
Axon-Targeting Motifs: Mechanisms and Applications of Enhancing Axonal Localisation of Transmembrane Proteins



Abstract
1. Introduction
2. Two Distinct Pathways Mediate Transmembrane Protein Trafficking in CNS Neurons
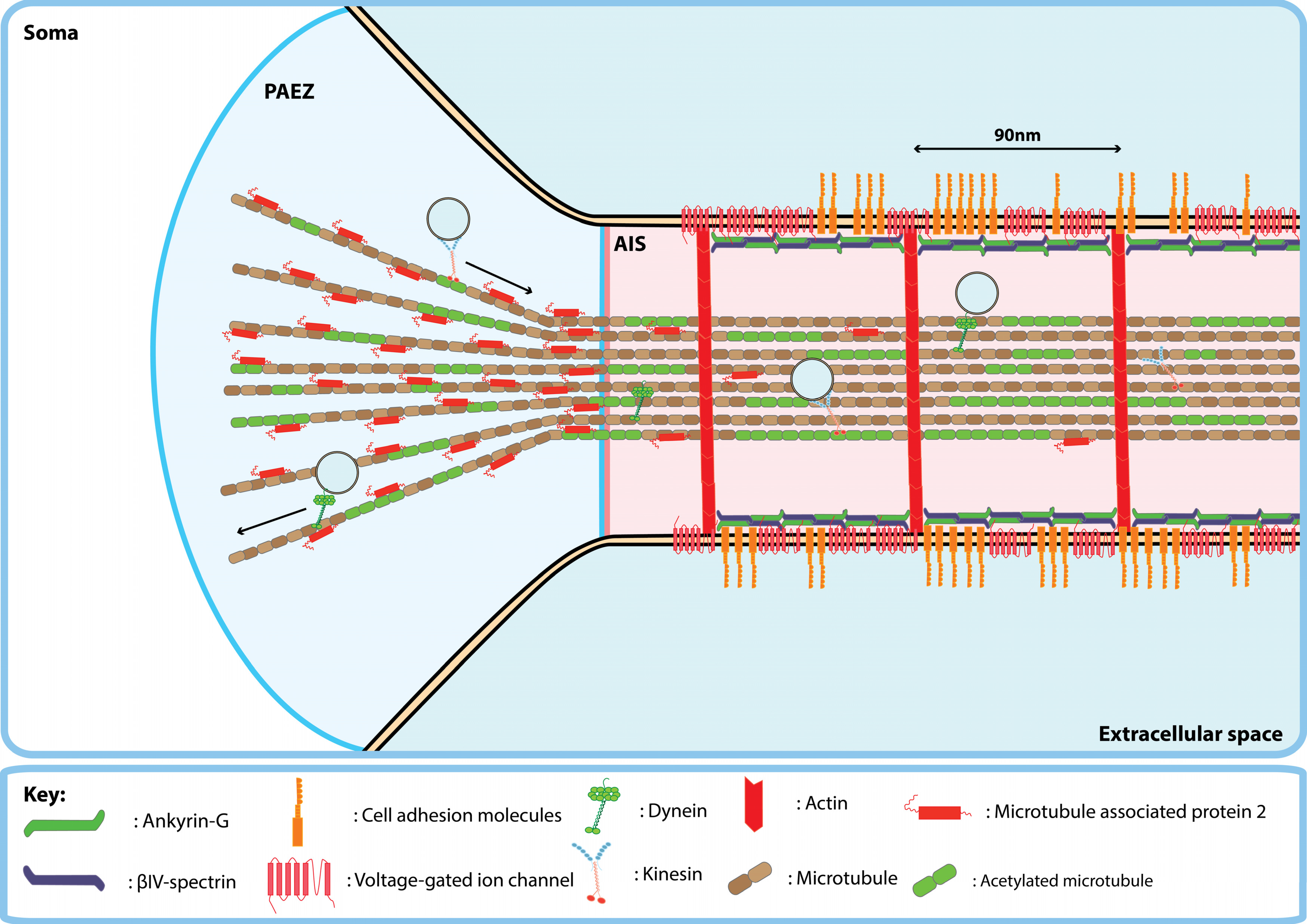
3. Maintenance of Transmembrane Protein Polarity by the Proximal Axon
4. Axon-Targeting Motifs Exploit Diverse Trafficking Pathways to Promote Axonal Localisation
5. Further Applications of ATMs
5.1. Improving Characterisation of Mammalian Neuronal Circuity
5.2. Improving Genetic Therapies for Axonopathies
6. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Smith, D.H. Stretch growth of integrated axon tracts: Extremes and exploitations. Prog. Neurobiol. 2009, 89, 231–239. [Google Scholar] [CrossRef] [PubMed]
- Esakakibara, A.; Ehatanaka, Y. Neuronal polarization in the developing cerebral cortex. Front. Neurosci. 2015, 9, 116. [Google Scholar] [CrossRef]
- Zempel, H.; Thies, E.; Mandelkow, E.-M. A Oligomers Cause Localized Ca2+ Elevation, Missorting of Endogenous Tau into Dendrites, Tau Phosphorylation, and Destruction of Microtubules and Spines. J. Neurosci. 2010, 30, 11938–11950. [Google Scholar] [CrossRef] [PubMed]
- Bilsland, L.G.; Sahai, E.; Kelly, G.; Golding, M.; Greensmith, L.; Schiavo, G. Deficits in axonal transport precede ALS symptoms in vivo. Proc. Natl. Acad. Sci. USA 2010, 107, 20523–20528. [Google Scholar] [CrossRef]
- Li, P.; Merrill, S.A.; Jorgensen, E.M.; Shen, K. Two Clathrin Adaptor Protein Complexes Instruct Axon-Dendrite Polarity. Neuron 2016, 90, 564–580. [Google Scholar] [CrossRef]
- Margeta, M.A.; Wang, G.J.; Shen, K. Clathrin adaptor AP-1 complex excludes multiple postsynaptic receptors from axons in C. elegans. Proc. Natl. Acad. Sci. USA 2009, 106, 1632–1637. [Google Scholar] [CrossRef]
- Burack, M.A.; Silverman, M.A.; Banker, G. The Role of Selective Transport in Neuronal Protein Sorting. Neuron 2000, 26, 465–472. [Google Scholar] [CrossRef]
- Zahavi, E.E.; Hummel, J.J.; Han, Y.; Bar, C.; Stucchi, R.; Altelaar, M.; Hoogenraad, C.C. Combined kinesin-1 and kinesin-3 activity drives axonal trafficking of TrkB receptors in Rab6 carriers. Dev. Cell 2021, 56, 494–508. [Google Scholar] [CrossRef]
- Sampo, B.; Kaech, S.; Kunz, S.; Banker, G. Two Distinct Mechanisms Target Membrane Proteins to the Axonal Surface. Neuron 2003, 37, 611–624. [Google Scholar] [CrossRef]
- Garrido, J.J.; Fernandes, F.; Giraud, P.; Mouret, I.; Pasqualini, E.; Fache, M.; Jullien, F.; Dargent, B. Identification of an axonal determinant in the C-terminus of the sodium channel Nav1.2. EMBO J. 2001, 20, 5950–5961. [Google Scholar] [CrossRef]
- Nabb, A.T.; Bentley, M. NgCAM and VAMP2 Reveal that Direct Delivery and Dendritic Degradation Maintain Axonal Polarity. Mol. Biol. Cell 2022, 33, ar3. [Google Scholar] [CrossRef] [PubMed]
