
Tendon Immune Regeneration: Insights on the Synergetic Role of Stem and Immune Cells during Tendon Regeneration

 ,
,  ,
,  ,
,  ,
,  ,
,  ,
, 
 ,
,  ,
, 
Abstract
:1. Introduction
2. Tendon Structure and Homeostasis
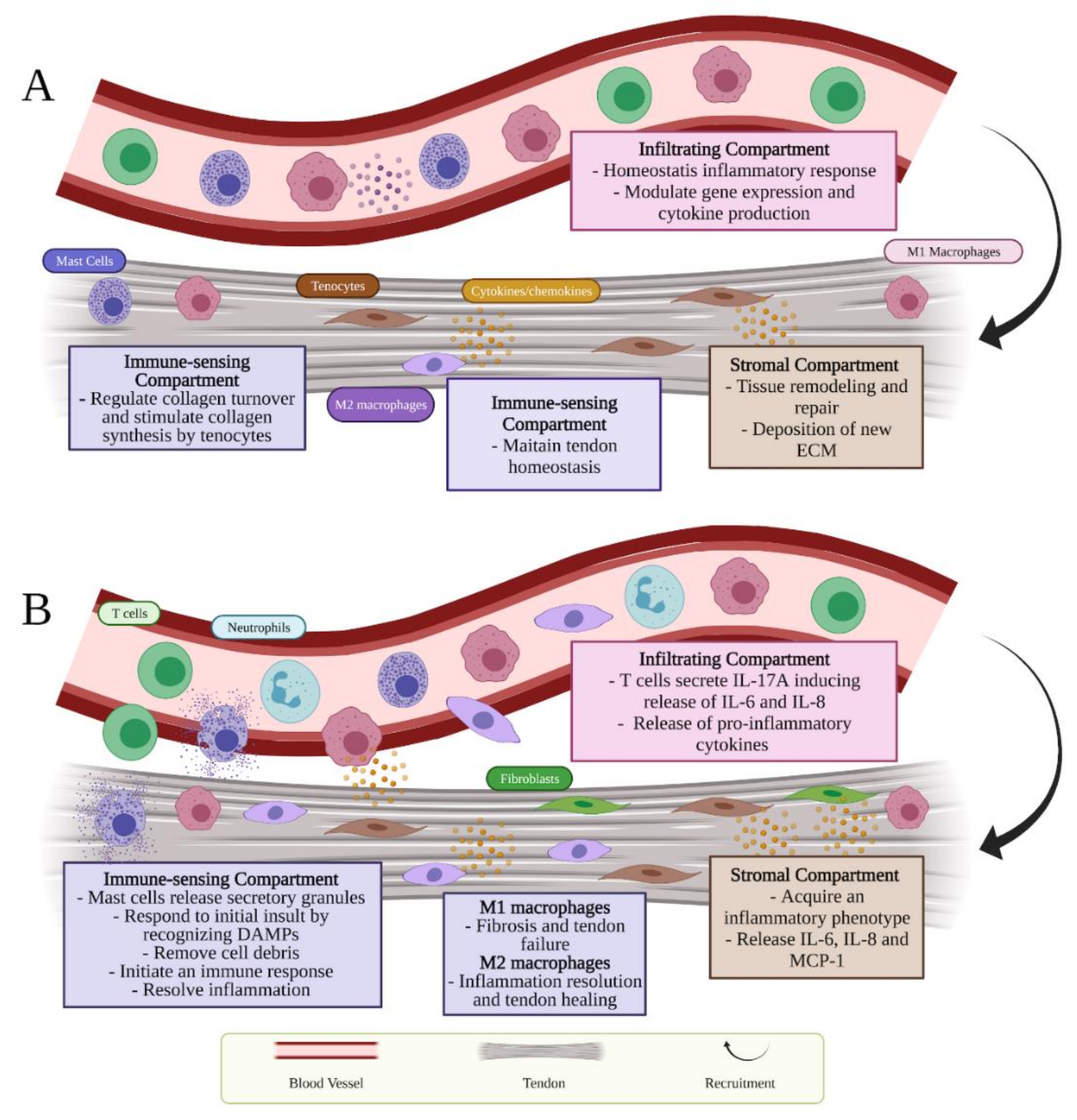
2.1. Tendon Stromal Compartment
2.2. Tendon Immune-Sensing Compartment
2.3. Infiltrating Compartment
3. Inflammatory Response Occurring during Tendon Injuries
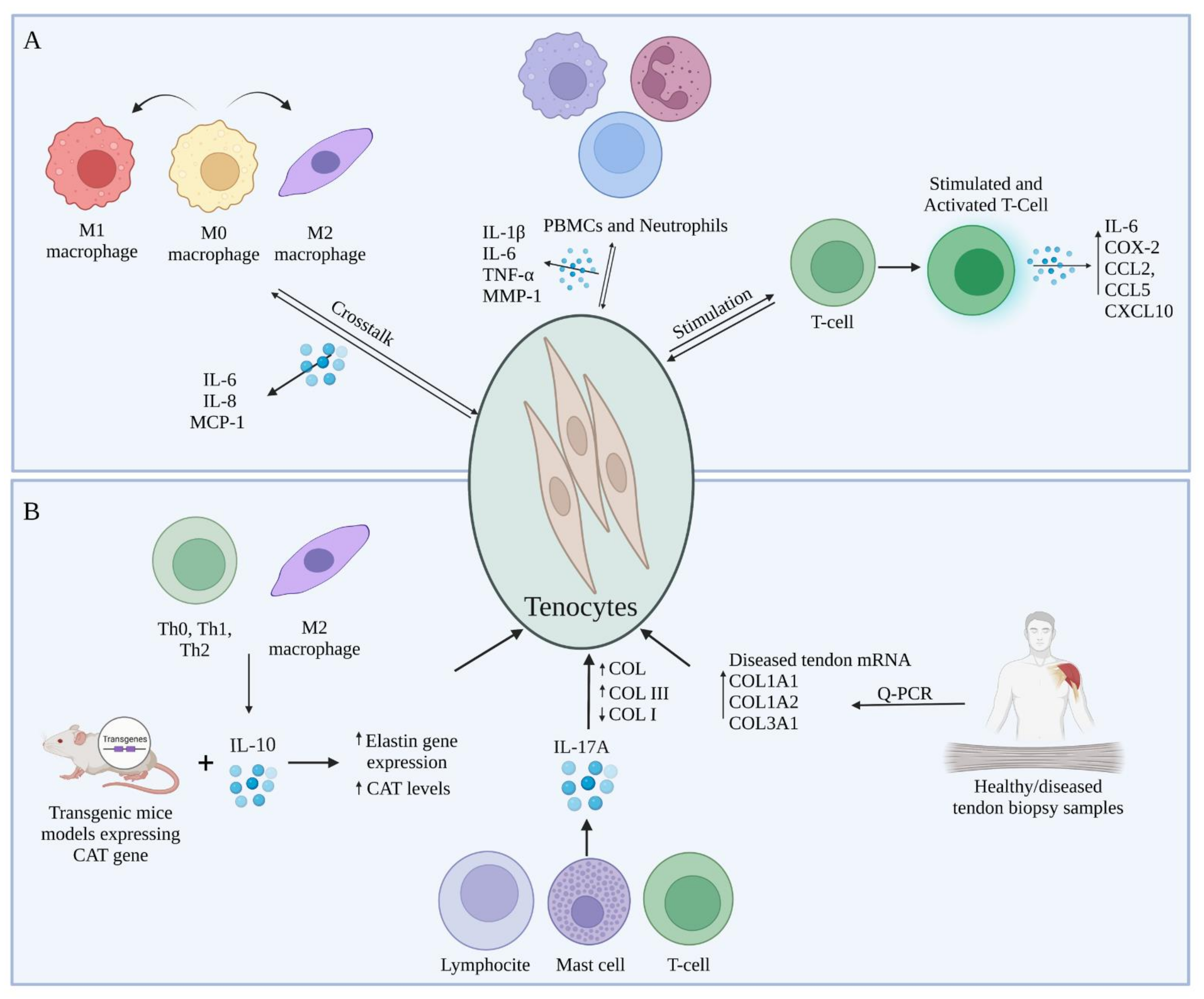
4. Crosstalk between Tenocytes and Immune Cells during Tendon Regeneration
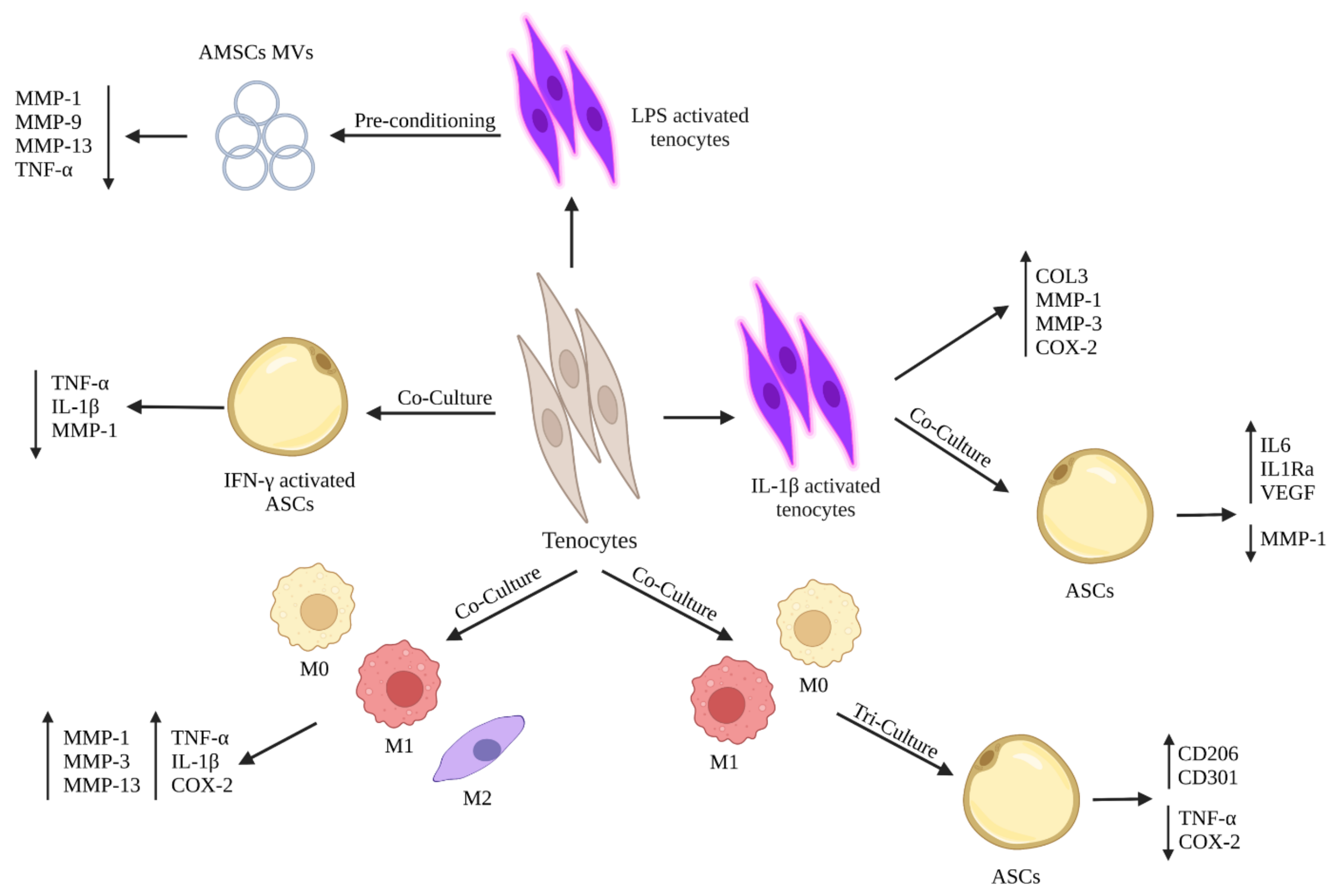
5. Crosstalk between Stem Cells and Tenocytes to Modulate Their Inflammatory Phenotype
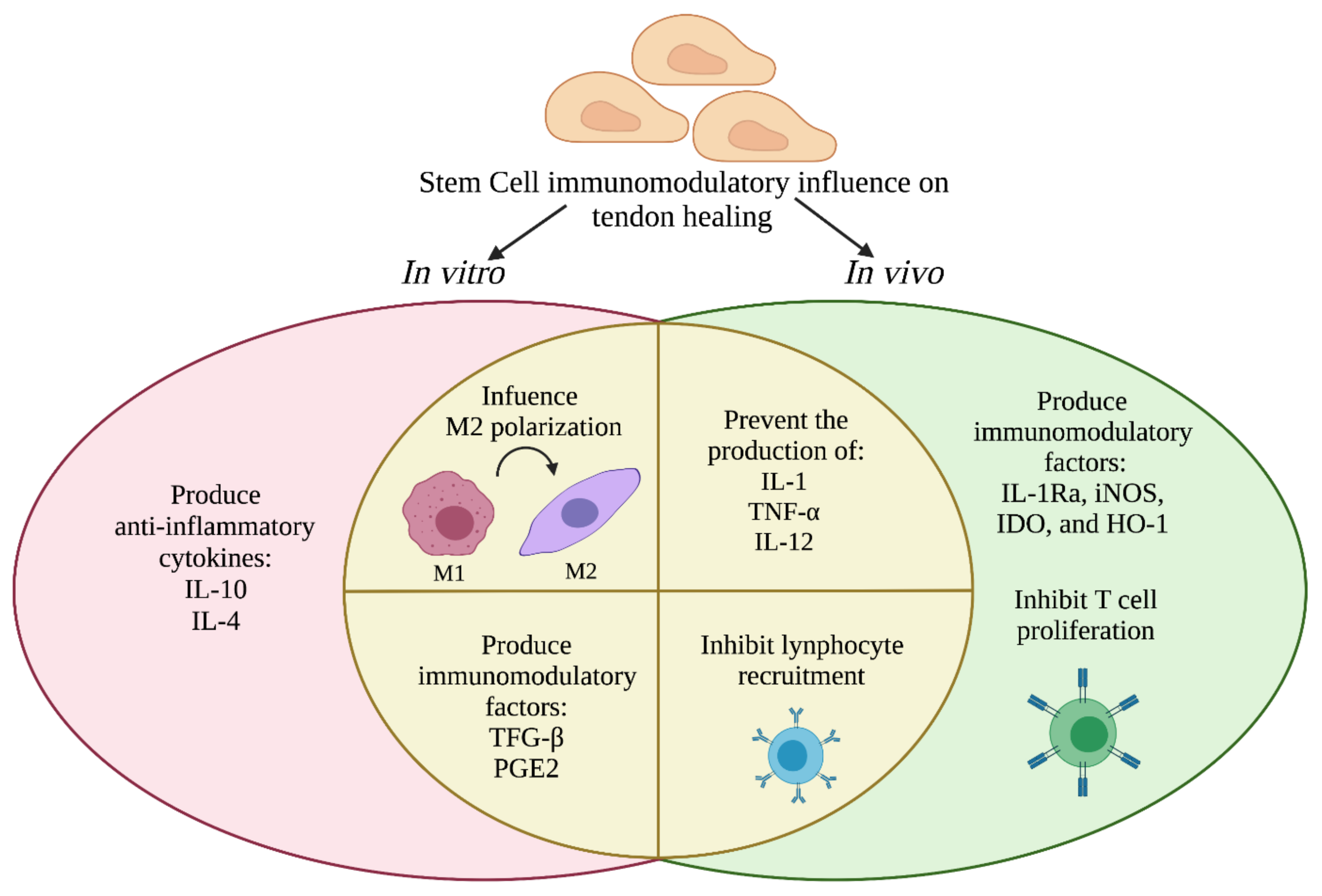
6. Crosstalk between Stem Cells and Immune Cells during Tendon Regeneration
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kirkendall, D.T.; Garrett, W.E. Function and Biomechanics of Tendons. Scand. J. Med. Sci. Sports 1997, 7, 62–66. [Google Scholar] [CrossRef]
- Lomas, A.J.; Ryan, C.N.M.; Sorushanova, A.; Shologu, N.; Sideri, A.I.; Tsioli, V.; Fthenakis, G.C.; Tzora, A.; Skoufos, I.; Quinlan, L.R.; et al. The Past, Present and Future in Scaffold-Based Tendon Treatments. Adv. Drug Deliv. Rev. 2015, 84, 257–277. [Google Scholar] [CrossRef]
- Millar, N.L.; Murrell, G.A.C.; Mcinnes, I.B. Inflammatory Mechanisms in Tendinopathy—Towards Translation. Nat. Rev. Rheumatol. 2017, 13, 110–122. [Google Scholar] [CrossRef] [PubMed]
- Aicale, R.; Tarantino, D.; Maffulli, N. Basic Science of Tendons. In Bio-Orthopaedics: A New Approach; Gobbi, A., Lane, J.G., Espregueira-Mendes, J., Karahan, M., Eds.; Springer: Berlin/Heidelberg, Germany, 2017; pp. 249–273. ISBN 9783662541814. [Google Scholar]
- Millar, N.L.; Hueber, A.J.; Reilly, J.H.; Yinghua, X.; Fazzi, U.G.; Murrell, G.A.C.; McInnes, I.B. Inflammation Is Present in Early Human Tendinopathy. Am. J. Sports Med. 2010, 38, 2085–2091. [Google Scholar] [CrossRef]
- Mosca, M.J.; Rashid, M.S.; Snelling, S.J.; Kirtley, S.; Carr, A.J.; Dakin, S.G. Trends in the Theory That Inflammation Plays a Causal Role in Tendinopathy: A Systematic Review and Quantitative Analysis of Published Reviews. BMJ Open Sport Exerc. Med. 2018, 4, e000332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, P.; Maffulli, N. Tendon Injury and Tendinopathy: Healing and Repair. J. Bone Jt. Surg.—Ser. A 2005, 87, 187–202. [Google Scholar] [CrossRef] [Green Version]
- Okabayashi, K.; Ashrafian, H.; Zacharakis, E.; Hasegawa, H.; Kitagawa, Y.; Athanasiou, T.; Darzi, A. Adhesions after Abdominal Surgery: A Systematic Review of the Incidence, Distribution and Severity. Surg. Today 2014, 44, 405–420. [Google Scholar] [CrossRef]
- Voleti, P.B.; Buckley, M.R.; Soslowsky, L.J. Tendon Healing: Repair and Regeneration. Annu. Rev. Biomed. Eng. 2012, 14, 47–71. [Google Scholar] [CrossRef] [PubMed]
- Drake, D.B.; Tilt, A.C.; DeGeorge, B.R. Acellular Flexor Tendon Allografts: A New Horizon for Tendon Reconstruction. J. Hand Surg. 2013, 38, 2491–2495. [Google Scholar] [CrossRef] [PubMed]
- Capella-Monsonís, H.; Kearns, S.; Kelly, J.; Zeugolis, D.I. Battling Adhesions: From Understanding to Prevention. BMC Biomed. Eng. 2019, 1, 5. [Google Scholar] [CrossRef] [Green Version]
- Thomopoulos, S.; Parks, W.C.; Rifkin, D.B.; Derwin, K.A. Mechanisms of Tendon Injury and Repair: Tendon Injury and Repair. J. Orthop. Res. 2015, 33, 832–839. [Google Scholar] [CrossRef] [Green Version]
- Maeda, E.; Shelton, J.C.; Bader, D.L.; Lee, D.A. Differential Regulation of Gene Expression in Isolated Tendon Fascicles Exposed to Cyclic Tensile Strain in Vitro. J. Appl. Physiol. 2009, 106, 506–512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wong, J.K.F.; Metcalfe, A.D.; Wong, R.; Bush, J.; Platt, C.; Garcon, A.; Goldspink, N.; McGrouther, D.A.; Ferguson, M.W.J. Reduction of Tendon Adhesions Following Administration of Adaprev, a Hypertonic Solution of Mannose-6-Phosphate: Mechanism of Action Studies. PLoS ONE 2014, 9, e112672. [Google Scholar] [CrossRef] [Green Version]
- Galloway, M.T.; Lalley, A.L.; Shearn, J.T. The Role of Mechanical Loading in Tendon Development, Maintenance, Injury, and Repair. J. Bone Jt. Surg. 2013, 95, 1620–1628. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, M. Structure and Metabolism of Tendons. Scand. J. Med. Sci. Sports 1997, 7, 55–61. [Google Scholar] [CrossRef]
- Ackermann, P.W. Tendinopathies in Sports: From Basic Research to the Field. In Sports Injuries: Prevention, Diagnosis, Treatment and Rehabilitation; Doral, M.N., Karlsson, J., Eds.; Springer: Berlin/Heidelberg, Germany, 2014; pp. 1–15. ISBN 9783642368011. [Google Scholar]
- Barboni, B.; Russo, V.; Berardinelli, P.; Mauro, A.; Valbonetti, L.; Sanyal, H.; Canciello, A.; Greco, L.; Muttini, A.; Gatta, V.; et al. Placental Stem Cells from Domestic Animals: Translational Potential and Clinical Relevance. Cell Transplant. 2018, 27, 93–116. [Google Scholar] [CrossRef]
- Barboni, B.; Russo, V.; Curini, V.; Mauro, A.; Martelli, A.; Muttini, A.; Bernabò, N.; Valbonetti, L.; Marchisio, M.; Di Giacinto, O.; et al. Achilles Tendon Regeneration Can Be Improved by Amniotic Epithelial Cell Allotransplantation. Cell Transplant. 2012, 21, 2377–2395. [Google Scholar] [CrossRef] [Green Version]
- Mauro, A.; Russo, V.; Di Marcantonio, L.; Berardinelli, P.; Martelli, A.; Muttini, A.; Mattioli, M.; Barboni, B. M1 and M2 Macrophage Recruitment during Tendon Regeneration Induced by Amniotic Epithelial Cell Allotransplantation in Ovine. Res. Vet. Sci. 2016, 105, 92–102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Russo, V.; Paolo, B.; Gatta, V.; Muttini, A.; Stuppia, L.; Parolini, O.; Mattioli, M.; Barboni, B. Cross-Talk between Human Amniotic Derived Cells and Host Tendon Supports Tissue Regeneration. J. Tissue Eng. Regen. Med. 2014, 8, 142. [Google Scholar]
- Ilaltdinov, A.W.; Gong, Y.; Leong, D.J.; Fung, D.T.; Sun, H.B. Tendon Homeostasis: Overview. In Encyclopedia of Bone Biology; Elsevier: Amsterdam, The Netherlands, 2020; pp. 270–293. ISBN 978-0-12-814082-6. [Google Scholar]
- Sensini, A.; Cristofolini, L. Biofabrication of Electrospun Scaffolds for the Regeneration of Tendons and Ligaments. Materials 2018, 11, 1963. [Google Scholar] [CrossRef] [Green Version]
- Citeroni, M.R.; Ciardulli, M.C.; Russo, V.; Della Porta, G.; Mauro, A.; El Khatib, M.; Di Mattia, M.; Galesso, D.; Barbera, C.; Forsyth, N.R.; et al. In Vitro Innovation of Tendon Tissue Engineering Strategies. Int. J. Mol. Sci. 2020, 21, 6726. [Google Scholar] [CrossRef]
- De Micheli, A.J.; Swanson, J.B.; Disser, N.P.; Martinez, L.M.; Walker, N.R.; Oliver, D.J.; Cosgrove, B.D.; Mendias, C.L. Single-Cell Transcriptomic Analysis Identifies Extensive Heterogeneity in the Cellular Composition of Mouse Achilles Tendons. Am. J. Physiol. Cell Physiol. 2020, 319, C885–C894. [Google Scholar] [CrossRef]
- Luesma, M.J.; Cantarero, I.; Sánchez-Cano, A.I.; Rodellar, C.; Junquera, C. Ultrastructural Evidence for Telocytes in Equine Tendon. J. Anat. 2021, 238, 527–535. [Google Scholar] [CrossRef]
- Nichols, A.E.C.; Muscat, S.N.; Miller, S.E.; Green, L.J.; Richards, M.S.; Loiselle, A.E. Impact of Isolation Method on Cellular Activation and Presence of Specific Tendon Cell Subpopulations during in Vitro Culture. FASEB J. 2021, 35. [Google Scholar] [CrossRef]
- Paolillo, C.; Londin, E.; Fortina, P. Single-Cell Genomics. Clin. Chem. 2019, 65, 972–985. [Google Scholar] [CrossRef]
