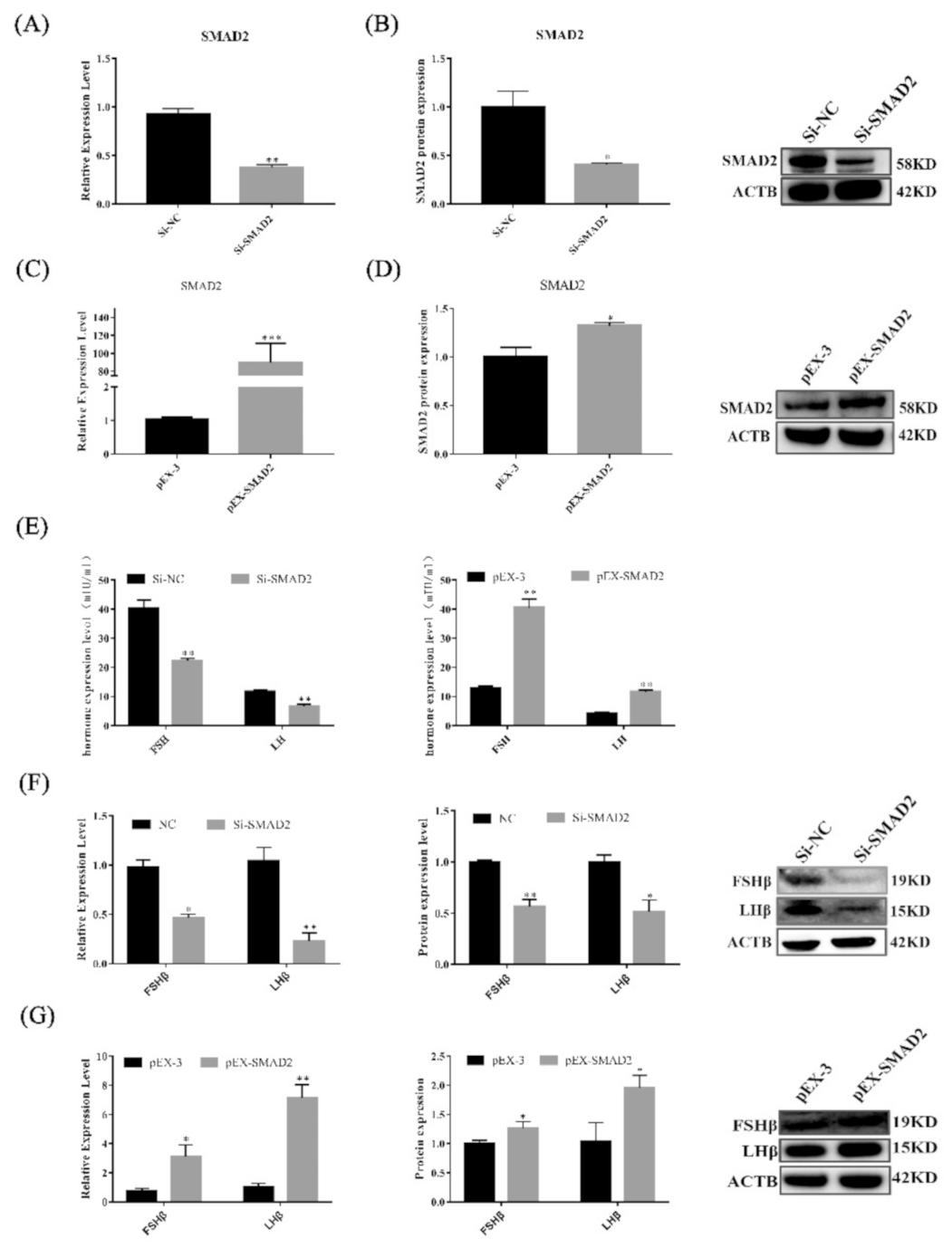
3.6. SMAD2 Overexpression Upregulates the Secretion of Gonadotropins in Pituitary Cells
3.7. SMAD2 Overexpression Upregulated Pituitary Cell Proliferation and Viability
3.8. lncRNA SM2 Is Involved in Pituitary Cells by Regulating the TGF-β/SMAD2 Pathway
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Takahashi, S. Intrapituitary regulatory system of proliferation of mammotrophs in the pituitary gland. Zoolog. Sci. 2004, 21, 601–611. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Reichlin, S. Neuroendocrinology of the pituitary gland. Toxicol. Pathol. 1989, 17, 250–255. [Google Scholar] [CrossRef] [PubMed]
- Farrow, P.; Simmons, J.G.; Pozzi, E.; Diaz-Arteche, C.; Richmond, S.; Bray, K.; Schwartz, O.; Whittle, S. Associations between early life stress and anterior pituitary gland volume development during late childhood. Psychoneuroendocrinology 2020, 122, 104868. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, K.; Yamamoto, K.; Kikuyama, S.; Tanaami, Y.; Machida, T.; Kobayashi, T. Delayed Postnatal Growth and Anterior Pituitary Development in Growth-Retarded (grt) Female Mice. Zoolog. Sci. 2021, 38, 238–246. [Google Scholar] [CrossRef] [PubMed]
- Shalev, D.; Melamed, P. The role of the hypothalamus and pituitary epigenomes in central activation of the reproductive axis at puberty. Mol. Cell Endocrinol. 2020, 518, 111031. [Google Scholar] [CrossRef] [PubMed]
- Ulloa-Aguirre, A.; Lira-Albarran, S. Clinical Applications of Gonadotropins in the Male. Prog. Mol. Biol. Transl. Sci. 2016, 143, 121–174. [Google Scholar] [CrossRef]
- Hillier, S.G. Gonadotropic control of ovarian follicular growth and development. Mol. Cell Endocrinol. 2001, 179, 39–46. [Google Scholar] [CrossRef]
- Kumar, T.R.; Wang, Y.; Lu, N.; Matzuk, M.M. Follicle stimulating hormone is required for ovarian follicle maturation but not male fertility. Nat. Genet 1997, 15, 201–204. [Google Scholar] [CrossRef]
- Qin, S.; Predescu, D.; Carman, B.; Patel, P.; Chen, J.; Kim, M.; Lahm, T.; Geraci, M.; Predescu, S.A. Up-Regulation of the Long Noncoding RNA X-Inactive-Specific Transcript and the Sex Bias in Pulmonary Arterial Hypertension. Am. J. Pathol. 2021, 191, 1135–1150. [Google Scholar] [CrossRef]
- Ni, W.; Yao, S.; Zhou, Y.; Liu, Y.; Huang, P.; Zhou, A.; Liu, J.; Che, L.; Li, J. Long noncoding RNA GAS5 inhibits progression of colorectal cancer by interacting with and triggering YAP phosphorylation and degradation and is negatively regulated by the m(6)A reader YTHDF3. Mol. Cancer 2019, 18, 143. [Google Scholar] [CrossRef]