- Wisco, D.; Anderson, E.D.; Chang, M.C.; Norden, C.; Boiko, T.; Folsch, H.; Winckler, B. Uncovering multiple axonal targeting pathways in hippocampal neurons. J. Cell Biol. 2003, 162, 1317–1328. [Google Scholar] [CrossRef] [PubMed]
- Ascaño, M.; Richmond, A.; Borden, P.; Kuruvilla, R. Axonal Targeting of Trk Receptors via Transcytosis Regulates Sensitivity to Neurotrophin Responses. J. Neurosci. 2009, 29, 11674–11685. [Google Scholar] [CrossRef] [PubMed]
- Farías, G.G.; Guardia, C.M.; Britt, D.J.; Guo, X.; Bonifacino, J.S. Sorting of Dendritic and Axonal Vesicles at the Pre-axonal Exclusion Zone. Cell Rep. 2015, 13, 1221–1232. [Google Scholar] [CrossRef]
- Gumy, L.F.; Katrukha, E.A.; Grigoriev, I.; Jaarsma, D.; Kapitein, L.C.; Akhmanova, A.; Hoogenraad, C.C. MAP2 Defines a Pre-axonal Filtering Zone to Regulate KIF1- versus KIF5-Dependent Cargo Transport in Sensory Neurons. Neuron 2017, 94, 347–362.e7. [Google Scholar] [CrossRef]
- Song, A.-H.; Wang, D.; Chen, G.; Li, Y.; Luo, J.; Duan, S.; Poo, M.-M. A Selective Filter for Cytoplasmic Transport at the Axon Initial Segment. Cell 2009, 136, 1148–1160. [Google Scholar] [CrossRef]
- Quistgaard, E.M.; Nissen, J.D.; Hansen, S.; Nissen, P. Mind the Gap: Molecular Architecture of the Axon Initial Segment—From Fold Prediction to a Mechanistic Model of Function? J. Mol. Biol. 2021, 433, 167176. [Google Scholar] [CrossRef]
- Leterrier, C.; Dargent, B. No Pasaran! Role of the axon initial segment in the regulation of protein transport and the maintenance of axonal identity. Semin. Cell Dev. Biol. 2013, 27, 44–51. [Google Scholar] [CrossRef]
- Hedstrom, K.L.; Ogawa, Y.; Rasband, M.N. AnkyrinG is required for maintenance of the axon initial segment and neuronal polarity. J. Cell Biol. 2008, 183, 635–640. [Google Scholar] [CrossRef]
- Franssen, E.H.P.; Zhao, R.-R.; Koseki, H.; Kanamarlapudi, V.; Hoogenraad, C.; Eva, R.; Fawcett, J. Exclusion of Integrins from CNS Axons Is Regulated by Arf6 Activation and the AIS. J. Neurosci. 2015, 35, 8359–8375. [Google Scholar] [CrossRef]
- Zhou, D.; Lambert, S.; Malen, P.L.; Carpenter, S.; Boland, L.; Bennett, V. AnkyrinG Is Required for Clustering of Voltage-gated Na Channels at Axon Initial Segments and for Normal Action Potential Firing. J. Cell Biol. 1998, 143, 1295–1304. [Google Scholar] [CrossRef] [PubMed]
- Jenkins, S.M.; Bennett, V. Ankyrin-G coordinates assembly of the spectrin-based membrane skeleton, voltage-gated sodium channels, and L1 CAMs at Purkinje neuron initial segments. J. Cell Biol. 2001, 155, 739–746. [Google Scholar] [CrossRef] [PubMed]
- Hedstrom, K.L.; Xu, X.; Ogawa, Y.; Frischknecht, R.; Seidenbecher, C.I.; Shrager, P.; Rasband, M.N. Neurofascin assembles a specialized extracellular matrix at the axon initial segment. J. Cell Biol. 2007, 178, 875–886. [Google Scholar] [CrossRef] [PubMed]