- Zhang, C.; Zhu, J.; Zhou, Y.; Thampatty, B.P.; Wang, J.H.-C. Tendon Stem/Progenitor Cells and Their Interactions with Extracellular Matrix and Mechanical Loading. Stem Cells Int. 2019, 2019, 3674647. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chuen, F.S.; Chuk, C.Y.; Ping, W.Y.; Nar, W.W.; Kim, H.L.; Ming, C.K. Immunohistochemical Characterization of Cells in Adult Human Patellar Tendons. J. Histochem. Cytochem. 2004, 52, 1151–1157. [Google Scholar] [CrossRef] [PubMed]
- Docheva, D.; Müller, S.A.; Majewski, M.; Evans, C.H. Biologics for Tendon Repair. Adv. Drug Deliv. Rev. 2015, 84, 222–239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Russo, V.; Mauro, A.; Martelli, A.; Di Giacinto, O.; Di Marcantonio, L.; Nardinocchi, D.; Berardinelli, P.; Barboni, B. Cellular and Molecular Maturation in Fetal and Adult Ovine Calcaneal Tendons. J. Anat. 2015, 226, 126–142. [Google Scholar] [CrossRef] [Green Version]
- McNeilly, C.M.; Banes, A.J.; Benjamin, M.; Ralphs, J.R. Tendon Cells in Vivo Form a Three Dimensional Network of Cell Processes Linked by Gap Junctions. J. Anat. 1996, 189 Pt 3, 593–600. [Google Scholar]
- Bi, Y.; Ehirchiou, D.; Kilts, T.M.; Inkson, C.A.; Embree, M.C.; Sonoyama, W.; Li, L.; Leet, A.I.; Seo, B.-M.; Zhang, L.; et al. Identification of Tendon Stem/Progenitor Cells and the Role of the Extracellular Matrix in Their Niche. Nat. Med. 2007, 13, 1219–1227. [Google Scholar] [CrossRef] [PubMed]
- Mienaltowski, M.J.; Adams, S.M.; Birk, D.E. Regional Differences in Stem Cell/Progenitor Cell Populations from the Mouse Achilles Tendon. Tissue Eng. Part A 2013, 19, 199–210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lui, P.P.Y. Identity of Tendon Stem Cells—How Much Do We Know? J. Cell. Mol. Med. 2013, 17, 55–64. [Google Scholar] [CrossRef] [PubMed]
- Salingcarnboriboon, R. Establishment of Tendon-Derived Cell Lines Exhibiting Pluripotent Mesenchymal Stem Cell-like Property. Exp. Cell Res. 2003, 287, 289–300. [Google Scholar] [CrossRef]
- Ruzzini, L.; Abbruzzese, F.; Rainer, A.; Longo, U.G.; Trombetta, M.; Maffulli, N.; Denaro, V. Characterization of Age-Related Changes of Tendon Stem Cells from Adult Human Tendons. Knee Surg. Sports Traumatol. Arthrosc. 2014, 22, 2856–2866. [Google Scholar] [CrossRef] [PubMed]
- Blondheim, N.R.; Levy, Y.S.; Ben-Zur, T.; Burshtein, A.; Cherlow, T.; Kan, I.; Barzilai, R.; Bahat-Stromza, M.; Barhum, Y.; Bulvik, S.; et al. Human Mesenchymal Stem Cells Express Neural Genes, Suggesting a Neural Predisposition. Stem Cells Dev. 2006, 15, 141–164. [Google Scholar] [CrossRef] [PubMed]
- Lehner, C.; Spitzer, G.; Gehwolf, R.; Wagner, A.; Weissenbacher, N.; Deininger, C.; Emmanuel, K.; Wichlas, F.; Tempfer, H.; Traweger, A. Tenophages: A Novel Macrophage-like Tendon Cell Population Expressing CX3CL1 and CX3CR1. DMM Dis. Models Mech. 2019, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murray, P.J.; Allen, J.E.; Biswas, S.K.; Fisher, E.A.; Gilroy, D.W.; Goerdt, S.; Gordon, S.; Hamilton, J.A.; Ivashkiv, L.B.; Lawrence, T.; et al. Macrophage Activation and Polarization: Nomenclature and Experimental Guidelines. Immunity 2014, 41, 14–20. [Google Scholar] [CrossRef] [Green Version]
- Kabashima, K.; Nakashima, C.; Nonomura, Y.; Otsuka, A.; Cardamone, C.; Parente, R.; De Feo, G.; Triggiani, M. Biomarkers for Evaluation of Mast Cell and Basophil Activation. Immunol. Rev. 2018, 282, 114–120. [Google Scholar] [CrossRef]
- Alim, M.A.; Ackermann, P.W.; Eliasson, P.; Blomgran, P.; Kristiansson, P.; Pejler, G.; Peterson, M. Increased Mast Cell Degranulation and Co-Localization of Mast Cells with the NMDA Receptor-1 during Healing after Achilles Tendon Rupture. Cell Tissue Res. 2017, 370, 451–460. [Google Scholar] [CrossRef] [Green Version]
- Krystel-Whittemore, M.; Dileepan, K.N.; Wood, J.G. Mast Cell: A Multi-Functional Master Cell. Front. Immunol. 2016, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alim, M.A.; Peterson, M.; Pejler, G. Do Mast Cells Have a Role in Tendon Healing and Inflammation? Cells 2020, 9, 1134. [Google Scholar] [CrossRef] [PubMed]
- D’Addona, A.; Maffulli, N.; Formisano, S.; Rosa, D. Inflammation in Tendinopathy. Surgeon 2017, 15, 297–302. [Google Scholar] [CrossRef] [PubMed]
- Thampatty, B.P.; Li, H.; Im, H.-J.; Wang, J.H.-C. EP4 Receptor Regulates Collagen Type-I, MMP-1, and MMP-3 Gene Expression in Human Tendon Fibroblasts in Response to IL-1β Treatment. Gene 2007, 386, 154–161. [Google Scholar] [CrossRef] [Green Version]
- Lavagnino, M.; Wall, M.E.; Little, D.; Banes, A.J.; Guilak, F.; Arnoczky, S.P. Tendon Mechanobiology: Current Knowledge and Future Research Opportunities. J. Orthop. Res. 2015, 33, 813–822. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eliasson, P.; Andersson, T.; Aspenberg, P. Rat Achilles Tendon Healing: Mechanical Loading and Gene Expression. J. Appl. Physiol. 2009, 107, 399–407. [Google Scholar] [CrossRef] [Green Version]
- Gaut, L.; Robert, N.; Delalande, A.; Bonnin, M.-A.; Pichon, C.; Duprez, D. EGR1 Regulates Transcription Downstream of Mechanical Signals during Tendon Formation and Healing. PLoS ONE 2016, 11, e0166237. [Google Scholar] [CrossRef] [Green Version]
- Guerquin, M.-J.; Charvet, B.; Nourissat, G.; Havis, E.; Ronsin, O.; Bonnin, M.-A.; Ruggiu, M.; Olivera-Martinez, I.; Robert, N.; Lu, Y.; et al. Transcription Factor EGR1 Directs Tendon Differentiation and Promotes Tendon Repair. J. Clin. Investig. 2013, 123, 3564–3576. [Google Scholar] [CrossRef] [Green Version]
- Lejard, V.; Blais, F.; Guerquin, M.-J.; Bonnet, A.; Bonnin, M.-A.; Havis, E.; Malbouyres, M.; Bidaud, C.B.; Maro, G.; Gilardi-Hebenstreit, P.; et al. EGR1 and EGR2 Involvement in Vertebrate Tendon Differentiation. J. Biol. Chem. 2011, 286, 5855–5867. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Zhang, C.; Zhu, S.; Lu, P.; Zhu, T.; Gong, X.; Zhang, Z.; Hu, J.; Yin, Z.; Heng, B.C.; et al. Mohawk Promotes the Tenogenesis of Mesenchymal Stem Cells Through Activation of the TGF β Signaling Pathway. Stem Cells 2015, 33, 443–455. [Google Scholar] [CrossRef]
- Pryce, B.A.; Watson, S.S.; Murchison, N.D.; Staverosky, J.A.; Dünker, N.; Schweitzer, R. Recruitment and Maintenance of Tendon Progenitors by TGFβ Signaling Are Essential for Tendon Formation. Development 2009, 136, 1351–1361. [Google Scholar] [CrossRef] [Green Version]
- Maeda, T.; Sakabe, T.; Sunaga, A.; Sakai, K.; Rivera, A.L.; Keene, D.R.; Sasaki, T.; Stavnezer, E.; Iannotti, J.; Schweitzer, R.; et al. Conversion of Mechanical Force into TGF-β-Mediated Biochemical Signals. Curr. Biol. 2011, 21, 933–941. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Suen, C.-W.; Zhang, J.; Li, G. Current Concepts on Tenogenic Differentiation and Clinical Applications. J. Orthop. Transl. 2017, 9, 28–42. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.J.; Chen, X.; Li, G.; Chan, K.M.; Heng, B.C.; Yin, Z.; Ouyang, H.W. Concise Review: Stem Cell Fate Guided By Bioactive Molecules for Tendon Regeneration. Stem Cells Transl. Med. 2018, 7, 404–414. [Google Scholar] [CrossRef] [PubMed]
- Gaut, L.; Duprez, D. Tendon Development and Diseases: Tendon Development and Diseases. Wiley Interdiscip. Rev. Dev. Biol. 2016, 5, 5–23. [Google Scholar] [CrossRef] [Green Version]
- Schneider, M.; Angele, P.; Järvinen, T.A.H.; Docheva, D. Rescue Plan for Achilles: Therapeutics Steering the Fate and Functions of Stem Cells in Tendon Wound Healing. Adv. Drug Deliv. Rev. 2018, 129, 352–375. [Google Scholar] [CrossRef]