- Kitagawa, M.; Kitagawa, K.; Kotake, Y.; Niida, H.; Ohhata, T. Cell cycle regulation by long non-coding RNAs. Cell Mol. Life Sci. 2013, 70, 4785–4794. [Google Scholar] [CrossRef] [PubMed]
- Cai, B.; Li, Z.; Ma, M.; Zhang, J.; Kong, S.; Abdalla, B.A.; Xu, H.; Jebessa, E.; Zhang, X.; Lawal, R.A.; et al. Long noncoding RNA SMUL suppresses SMURF2 production-mediated muscle atrophy via nonsense-mediated mRNA decay. Mol. Ther. Nucleic Acids 2021, 23, 512–526. [Google Scholar] [CrossRef] [PubMed]
- Hong, G.K.; Payne, S.C.; Jane, J.A., Jr. Anatomy, Physiology, and Laboratory Evaluation of the Pituitary Gland. Otolaryngol. Clin. N. Am. 2016, 49, 21–32. [Google Scholar] [CrossRef] [PubMed]
- Che, Q.; Liu, M.; Zhang, D.; Lu, Y.; Xu, J.; Lu, X.; Cao, X.; Liu, Y.; Dong, X.; Liu, S. Long Noncoding RNA HUPCOS Promotes Follicular Fluid Androgen Excess in PCOS Patients via Aromatase Inhibition. J. Clin. Endocrinol. Metab. 2020, 105. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Wang, H.; Zhou, D.; Shuang, T.; Zhao, H.; Chen, B. Up-Regulation of Long Noncoding RNA SRA Promotes Cell Growth, Inhibits Cell Apoptosis, and Induces Secretion of Estradiol and Progesterone in Ovarian Granular Cells of Mice. Med. Sci. Monit. 2018, 24, 2384–2390. [Google Scholar] [CrossRef]
- Salama, R.M.; Abd Elwahab, A.H.; Abd-Elgalil, M.M.; Elmongy, N.F.; Schaalan, M.F. LCZ696 (sacubitril/valsartan) protects against cyclophosphamide-induced testicular toxicity in rats: Role of neprilysin inhibition and lncRNA TUG1 in ameliorating apoptosis. Toxicology 2020, 437, 152439. [Google Scholar] [CrossRef]
- Wang, X.; Yang, C.; Guo, F.; Zhang, Y.; Ju, Z.; Jiang, Q.; Zhao, X.; Liu, Y.; Zhao, H.; Wang, J.; et al. Integrated analysis of mRNAs and long noncoding RNAs in the semen from Holstein bulls with high and low sperm motility. Sci. Rep. 2019, 9, 2092. [Google Scholar] [CrossRef]
- Peng, Y.; Chang, L.; Wang, Y.; Wang, R.; Hu, L.; Zhao, Z.; Geng, L.; Liu, Z.; Gong, Y.; Li, J.; et al. Genome-wide differential expression of long noncoding RNAs and mRNAs in ovarian follicles of two different chicken breeds. Genomics 2019, 111, 1395–1403. [Google Scholar] [CrossRef]
- Hou, J.; Long, H.; Zhou, C.; Zheng, S.; Wu, H.; Guo, T.; Wu, Q.; Zhong, T.; Wang, T. Long noncoding RNA Braveheart promotes cardiogenic differentiation of mesenchymal stem cells in vitro. Stem. Cell Res. Ther. 2017, 8, 4. [Google Scholar] [CrossRef]