- Nakada, C.; Ritchie, K.; Oba, Y.; Nakamura, M.; Hotta, Y.; Iino, R.; Kasai, R.; Yamaguchi, K.; Fujiwara, T.; Kusumi, A. Accumulation of anchored proteins forms membrane diffusion barriers during neuronal polarization. Nat. Cell Biol. 2003, 5, 626–632. [Google Scholar] [CrossRef]
- Albrecht, D.; Winterflood, C.M.; Sadeghi, M.; Tschager, T.; Noé, F.; Ewers, H. Nanoscopic compartmentalization of membrane protein motion at the axon initial segment. J. Cell Biol. 2016, 215, 37–46. [Google Scholar] [CrossRef]
- Al-Bassam, S.; Xu, M.; Wandless, T.J.; Arnold, D.B. Differential Trafficking of Transport Vesicles Contributes to the Localization of Dendritic Proteins. Cell Rep. 2012, 2, 89–100. [Google Scholar] [CrossRef]
- Eva, R.; Crisp, S.; Marland, J.; Norman, J.C.; Kanamarlapudi, V.; Ffrench-Constant, C.; Fawcett, J. ARF6 Directs Axon Transport and Traffic of Integrins and Regulates Axon Growth in Adult DRG Neurons. J. Neurosci. 2012, 32, 10352–10364. [Google Scholar] [CrossRef]
- Eva, R.; Koseki, H.; Kanamarlapudi, V.; Fawcett, J.W. EFA6 regulates selective polarised transport and axon regeneration from the axon initial segment. J. Cell Sci. 2017, 130, 3663–3675. [Google Scholar] [CrossRef]
- Kuijpers, M.; van de Willige, D.; Freal, A.; Chazeau, A.; Franker, M.A.; Hofenk, J.; Rodrigues, R.J.C.; Kapitein, L.C.; Akhmanova, A.; Jaarsma, D.; et al. Dynein Regulator NDEL1 Controls Polarized Cargo Transport at the Axon Initial Segment. Neuron 2016, 89, 461–471. [Google Scholar] [CrossRef]
- Lewis, T.L.; Mao, T.; Svoboda, K.; Arnold, D.B. Myosin-dependent targeting of transmembrane proteins to neuronal dendrites. Nat. Neurosci. 2009, 12, 568–576. [Google Scholar] [CrossRef]
- Andrews, M.R.; Soleman, S.; Cheah, M.; Tumbarello, D.A.; Mason, M.R.J.; Moloney, E.; Verhaagen, J.; Bensadoun, J.-C.; Schneider, B.; Aebischer, P.; et al. Axonal Localization of Integrins in the CNS Is Neuronal Type and Age Dependent. eNeuro 2016, 3, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Eom, T.; Oleynikov, Y.; Shenoy, S.; Liebelt, D.; Dictenberg, J.; Singer, R.; Bassell, G. Neurotrophin-Induced Transport of a β-Actin mRNP Complex Increases β-Actin Levels and Stimulates Growth Cone Motility. Neuron 2001, 31, 261–275. [Google Scholar] [CrossRef]
- Aronov, S.; Aranda, G.; Behar, L.; Ginzburg, I. Axonal Tau mRNA Localization Coincides with Tau Protein in Living Neuronal Cells and Depends on Axonal Targeting Signal. J. Neurosci. 2001, 21, 6577–6587. [Google Scholar] [CrossRef] [PubMed]
- Turner-Bridger, B.; Caterino, C.; Cioni, J.-M. Molecular mechanisms behind mRNA localization in axons. Open Biol. 2020, 10, 200177. [Google Scholar] [CrossRef]