- Jones, E.R.; Jones, G.C.; Legerlotz, K.; Riley, G.P. Corrigendum to “Cyclical Strain Modulates Metalloprotease and Matrix Gene Expression in Human Tenocytes via Activation of TGFβ” [Biochim. Biophys. Acta (2013) 2596–2607]. Biochim. Biophys. Acta BBA—Mol. Cell Res. 2013, 1833, 3445–3446. [Google Scholar] [CrossRef]
- Ito, Y.; Toriuchi, N.; Yoshitaka, T.; Ueno-Kudoh, H.; Sato, T.; Yokoyama, S.; Nishida, K.; Akimoto, T.; Takahashi, M.; Miyaki, S.; et al. The Mohawk Homeobox Gene Is a Critical Regulator of Tendon Differentiation. Proc. Natl. Acad. Sci. USA 2010, 107, 10538–10542. [Google Scholar] [CrossRef] [Green Version]
- Abate, M.; Gravare-Silbernagel, K.; Siljeholm, C.; Di Iorio, A.; De Amicis, D.; Salini, V.; Werner, S.; Paganelli, R. Pathogenesis of Tendinopathies: Inflammation or Degeneration? Arthritis Res. Ther. 2009, 11, 235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scopelliti, F.; Caterina, C.; Valentina, D.; Gianfranco, C.; Concetta, M.; Andrea, C. Platelet Lysate Converts M (IFNγ+LPS) Macrophages in CD206 + TGF-β + Arginase + M2-like Macrophages That Affect Fibroblast Activity and T Lymphocyte Migration. J. Tissue Eng. Regen. Med. 2021, 15, 788–797. [Google Scholar] [CrossRef]
- Rhee, I. Diverse Macrophages Polarization in Tumor Microenvironment. Arch. Pharm. Res. 2016, 39, 1588–1596. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Cao, Q.; Zheng, D.; Sun, Y.; Wang, C.; Yu, X.; Wang, Y.; Lee, V.W.S.; Zheng, G.; Tan, T.K.; et al. Discrete Functions of M 2a and M 2c Macrophage Subsets Determine Their Relative Efficacy in Treating Chronic Kidney Disease. Kidney Int. 2013, 84, 745–755. [Google Scholar] [CrossRef] [Green Version]
- Wermuth, P.J.; Jimenez, S.A. The Significance of Macrophage Polarization Subtypes for Animal Models of Tissue Fibrosis and Human Fibrotic Diseases. Clin. Transl. Med. 2015, 4, e2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moradi-Chaleshtori, M.; Bandehpour, M.; Heidari, N.; Mohammadi-Yeganeh, S.; Mahmoud Hashemi, S. Exosome-Mediated MiR-33 Transfer Induces M1 Polarization in Mouse Macrophages and Exerts Antitumor Effect in 4T1 Breast Cancer Cell Line. Int. Immunopharmacol. 2021, 90, 107198. [Google Scholar] [CrossRef] [PubMed]
- Moradi-Chaleshtori, M.; Shojaei, S.; Mohammadi-Yeganeh, S.; Hashemi, S.M. Transfer of MiRNA in Tumor-Derived Exosomes Suppresses Breast Tumor Cell Invasion and Migration by Inducing M1 Polarization in Macrophages. Life Sci. 2021, 282, 119800. [Google Scholar] [CrossRef]
- Oishi, S.; Takano, R.; Tamura, S.; Tani, S.; Iwaizumi, M.; Hamaya, Y.; Takagaki, K.; Nagata, T.; Seto, S.; Horii, T.; et al. M2 Polarization of Murine Peritoneal Macrophages Induces Regulatory Cytokine Production and Suppresses T-Cell Proliferation. Immunology 2016, 149, 320–328. [Google Scholar] [CrossRef]
- Zhao, H.-Y.; Zhang, Y.-Y.; Xing, T.; Tang, S.-Q.; Wen, Q.; Lyu, Z.-S.; Lv, M.; Wang, Y.; Xu, L.-P.; Zhang, X.-H.; et al. M2 Macrophages, but Not M1 Macrophages, Support Megakaryopoiesis by Upregulating PI3K-AKT Pathway Activity. Signal Transduct. Target. Ther. 2021, 6, 234. [Google Scholar] [CrossRef]
- Stolk, M.; Klatte-Schulz, F.; Schmock, A.; Minkwitz, S.; Wildemann, B.; Seifert, M. New Insights into Tenocyte-Immune Cell Interplay in an in Vitro Model of Inflammation. Sci. Rep. 2017, 7, 9801. [Google Scholar] [CrossRef]
- Akbar, M.; Gilchrist, D.S.; Kitson, S.M.; Nelis, B.; Crowe, L.A.N.; Garcia-Melchor, E.; Reilly, J.H.; Kerr, S.C.; Murrell, G.A.C.; McInnes, I.B.; et al. Targeting Danger Molecules in Tendinopathy: The HMGB1/TLR4 Axis. RMD Open 2017, 3, e000456. [Google Scholar] [CrossRef] [Green Version]
- Lambert, C.; Zappia, J.; Sanchez, C.; Florin, A.; Dubuc, J.-E.; Henrotin, Y. The Damage-Associated Molecular Patterns (DAMPs) as Potential Targets to Treat Osteoarthritis: Perspectives From a Review of the Literature. Front. Med. 2021, 7, 607186. [Google Scholar] [CrossRef]
- Millar, N.L.; Murrell, G.A.C.; McInnes, I.B. Alarmins in Tendinopathy: Unravelling New Mechanisms in a Common Disease. Rheumatology 2013, 52, 769–779. [Google Scholar] [CrossRef] [Green Version]
- Sunwoo, J.Y.; Eliasberg, C.D.; Carballo, C.B.; Rodeo, S.A. The Role of the Macrophage in Tendinopathy and Tendon Healing. J. Orthop. Res. 2020, 38, 1666–1675. [Google Scholar] [CrossRef]
- Tang, C.; Chen, Y.; Huang, J.; Zhao, K.; Chen, X.; Yin, Z.; Heng, B.C.; Chen, W.; Shen, W. The Roles of Inflammatory Mediators and Immunocytes in Tendinopathy. J. Orthop. Transl. 2018, 14, 23–33. [Google Scholar] [CrossRef]
- Klar, A.S.; Michalak-Mićka, K.; Biedermann, T.; Simmen-Meuli, C.; Reichmann, E.; Meuli, M. Characterization of M1 and M2 Polarization of Macrophages in Vascularized Human Dermo-Epidermal Skin Substitutes in Vivo. Pediatr. Surg. Int. 2018, 34, 129–135. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Zhang, S.; Wu, H.; Rong, X.; Guo, J. M2b Macrophage Polarization and Its Roles in Diseases. J. Leukoc. Biol. 2019, 106, 345–358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaggar, A.; Weathington, N. Bioactive Extracellular Matrix Fragments in Lung Health and Disease. J. Clin. Investig. 2016, 126, 3176–3184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, X.; Jackson, P.L.; Tanner, S.; Hardison, M.T.; Abdul Roda, M.; Blalock, J.E.; Gaggar, A. A Self-Propagating Matrix Metalloprotease-9 (MMP-9) Dependent Cycle of Chronic Neutrophilic Inflammation. PLoS ONE 2011, 6, e15781. [Google Scholar] [CrossRef] [PubMed]
- Buckley, C.D.; Pilling, D.; Lord, J.M.; Akbar, A.N.; Scheel-Toellner, D.; Salmon, M. Fibroblasts Regulate the Switch from Acute Resolving to Chronic Persistent Inflammation. Trends Immunol. 2001, 22, 199–204. [Google Scholar] [CrossRef]
- Crowley, T.; O’Neil, J.D.; Adams, H.; Thomas, A.M.; Filer, A.; Buckley, C.D.; Clark, A.R. Priming in Response to Pro-Inflammatory Cytokines Is a Feature of Adult Synovial but Not Dermal Fibroblasts. Arthritis Res. Ther. 2017, 19, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Dakin, S.G.; Buckley, C.D.; Al-Mossawi, M.H.; Hedley, R.; Martinez, F.O.; Wheway, K.; Watkins, B.; Carr, A.J. Persistent Stromal Fibroblast Activation Is Present in Chronic Tendinopathy. Arthritis Res. Ther. 2017, 19, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Dakin, S.G.; Martinez, F.O.; Yapp, C.; Wells, G.; Oppermann, U.; Dean, B.J.F.; Smith, R.D.J.; Wheway, K.; Watkins, B.; Roche, L.; et al. Inflammation Activation and Resolution in Human Tendon Disease. Sci. Transl. Med. 2015, 7, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dakin, S.G.; Ly, L.; Colas, R.A.; Oppermann, U.; Wheway, K.; Watkins, B.; Dalli, J.; Carr, A.J. Increased 15-PGDH Expression Leads to Dysregulated Resolution Responses in Stromal Cells from Patients with Chronic Tendinopathy. Sci. Rep. 2017, 7, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Dakin, S.G.; Colas, R.A.; Wheway, K.; Watkins, B.; Appleton, L.; Rees, J.; Gwilym, S.; Little, C.; Dalli, J.; Carr, A.J. Proresolving Mediators LXB4 and RvE1 Regulate Inflammation in Stromal Cells from Patients with Shoulder Tendon Tears. Am. J. Pathol. 2019, 189, 2258–2268. [Google Scholar] [CrossRef] [Green Version]
- Hart, D.A.; Frank, C.B.; Bray, R.C. Inflammatory Processes in Repetitive Motion and Overuse Syndromes; Potential Role of Neurogenic Mechanisms in Tendons and Ligaments. In Repetitive Motion Disorders of the Upper Extremity; American Academy of Orthopaedic Surgeons: Rosemont, IL, USA, 1995; pp. 247–262. ISBN 0-89203-143-3. [Google Scholar]
