
Reinfection of Transplanted Livers in HCV- and HCV/HIV-Infected Patients Is Characterized by a Different MicroRNA Expression Profile

 ,
,  , ,
, , 

Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. RNA Extraction and Library Preparation and Sequencing
2.3. smallRNA Bioinformatics Analysis
2.4. miRNAs Functional Enrichment Analysis
3. Results
3.1. Gene Expression Analyses Highlighted Specific miRNAs Differentially Expressed between Healthy Livers and HCV- as Well as HCV/HIV-Infected Biopsies
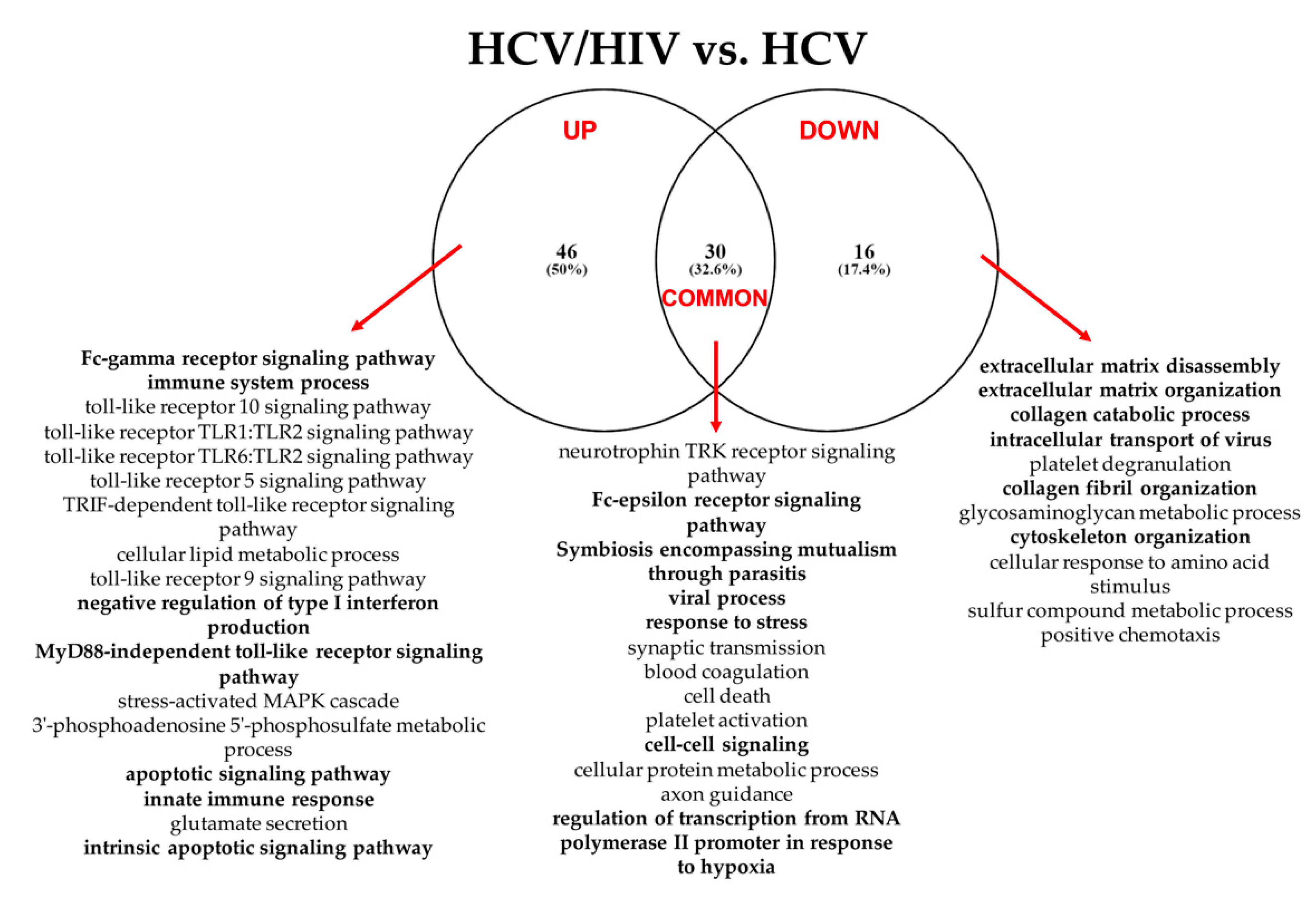
3.2. Comparative Functional Pathway Analyses of the Differentially Expressed miRNAs
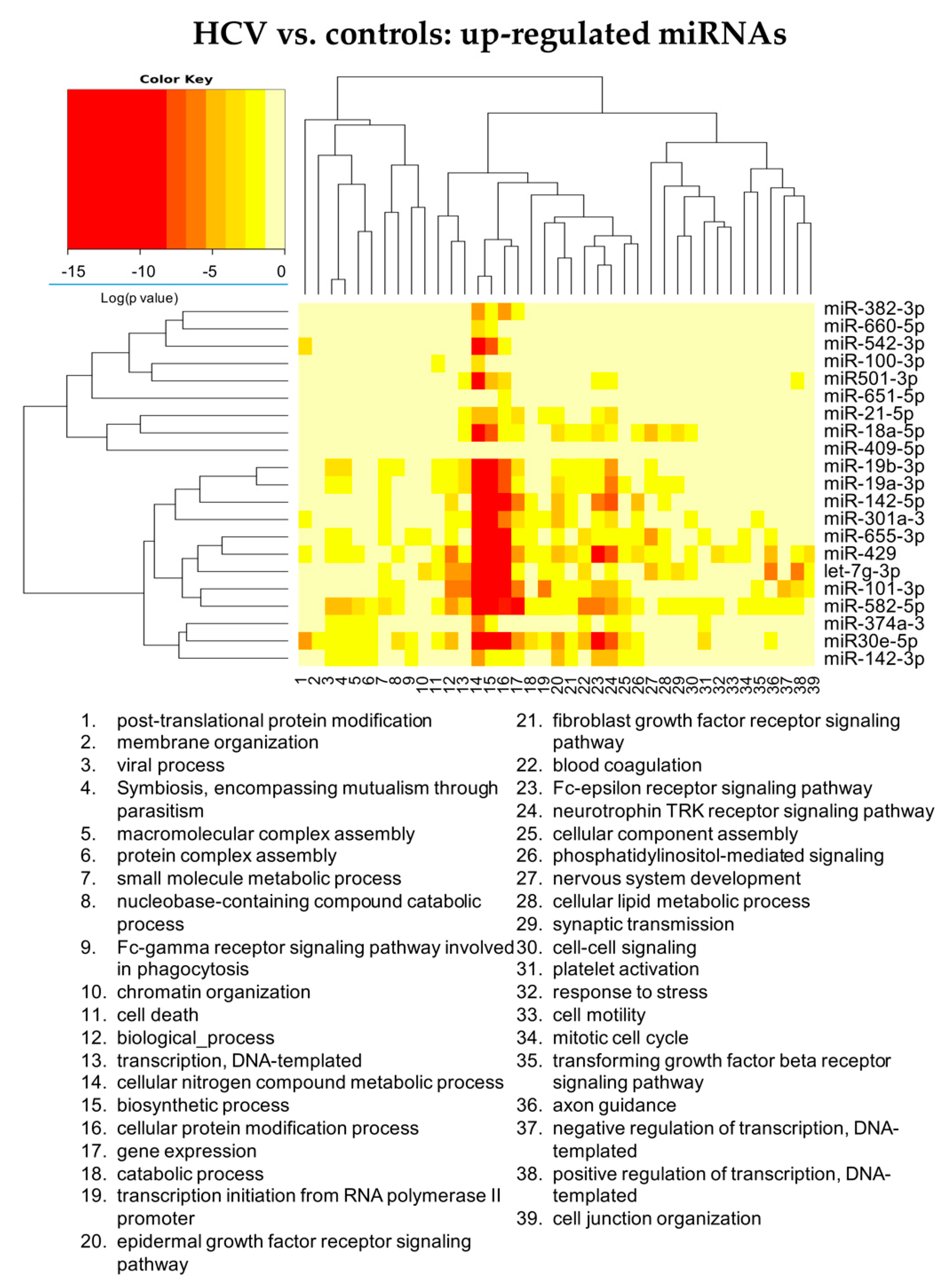
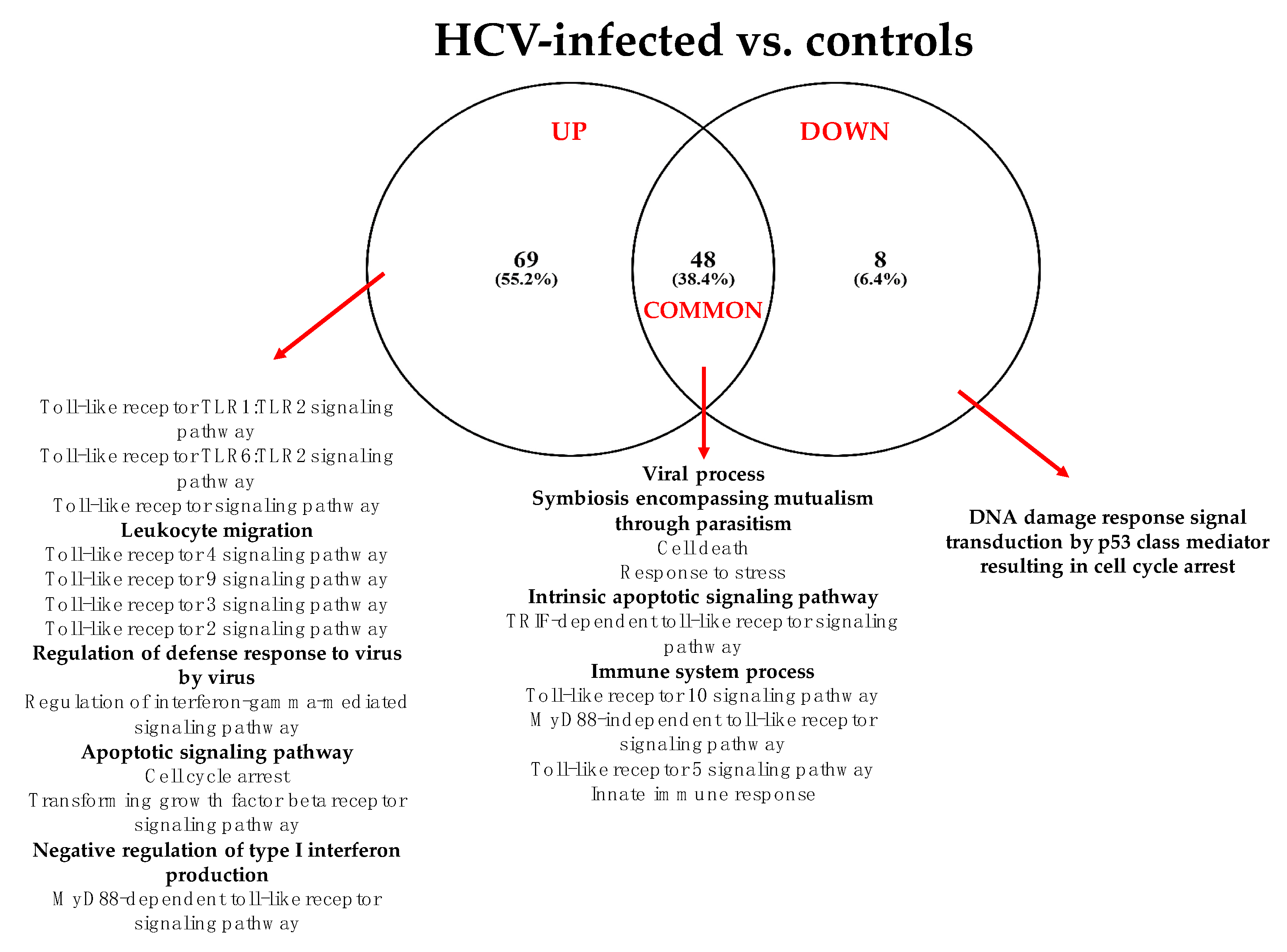
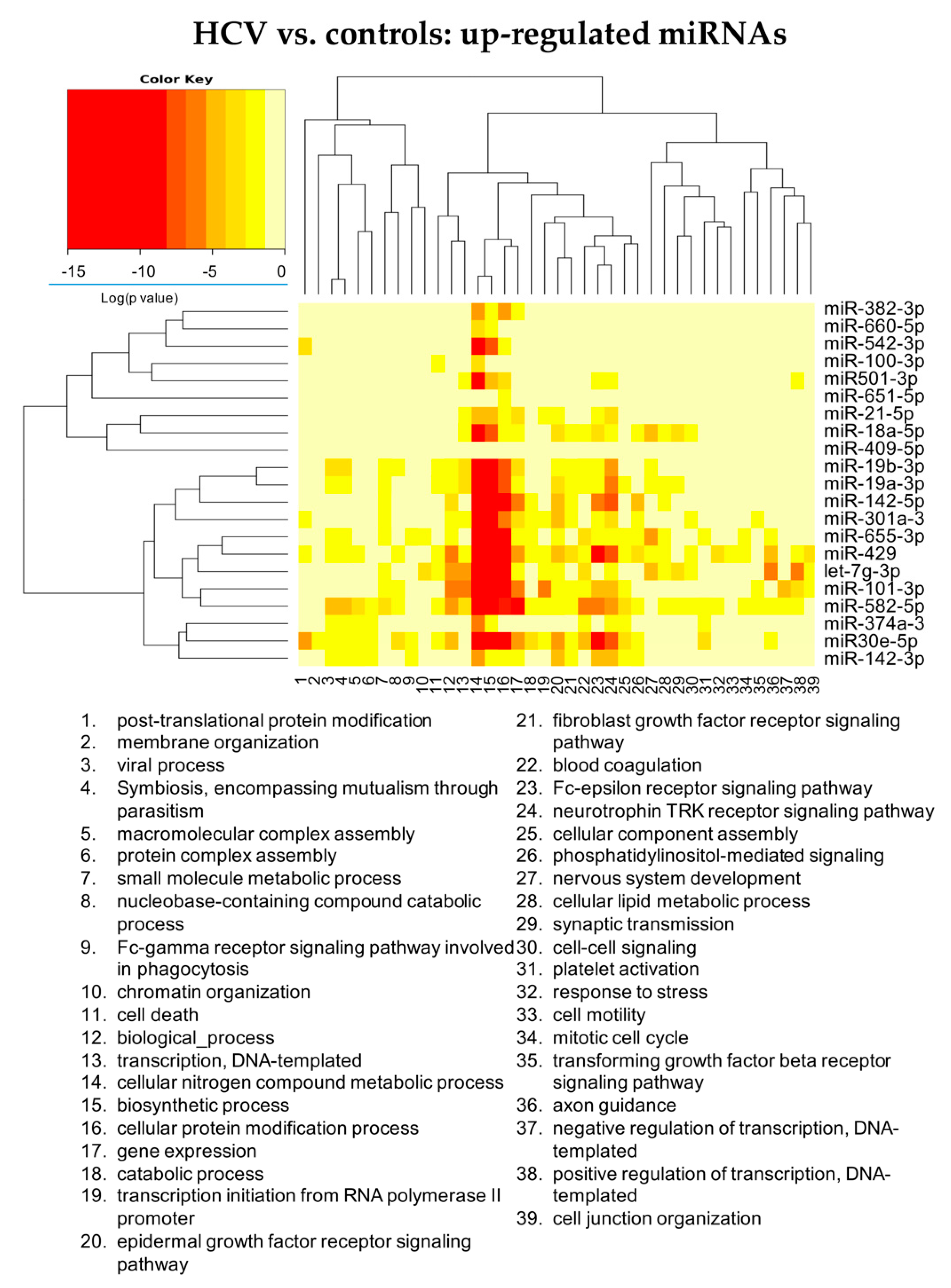
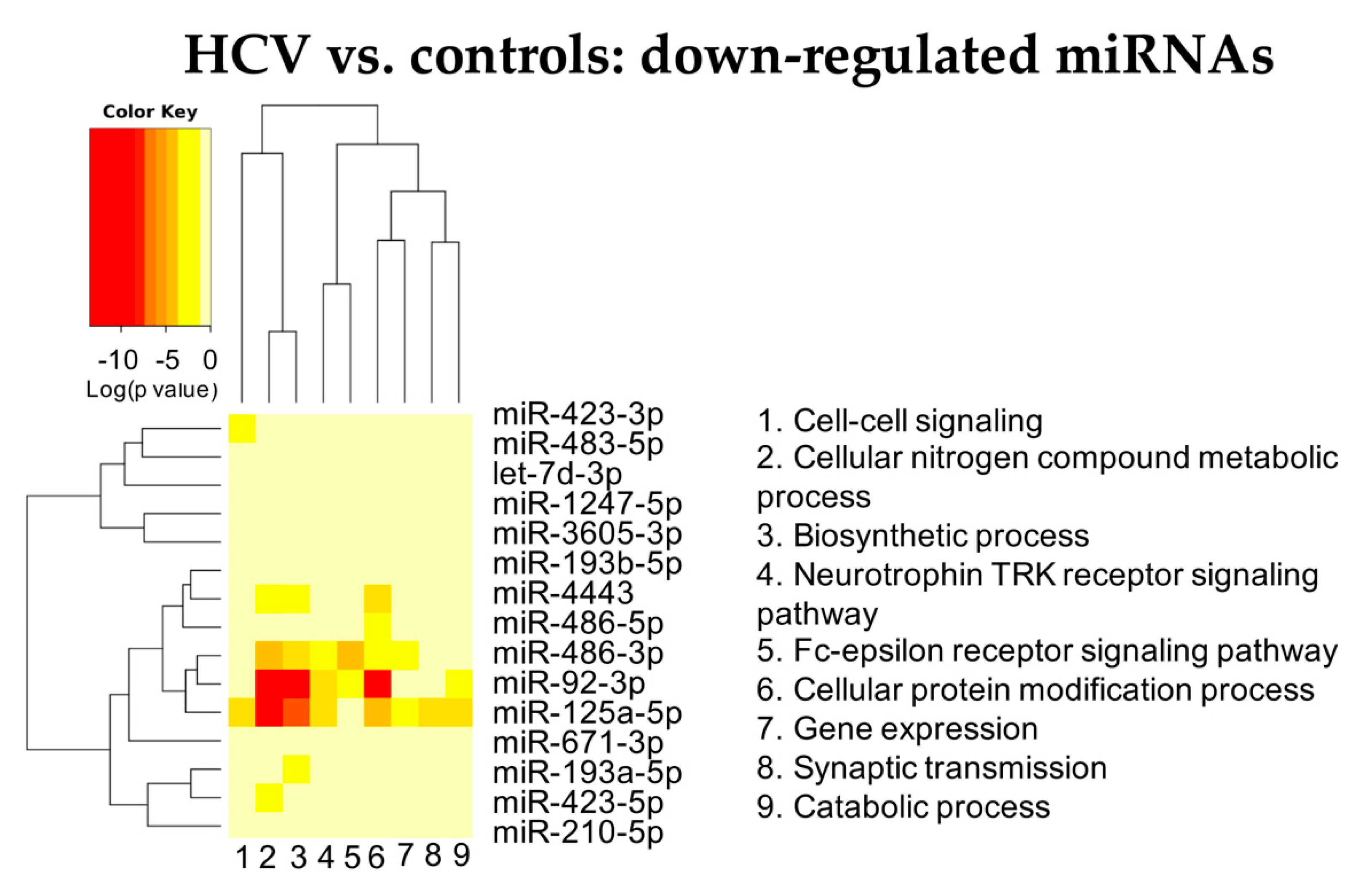
3.2.1. HCV vs. Controls
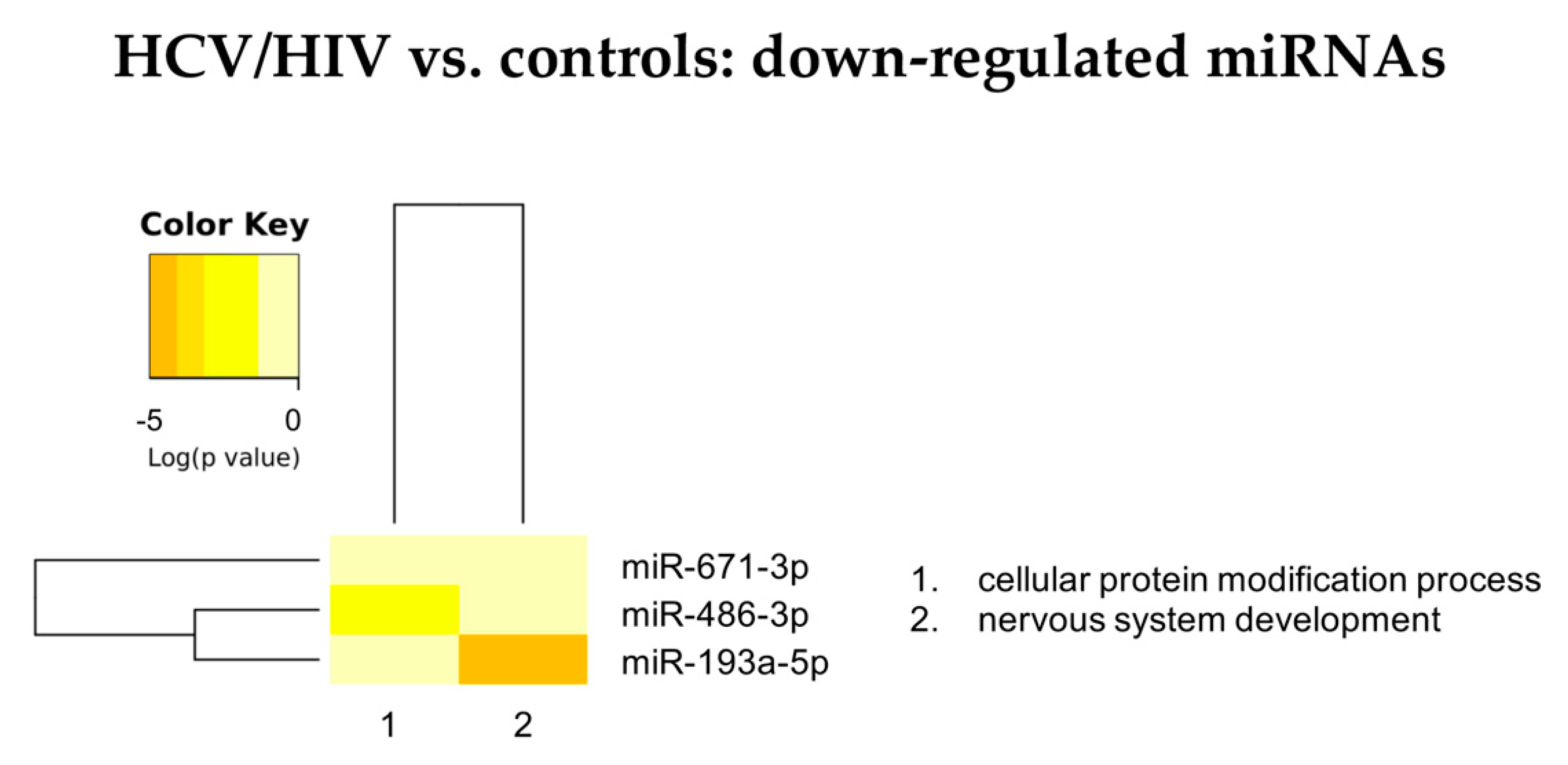
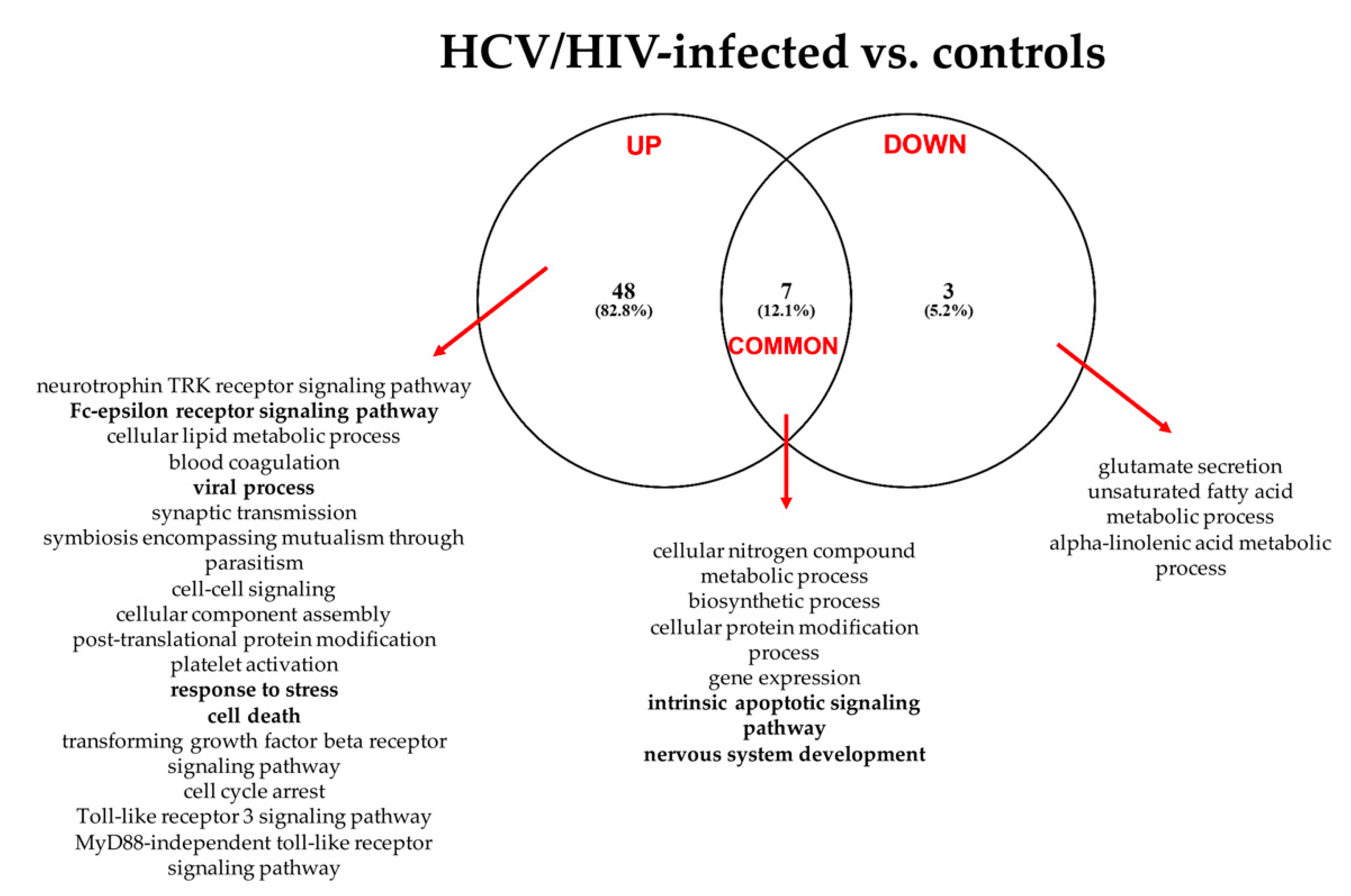
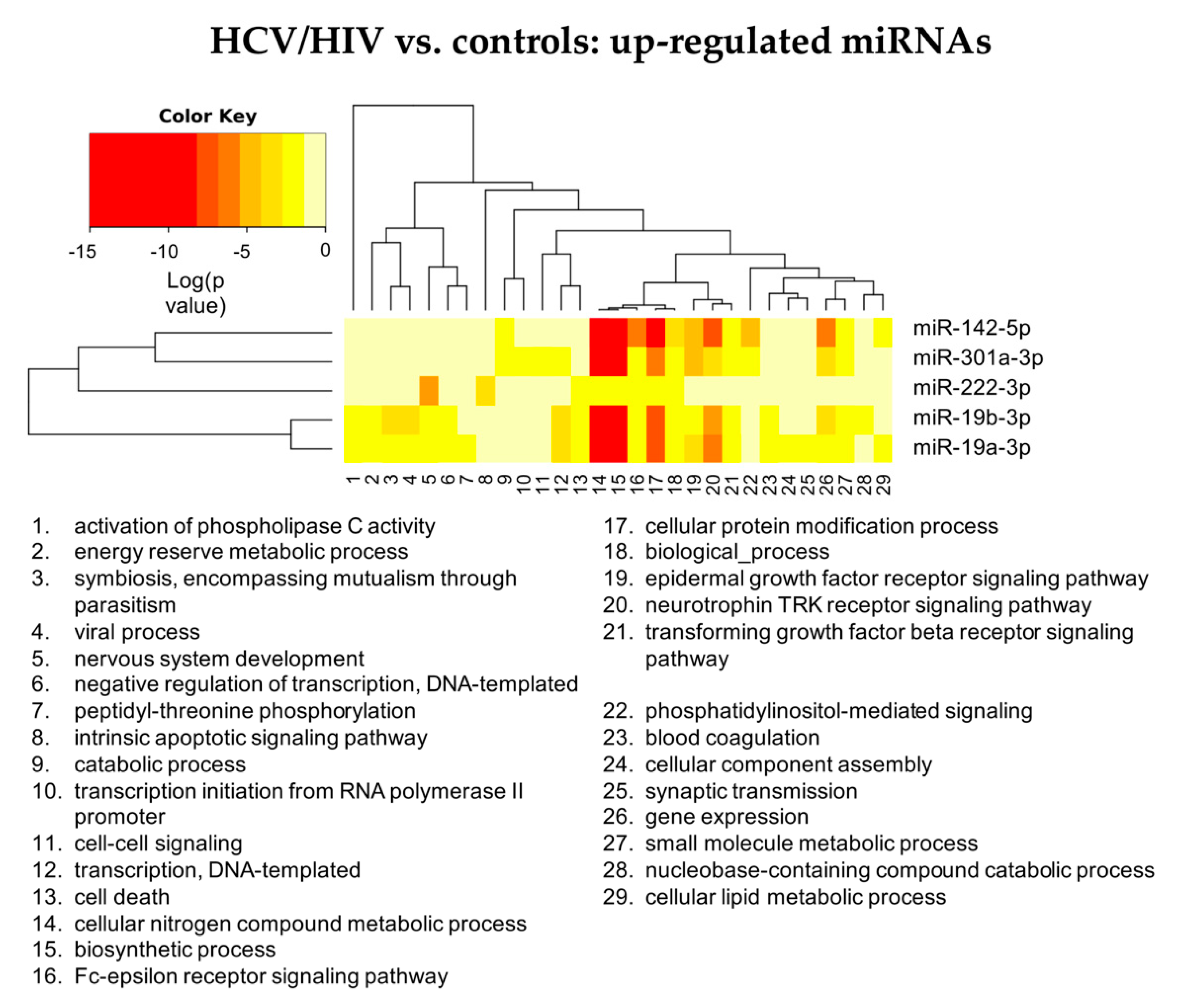
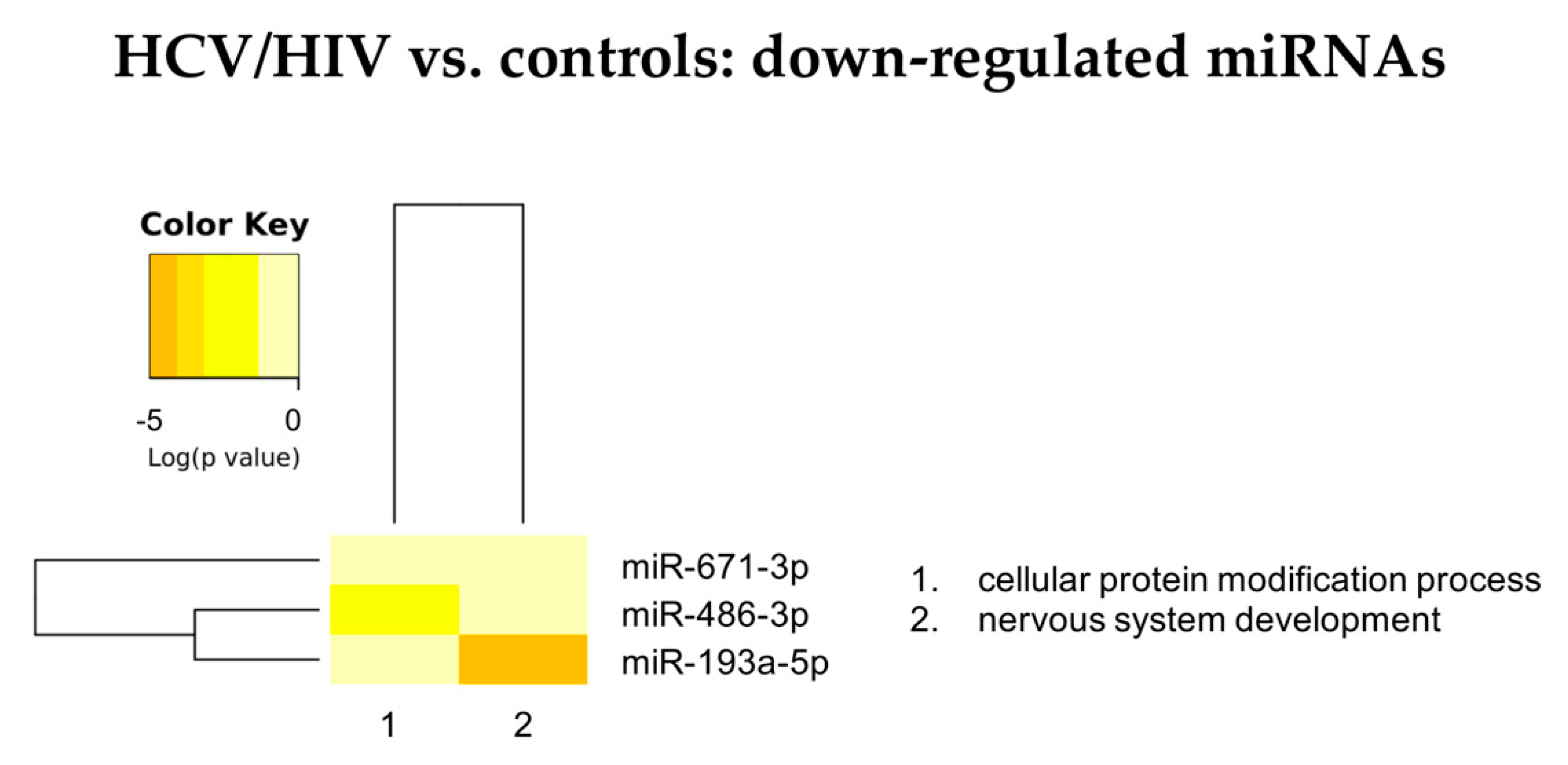
3.2.2. HCV/HIV vs. Controls
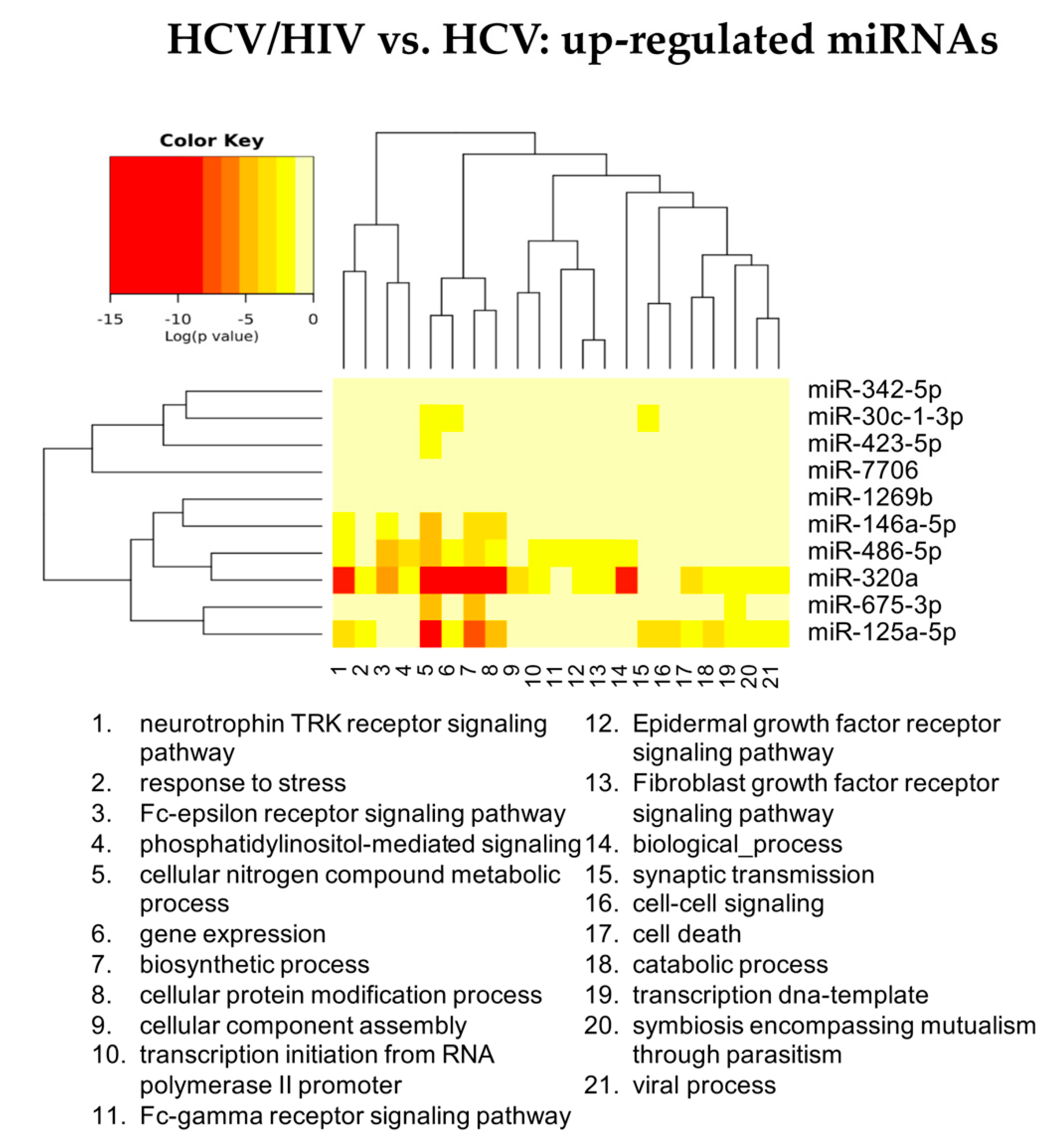
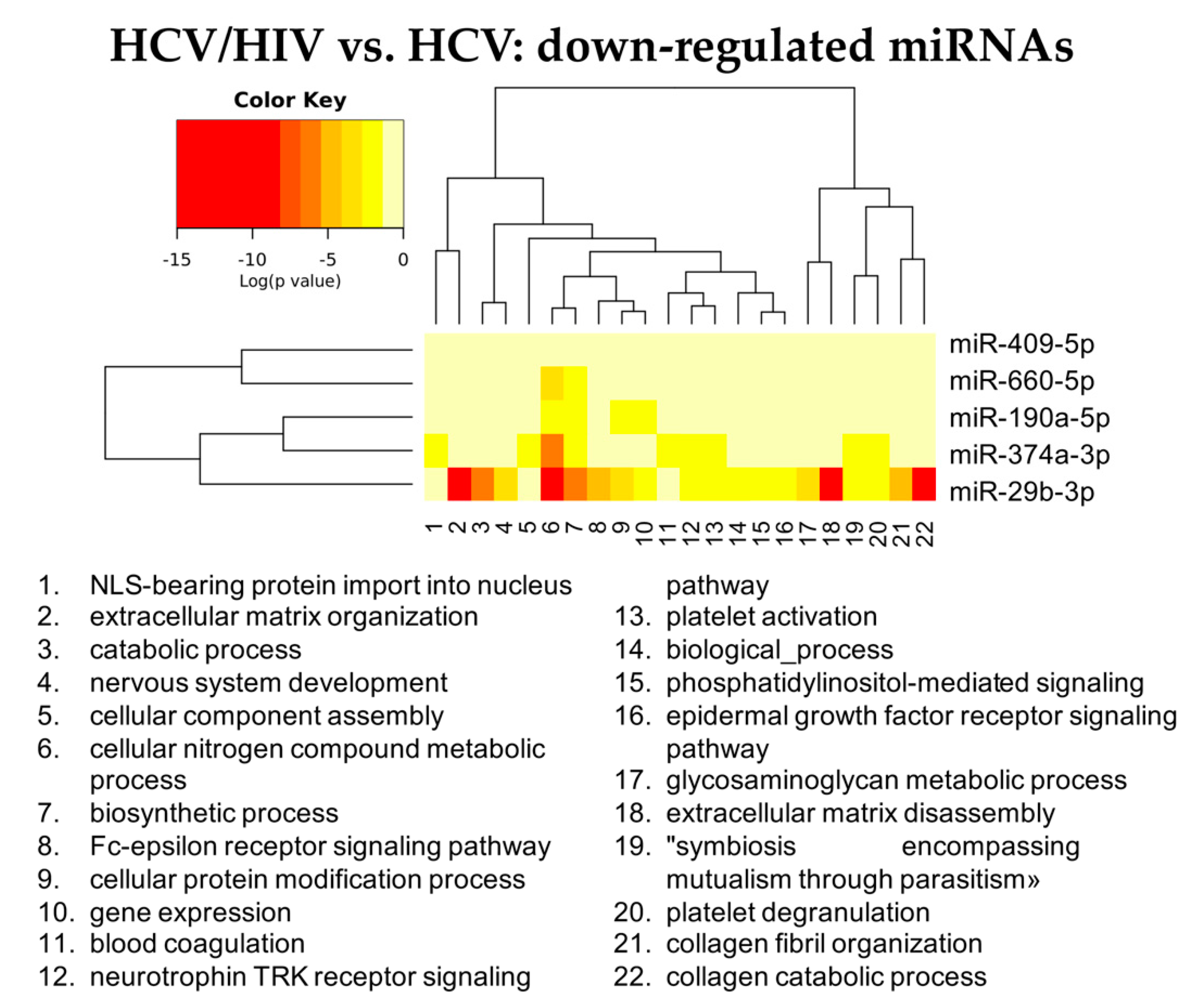
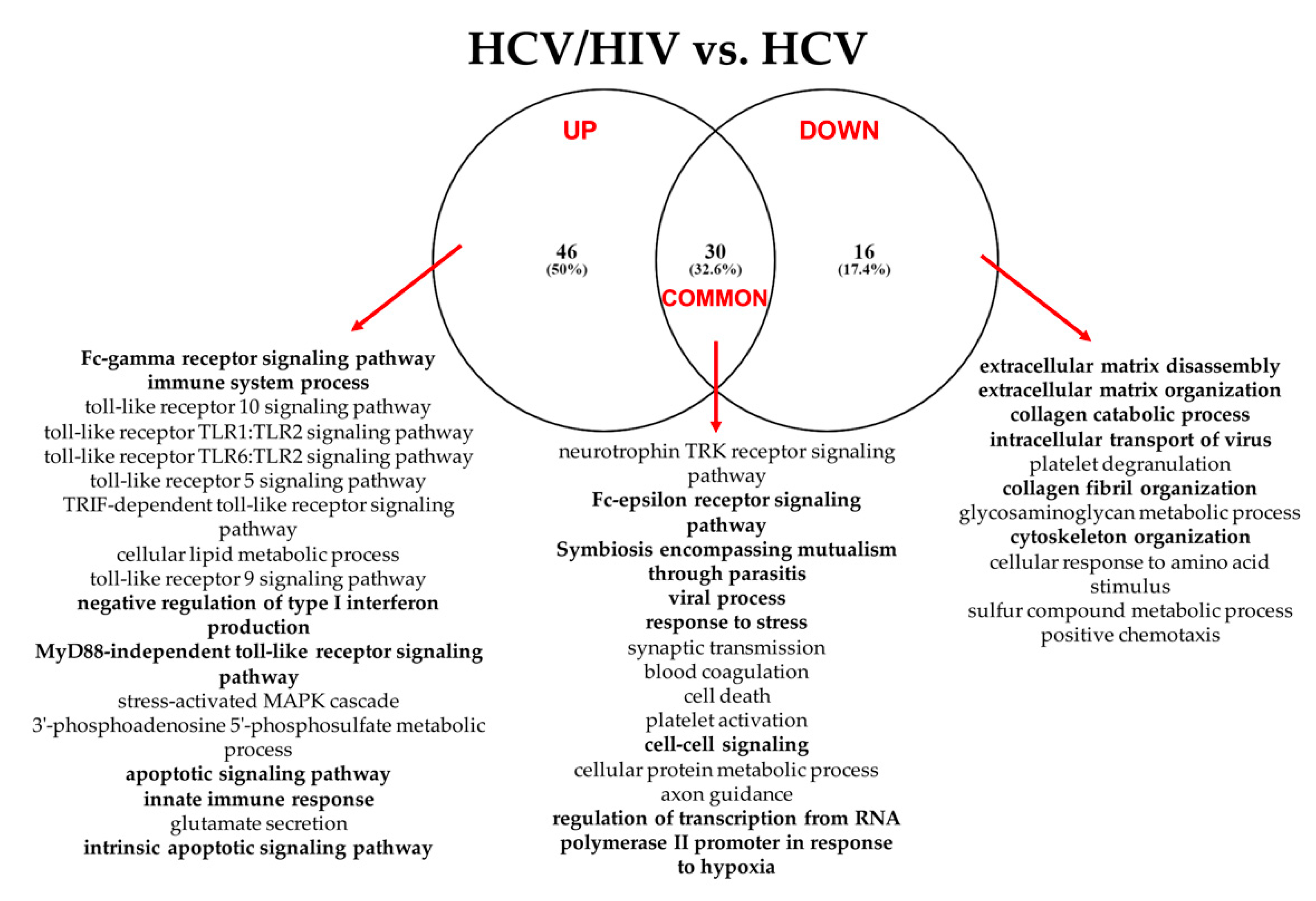
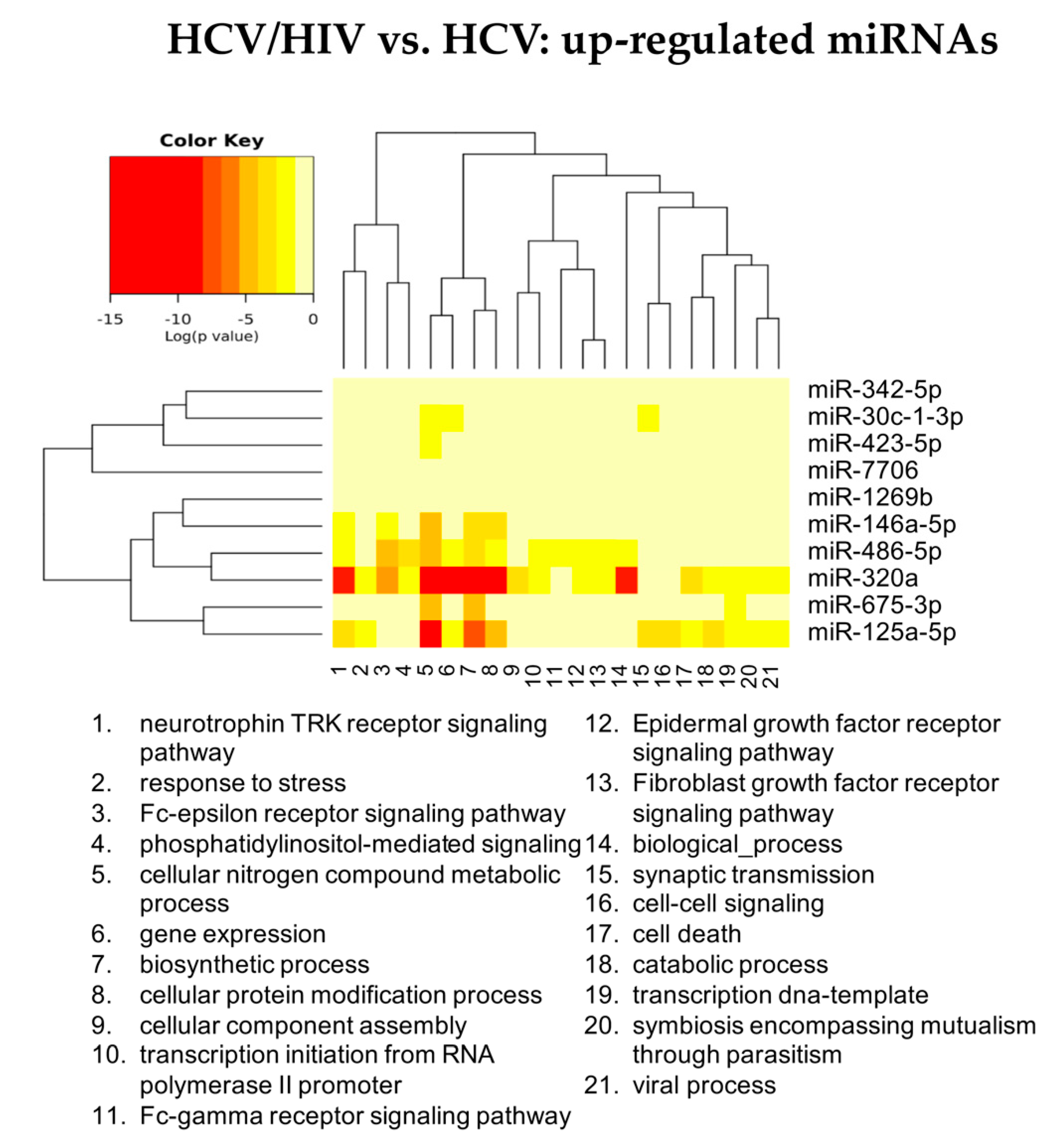
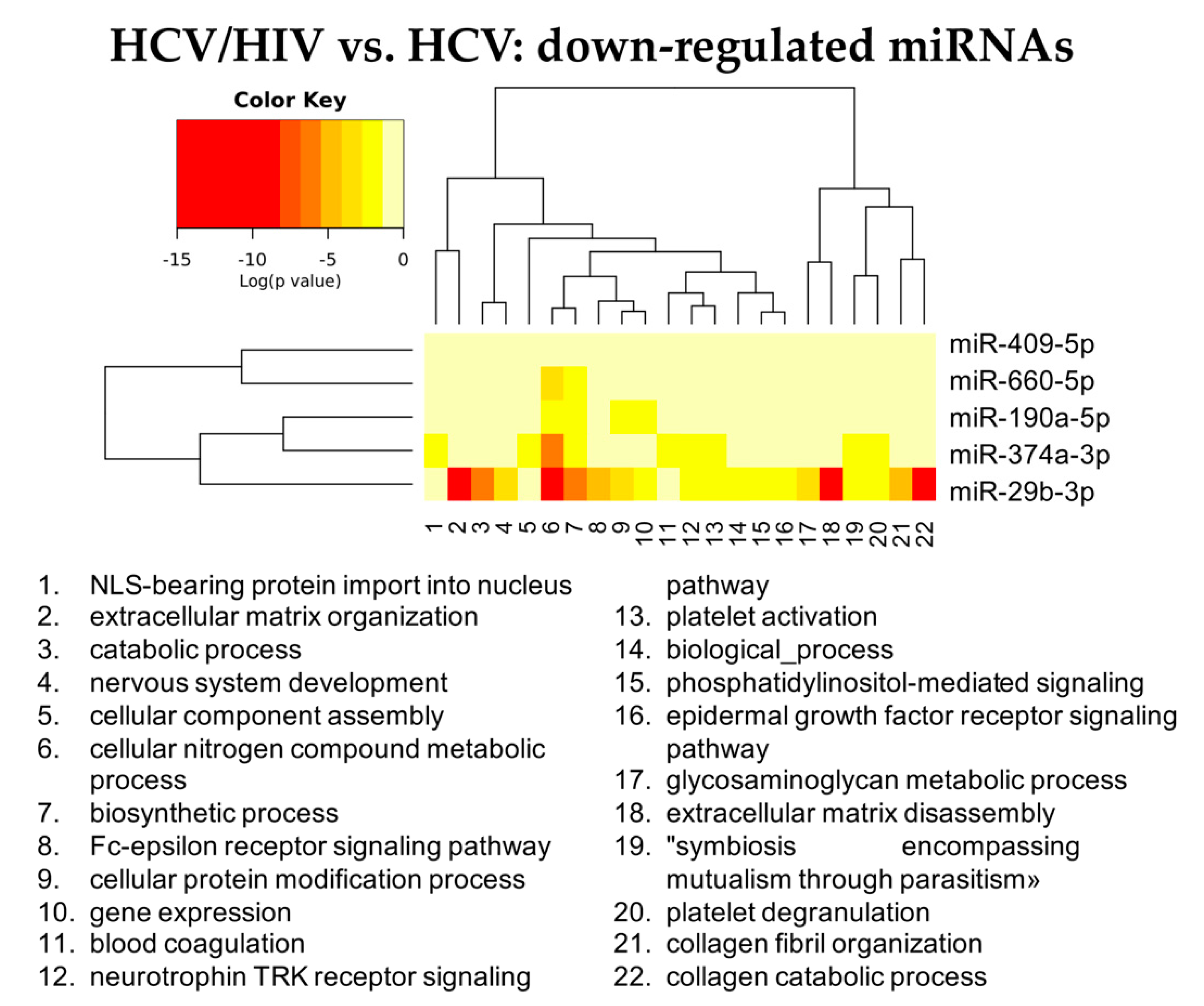
3.2.3. HCV/HIV Co-Infected vs. HCV Mono-Infected
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Baccarani, U.; Pravisani, R.; Isola, M.; Mocchegiani, F.; Lauterio, A.; Righi, E.; Magistri, P.; Corno, V.; Adani, G.L.; Lorenzin, D.; et al. Early post-liver transplant surgical morbidity in HIV-infected recipients: Risk factor for overall survival? A nationwide retrospective study. Transpl. Int. 2019, 32, 1044–1052. [Google Scholar] [CrossRef]
- Bulfoni, M.; Pravisani, R.; Dalla, E.; Cesselli, D.; Hidaka, M.; Di Loreto, C.; Eguchi, S.; Baccarani, U. miRNA expression profiles in liver grafts of HCV and HIV/HCV-infected recipients, 6 months after liver transplantation. J. Med. Virol. 2021, 93, 4992–5000. [Google Scholar] [CrossRef]
- Smith, C.J.; Ryom, L.; Weber, R.; Morlat, P.; Pradier, C.; Reiss, P.; Kowalska, J.D.; de Wit, S.; Law, M.; el Sadr, W.; et al. Trends in underlying causes of death in people with HIV from 1999 to 2011 (D:A:D): A multicohort collaboration. Lancet 2014, 384, 241–248. [Google Scholar] [CrossRef] [Green Version]
- Gobran, S.T.; Ancuta, P.; Shoukry, N.H. A Tale of Two Viruses: Immunological Insights Into HCV/HIV Coinfection. Front. Immunol. 2021, 12, 726419. [Google Scholar] [CrossRef] [PubMed]
- Grassi, A.; Ballardini, G. Post-liver transplant hepatitis C virus recurrence: An unresolved thorny problem. World J. Gastroenterol. 2014, 20, 11095–11115. [Google Scholar] [CrossRef] [PubMed]
- Naggie, S. Hepatitis C Virus, Inflammation, and Cellular Aging: Turning Back Time. Top. Antivir. Med. 2017, 25, 3–6. [Google Scholar] [PubMed]