- Padmanabhan, S.; Kareva, T.; Kholodilov, N.; E Burke, R. Quantitative morphological comparison of axon-targeting strategies for gene therapies directed to the nigro-striatal projection. Gene Ther. 2013, 21, 115–122. [Google Scholar] [CrossRef]
- Babetto, E.; Beirowski, B.; Janeckova, L.; Brown, R.; Gilley, J.; Thomson, D.; Ribchester, R.R.; Coleman, M.P. Targeting NMNAT1 to Axons and Synapses Transforms Its Neuroprotective Potency In Vivo. J. Neurosci. 2010, 30, 13291–13304. [Google Scholar] [CrossRef]
- Satpute-Krishnan, P.; DeGiorgis, J.A.; Conley, M.P.; Jang, M.; Bearer, E.L. A peptide zipcode sufficient for anterograde transport within amyloid precursor protein. Proc. Natl. Acad. Sci. USA 2006, 103, 16532–16537. [Google Scholar] [CrossRef]
- El-Husseini, A.E.-D.; Craven, S.E.; Brock, S.C.; Bredt, D.S. Polarized Targeting of Peripheral Membrane Proteins in Neurons. J. Biol. Chem. 2001, 276, 44984–44992. [Google Scholar] [CrossRef]
- Zuber, M.X.; Strittmatter, S.M.; Fishman, M.C. A membrane-targeting signal in the amino terminus of the neuronal protein GAP-43. Nature 1989, 341, 345–348. [Google Scholar] [CrossRef]
- Xu, M.; Cao, R.; Xiao, R.; Zhu, M.X.; Gu, C. The Axon Dendrite Targeting of Kv3 (Shaw) Channels Is Determined by a Targeting Motif That Associates with the T1 Domain and Ankyrin G. J. Neurosci. 2007, 27, 14158–14170. [Google Scholar] [CrossRef]
- Gasser, A.; Ho, T.S.-Y.; Cheng, X.; Chang, K.-J.; Waxman, S.G.; Rasband, M.N.; Dib-Hajj, S.D. An AnkyrinG-Binding Motif Is Necessary and Sufficient for Targeting Nav1.6 Sodium Channels to Axon Initial Segments and Nodes of Ranvier. J. Neurosci. 2012, 32, 7232–7243. [Google Scholar] [CrossRef] [PubMed]
- Lemaillet, G.; Walker, B.; Lambert, S. Identification of a Conserved Ankyrin-binding Motif in the Family of Sodium Channel α Subunits. J. Biol. Chem. 2003, 278, 27333–27339. [Google Scholar] [CrossRef] [PubMed]
- Lewis, T.L., Jr.; Mao, T.; Arnold, D.B. A Role for Myosin VI in the Localization of Axonal Proteins. PLoS Biol. 2011, 9, e1001021. [Google Scholar] [CrossRef]
- Fairless, R.; Masius, H.; Rohlmann, A.; Heupel, K.; Ahmad, M.; Reissner, C.; Dresbach, T.; Missler, M. Polarized Targeting of Neurexins to Synapses Is Regulated by their C-Terminal Sequences. J. Neurosci. 2008, 28, 12969–12981. [Google Scholar] [CrossRef]
- Xu, J.; Zhu, Y.; Heinemann, S.F. Identification of Sequence Motifs That Target Neuronal Nicotinic Receptors to Dendrites and Axons. J. Neurosci. 2006, 26, 9780–9793. [Google Scholar] [CrossRef]
- Bel, C.; Oguievetskaia, K.; Pitaval, C.; Goutebroze, L.; Faivre-Sarrailh, C. Axonal targeting of Caspr2 in hippocampal neurons via selective somatodendritic endocytosis. J. Cell Sci. 2009, 122, 3403–3413. [Google Scholar] [CrossRef]
