
Reproductive Toxicity of Furfural Acetone in Meloidogyne incognita and Caenorhabditis elegans



Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemical
2.2. Nematodes
2.3. Pot Experiment
2.4. Effect of FAc on Nematode Reproduction
2.5. Nematode Germ Cells Determination
2.6. Nematode Gonadal Development Assay
2.7. Apoptosis Assays
2.8. Quantitative Real-Time PCR
2.9. Statistical Analysis
3. Results
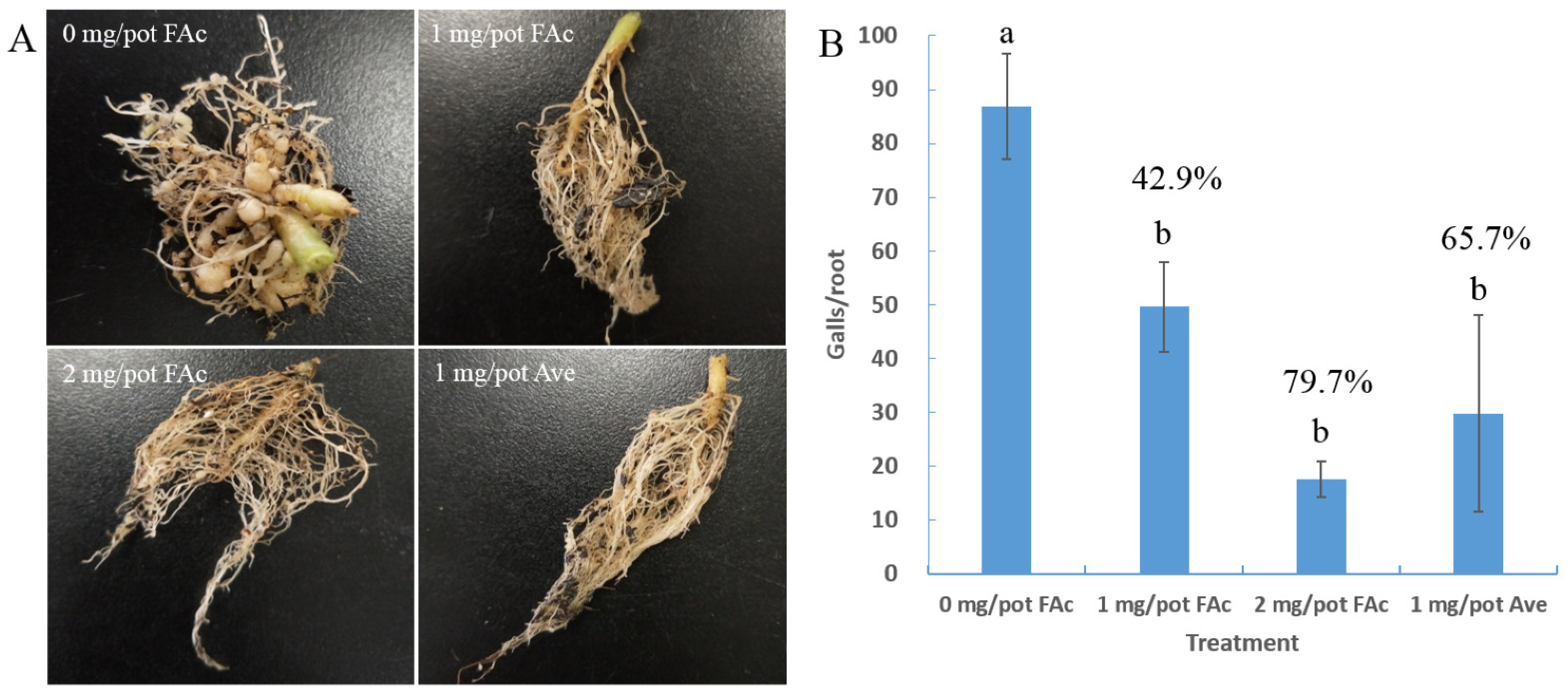
3.1. Efficiency of FAc against M. incognita in Pots