- Medrano, L.M.; Berenguer, J.; Salgüero, S.; González-García, J.; Díez, C.; Hontañón, V.; Garcia-Broncano, P.; Ibañez-Samaniego, L.; Bellón, J.M.; Jiménez-Sousa, M.A.; et al. Successful HCV Therapy Reduces Liver Disease Severity and Inflammation Biomarkers in HIV/HCV-Coinfected Patients With Advanced Cirrhosis: A Cohort Study. Front. Med. 2021, 8, 17. [Google Scholar] [CrossRef] [PubMed]
- Cotter, T.G.; Wang, J.; Lieber, S.R.; Odenwald, M.A.; Rich, N.E.; Marrero, J.A.; Singal, A.G.; Mitchell, M.C.; Aronsohn, A.; Charlton, M.; et al. “Raising HOPE”: Improved Outcomes for HIV/HCV-coinfected Liver Transplant Recipients in the Direct-acting Antiviral Era. Transplant. Direct 2021, 7, e707. [Google Scholar] [CrossRef] [PubMed]
- Grottenthaler, J.M.; Werner, C.R.; Steurer, M.; Spengler, U.; Berg, T.; Engelmann, C.; Wedemeyer, H.; von Hahn, T.; Stremmel, W.; Pathil, A.; et al. Successful direct acting antiviral (DAA) treatment of HCV/HIV-coinfected patients before and after liver transplantation. PLoS ONE 2018, 13, e0197544. [Google Scholar] [CrossRef]
- Locke, J.E.; Durand, C.; Reed, R.D.; MacLennan, P.A.; Mehta, S.; Massie, A.; Nellore, A.; DuBay, D.; Segev, D.L. Long-term Outcomes after Liver Transplantation among Human Immunodeficiency Virus-Infected Recipients. Transplantation 2016, 100, 141–146. [Google Scholar] [CrossRef]
- Brochado-Kith, O.; Martinez, I.; Berenguer, J.; Gonzalez-Garcia, J.; Salguero, S.; Sepulveda-Crespo, D.; Diez, C.; Hontanon, V.; Ibanez-Samaniego, L.; Perez-Latorre, L.; et al. HCV Cure with Direct-Acting Antivirals Improves Liver and Immunological Markers in HIV/HCV-Coinfected Patients. Front. Immunol. 2021, 12, 723196. [Google Scholar] [CrossRef] [PubMed]
- Ghiglione, Y.; Polo, M.L.; Urioste, A.; Rhodes, A.; Czernikier, A.; Trifone, C.; Quiroga, M.F.; Sisto, A.; Patterson, P.; Salomon, H.; et al. Hepatitis C Virus (HCV) Clearance After Treatment With Direct-Acting Antivirals in Human Immunodeficiency Virus (HIV)-HCV Coinfection Modulates Systemic Immune Activation and HIV Transcription on Antiretroviral Therapy. Open Forum Infect. Dis. 2020, 7, ofaa115. [Google Scholar] [CrossRef]
- Hamdane, N.; Juhling, F.; Crouchet, E.; El Saghire, H.; Thumann, C.; Oudot, M.A.; Bandiera, S.; Saviano, A.; Ponsolles, C.; Roca Suarez, A.A.; et al. HCV-Induced Epigenetic Changes Associated with Liver Cancer Risk Persist After Sustained Virologic Response. Gastroenterology 2019, 156, 2313–2329.e7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perez, S.; Kaspi, A.; Domovitz, T.; Davidovich, A.; Lavi-Itzkovitz, A.; Meirson, T.; Alison Holmes, J.; Dai, C.Y.; Huang, C.F.; Chung, R.T.; et al. Hepatitis C virus leaves an epigenetic signature post cure of infection by direct-acting antivirals. PLoS Genet. 2019, 15, e1008181. [Google Scholar] [CrossRef] [PubMed]