- Wernersson, S.; Pejler, G. Mast Cell Secretory Granules: Armed for Battle. Nat. Rev. Immunol. 2014, 14, 478–494. [Google Scholar] [CrossRef]
- Marsolais, D.; Côté, C.H.; Frenette, J. Neutrophils and Macrophages Accumulate Sequentially Following Achilles Tendon Injury. J. Orthop. Res. 2001, 19, 1203–1209. [Google Scholar] [CrossRef]
- Millar, N.L.; Akbar, M.; Campbell, A.L.; Reilly, J.H.; Kerr, S.C.; McLean, M.; Frleta-Gilchrist, M.; Fazzi, U.G.; Leach, W.J.; Rooney, B.P.; et al. IL-17A Mediates Inflammatory and Tissue Remodelling Events in Early Human Tendinopathy. Sci. Rep. 2016, 6, 27149. [Google Scholar] [CrossRef] [PubMed]
- Magnan, B.; Bondi, M.; Pierantoni, S.; Samaila, E. The Pathogenesis of Achilles Tendinopathy: A Systematic Review. Foot Ankle Surg. 2014, 20, 154–159. [Google Scholar] [CrossRef] [PubMed]
- Greve, K.; Domeij-Arverud, E.; Labruto, F.; Edman, G.; Bring, D.; Nilsson, G.; Ackermann, P.W. Metabolic Activity in Early Tendon Repair Can Be Enhanced by Intermittent Pneumatic Compression: IPC Increases Metabolites in Tendon Repair. Scand. J. Med. Sci. Sports 2012, 22, e55–e63. [Google Scholar] [CrossRef]
- Andersson, G.; Backman, L.J.; Scott, A.; Lorentzon, R.; Forsgren, S.; Danielson, P. Substance P Accelerates Hypercellularity and Angiogenesis in Tendon Tissue and Enhances Paratendinitis in Response to Achilles Tendon Overuse in a Tendinopathy Model. Br. J. Sports Med. 2011, 45, 1017–1022. [Google Scholar] [CrossRef] [Green Version]
- Ackermann, P.W. Neuronal Regulation of Tendon Homoeostasis. Int. J. Exp. Pathol. 2013, 94, 271–286. [Google Scholar] [CrossRef]
- Dakin, S.G.; Dudhia, J.; Smith, R.K.W. Resolving an Inflammatory Concept: The Importance of Inflammation and Resolution in Tendinopathy. Vet. Immunol. Immunopathol. 2014, 158, 121–127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Costa-Almeida, R.; Calejo, I.; Gomes, M.E. Mesenchymal Stem Cells Empowering Tendon Regenerative Therapies. Int. J. Mol. Sci. 2019, 20, 3002. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dakin, S.G.; Werling, D.; Hibbert, A.; Abayasekara, D.R.E.; Young, N.J.; Smith, R.K.W.; Dudhia, J. Macrophage Sub-Populations and the Lipoxin A4 Receptor Implicate Active Inflammation during Equine Tendon Repair. PLoS ONE 2012, 7, e32333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Sadi, O.; Schulze-Tanzil, G.; Kohl, B.; Lohan, A.; Lemke, M.; Ertel, W.; John, T. Tenocytes, pro-Inflammatory Cytokines and Leukocytes: A Relationship? Muscles Ligaments Tendons J. 2012, 1, 68–76. [Google Scholar] [PubMed]
- Scott, A. Neuropeptides in Tendinopathy. Front. Biosci. 2009, 26, 2203–2211. [Google Scholar] [CrossRef] [Green Version]
- Rees, J.D.; Stride, M.; Scott, A. Tendons—Time to Revisit Inflammation. Br. J. Sports Med. 2014, 48, 1553–1557. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Melchor, E.; Cafaro, G.; MacDonald, L.; Crowe, L.A.N.; Sood, S.; McLean, M.; Fazzi, U.G.; McInnes, I.B.; Akbar, M.; Millar, N.L. Novel Self-Amplificatory Loop between T Cells and Tenocytes as a Driver of Chronicity in Tendon Disease. Ann. Rheum. Dis. 2021, 80, 1075–1085. [Google Scholar] [CrossRef]
- Matthews, T.J.W.; Hand, G.C.; Rees, J.L.; Athanasou, N.A.; Carr, A.J. Pathology of the Torn Rotator Cuff Tendon: Reduction in Potential for Repair as Tear Size Increases. J. Bone Joint Surg. Br. 2006, 88-B, 489–495. [Google Scholar] [CrossRef]
- Noah, A.C.; Li, T.M.; Martinez, L.M.; Wada, S.; Swanson, J.B.; Disser, N.P.; Sugg, K.B.; Rodeo, S.A.; Lu, T.T.; Mendias, C.L. Adaptive and Innate Immune Cell Responses in Tendons and Lymph Nodes after Tendon Injury and Repair. J. Appl. Physiol. 2020, 128, 473–482. [Google Scholar] [CrossRef]
- Morita, W.; Dakin, S.G.; Snelling, S.J.B.; Carr, A.J. Cytokines in Tendon Disease: A Systematic Review. Bone Jt. Res. 2017, 6, 656–664. [Google Scholar] [CrossRef] [Green Version]
- Mobasheri, A.; Shakibaei, M. Is Tendinitis an Inflammatory Disease Initiated and Driven by Pro-Inflammatory Cytokines Such as Interleukin 1ß? Histol. Histopathol. 2013, 28, 955–964. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Castejon, G.; Brough, D. Understanding the Mechanism of IL-1β Secretion. Cytokine Growth Factor Rev. 2011, 22, 189–195. [Google Scholar] [CrossRef]
- Ziegler, S.F.; Ramsdell, F.; Alderson, M.R. The Activation Antigen CD69. Stem Cells 1994, 12, 456–465. [Google Scholar] [CrossRef] [PubMed]
- Srenathan, U.; Steel, K.; Taams, L.S. IL-17+ CD8+ T Cells: Differentiation, Phenotype and Role in Inflammatory Disease. Immunol. Lett. 2016, 178, 20–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taylor, P.R.; Roy, S.; Leal, S.M.; Sun, Y.; Howell, S.J.; Cobb, B.A.; Li, X.; Pearlman, E. Activation of Neutrophils by Autocrine IL-17A–IL-17RC Interactions during Fungal Infection Is Regulated by IL-6, IL-23, RORγt and Dectin-2. Nat. Immunol. 2014, 15, 143–151. [Google Scholar] [CrossRef] [Green Version]
- Lin, A.M.; Rubin, C.J.; Khandpur, R.; Wang, J.Y.; Riblett, M.; Yalavarthi, S.; Villanueva, E.C.; Shah, P.; Kaplan, M.J.; Bruce, A.T. Mast Cells and Neutrophils Release IL-17 through Extracellular Trap Formation in Psoriasis. J. Immunol. 2011, 187, 490–500. [Google Scholar] [CrossRef] [Green Version]
- Song, C.; Luo, L.; Lei, Z.; Li, B.; Liang, Z.; Liu, G.; Li, D.; Zhang, G.; Huang, B.; Feng, Z.-H. IL-17-Producing Alveolar Macrophages Mediate Allergic Lung Inflammation Related to Asthma. J. Immunol. 2008, 181, 6117–6124. [Google Scholar] [CrossRef] [Green Version]
- Fossiez, F.; Djossou, O.; Chomarat, P.; Flores-Romo, L.; Ait-Yahia, S.; Maat, C.; Pin, J.J.; Garrone, P.; Garcia, E.; Saeland, S.; et al. T Cell Interleukin-17 Induces Stromal Cells to Produce Proinflammatory and Hematopoietic Cytokines. J. Exp. Med. 1996, 183, 2593–2603. [Google Scholar] [CrossRef] [Green Version]
- Lu, H.-L.; Huang, X.-Y.; Luo, Y.-F.; Tan, W.-P.; Chen, P.-F.; Guo, Y.-B. Activation of M1 Macrophages Plays a Critical Role in the Initiation of Acute Lung Injury. Biosci. Rep. 2018, 13, BSR20171555. [Google Scholar] [CrossRef] [Green Version]
- John, T.; Lodka, D.; Kohl, B.; Ertel, W.; Jammrath, J.; Conrad, C.; Stoll, C.; Busch, C.; Schulze-Tanzil, G. Effect of Pro-Inflammatory and Immunoregulatory Cytokines on Human Tenocytes. J. Orthop. Res. 2010, 28, 1071–1077. [Google Scholar] [CrossRef]
- Pan, H.; Halper, J. Regulation of Heat Shock Protein 47 and Type I Procollagen Expression in Avian Tendon Cells. Cell Tissue Res. 2003, 311, 373–382. [Google Scholar] [CrossRef]
- Behzad, H.; Sharma, A.; Mousavizadeh, R.; Lu, A.; Scott, A. Mast Cells Exert Pro-Inflammatory Effects of Relevance to the Pathophyisology of Tendinopathy. Arthritis Res. Ther. 2013, 15, R184. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Behzad, H.; Tsai, S.-H.; Nassab, P.; Mousavizadeh, R.; McCormack, R.G.; Scott, A. Mechanism of Mast Cell Adhesion to Human Tenocytes in Vitro. J. Orthop. Res. 2015, 33, 9–16. [Google Scholar] [CrossRef]
- Riquet, F.B.; Lai, W.-F.T.; Birkhead, J.R.; Suen, L.-F.; Karsenty, G.; Goldring, M.B. Suppression of Type I Collagen Gene Expression by Prostaglandins in Fibroblasts Is Mediated at the Transcriptional Level. Mol. Med. 2000, 6, 705–719. [Google Scholar] [CrossRef] [Green Version]