- Salmena, L.; Poliseno, L.; Tay, Y.; Kats, L.; Pandolfi, P.P. A ceRNA hypothesis: The Rosetta Stone of a hidden RNA language? Cell 2011, 146, 353–358. [Google Scholar] [CrossRef]
- Zheng, X.; Zhao, K.; Liu, T.; Liu, L.; Zhou, C.; Xu, M. Long noncoding RNA PVT1 promotes laryngeal squamous cell carcinoma development by acting as a molecular sponge to regulate miR-519d-3p. J. Cell Biochem. 2019, 120, 3911–3921. [Google Scholar] [CrossRef] [PubMed]
- Yao, X.; Gao, X.; Bao, Y.; El-Samahy, M.A.; Yang, J.; Wang, Z.; Li, X.; Zhang, G.; Zhang, Y.; Liu, W.; et al. lncRNA FDNCR promotes apoptosis of granulosa cells by targeting the miR-543-3p/DCN/TGF-beta signaling pathway in Hu sheep. Mol. Ther. Nucleic Acids 2021, 24, 223–240. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.J.; Horlbeck, M.A.; Cho, S.W.; Birk, H.S.; Malatesta, M.; He, D.; Attenello, F.J.; Villalta, J.E.; Cho, M.Y.; Chen, Y.; et al. CRISPRi-based genome-scale identification of functional long noncoding RNA loci in human cells. Science 2017, 355, aah7111. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Lan, Y.; Xie, A.; Shi, J.; Zhao, H.; Xu, L.; Zhu, S.; Luo, T.; Zhao, T.; Xiao, Y.; et al. Comprehensive analysis of long noncoding RNA (lncRNA)-chromatin interactions reveals lncRNA functions dependent on binding diverse regulatory elements. J. Biol. Chem. 2019, 294, 15613–15622. [Google Scholar] [CrossRef]
- Han, D.X.; Sun, X.L.; Wang, C.J.; Yu, Z.W.; Zheng, Y.; Huang, Y.J.; Wang, W.H.; Jiang, H.; Gao, Y.; Yuan, B.; et al. Differentially expressed lncRNA-m433s1 regulates FSH secretion by functioning as a miRNA sponge in male rat anterior pituitary cellsdagger. Biol. Reprod. 2019, 101, 416–425. [Google Scholar] [CrossRef]
- Du, Q.; Hoover, A.R.; Dozmorov, I.; Raj, P.; Khan, S.; Molina, E.; Chang, T.C.; de la Morena, M.T.; Cleaver, O.B.; Mendell, J.T.; et al. MIR205HG Is a Long Noncoding RNA that Regulates Growth Hormone and Prolactin Production in the Anterior Pituitary. Dev. Cell 2019, 49, 618–631.e5. [Google Scholar] [CrossRef]
- Tang, H.; Zhu, D.; Zhang, G.; Luo, X.; Xie, W. AFAP1-AS1 Promotes Proliferation of Pituitary Adenoma Cells through miR-103a-3p to Activate PI3K/AKT Signaling Pathway. World Neurosurg. 2019, 130, e888–e898. [Google Scholar] [CrossRef]
- D’Angelo, D.; Mussnich, P.; Sepe, R.; Raia, M.; Del Vecchio, L.; Cappabianca, P.; Pellecchia, S.; Petrosino, S.; Saggio, S.; Solari, D.; et al. RPSAP52 lncRNA is overexpressed in pituitary tumors and promotes cell proliferation by acting as miRNA sponge for HMGA proteins. J. Mol. Med. 2019, 97, 1019–1032. [Google Scholar] [CrossRef]