- Gupta, A.; Swaminathan, G.; Martin-Garcia, J.; Navas-Martin, S. MicroRNAs, hepatitis C virus, and HCV/HIV-1 co-infection: New insights in pathogenesis and therapy. Viruses 2012, 4, 2485–2513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyaaki, H.; Takatsuki, M.; Ichikawa, T.; Hidaka, M.; Soyama, A.; Ohdan, H.; Inomata, Y.; Uemoto, S.; Kokudo, N.; Nakao, K.; et al. Intrahepatic MicroRNA Profile of Liver Transplant Recipients with Hepatitis C Virus Co-Infected with Human Immunodeficiency Virus. Ann. Transplant. 2017, 22, 701–706. [Google Scholar] [CrossRef]
- van der Meer, A.J.; Farid, W.R.; Sonneveld, M.J.; de Ruiter, P.E.; Boonstra, A.; van Vuuren, A.J.; Verheij, J.; Hansen, B.E.; de Knegt, R.J.; van der Laan, L.J.; et al. Sensitive detection of hepatocellular injury in chronic hepatitis C patients with circulating hepatocyte-derived microRNA-122. J. Viral. Hepat. 2013, 20, 158–166. [Google Scholar] [CrossRef] [PubMed]
- Ha, M.; Kim, V.N. Regulation of microRNA biogenesis. Nat. Rev. Mol. Cell Biol. 2014, 15, 509–524. [Google Scholar] [CrossRef]
- Moghoofei, M.; Najafipour, S.; Mostafaei, S.; Tavakoli, A.; Bokharaei-Salim, F.; Ghorbani, S.; Javanmard, D.; Ghaffari, H.; Monavari, S.H. MicroRNAs Profiling in HIV, HCV, and HIV/HCV Co-Infected Patients. Curr. HIV Res. 2021, 19, 27–34. [Google Scholar] [CrossRef]
- Baccarani, U.; Righi, E.; Adani, G.L.; Lorenzin, D.; Pasqualucci, A.; Bassetti, M.; Risaliti, A. Pros and cons of liver transplantation in human immunodeficiency virus infected recipients. World J. Gastroenterol. 2014, 20, 5353–5362. [Google Scholar] [CrossRef]
- Ishak, K.; Baptista, A.; Bianchi, L.; Callea, F.; De Groote, J.; Gudat, F.; Denk, H.; Desmet, V.; Korb, G.; MacSween, R.N.; et al. Histological grading and staging of chronic hepatitis. J. Hepatol. 1995, 22, 696–699. [Google Scholar] [CrossRef]
- Vodkin, I.; Kuo, A. Extended Criteria Donors in Liver Transplantation. Clin. Liver Dis. 2017, 21, 289–301. [Google Scholar] [CrossRef] [PubMed]
- Martin, M. Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet. J. Next Gener. Seq. Data Anal. 2011, 17, 10–12. [Google Scholar] [CrossRef]
- Kozomara, A.; Griffiths-Jones, S. miRBase: Integrating microRNA annotation and deep-sequencing data. Nucleic Acids Res. 2011, 39, D152–D157. [Google Scholar] [CrossRef] [Green Version]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [Green Version]
- Vlachos, I.S.; Zagganas, K.; Paraskevopoulou, M.D.; Georgakilas, G.; Karagkouni, D.; Vergoulis, T.; Dalamagas, T.; Hatzigeorgiou, A.G. DIANA-miRPath v3.0: Deciphering microRNA function with experimental support. Nucleic Acids Res. 2015, 43, W460–W466. [Google Scholar] [CrossRef]
- Hernandez, M.D.; Sherman, K.E. HIV/hepatitis C coinfection natural history and disease progression. Curr. Opin. HIV AIDS 2011, 6, 478–482. [Google Scholar] [CrossRef] [Green Version]