- Rőszer, T. Understanding the Mysterious M2 Macrophage through Activation Markers and Effector Mechanisms. Mediators Inflamm. 2015, 2015, 816460. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, W.C.; Woods, V.M.; Chitko-McKown, C.G.; Hash, S.M.; Rice-Ficht, A.C. Interleukin-10 Is Expressed by Bovine Type 1 Helper, Type 2 Helper, and Unrestricted Parasite-Specific T-Cell Clones and Inhibits Proliferation of All Three Subsets in an Accessory-Cell-Dependent Manner. Infect. Immun. 1994, 62, 4697–4708. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reitamo, S.; Remitz, A.; Tamai, K.; Ledo, I.; Uitto, J. Interleukin 10 Up-Regulates Elastin Gene Expression In Vivo and In Vitro at the Transcriptional Level. Biochem. J. 1994, 302, 331–333. [Google Scholar] [CrossRef] [Green Version]
- Kishimoto, T.; Akira, S.; Narazaki, M.; Taga, T. Interleukin-6 Family of Cytokines and Gp130. Blood 1995, 86, 1243–1254. [Google Scholar] [CrossRef] [Green Version]
- Ropelle, E.R.; Flores, M.B.; Cintra, D.E.; Rocha, G.Z.; Pauli, J.R.; Morari, J.; de Souza, C.T.; Moraes, J.C.; Prada, P.O.; Guadagnini, D.; et al. IL-6 and IL-10 Anti-Inflammatory Activity Links Exercise to Hypothalamic Insulin and Leptin Sensitivity through IKKβ and ER Stress Inhibition. PLoS Biol. 2010, 8, e1000465. [Google Scholar] [CrossRef]
- Lin, T.W.; Cardenas, L.; Glaser, D.L.; Soslowsky, L.J. Tendon Healing in Interleukin-4 and Interleukin-6 Knockout Mice. J. Biomech. 2006, 39, 61–69. [Google Scholar] [CrossRef] [PubMed]
- Jelinsky, S.A.; Rodeo, S.A.; Li, J.; Gulotta, L.V.; Archambault, J.M.; Seeherman, H.J. Regulation of Gene Expression in Human Tendinopathy. BMC Musculoskelet. Disord. 2011, 12, 86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsuzaki, M.; Guyton, G.; Garrett, W.; Archambault, J.M.; Herzog, W.; Almekinders, L.; Bynum, D.; Yang, X.; Banes, A.J. IL-1β Induces COX2, MMP-1, -3 and -13, ADAMTS-4, IL-1β and IL-6 in Human Tendon Cells. J. Orthop. Res. 2003, 21, 256–264. [Google Scholar] [CrossRef]
- De Castro, L.L.; Lopes-Pacheco, M.; Weiss, D.J.; Cruz, F.F.; Rocco, P.R.M. Current Understanding of the Immunosuppressive Properties of Mesenchymal Stromal Cells. J. Mol. Med. Berl. Ger. 2019, 97, 605–618. [Google Scholar] [CrossRef]
- Burk, J. Mechanisms of Action of Multipotent Mesenchymal Stromal Cells in Tendon Disease. In Tendons; Sözen, H., Ed.; IntechOpen: London, UK, 2019; ISBN 978-1-83962-985-3. [Google Scholar]
- Barboni, B.; Russo, V.; Gatta, V.; Bernabò, N.; Berardinelli, P.; Mauro, A.; Martelli, A.; Valbonetti, L.; Muttini, A.; Di Giacinto, O.; et al. Therapeutic Potential of HAECs for Early Achilles Tendon Defect Repair through Regeneration. J. Tissue Eng. Regen. Med. 2018, 12, e1594–e1608. [Google Scholar] [CrossRef] [PubMed]
- Manuelpillai, U.; Lourensz, D.; Vaghjiani, V.; Tchongue, J.; Lacey, D.; Tee, J.-Y.; Murthi, P.; Chan, J.; Hodge, A.; Sievert, W. Human Amniotic Epithelial Cell Transplantation Induces Markers of Alternative Macrophage Activation and Reduces Established Hepatic Fibrosis. PLoS ONE 2012, 7, e38631. [Google Scholar] [CrossRef] [Green Version]
- Miki, T. A Rational Strategy for the Use of Amniotic Epithelial Stem Cell Therapy for Liver Diseases. Stem Cells Transl. Med. 2016, 5, 405–409. [Google Scholar] [CrossRef] [Green Version]
- Miki, T. Amnion-Derived Stem Cells: In Quest of Clinical Applications. Stem Cell Res. Ther. 2011, 2, 25. [Google Scholar] [CrossRef] [Green Version]
- Liu, Q.-W.; Huang, Q.-M.; Wu, H.-Y.; Zuo, G.-S.-L.; Gu, H.-C.; Deng, K.-Y.; Xin, H.-B. Characteristics and Therapeutic Potential of Human Amnion-Derived Stem Cells. Int. J. Mol. Sci. 2021, 22, 970. [Google Scholar] [CrossRef]
- Lange-Consiglio, A.; Lazzari, B.; Perrini, C.; Pizzi, F.; Stella, A.; Cremonesi, F.; Capra, E. MicroRNAs of Equine Amniotic Mesenchymal Cell-Derived Microvesicles and Their Involvement in Anti-Inflammatory Processes. Cell Transplant. 2018, 27, 45–54. [Google Scholar] [CrossRef] [Green Version]
- Manning, C.N.; Martel, C.; Sakiyama-Elbert, S.E.; Silva, M.J.; Shah, S.; Gelberman, R.H.; Thomopoulos, S. Adipose-Derived Mesenchymal Stromal Cells Modulate Tendon Fibroblast Responses to Macrophage-Induced Inflammation in Vitro. Stem Cell Res. Ther. 2015, 6, 74. [Google Scholar] [CrossRef] [Green Version]
- Viganò, M.; Lugano, G.; Perucca Orfei, C.; Menon, A.; Ragni, E.; Colombini, A.; De Luca, P.; Randelli, P.; de Girolamo, L. Autologous Microfragmented Adipose Tissue Reduces the Catabolic and Fibrosis Response in an In Vitro Model of Tendon Cell Inflammation. Stem Cells Int. 2019, 2019, 5620286. [Google Scholar] [CrossRef] [PubMed]
- Lange-Consiglio, A.; Rossi, D.; Tassan, S.; Perego, R.; Cremonesi, F.; Parolini, O. Conditioned Medium from Horse Amniotic Membrane-Derived Multipotent Progenitor Cells: Immunomodulatory Activity In Vitro and First Clinical Application in Tendon and Ligament Injuries In Vivo. Stem Cells Dev. 2013, 22, 3015–3024. [Google Scholar] [CrossRef]
- Lange-Consiglio, A.; Perrini, C.; Tasquier, R.; Deregibus, M.C.; Camussi, G.; Pascucci, L.; Marini, M.G.; Corradetti, B.; Bizzaro, D.; De Vita, B.; et al. Equine Amniotic Microvesicles and Their Anti-Inflammatory Potential in a Tenocyte Model In Vitro. Stem Cells Dev. 2016, 25, 610–621. [Google Scholar] [CrossRef] [Green Version]
- Berthiaume, F.; Maguire, T.J.; Yarmush, M.L. Tissue Engineering and Regenerative Medicine: History, Progress, and Challenges. Annu. Rev. Chem. Biomol. Eng. 2011, 2, 403–430. [Google Scholar] [CrossRef]
- Buschmann, J.; Yamada, Y.; Schulz-Schönhagen, K.; Hess, S.C.; Stark, W.J.; Opelz, C.; Bürgisser, G.M.; Weder, W.; Jungraithmayr, W. Hybrid Nanocomposite as a Chest Wall Graft with Improved Integration by Adipose-Derived Stem Cells. Sci. Rep. 2019, 9, 10910. [Google Scholar] [CrossRef]
- Gaspar, D.; Spanoudes, K.; Holladay, C.; Pandit, A.; Zeugolis, D. Progress in Cell-Based Therapies for Tendon Repair. Adv. Drug Deliv. Rev. 2015, 84, 240–256. [Google Scholar] [CrossRef]
- Gonçalves, A.I.; Costa-Almeida, R.; Gershovich, P.; Rodrigues, M.T.; Reis, R.L.; Gomes, M.E. Cell-Based Approaches for Tendon Regeneration. In Tendon Regeneration; Elsevier: Amsterdam, The Netherlands, 2015; ISBN 978-0-12-801590-2. [Google Scholar]
- Kaux, J.-F.; Crielaard, J.-M. Platelet-Rich Plasma Application in the Management of Chronic Tendinopathies. Acta Orthop. Belg. 2013, 79, 10–15. [Google Scholar]
- Filardo, G.; Di Matteo, B.; Kon, E.; Merli, G.; Marcacci, M. Platelet-Rich Plasma in Tendon-Related Disorders: Results and Indications. Knee Surg. Sports Traumatol. Arthrosc. 2018, 26, 1984–1999. [Google Scholar] [CrossRef] [PubMed]
- Tang, J.B.; Zhou, Y.L.; Wu, Y.F.; Liu, P.Y.; Wang, X.T. Gene Therapy Strategies to Improve Strength and Quality of Flexor Tendon Healing. Expert Opin. Biol. Ther. 2016, 16, 291–301. [Google Scholar] [CrossRef] [PubMed]
- Freedman, B.R.; Mooney, D.J. Biomaterials to Mimic and Heal Connective Tissues. Adv. Mater. 2019, 31, e1806695. [Google Scholar] [CrossRef]
- Lin, J.; Zhou, W.; Han, S.; Bunpetch, V.; Zhao, K.; Liu, C.; Yin, Z.; Ouyang, H. Cell-Material Interactions in Tendon Tissue Engineering. Acta Biomater. 2018, 70, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Lim, W.L.; Liau, L.L.; Ng, M.H.; Chowdhury, S.R.; Law, J.X. Current Progress in Tendon and Ligament Tissue Engineering. Tissue Eng. Regen. Med. 2019, 16, 549–571. [Google Scholar] [CrossRef]
- Lui, P.P. Stem Cell Technology for Tendon Regeneration: Current Status, Challenges, and Future Research Directions. Stem Cells Cloning Adv. Appl. 2015, 2015, 163–174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galatz, L.M.; Gerstenfeld, L.; Heber-Katz, E.; Rodeo, S.A. Tendon Regeneration and Scar Formation: The Concept of Scarless Healing. J. Orthop. Res. 2015, 33, 823–831. [Google Scholar] [CrossRef]