- Feng, X.; Li, F.; Wang, F.; Zhang, G.; Pang, J.; Ren, C.; Zhang, T.; Yang, H.; Wang, Z.; Zhang, Y. Genome-wide differential expression profiling of mRNAs and lncRNAs associated with prolificacy in Hu sheep. Biosci. Rep. 2018, 38. [Google Scholar] [CrossRef]
- Guthrie, H.D.; Cooper, B.S. Follicular Atresia, follicular fluid hormones, and circulating hormones during the midluteal phase of the estrous cycle in pigs. Biol. Reprod. 1996, 55, 543–547. [Google Scholar] [CrossRef]
- Li, Y.; Schang, G.; Boehm, U.; Deng, C.X.; Graff, J.; Bernard, D.J. SMAD3 Regulates Follicle-stimulating Hormone Synthesis by Pituitary Gonadotrope Cells in Vivo. J. Biol. Chem. 2017, 292, 2301–2314. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Zhang, Y.; Xu, X.; Li, J.; Yuan, F.; Bo, S.; Qiao, J.; Xia, G.; Su, Y.; Zhang, M. Transforming growth factor-beta is involved in maintaining oocyte meiotic arrest by promoting natriuretic peptide type C expression in mouse granulosa cells. Cell Death Dis. 2019, 10, 558. [Google Scholar] [CrossRef] [PubMed]
- Kotarba, G.; Zielinska-Gorska, M.; Biernacka, K.; Gajewska, A. Gonadotropin-releasing hormone-Cu complex (Cu-GnRH) transcriptional activity in vivo in the female rat anterior pituitary gland. Brain Res. Bull. 2020, 156, 67–75. [Google Scholar] [CrossRef] [PubMed]
- Lajko, E.; Pallinger, E.; Kovacs, Z.; Szabo, I.; Kohidai, L. Effects of Gonadotropin-Releasing Hormone (GnRH) and Its Analogues on the Physiological Behaviors and Hormone Content of Tetrahymena pyriformis. Int. J. Mol. Sci. 2019, 20, 5711. [Google Scholar] [CrossRef] [PubMed]
- Park, S.R.; Cho, A.; Park, S.T.; Park, C.H.; Lim, S.; Jin, M.; Lee, H.Y.; Hong, I.S. Double-edged sword of gonadotropin-releasing hormone (GnRH): A novel role of GnRH in the multiple beneficial functions of endometrial stem cells. Cell Death Dis. 2018, 9, 828. [Google Scholar] [CrossRef] [PubMed]
- Deng, M.; Liu, Z.; Ren, C.; An, S.; Wan, Y.; Wang, F. Highly methylated Xist in SCNT embryos was retained in deceased cloned female goats. Reprod. Fertil. Dev. 2019, 31, 855–866. [Google Scholar] [CrossRef]
- Li, D.; Liu, H.; Li, Y.; Yang, M.; Qu, C.; Zhang, Y.; Liu, Y.; Zhang, X. Identification of suitable endogenous control genes for quantitative RT-PCR analysis of miRNA in bovine solid tissues. Mol. Biol. Rep. 2014, 41, 6475–6480. [Google Scholar] [CrossRef]