- Liu, A.; Xu, X. MicroRNA isolation from formalin-fixed, paraffin-embedded tissues. Methods Mol. Biol. 2011, 724, 259–267. [Google Scholar] [CrossRef] [Green Version]
- Azzalini, E.; De Martino, E.; Fattorini, P.; Canzonieri, V.; Stanta, G.; Bonin, S. Reliability of miRNA Analysis from Fixed and Paraffin-Embedded Tissues. Int. J. Mol. Sci. 2019, 20, 4819. [Google Scholar] [CrossRef] [Green Version]
- Kawasaki, T.; Kawai, T. Toll-like receptor signaling pathways. Front. Immunol. 2014, 5, 461. [Google Scholar] [CrossRef] [Green Version]
- Crane, M.; Visvanathan, K.; Lewin, S.R. HIV Infection and TLR Signalling in the Liver. Gastroenterol. Res. Pract. 2012, 2012, 473925. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seki, E.; De Minicis, S.; Osterreicher, C.H.; Kluwe, J.; Osawa, Y.; Brenner, D.A.; Schwabe, R.F. TLR4 enhances TGF-beta signaling and hepatic fibrosis. Nat. Med. 2007, 13, 1324–1332. [Google Scholar] [CrossRef] [PubMed]
- Berzsenyi, M.D.; Roberts, S.K.; Preiss, S.; Woollard, D.J.; Beard, M.R.; Skinner, N.A.; Bowden, D.S.; Visvanathan, K. Hepatic TLR2 & TLR4 expression correlates with hepatic inflammation and TNF-alpha in HCV & HCV/HIV infection. J. Viral. Hepat. 2011, 18, 852–860. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; He, Z.; Hu, Y.; Wen, W.; Lin, C.; Yu, J.; Pan, J.; Li, R.; Deng, H.; Liao, S.; et al. MicroRNA-30e* suppresses dengue virus replication by promoting NF-kappaB-dependent IFN production. PLoS Negl. Trop. Dis. 2014, 8, e3088. [Google Scholar] [CrossRef] [PubMed]
- Mao, J.; Hu, X.; Pang, P.; Zhou, B.; Li, D.; Shan, H. miR-30e acts as a tumor suppressor in hepatocellular carcinoma partly via JAK1/STAT3 pathway. Oncol. Rep. 2017, 38, 393–401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chou, W.W.; Huang, C.F.; Yeh, M.L.; Tsai, Y.S.; Hsieh, M.Y.; Huang, C.I.; Huang, J.F.; Tsai, P.C.; Hsi, E.; Juo, S.H.; et al. MicroRNA let-7g cooperates with interferon/ribavirin to repress hepatitis C virus replication. J. Mol. Med. 2016, 94, 311–320. [Google Scholar] [CrossRef]
- Zhang, M.; Dong, B.B.; Lu, M.; Zheng, M.J.; Chen, H.; Ding, J.Z.; Xu, A.M.; Xu, Y.H. miR-429 functions as a tumor suppressor by targeting FSCN1 in gastric cancer cells. Onco Targets Ther. 2016, 9, 1123–1133. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Wu, Z.; Huang, P. The function of miRNAs in hepatocarcinogenesis induced by hepatitis B virus X protein (Review). Oncol. Rep. 2017, 38, 652–664. [Google Scholar] [CrossRef]
- Sanchez-Quijano, A.; Andreu, J.; Gavilan, F.; Luque, F.; Abad, M.A.; Soto, B.; Munoz, J.; Aznar, J.M.; Leal, M.; Lissen, E. Influence of human immunodeficiency virus type 1 infection on the natural course of chronic parenterally acquired hepatitis C. Eur. J. Clin. Microbiol. Infect. Dis. 1995, 14, 949–953. [Google Scholar] [CrossRef]