- Gauthier-Kemper, A.; Igaev, M.; Sündermann, F.; Janning, D.; Brühmann, J.; Moschner, K.; Reyher, H.-J.; Junge, W.; Glebov, K.; Walter, J.; et al. Interplay between phosphorylation and palmitoylation mediates plasma membrane targeting and sorting of GAP43. Mol. Biol. Cell 2014, 25, 3284–3299. [Google Scholar] [CrossRef]
- Tortosa, E.; Adolfs, Y.; Fukata, M.; Pasterkamp, R.J.; Kapitein, L.C.; Hoogenraad, C.C. Dynamic Palmitoylation Targets MAP6 to the Axon to Promote Microtubule Stabilization during Neuronal Polarization. Neuron 2017, 94, 809–825.e7. [Google Scholar] [CrossRef]
- Broussard, G.; Liang, Y.; Fridman, M.; Unger, E.; Meng, G.; Xiao, X.; Ji, N.; Petreanu, L.; Tian, L. In vivo measurement of afferent activity with axon-specific calcium imaging. Nat. Neurosci. 2018, 21, 1272–1280. [Google Scholar] [CrossRef]
- Inomata, H.; Nakamura, Y.; Hayakawa, A.; Takata, H.; Suzuki, T.; Miyazawa, K.; Kitamura, N. A Scaffold Protein JIP-1b Enhances Amyloid Precursor Protein Phosphorylation by JNK and Its Association with Kinesin Light Chain 1. J. Biol. Chem. 2003, 278, 22946–22955. [Google Scholar] [CrossRef]
- Barry, J.; Gu, Y.; Jukkola, P.; O’Neill, B.; Gu, H.; Mohler, P.J.; Rajamani, K.T.; Gu, C. Ankyrin-G Directly Binds to Kinesin-1 to Transport Voltage-Gated Na+ Channels into Axons. Dev. Cell 2014, 28, 117–131. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Shui, B.; Kotlikoff, M.I.; Sondermann, H. Structural Basis for Calcium Sensing by GCaMP2. Structure 2008, 16, 1817–1827. [Google Scholar] [CrossRef] [PubMed]
- Broussard, G.; Liang, R.; Etian, L. Monitoring activity in neural circuits with genetically encoded indicators. Front. Mol. Neurosci. 2014, 7, 97. [Google Scholar] [CrossRef] [PubMed]
- Dreosti, E.; Odermatt, B.; Dorostkar, M.; Lagnado, L. A genetically encoded reporter of synaptic activity in vivo. Nat. Methods 2009, 6, 883–889. [Google Scholar] [CrossRef]
- Waller, A. Experiments on the Section of the Glosso-Pharyngeal and Hypoglossal Nerves of the Frog, and Observations of the Alterations Produced Thereby in the Structure of Their Primitive Fibres. Edinb. Med Surg. J. 1851, 76, 369–376. [Google Scholar] [PubMed]
- Perry, V.H.; Brown, M.C.; Lunn, E.R. Very Slow Retrograde and Wallerian Degeneration in the CNS of C57BL/Ola Mice. Eur. J. Neurosci. 1991, 3, 102–105. [Google Scholar] [CrossRef]
- Mack, T.G.A.; Reiner, M.; Beirowski, B.; Mi, W.; Emanuelli, M.; Wagner, D.; Thomson, D.; Gillingwater, T.; Court, F.; Conforti, L.; et al. Wallerian degeneration of injured axons and synapses is delayed by a Ube4b/Nmnat chimeric gene. Nat. Neurosci. 2001, 4, 1199–1206. [Google Scholar] [CrossRef]
- Beirowski, B.; Babetto, E.; Gilley, J.; Mazzola, F.; Conforti, L.; Janeckova, L.; Magni, G.; Ribchester, R.R.; Coleman, M.P. Non-Nuclear WldS Determines Its Neuroprotective Efficacy for Axons and Synapses In Vivo. J. Neurosci. 2009, 29, 653–668. [Google Scholar] [CrossRef]