- Nygard, A.B.; Jorgensen, C.B.; Cirera, S.; Fredholm, M. Selection of reference genes for gene expression studies in pig tissues using SYBR green qPCR. BMC Mol. Biol. 2007, 8, 67. [Google Scholar] [CrossRef]
- Zheng, J.; Wang, Z.; Yang, H.; Yao, X.; Yang, P.; Ren, C.; Wang, F.; Zhang, Y. Pituitary Transcriptomic Study Reveals the Differential Regulation of lncRNAs and mRNAs Related to Prolificacy in Different FecB Genotyping Sheep. Genes 2019, 10, 157. [Google Scholar] [CrossRef]
- Liu, X.H.; Sun, M.; Nie, F.Q.; Ge, Y.B.; Zhang, E.B.; Yin, D.D.; Kong, R.; Xia, R.; Lu, K.H.; Li, J.H.; et al. Lnc RNA HOTAIR functions as a competing endogenous RNA to regulate HER2 expression by sponging miR-331-3p in gastric cancer. Mol. Cancer 2014, 13, 92. [Google Scholar] [CrossRef]
- Wang, W.T.; Ye, H.; Wei, P.P.; Han, B.W.; He, B.; Chen, Z.H.; Chen, Y.Q. LncRNAs H19 and HULC, activated by oxidative stress, promote cell migration and invasion in cholangiocarcinoma through a ceRNA manner. J. Hematol. Oncol. 2016, 9, 117. [Google Scholar] [CrossRef] [PubMed]
- Im, H.I.; Kenny, P.J. MicroRNAs in neuronal function and dysfunction. Trends Neurosci. 2012, 35, 325–334. [Google Scholar] [CrossRef] [PubMed]
- Cao, C.; Ding, Y.; Kong, X.; Feng, G.; Xiang, W.; Chen, L.; Yang, F.; Zhang, K.; Chu, M.; Wang, P.; et al. Reproductive role of miRNA in the hypothalamic-pituitary axis. Mol. Cell Neurosci. 2018, 88, 130–137. [Google Scholar] [CrossRef] [PubMed]
- Bak, M.; Silahtaroglu, A.; Moller, M.; Christensen, M.; Rath, M.F.; Skryabin, B.; Tommerup, N.; Kauppinen, S. MicroRNA expression in the adult mouse central nervous system. RNA 2008, 14, 432–444. [Google Scholar] [CrossRef] [PubMed]
- Ye, R.S.; Li, M.; Li, C.Y.; Qi, Q.E.; Chen, T.; Cheng, X.; Wang, S.B.; Shu, G.; Wang, L.N.; Zhu, X.T.; et al. miR-361-3p regulates FSH by targeting FSHB in a porcine anterior pituitary cell model. Reproduction 2017, 153, 341–349. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Li, H.; Zhang, D.; Xu, G.; Zhang, J.; Cui, S. miR-7 mediates the signaling pathway of NE affecting FSH and LH synthesis in pig pituitary. J. Endocrinol. 2020, 244, 459–471. [Google Scholar] [CrossRef] [PubMed]
- Fortin, J.; Boehm, U.; Deng, C.X.; Treier, M.; Bernard, D.J. Follicle-stimulating hormone synthesis and fertility depend on SMAD4 and FOXL2. FASEB J. 2014, 28, 3396–3410. [Google Scholar] [CrossRef]
- Gueripel, X.; Benahmed, M.; Gougeon, A. Sequential gonadotropin treatment of immature mice leads to amplification of transforming growth factor beta action, via upregulation of receptor-type 1, Smad 2 and 4, and downregulation of Smad 6. Biol. Reprod. 2004, 70, 640–648. [Google Scholar] [CrossRef][Green Version]