- Lodge, R.; Ferreira Barbosa, J.A.; Lombard-Vadnais, F.; Gilmore, J.C.; Deshiere, A.; Gosselin, A.; Wiche Salinas, T.R.; Bego, M.G.; Power, C.; Routy, J.P.; et al. Host MicroRNAs-221 and -222 Inhibit HIV-1 Entry in Macrophages by Targeting the CD4 Viral Receptor. Cell Rep. 2017, 21, 141–153. [Google Scholar] [CrossRef] [Green Version]
- Wang, D.; Sang, Y.; Sun, T.; Kong, P.; Zhang, L.; Dai, Y.; Cao, Y.; Tao, Z.; Liu, W. Emerging roles and mechanisms of microRNA2223p in human cancer (Review). Int. J. Oncol. 2021, 58, 20. [Google Scholar] [CrossRef] [PubMed]
- Pineau, P.; Volinia, S.; McJunkin, K.; Marchio, A.; Battiston, C.; Terris, B.; Mazzaferro, V.; Lowe, S.W.; Croce, C.M.; Dejean, A. miR-221 overexpression contributes to liver tumorigenesis. Proc. Natl. Acad. Sci. USA 2010, 107, 264–269. [Google Scholar] [CrossRef] [Green Version]
- Wong, Q.W.; Ching, A.K.; Chan, A.W.; Choy, K.W.; To, K.F.; Lai, P.B.; Wong, N. MiR-222 overexpression confers cell migratory advantages in hepatocellular carcinoma through enhancing AKT signaling. Clin. Cancer Res. 2010, 16, 867–875. [Google Scholar] [CrossRef] [Green Version]
- Ogawa, T.; Enomoto, M.; Fujii, H.; Sekiya, Y.; Yoshizato, K.; Ikeda, K.; Kawada, N. MicroRNA-221/222 upregulation indicates the activation of stellate cells and the progression of liver fibrosis. Gut 2012, 61, 1600–1609. [Google Scholar] [CrossRef] [PubMed]
- Song, J.; Ouyang, Y.; Che, J.; Li, X.; Zhao, Y.; Yang, K.; Zhao, X.; Chen, Y.; Fan, C.; Yuan, W. Potential Value of miR-221/222 as Diagnostic, Prognostic, and Therapeutic Biomarkers for Diseases. Front. Immunol. 2017, 8, 56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cesselli, D.; Parisse, P.; Aleksova, A.; Veneziano, C.; Cervellin, C.; Zanello, A.; Beltrami, A.P. Extracellular Vesicles: How Drug and Pathology Interfere With Their Biogenesis and Function. Front. Physiol. 2018, 9, 1394. [Google Scholar] [CrossRef] [PubMed]
- Parisse, P.; Rago, I.; Ulloa Severino, L.; Perissinotto, F.; Ambrosetti, E.; Paoletti, P.; Ricci, M.; Beltrami, A.P.; Cesselli, D.; Casalis, L. Atomic force microscopy analysis of extracellular vesicles. Eur. Biophys. J. 2017, 46, 813–820. [Google Scholar] [CrossRef] [PubMed]
- Wijarnpreecha, K.; Thongprayoon, C.; Panjawatanan, P.; Ungprasert, P. Hepatitis C Virus Infection and Risk of Venous Thromboembolism: A Systematic Review and Meta-Analysis. Ann. Hepatol. 2017, 16, 514–520. [Google Scholar] [CrossRef]
- Rokx, C.; Borjas Howard, J.F.; Smit, C.; Wit, F.W.; Pieterman, E.D.; Reiss, P.; Cannegieter, S.C.; Lijfering, W.M.; Meijer, K.; Bierman, W.; et al. Risk of recurrent venous thromboembolism in patients with HIV infection: A nationwide cohort study. PLoS Med. 2020, 17, e1003101. [Google Scholar] [CrossRef]