- Yamagata, T.; Saito, H.; Habuchi, O.; Suzuki, S. Purification and Properties of Bacterial Chondroitinases and Chondrosulfatases. J. Biol. Chem. 1968, 243, 1523–1535. [Google Scholar] [CrossRef]
- Hussein, R.K.; Mencio, C.P.; Katagiri, Y.; Brake, A.M.; Geller, H.M. Role of Chondroitin Sulfation Following Spinal Cord Injury. Front. Cell. Neurosci. 2020, 14, 208. [Google Scholar] [CrossRef]
- Bradbury, E.J.; Moon, L.; Popat, R.J.; King, V.R.; Bennett, G.S.; Patel, P.N.; Fawcett, J.; McMahon, S. Chondroitinase ABC promotes functional recovery after spinal cord injury. Nature 2002, 416, 636–640. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.; McKeon, R.J.; Bellamkonda, R.V. Sustained delivery of thermostabilized chABC enhances axonal sprouting and functional recovery after spinal cord injury. Proc. Natl. Acad. Sci. USA 2009, 107, 3340–3345. [Google Scholar] [CrossRef] [PubMed]
- Burnside, E.R.; De Winter, F.; Didangelos, A.; James, N.D.; Andreica, E.-C.; Horsfall, H.L.; Muir, E.M.; Verhaagen, J.; Bradbury, E. Immune-evasive gene switch enables regulated delivery of chondroitinase after spinal cord injury. Brain 2018, 141, 2362–2381. [Google Scholar] [CrossRef] [PubMed]
- Muir, E.M.; Fyfe, I.; Gardiner, S.; Li, L.; Warren, P.; Fawcett, J.W.; Keynes, R.J.; Rogers, J.H. Modification of N-glycosylation sites allows secretion of bacterial chondroitinase ABC from mammalian cells. J. Biotechnol. 2010, 145, 103–110. [Google Scholar] [CrossRef] [PubMed]
- Day, P.; Alves, N.; Daniell, E.; Dasgupta, D.; Ogborne, R.; Steeper, A.; Raza, M.; Ellis, C.; Fawcett, J.; Keynes, R.; et al. Targeting chondroitinase ABC to axons enhances the ability of chondroitinase to promote neurite outgrowth and sprouting. PLoS ONE 2020, 15, e0221851. [Google Scholar] [CrossRef] [PubMed]
- Hollis, E.R.; Lu, P.; Blesch, A.; Tuszynski, M.H. IGF-I gene delivery promotes corticospinal neuronal survival but not regeneration after adult CNS injury. Exp. Neurol. 2009, 215, 53–59. [Google Scholar] [CrossRef][Green Version]
- Hollis, E.; Jamshidi, P.; Löw, K.; Blesch, A.; Tuszynski, M.H. Induction of corticospinal regeneration by lentiviral trkB-induced Erk activation. Proc. Natl. Acad. Sci. USA 2009, 106, 7215–7220. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Protein of Origin | Region | Peptide | Putative Mechanism of Targeting | Model | Source |
|---|---|---|---|---|---|
| Amyloid precursor protein (APP) | C-terminus | GYENPTYKFFEQMQN | Promotes interaction with KLC1 and NPTY motifs recruits JIP-1b which interacts with KLC1 to promote association with kinesin-1. | Giant squid axon, primary E18 rat hippocampal neurons, and primary E14-16 murine dorsal root ganglion neurons | [35,36,37] |