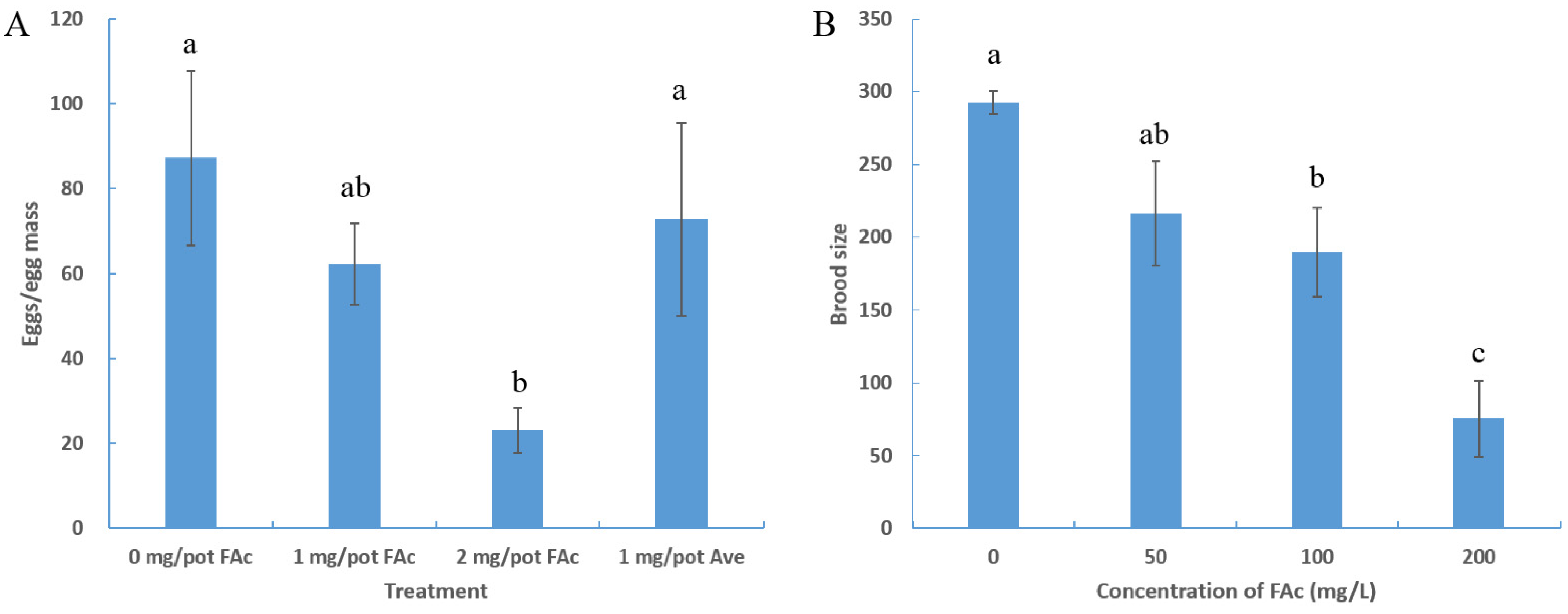
3.2. FAc Reduced the Brood Size of Nematode
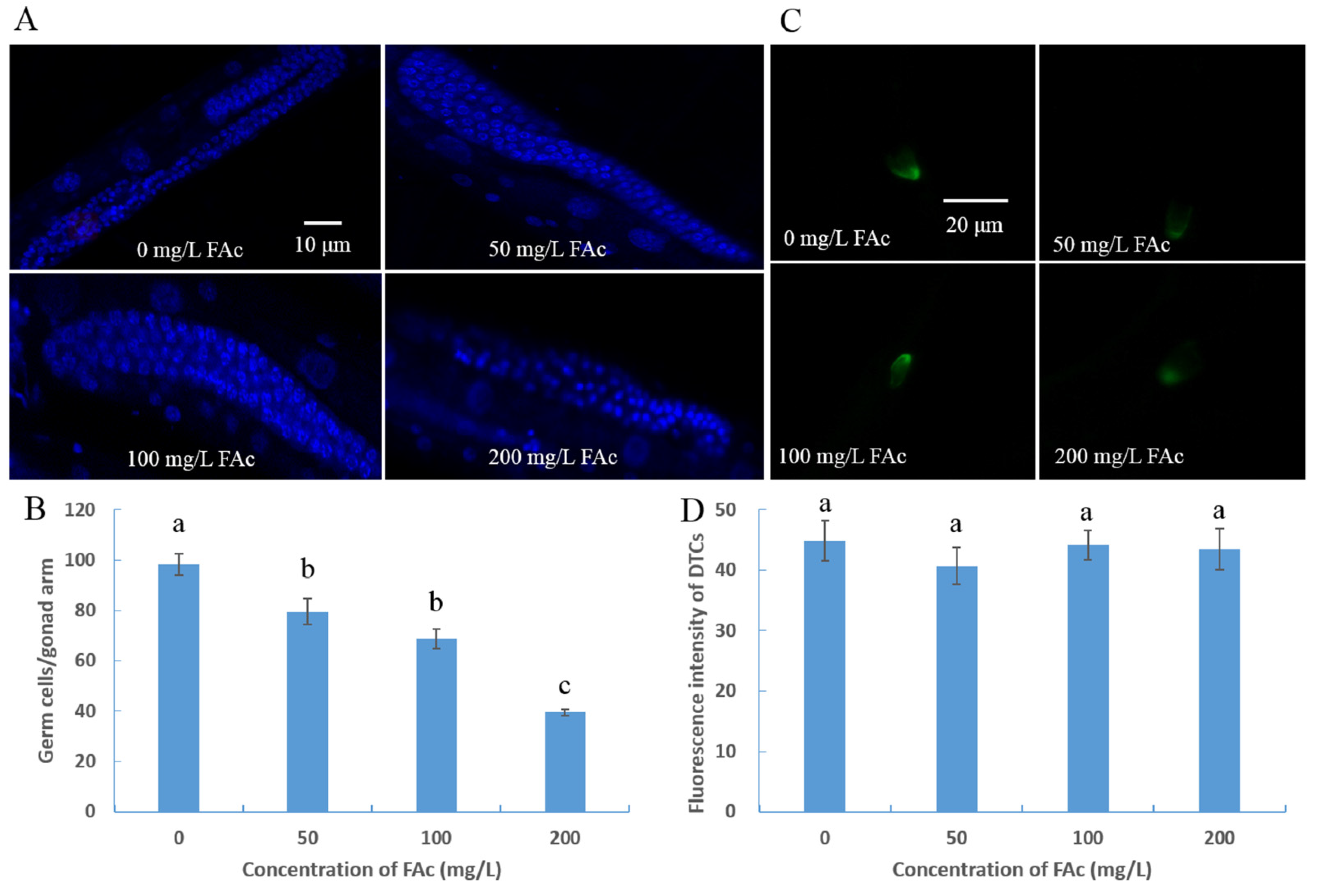
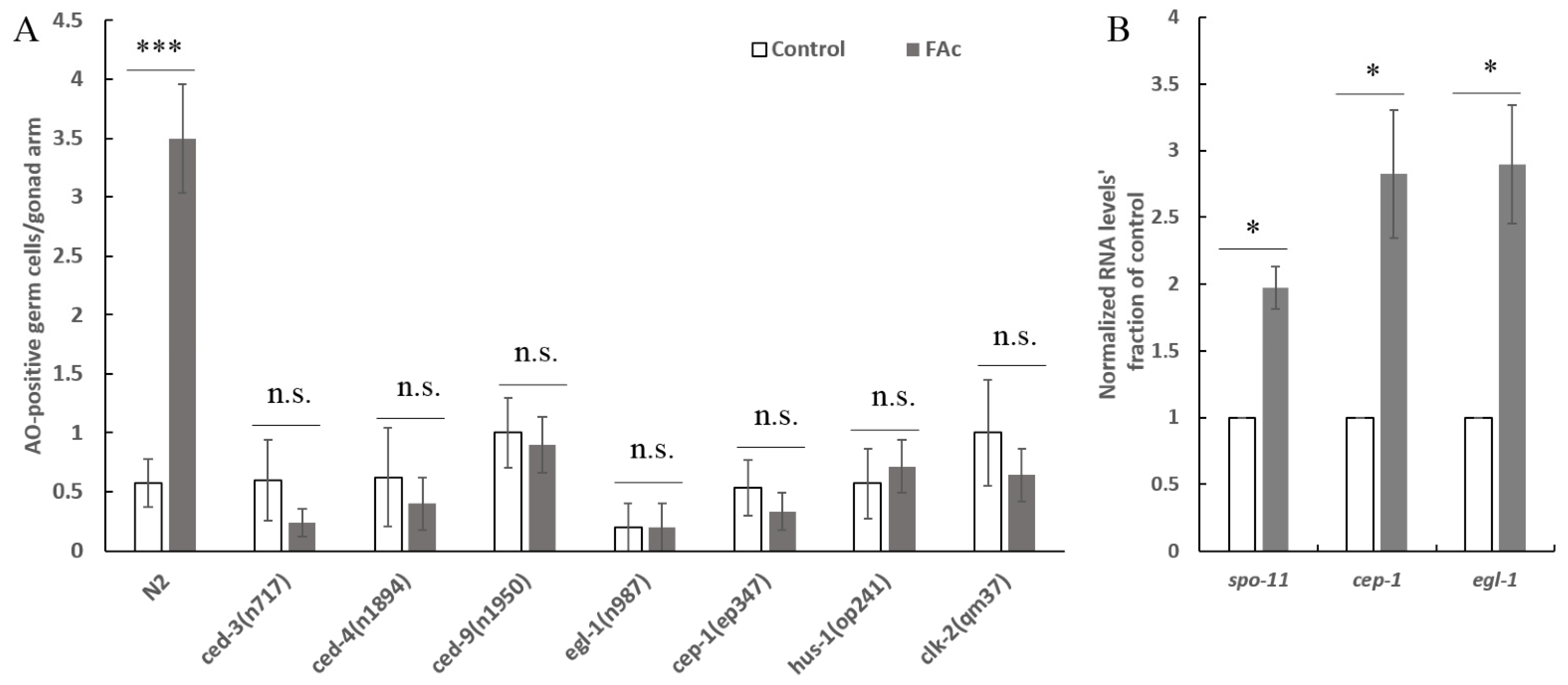
3.3. FAc Exposure Induced Germ-Cell Reduction of C. elegans
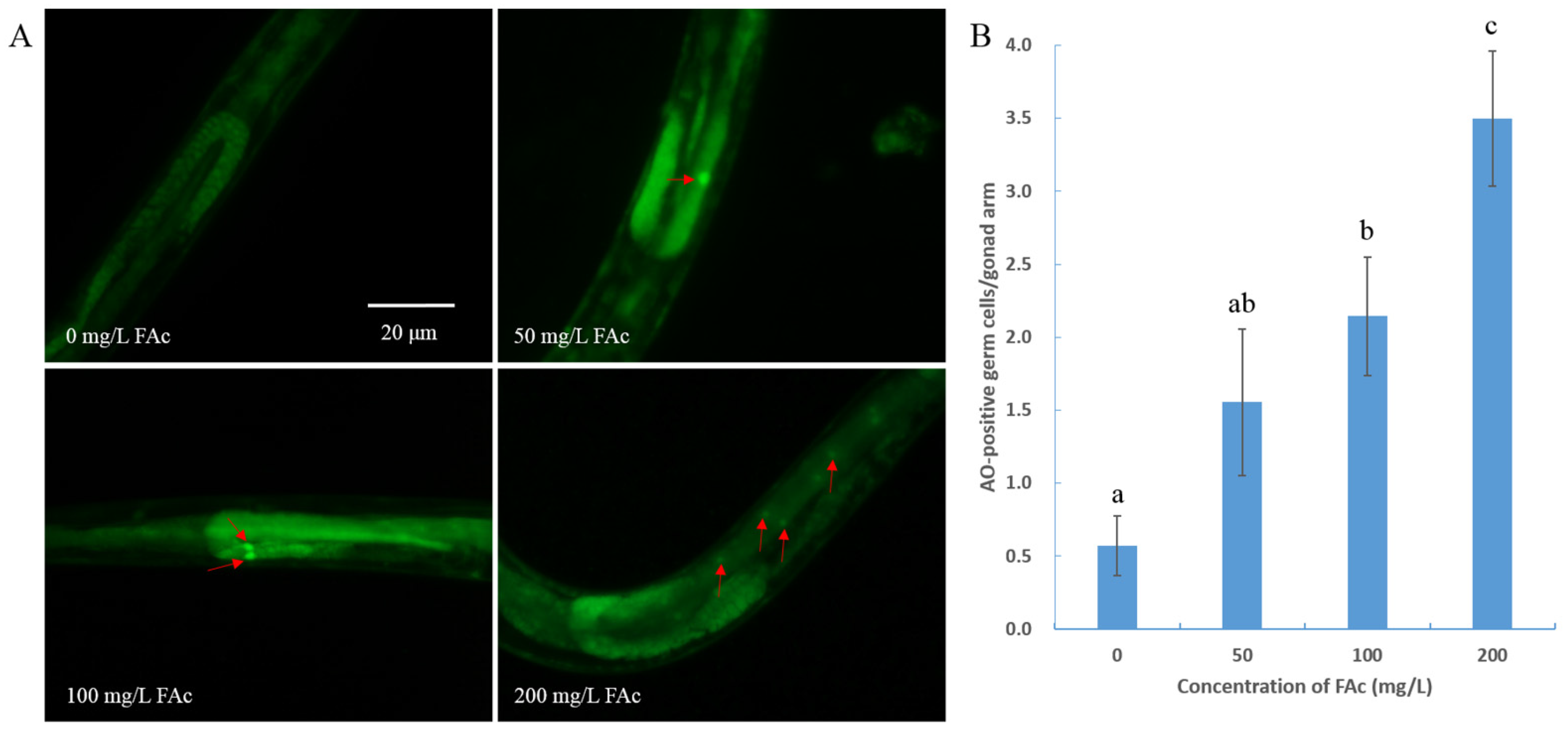
3.4. FAc Exposure Induced the Germ-Cell Apoptosis of C. elegans
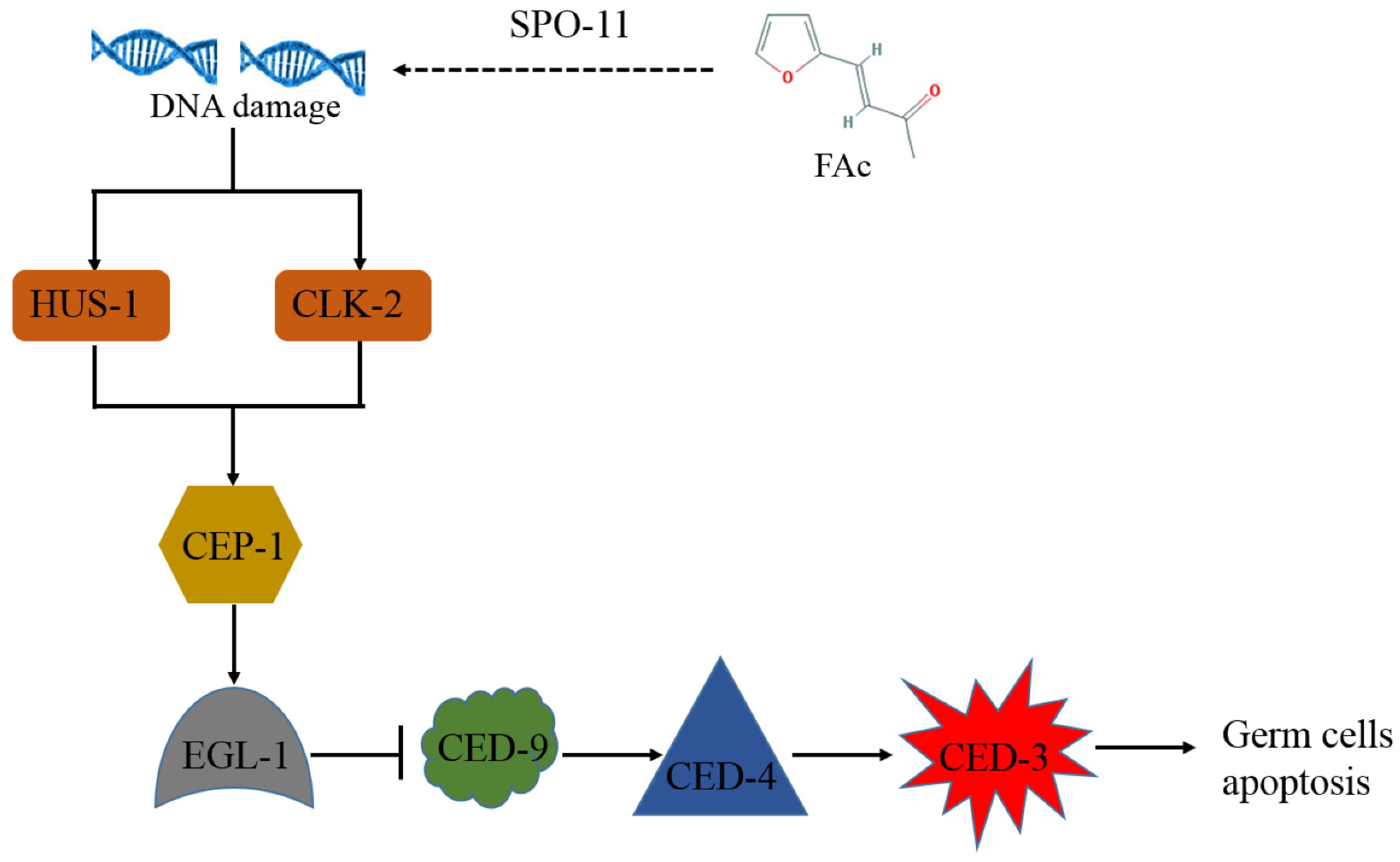
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Decraemer, W.; Hunt, D.J. Structure and classification. In Plant