- Pergoli, L.; Cantone, L.; Favero, C.; Angelici, L.; Iodice, S.; Pinatel, E.; Hoxha, M.; Dioni, L.; Letizia, M.; Albetti, B.; et al. Extracellular vesicle-packaged miRNA release after short-term exposure to particulate matter is associated with increased coagulation. Part. Fibre Toxicol. 2017, 14, 32. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Liu, C.; Wei, W.; Chen, W. Predictive value of circulating coagulation related microRNAs expressions for major adverse cardiac and cerebral event risk in patients undergoing continuous ambulatory peritoneal dialysis: A cohort study. J. Nephrol. 2020, 33, 157–165. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Wen, Q.; Chen, H.; Wu, X.; Liu, B.; Li, H.; Su, L.; Tong, H. Exosomes Derived From Heat Stroke Cases Carry miRNAs Associated With Inflammation and Coagulation Cascade. Front. Immunol. 2021, 12, 624753. [Google Scholar] [CrossRef] [PubMed]
- Chettimada, S.; Lorenz, D.R.; Misra, V.; Dillon, S.T.; Reeves, R.K.; Manickam, C.; Morgello, S.; Kirk, G.D.; Mehta, S.H.; Gabuzda, D. Exosome markers associated with immune activation and oxidative stress in HIV patients on antiretroviral therapy. Sci. Rep. 2018, 8, 7227. [Google Scholar] [CrossRef] [PubMed]
- Welch, J.L.; Stapleton, J.T.; Okeoma, C.M. Vehicles of intercellular communication: Exosomes and HIV-1. J. Gen. Virol. 2019, 100, 350–366. [Google Scholar] [CrossRef]
- Faccioli, J.; Nardelli, S.; Gioia, S.; Riggio, O.; Ridola, L. Neurological and psychiatric effects of hepatitis C virus infection. World J. Gastroenterol. 2021, 27, 4846–4861. [Google Scholar] [CrossRef]
- Nawaz, R.; Zahid, S.; Idrees, M.; Rafique, S.; Shahid, M.; Ahad, A.; Amin, I.; Almas, I.; Afzal, S. HCV-induced regulatory alterations of IL-1beta, IL-6, TNF-alpha, and IFN-Upsilon operative, leading liver en-route to non-alcoholic steatohepatitis. Inflamm. Res. 2017, 66, 477–486. [Google Scholar] [CrossRef]
- Thakur, K.T.; Boubour, A.; Saylor, D.; Das, M.; Bearden, D.R.; Birbeck, G.L. Global HIV neurology: A comprehensive review. AIDS 2019, 33, 163–184. [Google Scholar] [CrossRef]
- Alford, K.; Vera, J.H. Cognitive Impairment in people living with HIV in the ART era: A Review. Br. Med. Bull. 2018, 127, 55–68. [Google Scholar] [CrossRef] [PubMed]
- O’Meara, T.; Kong, Y.; Chiarella, J.; Price, R.W.; Chaudhury, R.; Liu, X.; Spudich, S.; Robertson, K.; Emu, B.; Lu, L. Exosomal MicroRNAs Associate With Neuropsychological Performance in Individuals with HIV Infection on Antiretroviral Therapy. J. Acquir. Immune Defic. Syndr. 2019, 82, 514–522. [Google Scholar] [CrossRef] [PubMed]
- Kutchy, N.A.; Peeples, E.S.; Sil, S.; Liao, K.; Chivero, E.T.; Hu, G.; Buch, S. Extracellular Vesicles in Viral Infections of the Nervous System. Viruses 2020, 12, 700. [Google Scholar] [CrossRef]
- Zhang, H.; Gao, S.; Pei, R.; Chen, X.; Li, C. Hepatitis C virus-induced prion protein expression facilitates hepatitis C virus replication. Virol. Sin. 2017, 32, 503–510. [Google Scholar] [CrossRef] [PubMed]