- Im, G.-I.; Kim, T.-K. Stem Cells for the Regeneration of Tendon and Ligament: A Perspective. Int. J. Stem Cells 2020, 13, 335–341. [Google Scholar] [CrossRef] [PubMed]
- Migliorini, F.; Tingart, M.; Maffulli, N. Progress with Stem Cell Therapies for Tendon Tissue Regeneration. Expert Opin. Biol. Ther. 2020, 20, 1373–1379. [Google Scholar] [CrossRef]
- Hosaka, Y.; Kirisawa, R.; Yamamoto, E.; Ueda, H.; Iwai, H.; Takehana, K. Localization of Cytokines in Tendinocytes of the Superficial Digital Flexor Tendon in the Horse. J. Vet. Med. Sci. 2002, 64, 945–947. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clegg, P.D.; Strassburg, S.; Smith, R.K. Cell Phenotypic Variation in Normal and Damaged Tendons. Int. J. Exp. Pathol. 2007, 88, 227–235. [Google Scholar] [CrossRef]
- Han, Y.; Li, X.; Zhang, Y.; Han, Y.; Chang, F.; Ding, J. Mesenchymal Stem Cells for Regenerative Medicine. Cells 2019, 8, 886. [Google Scholar] [CrossRef] [Green Version]
- Cao, W.; Cao, K.; Cao, J.; Wang, Y.; Shi, Y. Mesenchymal Stem Cells and Adaptive Immune Responses. Immunol. Lett. 2015, 168, 147–153. [Google Scholar] [CrossRef]
- Davies, L.C.; Boberg, E.; Le Blanc, K. Commentary: Role of Mesenchymal Stromal Cell–Mediated Crosstalk with Macrophages in Graft-versus-Host Disease and Tissue Repair. Biol. Blood Marrow Transplant. 2017, 23, 861–862. [Google Scholar] [CrossRef] [Green Version]
- Cho, D.-I.; Kim, M.R.; Jeong, H.; Jeong, H.C.; Jeong, M.H.; Yoon, S.H.; Kim, Y.S.; Ahn, Y. Mesenchymal Stem Cells Reciprocally Regulate the M1/M2 Balance in Mouse Bone Marrow-Derived Macrophages. Exp. Mol. Med. 2014, 46, e70. [Google Scholar] [CrossRef]
- Geng, Y.; Zhang, L.; Fu, B.; Zhang, J.; Hong, Q.; Hu, J.; Li, D.; Luo, C.; Cui, S.; Zhu, F.; et al. Mesenchymal Stem Cells Ameliorate Rhabdomyolysis-Induced Acute Kidney Injury via the Activation of M2 Macrophages. Stem Cell Res. Ther. 2014, 5, 80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, L.; Tredget, E.E.; Wu, P.Y.G.; Wu, Y. Paracrine Factors of Mesenchymal Stem Cells Recruit Macrophages and Endothelial Lineage Cells and Enhance Wound Healing. PLoS ONE 2008, 3, e1886. [Google Scholar] [CrossRef] [Green Version]
- Scher, J.U.; Pillinger, M.H. The Anti-Inflammatory Effects of Prostaglandins. J. Investig. Med. 2009, 57, 703–708. [Google Scholar] [CrossRef]
- Vasandan, A.B.; Jahnavi, S.; Shashank, C.; Prasad, P.; Kumar, A.; Prasanna, S.J. Human Mesenchymal Stem Cells Program Macrophage Plasticity by Altering Their Metabolic Status via a PGE2-Dependent Mechanism. Sci. Rep. 2016, 6, 38308. [Google Scholar] [CrossRef] [Green Version]
- Taylor, A.; Verhagen, J.; Blaser, K.; Akdis, M.; Akdis, C.A. Mechanisms of Immune Suppression by Interleukin-10 and Transforming Growth Factor-Beta: The Role of T Regulatory Cells. Immunology 2006, 117, 433–442. [Google Scholar] [CrossRef] [PubMed]
- Ackermann, P.W.; Hart, D.A. (Eds.) Metabolic Influences on Risk for Tendon Disorders; Advances in Experimental Medicine and Biology; Springer International Publishing: Cham, Schwitzerland, 2016; Volume 920, ISBN 978-3-319-33941-2. [Google Scholar]
- Digiacomo, G.; Ziche, M.; Dello Sbarba, P.; Donnini, S.; Rovida, E. Prostaglandin E2 Transactivates the Colony-stimulating Factor-1 Receptor and Synergizes with Colony-stimulating Factor-1 in the Induction of Macrophage Migration via the Mitogen-activated Protein Kinase ERK1/2. FASEB J. 2015, 29, 2545–2554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rossi, D.; Pianta, S.; Magatti, M.; Sedlmayr, P.; Parolini, O. Characterization of the Conditioned Medium from Amniotic Membrane Cells: Prostaglandins as Key Effectors of Its Immunomodulatory Activity. PLoS ONE 2012, 7, e46956. [Google Scholar] [CrossRef]
- Lee, K.-C.; Lin, H.-C.; Huang, Y.-H.; Hung, S.-C. Allo-Transplantation of Mesenchymal Stem Cells Attenuates Hepatic Injury through IL1Ra Dependent Macrophage Switch in a Mouse Model of Liver Disease. J. Hepatol. 2015, 63, 1405–1412. [Google Scholar] [CrossRef]
- Wang, X.; Gu, H.; Qin, D.; Yang, L.; Huang, W.; Essandoh, K.; Wang, Y.; Caldwell, C.C.; Peng, T.; Zingarelli, B.; et al. Exosomal MiR-223 Contributes to Mesenchymal Stem Cell-Elicited Cardioprotection in Polymicrobial Sepsis. Sci. Rep. 2015, 5, 13721. [Google Scholar] [CrossRef] [Green Version]
- Shen, B.; Liu, J.; Zhang, F.; Wang, Y.; Qin, Y.; Zhou, Z.; Qiu, J.; Fan, Y. CCR2 Positive Exosome Released by Mesenchymal Stem Cells Suppresses Macrophage Functions and Alleviates Ischemia/Reperfusion-Induced Renal Injury. Stem Cells Int. 2016, 2016, 1240301. [Google Scholar] [CrossRef] [Green Version]
- Watanabe, Y.; Tsuchiya, A.; Seino, S.; Kawata, Y.; Kojima, Y.; Ikarashi, S.; Starkey Lewis, P.J.; Lu, W.-Y.; Kikuta, J.; Kawai, H.; et al. Mesenchymal Stem Cells and Induced Bone Marrow-Derived Macrophages Synergistically Improve Liver Fibrosis in Mice. Stem Cells Transl. Med. 2019, 8, 271–284. [Google Scholar] [CrossRef] [Green Version]
- English, K. Mechanisms of Mesenchymal Stromal Cell Immunomodulation. Immunol. Cell Biol. 2013, 91, 19–26. [Google Scholar] [CrossRef] [Green Version]
- Chiossone, L.; Conte, R.; Spaggiari, G.M.; Serra, M.; Romei, C.; Bellora, F.; Becchetti, F.; Andaloro, A.; Moretta, L.; Bottino, C. Mesenchymal Stromal Cells Induce Peculiar Alternatively Activated Macrophages Capable of Dampening Both Innate and Adaptive Immune Responses: MSC-Induced Macrophages Inhibit NK and T Cells. Stem Cells 2016, 34, 1909–1921. [Google Scholar] [CrossRef] [Green Version]
- Su, J.; Chen, X.; Huang, Y.; Li, W.; Li, J.; Cao, K.; Cao, G.; Zhang, L.; Li, F.; Roberts, A.I.; et al. Phylogenetic Distinction of INOS and IDO Function in Mesenchymal Stem Cell-Mediated Immunosuppression in Mammalian Species. Cell Death Differ. 2014, 21, 388–396. [Google Scholar] [CrossRef] [Green Version]
- Chabannes, D.; Hill, M.; Merieau, E.; Rossignol, J.; Brion, R.; Soulillou, J.P.; Anegon, I.; Cuturi, M.C. A Role for Heme Oxygenase-1 in the Immunosuppressive Effect of Adult Rat and Human Mesenchymal Stem Cells. Blood 2007, 110, 3691–3694. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akiyama, K.; Chen, C.; Wang, D.; Xu, X.; Qu, C.; Yamaza, T.; Cai, T.; Chen, W.; Sun, L.; Shi, S. Mesenchymal-Stem-Cell-Induced Immunoregulation Involves FAS-Ligand-/FAS-Mediated T Cell Apoptosis. Cell Stem Cell 2012, 10, 544–555. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, R.H.; Yoon, N.; Reneau, J.C.; Prockop, D.J. Preactivation of Human MSCs with TNF-α Enhances Tumor-Suppressive Activity. Cell Stem Cell 2012, 11, 825–835. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Del Papa, B.; Sportoletti, P.; Cecchini, D.; Rosati, E.; Balucani, C.; Baldoni, S.; Fettucciari, K.; Marconi, P.; Martelli, M.F.; Falzetti, F.; et al. Notch1 Modulates Mesenchymal Stem Cells Mediated Regulatory T-Cell Induction. Eur. J. Immunol. 2013, 43, 182–187. [Google Scholar] [CrossRef]
- Lever, M.; Maini, P.K.; van der Merwe, P.A.; Dushek, O. Phenotypic Models of T Cell Activation. Nat. Rev. Immunol. 2014, 14, 619–629. [Google Scholar] [CrossRef] [Green Version]
- Le Blanc, K.; Mougiakakos, D. Multipotent Mesenchymal Stromal Cells and the Innate Immune System. Nat. Rev. Immunol. 2012, 12, 383–396. [Google Scholar] [CrossRef] [PubMed]