| Paralemmin | C-terminus | DMKKHRCKCCSIM | Dicysteine palmitoylation motif with nearby basic amino acids sufficient for targeting to secretory pathway, likely through association with lipid rafts in trans-Golgi network. | Primary E18 rat hippocampal neurons | [38] |
| Growth associated protein-43 (GAP-43) | N-terminus | MLCCMRRTKQV | [35,38,39] | ||
| Kv3.1 | C-terminus | MAKQKLPKKKKHIPRRP | Interacts with T1 tetramerisation domain and Ankyrin-G binding motif. | Primary E18 rat hippocampal neurons | [40] |
| Nav1.2 | C-terminus | CLDILFAFT | Stimulates clathrin-dependent somatodendritic endocytosis. | Primary E18 rat hippocampal neurons | [10] |
| Voltage gated sodium channel α subunits | Intracellular loop II-III | (V/A)P(I/L)AxxE(S/D)D | Ankyrin-G binding motif. | Primary dorsal root ganglion neuron-Schwann cell myelinating coculture | [41,42] |
| Optineurin (OPTN) | Myosin VI-binding domains | OPTN AAs 420-526 | Association with actin-based minus-end directed myosin VI stimulates somatodendritic endocytosis. | Primary E18 rat cortical neurons | [43] |
| Disabled homologue 2 (DAB2) | DAB2 AAs 649-719 | ||||
| Neurexin-1α (Nxn1α) | C-terminus | Nxn-1α AAs 1420-1477 | PDZ recognition motif is required for Golgi exit and sorting into secretory vesicles, preferential exocytosis onto axon membrane. | Primary P0 murine hippocampal neurons | [44] |
| Acetylcholine receptor α4 subunit | M3-M4 loop | [D/E]xxxL[L/I] | AP-2 and -3 binding motif stimulates somatodendritic endocytosis. | Primary P0 rat hippocampal neurons | [45] |
| Contactin-associated protein-like 2 (Caspr2) | 4.1 binding domain | RYMFRHKGT | Protein kinase C phosphorylation of [R/K]X[pS/pT] motif increases somatodendritic endocytosis. | Primary E18 rat hippocampal neurons | [46] |
| SifA and kinesin-interacting protein (SKIP) | Kinesin light chain binding sequence | TNLEWDDSAI | KLC1 binding motif promotes association with kinesin-1. | Primary E18 rat hippocampal neurons | [14] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Steele-Nicholson, L.J.; Andrews, M.R. Axon-Targeting Motifs: Mechanisms and Applications of Enhancing Axonal Localisation of Transmembrane Proteins. Cells 2022, 11, 937. https://doi.org/10.3390/cells11060937
Steele-Nicholson LJ, Andrews MR. Axon-Targeting Motifs: Mechanisms and Applications of Enhancing Axonal Localisation of Transmembrane Proteins. Cells. 2022; 11(6):937. https://doi.org/10.3390/cells11060937
Chicago/Turabian StyleSteele-Nicholson, Lloyd J., and Melissa R. Andrews. 2022. "Axon-Targeting Motifs: Mechanisms and Applications of Enhancing Axonal Localisation of Transmembrane Proteins" Cells 11, no. 6: 937. https://doi.org/10.3390/cells11060937
APA StyleSteele-Nicholson, L. J., & Andrews, M. R. (2022). Axon-Targeting Motifs: Mechanisms and Applications of Enhancing Axonal Localisation of Transmembrane Proteins. Cells, 11(6), 937. https://doi.org/10.3390/cells11060937