- Megra, B.W.; Eugenin, E.A.; Berman, J.W. The Role of Shed PrP(c) in the Neuropathogenesis of HIV Infection. J. Immunol. 2017, 199, 224–232. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.; Zhang, Y.; Su, Y.; Tian, L.; Qin, P.; Xu, X.; Zhou, Y. microRNA-125a targets MAVS and TRAF6 to modulate interferon signaling and promote HCV infection. Virus Res. 2021, 296, 198336. [Google Scholar] [CrossRef] [PubMed]
- Youness, R.A.; El-Tayebi, H.M.; Assal, R.A.; Hosny, K.; Esmat, G.; Abdelaziz, A.I. MicroRNA-486-5p enhances hepatocellular carcinoma tumor suppression through repression of IGF-1R and its downstream mTOR, STAT3 and c-Myc. Oncol. Lett. 2016, 12, 2567–2573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gragnani, L.; Piluso, A.; Fognani, E.; Zignego, A.L. MicroRNA expression in hepatitis C virus-related malignancies: A brief review. World J. Gastroenterol. 2015, 21, 8562–8568. [Google Scholar] [CrossRef] [PubMed]
- Roderburg, C.; Urban, G.W.; Bettermann, K.; Vucur, M.; Zimmermann, H.; Schmidt, S.; Janssen, J.; Koppe, C.; Knolle, P.; Castoldi, M.; et al. Micro-RNA profiling reveals a role for miR-29 in human and murine liver fibrosis. Hepatology 2011, 53, 209–218. [Google Scholar] [CrossRef]
- Jiang, X.P.; Ai, W.B.; Wan, L.Y.; Zhang, Y.Q.; Wu, J.F. The roles of microRNA families in hepatic fibrosis. Cell Biosci. 2017, 7, 34. [Google Scholar] [CrossRef] [Green Version]
- Kwon, J.J.; Factora, T.D.; Dey, S.; Kota, J. A Systematic Review of miR-29 in Cancer. Mol. Ther. Oncolytics 2019, 12, 173–194. [Google Scholar] [CrossRef] [Green Version]
- Hayes, C.N.; Chayama, K. MicroRNAs as Biomarkers for Liver Disease and Hepatocellular Carcinoma. Int. J. Mol. Sci. 2016, 17, 280. [Google Scholar] [CrossRef]
- Pereira, P.A.; Tomas, J.F.; Queiroz, J.A.; Figueiras, A.R.; Sousa, F. Recombinant pre-miR-29b for Alzheimer s disease therapeutics. Sci. Rep. 2016, 6, 19946. [Google Scholar] [CrossRef]
- Li, J.; Wang, L.; Hua, X.; Tang, H.; Chen, R.; Yang, T.; Das, S.; Xiao, J. CRISPR/Cas9-Mediated miR-29b Editing as a Treatment of Different Types of Muscle Atrophy in Mice. Mol. Ther. 2020, 28, 1359–1372. [Google Scholar] [CrossRef] [Green Version]
- Shin, J.; Shim, H.G.; Hwang, T.; Kim, H.; Kang, S.H.; Dho, Y.S.; Park, S.H.; Kim, S.J.; Park, C.K. Restoration of miR-29b exerts anti-cancer effects on glioblastoma. Cancer Cell Int. 2017, 17, 104. [Google Scholar] [CrossRef] [PubMed]
- Jahangard, Y.; Monfared, H.; Moradi, A.; Zare, M.; Mirnajafi-Zadeh, J.; Mowla, S.J. Therapeutic Effects of Transplanted Exosomes Containing miR-29b to a Rat Model of Alzheimer’s Disease. Front. Neurosci. 2020, 14, 564. [Google Scholar] [CrossRef] [PubMed]