- Le Blanc, K.; Davies, L.C. Mesenchymal Stromal Cells and the Innate Immune Response. Immunol. Lett. 2015, 168, 140–146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ouyang, L.; Cao, J.; Dai, Q.; Qiu, D. New Insight of Immuno-Engineering in Osteoimmunomodulation for Bone Regeneration. Regen. Ther. 2021, 18, 24–29. [Google Scholar] [CrossRef]
- Liu, J.; Feng, B.; Xu, Y.; Zhu, J.; Feng, X.; Chen, W.; Sheng, X.; Shi, X.; Pan, Q.; Yu, J.; et al. Immunomodulatory Effect of Mesenchymal Stem Cells in Chemical-Induced Liver Injury: A High-Dimensional Analysis. Stem Cell Res. Ther. 2019, 10, 262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aktas, E.; Chamberlain, C.S.; Saether, E.E.; Duenwald-Kuehl, S.E.; Kondratko-Mittnacht, J.; Stitgen, M.; Lee, J.S.; Clements, A.E.; Murphy, W.L.; Vanderby, R. Immune Modulation with Primed Mesenchymal Stem Cells Delivered via Biodegradable Scaffold to Repair an Achilles Tendon Segmental Defect. J. Orthop. Res. 2017, 35, 269–280. [Google Scholar] [CrossRef] [PubMed]
- Saether, E.E.; Chamberlain, C.S.; Leiferman, E.M.; Kondratko-Mittnacht, J.R.; Li, W.J.; Brickson, S.L.; Vanderby, R. Enhanced Medial Collateral Ligament Healing Using Mesenchymal Stem Cells: Dosage Effects on Cellular Response and Cytokine Profile. Stem Cell Rev. Rep. 2014, 10, 86–96. [Google Scholar] [CrossRef] [Green Version]
- Rui, Y.-F.; Lui, P.P.Y.; Li, G.; Fu, S.C.; Lee, Y.W.; Chan, K.M. Isolation and Characterization of Multipotent Rat Tendon-Derived Stem Cells. Tissue Eng. Part A 2010, 16, 1549–1558. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Wang, J.H.-C. Characterization of Differential Properties of Rabbit Tendon Stem Cells and Tenocytes. BMC Musculoskelet. Disord. 2010, 11, 10. [Google Scholar] [CrossRef] [Green Version]
- Lovati, A.B.; Corradetti, B.; Lange Consiglio, A.; Recordati, C.; Bonacina, E.; Bizzaro, D.; Cremonesi, F. Characterization and Differentiation of Equine Tendon-Derived Progenitor Cells. J. Biol. Regul. Homeost. Agents 2011, 25, S75–S84. [Google Scholar]
- Guo, J.; Chan, K.-M.; Zhang, J.-F.; Li, G. Tendon-Derived Stem Cells Undergo Spontaneous Tenogenic Differentiation. Exp. Cell Res. 2016, 341, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Mazzocca, A.D.; Chowaniec, D.; McCarthy, M.B.; Beitzel, K.; Cote, M.P.; McKinnon, W.; Arciero, R. In Vitro Changes in Human Tenocyte Cultures Obtained from Proximal Biceps Tendon: Multiple Passages Result in Changes in Routine Cell Markers. Knee Surg. Sports Traumatol. Arthrosc. 2012, 20, 1666–1672. [Google Scholar] [CrossRef] [PubMed]
- Lui, P.P.Y.; Kong, S.K.; Lau, P.M.; Wong, Y.M.; Lee, Y.W.; Tan, C.; Wong, O.T. Immunogenicity and Escape Mechanisms of Allogeneic Tendon-Derived Stem Cells. Tissue Eng. Part A 2014, 20, 3010–3020. [Google Scholar] [CrossRef]
- Shen, W.; Chen, J.; Yin, Z.; Chen, X.; Liu, H.; Heng, B.C.; Chen, W.; Ouyang, H.-W. Allogenous Tendon Stem/Progenitor Cells in Silk Scaffold for Functional Shoulder Repair. Cell Transplant. 2012, 21, 943–958. [Google Scholar] [CrossRef] [Green Version]
- Tarafder, S.; Chen, E.; Jun, Y.; Kao, K.; Sim, K.H.; Back, J.; Lee, F.Y.; Lee, C.H. Tendon Stem/Progenitor Cells Regulate Inflammation in Tendon Healing via JNK and STAT3 Signaling. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2017, 31, 3991–3998. [Google Scholar] [CrossRef] [Green Version]
- Sabat, R.; Grütz, G.; Warszawska, K.; Kirsch, S.; Witte, E.; Wolk, K.; Geginat, J. Biology of Interleukin-10. Cytokine Growth Factor Rev. 2010, 21, 331–344. [Google Scholar] [CrossRef] [Green Version]
- Opal, S.M.; DePalo, V.A. Anti-Inflammatory Cytokines. Chest 2000, 117, 1162–1172. [Google Scholar] [CrossRef] [Green Version]
- Legerlotz, K.; Jones, E.R.; Screen, H.R.C.; Riley, G.P. Increased Expression of IL-6 Family Members in Tendon Pathology. Rheumatol. Oxf. Engl. 2012, 51, 1161–1165. [Google Scholar] [CrossRef] [Green Version]
- Murray, P.J. The Primary Mechanism of the IL-10-Regulated Antiinflammatory Response Is to Selectively Inhibit Transcription. Proc. Natl. Acad. Sci. USA 2005, 102, 8686–8691. [Google Scholar] [CrossRef] [Green Version]
- Staples, K.J.; Smallie, T.; Williams, L.M.; Foey, A.; Burke, B.; Foxwell, B.M.J.; Ziegler-Heitbrock, L. IL-10 Induces IL-10 in Primary Human Monocyte-Derived Macrophages via the Transcription Factor Stat3. J. Immunol. 2007, 178, 4779–4785. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hutchins, A.P.; Diez, D.; Miranda-Saavedra, D. The IL-10/STAT3-Mediated Anti-Inflammatory Response: Recent Developments and Future Challenges. Brief. Funct. Genomics 2013, 12, 489–498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, F.; Murrell, G.A.C.; Wang, M.-X. Oxidative Stress-Induced c-Jun N-Terminal Kinase (JNK) Activation in Tendon Cells Upregulates MMP1 MRNA and Protein Expression. J. Orthop. Res. Off. Publ. Orthop. Res. Soc. 2007, 25, 378–389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muttini, A.; Valbonetti, L.; Abate, M.; Colosimo, A.; Curini, V.; Mauro, A.; Berardinelli, P.; Russo, V.; Cocciolone, D.; Marchisio, M.; et al. Ovine Amniotic Epithelial Cells: In Vitro Characterization and Transplantation into Equine Superficial Digital Flexor Tendon Spontaneous Defects. Res. Vet. Sci. 2013, 94, 158–169. [Google Scholar] [CrossRef] [PubMed]
- Wassmer, C.-H.; Berishvili, E. Immunomodulatory Properties of Amniotic Membrane Derivatives and Their Potential in Regenerative Medicine. Curr. Diab. Rep. 2020, 20, 31. [Google Scholar] [CrossRef]
- Barboni, B.; Russo, V.; Curini, V.; Martelli, A.; Berardinelli, P.; Mauro, A.; Mattioli, M.; Marchisio, M.; Bonassi Signoroni, P.; Parolini, O.; et al. Gestational Stage Affects Amniotic Epithelial Cells Phenotype, Methylation Status, Immunomodulatory and Stemness Properties. Stem Cell Rev. Rep. 2014, 10, 725–741. [Google Scholar] [CrossRef] [Green Version]
- Magatti, M.; Caruso, M.; De Munari, S.; Vertua, E.; De, D.; Manuelpillai, U.; Parolini, O. Human Amniotic Membrane-Derived Mesenchymal and Epithelial Cells Exert Different Effects on Monocyte-Derived Dendritic Cell Differentiation and Function. Cell Transplant. 2015, 24, 1733–1752. [Google Scholar] [CrossRef]
- Wynn, T.; Barron, L. Macrophages: Master Regulators of Inflammation and Fibrosis. Semin. Liver Dis. 2010, 30, 245–257. [Google Scholar] [CrossRef] [Green Version]
- Cargnoni, A.; Gibelli, L.; Tosini, A.; Signoroni, P.B.; Nassuato, C.; Arienti, D.; Lombardi, G.; Albertini, A.; Wengler, G.S.; Parolini, O. Transplantation of Allogeneic and Xenogeneic Placenta-Derived Cells Reduces Bleomycin-Induced Lung Fibrosis. Cell Transplant. 2009, 18, 405–422. [Google Scholar] [CrossRef] [Green Version]
- Abumaree, M.H.; Al Jumah, M.A.; Kalionis, B.; Jawdat, D.; Al Khaldi, A.; Abomaray, F.M.; Fatani, A.S.; Chamley, L.W.; Knawy, B.A. Human Placental Mesenchymal Stem Cells (PMSCs) Play a Role as Immune Suppressive Cells by Shifting Macrophage Differentiation from Inflammatory M1 to Anti-Inflammatory M2 Macrophages. Stem Cell Rev. Rep. 2013, 9, 620–641. [Google Scholar] [CrossRef]
- Tan, J.L.; Chan, S.T.; Wallace, E.M.; Lim, R. Human Amnion Epithelial Cells Mediate Lung Repair by Directly Modulating Macrophage Recruitment and Polarization. Cell Transplant. 2014, 23, 319–328. [Google Scholar] [CrossRef] [Green Version]
- Mosser, D.M.; Edwards, J.P. Exploring the Full Spectrum of Macrophage Activation. Nat. Rev. Immunol. 2008, 8, 958–969. [Google Scholar] [CrossRef]