- Yang, H.; Wan, Z.; Jin, Y.; Wang, F.; Zhang, Y. SMAD2 regulates testicular development and testosterone synthesis in Hu sheep. Theriogenology 2021, 174, 139–148. [Google Scholar] [CrossRef]
- Luo, X.; Xu, J.; Chegini, N. Gonadotropin releasing hormone analogue (GnRHa) alters the expression and activation of Smad in human endometrial epithelial and stromal cells. Reprod. Biol. Endocrinol. 2003, 1, 125. [Google Scholar] [CrossRef]
- Fortin, J.; Ongaro, L.; Li, Y.; Tran, S.; Lamba, P.; Wang, Y.; Zhou, X.; Bernard, D.J. Minireview: Activin Signaling in Gonadotropes: What Does the FOX say... to the SMAD? Mol. Endocrinol. 2015, 29, 963–977. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Items | Primer Sequence (5′-3′) |
|---|---|
| siRNA-NC | S: 5′-UUCUCCGAACGUGUCACGUTT-3′ |
| A: 5′-ACGUGACACGUUCGGAGAATT-3′ | |
| siRNA-LncSM2-1 (3415) | S: 5′-GCGGCCAGAAUGUAAUUAATT-3′ |
| A: 5′-UUAAUUACAUUCUGGCCGCTT-3′ | |
| siRNA-LncSM2-2 (2592) | S: 5′-GCUGGAGAAUAAAGCAUAUTT-3′ |
| A: 5′-AUAUGCUUUAUUCUCCAGCTT-3′ | |
| siRNA-LncSM2-3 (3794) | S: 5′-CUCCAACACUAGGCAAAUATT-3′ |
| A: 5′-UAUUUGCCUAGUGUUGGAGTT-3′ | |
| siRNA-SMAD2-531 | S: 5′-GCCUGAUCUUCACAGUCAUTT-3′ |
| A: 5′-AUGACUGUGAAGAUCAGGCTT-3′ | |
| oar-miR-16b mimics | S: 5′-UAGCAGCACGUAAAUAUUGG-3′ |
| A: 5′-AAUAUUUACGUGCUGCUAUU-3′ | |
| NC mimics | S: 5′-UUCUCCGAACGUGUCACGUTT-3′ |
| A: 5′-ACGUGACACGUUCGGAGAATT-3′ | |
| oar-miR-16b inhibitor | 5’-CCAAUAUUUACGUGCUGCUA-3’ |
| NC inhibitor | 5’-CAGUACUUUUGUGUAGUACAA-3’ |
| Gene Name | Forward Primer (5′-3′) | Reverse Primer (5′-3′) | Usage |
|---|---|---|---|
| ACTB | CGCAAGTACTCCGTGTGGAT | TAACGCAGCTAACAGTCCGC | RT-qPCR |
| LncSM2 | TCTAGAACGCAAGTGCCTGG | GTCTGCAAGGAAAAGGCGAC | RT-qPCR |
| SMAD2 | GCAATCTTTGTGCAGAGCCC | TGCTTGTTACCGTCTGCCTT | RT-qPCR |
| PCNA | CCTTGGTGCAGCTAACCCTT | GCCAAGGTGTCCGCATTATC | RT-qPCR |
| BAX | TTGGCTGAGTCGCTGAAGAGC | AACTCCCATGGCCCCCAAAT | RT-qPCR |
| BCL2 | CCTTTCGTTTGCTCGTGCTC | ACCTCCTCCGTGATGTGGTAT | RT-qPCR |
| FSHB | GCTATTGCTACACCCGGGAC | AGTGGCTACTGGGTACGTGT | RT-qPCR |
| LHB | CCGCTCCCAGATATCCTCTTC | TTATTGGGAAGGGAGGGGAGG | RT-qPCR |
| GnRHR | GCTGCCTCTTCATCATCCCTCTTC | CCTCAGCCGAGCTTGTGGTATATTG | RT-qPCR |
| TGF-β1 | ACCGGCCCTTCCTGCTCCTCAT | GGAGCGCACGATCATGTTGGA | RT-qPCR |
| TGF-β2 | CCCCAGAAGACTACCTCG | AGTATTCCTCGTCGCTCC | RT-qPCR |
| TGF-β3 | CCACCTTGGACTTCAACC | CGGGTGCTGTTGTAAAGA | RT-qPCR |