- Franco, S.; Buccione, D.; Tural, C.; Martinez, M.A. Circulating microRNA signatures that predict liver fibrosis progression in patients with HIV-1/hepatitis C virus coinfections. AIDS 2021, 35, 1355–1363. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| HCV Mono-Infected vs. Controls | HCV/HIV Co-Infected vs. Controls | HCV/HIV Co-Infected vs. HCV Mono-Infected | |||
|---|---|---|---|---|---|
| Up Regulated | Down Regulated | Up Regulated | Down Regulated | Up Regulated | Down Regulated |
| hsa-miR-18a-5p | hsa-let-7d-3p | hsa-miR-301a-3p | hsa-miR-193b-5p | hsa-miR-675-3p | hsa-miR-409-5p |
| hsa-miR-382-3p | hsa-miR-423-3p | hsa-miR-142-5p | hsa-miR-671-3p | hsa-miR-1269b | hsa-miR-660-5p |
| hsa-miR-651 5p | hsa-miR-193a-5p | hsa-miR-19a-3p | hsa-miR-486-3p | hsa-miR-342-5p | hsa-miR-190a-5p |
| hsa-miR-655-3p | hsa-miR-483-5p | hsa-miR-222-3p | hsa-miR-7706 | hsa-miR-29b-3p | |
| hsa-miR-301a-3p | hsa-miR-423-5p | hsa-miR-19b-3p | hsa-miR-30c-1-3p | hsa-miR-374a-3p | |
| hsa-let-7g-3p | hsa-miR-92b-3p | hsa-miR-423-5p | |||
| hsa-miR-100-3p | hsa-miR-193b-5p | hsa-miR-125a-5p | |||
| hsa-miR-409-5p | hsa-miR-671-3p | hsa-miR-320a-3p | |||
| hsa-miR-374a-3p | hsa-miR-486-5p | hsa-miR-486-5p | |||
| hsa-miR-19a-3p | hsa-miR-125a-5p | hsa-miR-146a-5p | |||
| hsa-miR-19b-3p | hsa-miR-210-5p | ||||
| hsa-miR-142-5p | hsa-miR-486-3p | ||||
| hsa-miR-582-5p | hsa-miR-1247-5p | ||||
| hsa-miR-429 | hsa-miR-3605-3p | ||||
| hsa-miR-660-5p | hsa-miR-4443 | ||||
| hsa-miR-21-5p | |||||
| hsa-miR-542-3p | |||||
| hsa-miR-142-3p | |||||
| hsa-miR-101-3p | |||||
| hsa-miR-501-3p | |||||
| hsa-miR-30e-5p | |||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dalla, E.; Bulfoni, M.; Cesselli, D.; Pravisani, R.; Hidaka, M.; Eguchi, S.; Baccarani, U. Reinfection of Transplanted Livers in HCV- and HCV/HIV-Infected Patients Is Characterized by a Different MicroRNA Expression Profile. Cells 2022, 11, 690. https://doi.org/10.3390/cells11040690
Dalla E, Bulfoni M, Cesselli D, Pravisani R, Hidaka M, Eguchi S, Baccarani U. Reinfection of Transplanted Livers in HCV- and HCV/HIV-Infected Patients Is Characterized by a Different MicroRNA Expression Profile. Cells. 2022; 11(4):690. https://doi.org/10.3390/cells11040690
Chicago/Turabian StyleDalla, Emiliano, Michela Bulfoni, Daniela Cesselli, Riccardo Pravisani, Masaaki Hidaka, Susumu Eguchi, and Umberto Baccarani. 2022. "Reinfection of Transplanted Livers in HCV- and HCV/HIV-Infected Patients Is Characterized by a Different MicroRNA Expression Profile" Cells 11, no. 4: 690. https://doi.org/10.3390/cells11040690
APA StyleDalla, E., Bulfoni, M., Cesselli, D., Pravisani, R., Hidaka, M., Eguchi, S., & Baccarani, U. (2022). Reinfection of Transplanted Livers in HCV- and HCV/HIV-Infected Patients Is Characterized by a Different MicroRNA Expression Profile. Cells, 11(4), 690. https://doi.org/10.3390/cells11040690