- Labonte, A.C.; Tosello-Trampont, A.-C.; Hahn, Y.S. The Role of Macrophage Polarization in Infectious and Inflammatory Diseases. Mol. Cells 2014, 37, 275–285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, T.J.; Jones, C.A.; Miles, E.A.; Warner, J.O.; Warner, J.A. Fetal and Neonatal IL-13 Production during Pregnancy and at Birth and Subsequent Development of Atopic Symptoms. J. Allergy Clin. Immunol. 2000, 105, 951–959. [Google Scholar] [CrossRef] [PubMed]
- Bowen, J.M.; Chamley, L.; Keelan, J.A.; Mitchell, M.D. Cytokines of the Placenta and Extra-Placental Membranes: Roles and Regulation during Human Pregnancy and Parturition. Placenta 2002, 23, 257–273. [Google Scholar] [CrossRef] [PubMed]
- Fiorentino, D.F.; Zlotnik, A.; Vieira, P.; Mosmann, T.R.; Howard, M.; Moore, K.W.; O’Garra, A. IL-10 Acts on the Antigen-Presenting Cell to Inhibit Cytokine Production by Th1 Cells. J. Immunol. 1991, 146, 3444–3451. [Google Scholar]
- D’Andrea, A.; Aste-Amezaga, M.; Valiante, N.M.; Ma, X.; Kubin, M.; Trinchieri, G. Interleukin 10 (IL-10) Inhibits Human Lymphocyte Interferon Gamma-Production by Suppressing Natural Killer Cell Stimulatory Factor/IL-12 Synthesis in Accessory Cells. J. Exp. Med. 1993, 178, 1041–1048. [Google Scholar] [CrossRef] [PubMed]
- Hedrich, C.M.; Bream, J.H. Cell Type-Specific Regulation of IL-10 Expression in Inflammation and Disease. Immunol. Res. 2010, 47, 185–206. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Koike-Soko, C.; Sugimoto, J.; Yoshida, T.; Okabe, M.; Nikaido, T. Human Amnion-Derived Stem Cells Have Immunosuppressive Properties on NK Cells and Monocytes. Cell Transplant. 2015, 24, 2065–2076. [Google Scholar] [CrossRef] [Green Version]
- Ariel, A.; Timor, O. Hanging in the Balance: Endogenous Anti-Inflammatory Mechanisms in Tissue Repair and Fibrosis. J. Pathol. 2013, 229, 250–263. [Google Scholar] [CrossRef]
- Mantovani, A.; Biswas, S.K.; Galdiero, M.R.; Sica, A.; Locati, M. Macrophage Plasticity and Polarization in Tissue Repair and Remodelling. J. Pathol. 2013, 229, 176–185. [Google Scholar] [CrossRef]
- Zhou, Y.; Wang, J.H.-C. PRP Treatment Efficacy for Tendinopathy: A Review of Basic Science Studies. BioMed Res. Int. 2016, 2016, 9103792. [Google Scholar] [CrossRef] [Green Version]
- Rhatomy, S.; Prasetyo, T.E.; Setyawan, R.; Soekarno, N.R.; Romaniyanto, F.; Sedjati, A.P.; Sumarwoto, T.; Utomo, D.N.; Suroto, H.; Mahyudin, F.; et al. Prospect of Stem Cells Conditioned Medium (Secretome) in Ligament and Tendon Healing: A Systematic Review. Stem Cells Transl. Med. 2020, 9, 895–902. [Google Scholar] [CrossRef] [Green Version]
- Pinho, A.G.; Cibrão, J.R.; Silva, N.A.; Monteiro, S.; Salgado, A.J. Cell Secretome: Basic Insights and Therapeutic Opportunities for CNS Disorders. Pharmaceuticals 2020, 13, 31. [Google Scholar] [CrossRef] [Green Version]
- Citeroni, M.R.; Mauro, A.; Ciardulli, M.C.; Di Mattia, M.; El Khatib, M.; Russo, V.; Turriani, M.; Santer, M.; Della Porta, G.; Maffulli, N.; et al. Amnion-Derived Teno-Inductive Secretomes: A Novel Approach to Foster Tendon Differentiation and Regeneration in an Ovine Model. Front. Bioeng. Biotechnol. 2021, 9, 649288. [Google Scholar] [CrossRef] [PubMed]
- Lange-Consiglio, A.; Tassan, S.; Corradetti, B.; Meucci, A.; Perego, R.; Bizzaro, D.; Cremonesi, F. Investigating the Efficacy of Amnion-Derived Compared with Bone Marrow–Derived Mesenchymal Stromal Cells in Equine Tendon and Ligament Injuries. Cytotherapy 2013, 15, 1011–1020. [Google Scholar] [CrossRef] [PubMed]
- Plock, J.A.; Schnider, J.T.; Solari, M.G.; Zheng, X.X.; Gorantla, V.S. Perspectives on the Use of Mesenchymal Stem Cells in Vascularized Composite Allotransplantation. Front. Immunol. 2013, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nishio, H.; Saita, Y.; Kobayashi, Y.; Takaku, T.; Fukusato, S.; Uchino, S.; Wakayama, T.; Ikeda, H.; Kaneko, K. Platelet-Rich Plasma Promotes Recruitment of Macrophages in the Process of Tendon Healing. Regen. Ther. 2020, 14, 262–270. [Google Scholar] [CrossRef]
- Lee, B.-C.; Kang, K.-S. Functional Enhancement Strategies for Immunomodulation of Mesenchymal Stem Cells and Their Therapeutic Application. Stem Cell Res. Ther. 2020, 11, 397. [Google Scholar] [CrossRef] [PubMed]
- Seo, Y.; Kang, M.-J.; Kim, H.-S. Strategies to Potentiate Paracrine Therapeutic Efficacy of Mesenchymal Stem Cells in Inflammatory Diseases. Int. J. Mol. Sci. 2021, 22, 3397. [Google Scholar] [CrossRef]
- Meng, X.; Zheng, M.; Yu, M.; Bai, W.; Zuo, L.; Bu, X.; Liu, Y.; Xia, L.; Hu, J.; Liu, L.; et al. Transplantation of CRISPRa System Engineered IL10-Overexpressing Bone Marrow-Derived Mesenchymal Stem Cells for the Treatment of Myocardial Infarction in Diabetic Mice. J. Biol. Eng. 2019, 13, 49. [Google Scholar] [CrossRef] [Green Version]
- Lian, W.-S.; Cheng, W.T.-K.; Cheng, C.-C.; Hsiao, F.S.-H.; Chen, J.-J.; Cheng, C.-F.; Wu, S.-C. In Vivo Therapy of Myocardial Infarction with Mesenchymal Stem Cells Modified with Prostaglandin I Synthase Gene Improves Cardiac Performance in Mice. Life Sci. 2011, 88, 455–464. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.-X.; Zhang, N.; Wang, H.-W.; Gao, P.; Yang, Q.-P.; Wen, Q.-P. CXCR4 Receptor Overexpression in Mesenchymal Stem Cells Facilitates Treatment of Acute Lung Injury in Rats. J. Biol. Chem. 2015, 290, 1994–2006. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meeremans, M.; Van de Walle, G.R.; Van Vlierberghe, S.; De Schauwer, C. The Lack of a Representative Tendinopathy Model Hampers Fundamental Mesenchymal Stem Cell Research. Front. Cell Dev. Biol. 2021, 9, 651164. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| DAMPs | Receptors | Biological Activity | Reference |
|---|---|---|---|
| HMGB1 | RAGE, TLR-2, TLR-4, TLR-9 | ↑ Pro-inflammatory cytokines and stromal cell compartments responses | [3,72] |
| HSPs | TLR-4 | Cytokine and chemokine release and activation of NK cells | [74] |
| S100A8/A9 | TLR-4 | Attraction of T cells, neutrophils, and macrophages | [73] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Russo, V.; El Khatib, M.; Prencipe, G.; Citeroni, M.R.; Faydaver, M.; Mauro, A.; Berardinelli, P.; Cerveró-Varona, A.; Haidar-Montes, A.A.; Turriani, M.; et al. Tendon Immune Regeneration: Insights on the Synergetic Role of Stem and Immune Cells during Tendon Regeneration. Cells 2022, 11, 434. https://doi.org/10.3390/cells11030434
Russo V, El Khatib M, Prencipe G, Citeroni MR, Faydaver M, Mauro A, Berardinelli P, Cerveró-Varona A, Haidar-Montes AA, Turriani M, et al. Tendon Immune Regeneration: Insights on the Synergetic Role of Stem and Immune Cells during Tendon Regeneration. Cells. 2022; 11(3):434. https://doi.org/10.3390/cells11030434
Chicago/Turabian StyleRusso, Valentina, Mohammad El Khatib, Giuseppe Prencipe, Maria Rita Citeroni, Melisa Faydaver, Annunziata Mauro, Paolo Berardinelli, Adrián Cerveró-Varona, Arlette A. Haidar-Montes, Maura Turriani, and et al. 2022. "Tendon Immune Regeneration: Insights on the Synergetic Role of Stem and Immune Cells during Tendon Regeneration" Cells 11, no. 3: 434. https://doi.org/10.3390/cells11030434
APA StyleRusso, V., El Khatib, M., Prencipe, G., Citeroni, M. R., Faydaver, M., Mauro, A., Berardinelli, P., Cerveró-Varona, A., Haidar-Montes, A. A., Turriani, M., Di Giacinto, O., Raspa, M., Scavizzi, F., Bonaventura, F., Stöckl, J., & Barboni, B. (2022). Tendon Immune Regeneration: Insights on the Synergetic Role of Stem and Immune Cells during Tendon Regeneration. Cells, 11(3), 434. https://doi.org/10.3390/cells11030434