| TGF-R1 | CTGTCGGAAAGCCGTCATCT | TCCTCTTCACTTGGCACTCG | RT-qPCR |
| TGF-R2 | TCGCCGAGGTCTACAAGG | GCGTGGAAGGCAGTGATG | RT-qPCR |
| U6 | CTCGCTTCGGCAGCACA | AACGCTTCACGAATTTGCGT | RT-qPCR |
| Genes | Primer Sequence (5′-3′) | Usage |
|---|---|---|
| oar-miR-16b | GTCGTATCCAGTGCAGGGTCCGAGGTATTCGCACTGGATACGACCCAATA | RT-qPCR |
| oar-miR-16b | F: GCGCGTAGCAGCACGTAAA | RT-qPCR |
| R: AGTGCAGGGTCCGAGGTATT | RT-qPCR |
| Antibodies | Cat No. | Source | Dilution of IF | Dilution of WB |
|---|---|---|---|---|
| SMAD2 | 12570-1-AP | Proteintech, Wuhan, China | 1:100 | 1:1000 |
| PCNA | AF0239 | AFFinity, Sterling, VA, USA | 1:1000 | |
| BAX | 50599-2-lg | Proteintech, Wuhan, China | 1:1000 | |
| BCL2 | 12789-1-AP | Proteintech, Wuhan, China | 1:500 | |
| FSHβ | AB180489 | Abcam, Cambridge, UK | 1:100 | 1:1000 |
| LHβ | AB150416 | Abcam, Cambridge, UK | 1:1000 | |
| LHβ | DF7140 | AFFinity, Sterling, VA, USA | 1:100 | |
| β-actin | bs-0061R | Proteintech, Wuhan, China | 1:1000 | |
| Goat anti-mouse lgG | 20536-1-AP | Proteintech, Wuhan, China | 1:2000 | |
| Goat anti-Rabbit lgG | SA00001-2 | Proteintech, Wuhan, China | 1:1000 | 1:2000 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wan, Z.; Yang, H.; Chen, P.; Wang, Z.; Cai, Y.; Yao, X.; Wang, F.; Zhang, Y. The Novel Competing Endogenous Long Noncoding RNA SM2 Regulates Gonadotropin Secretion in the Hu Sheep Anterior Pituitary by Targeting the Oar-miR-16b/TGF-β/SMAD2 Signaling Pathway. Cells 2022, 11, 985. https://doi.org/10.3390/cells11060985
Wan Z, Yang H, Chen P, Wang Z, Cai Y, Yao X, Wang F, Zhang Y. The Novel Competing Endogenous Long Noncoding RNA SM2 Regulates Gonadotropin Secretion in the Hu Sheep Anterior Pituitary by Targeting the Oar-miR-16b/TGF-β/SMAD2 Signaling Pathway. Cells. 2022; 11(6):985. https://doi.org/10.3390/cells11060985
Chicago/Turabian StyleWan, Zhen, Hua Yang, Peiyong Chen, Zhibo Wang, Yu Cai, Xiaolei Yao, Feng Wang, and Yanli Zhang. 2022. "The Novel Competing Endogenous Long Noncoding RNA SM2 Regulates Gonadotropin Secretion in the Hu Sheep Anterior Pituitary by Targeting the Oar-miR-16b/TGF-β/SMAD2 Signaling Pathway" Cells 11, no. 6: 985. https://doi.org/10.3390/cells11060985
APA StyleWan, Z., Yang, H., Chen, P., Wang, Z., Cai, Y., Yao, X., Wang, F., & Zhang, Y. (2022). The Novel Competing Endogenous Long Noncoding RNA SM2 Regulates Gonadotropin Secretion in the Hu Sheep Anterior Pituitary by Targeting the Oar-miR-16b/TGF-β/SMAD2 Signaling Pathway. Cells, 11(6), 985. https://doi.org/10.3390/cells11060985