Nematology; Perry, R.N., Moens, M., Eds.; CAB International: Wallingford, UK, 2006; pp. 3–32. [Google Scholar]
- Jones, J.T.; Haegeman, A.; Danchin, E.G.J.; Gaur, H.S.; Helder, J.; Jones, M.G.K.; Kikuchi, T.; Manzanilla-Lopez, R.; Palomares-Rius, J.E.; Wesemael, W.M.L. Top 10 plant-parasitic nematodes in molecular plant pathology. Mol. Plant Pathol. 2013, 14, 946–961. [Google Scholar] [CrossRef] [PubMed]
- Abad, P.; Gouzy, J.; Aury, J.M.; Castagnone-Sereno, P.; Danchin, E.G.J.; Deleury, E.; Perfus-Barbeoch, L.; Anthouard, V.; Artiguenave, F.; Blok, V.C.; et al. Genome sequence of the metazoan plant-parasitic nematode Meloidogyne incognita. Nat. Biotechnol. 2008, 26, 909–915. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, T.Y.; Jang, J.Y.; Jeon, S.J.; Lee, H.W.; Bae, C.H.; Yeo, J.H.; Lee, H.B.; Kim, I.S.; Park, H.W.; Kim, J.C. Nematicidal activity of kojic acid produced by Aspergillus oryzae against Meloidogyne incognita. J. Microbiol. Biotechnol. 2016, 26, 1383–1391. [Google Scholar] [CrossRef]
- Abdelnabby, H.; Wang, Y.H.; Xiao, X.Q.; Wang, G.F.; Yang, F.; Xiao, Y.N. Impact of direct and indirect application of rising furfural concentrations on viability, infectivity and reproduction of the root knot nematode, Meloidogyne incognita in Pisum sativum. Microb. Pathog. 2016, 96, 26–34. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.X.; Li, Q.X.; Song, B.A. Chemical Nematicides: Recent research progress and outlook. J. Agric. Food Chem. 2020, 68, 12175–12188. [Google Scholar] [CrossRef] [PubMed]
- Coyne, D.L.; Cortada, L.; Dalzell, J.J.; Claudius-cole, A.O.; Haukeland, S.; Luambano, N.; Talwana, H. Plant-parasitic nematodes and food security in Sub-Saharan Africa. Annu. Rev. Phytopathol. 2018, 56, 381–403. [Google Scholar] [CrossRef] [Green Version]
- Hu, H.J.; Gao, Y.; Li, X.; Chen, S.L.; Yan, S.Z.; Tian, X.J. Identification and nematicidal characterization of proteases secreted by endophytic bacteria Bacillus cereus BCM2. Phytopathology 2020, 110, 336–344. [Google Scholar] [CrossRef]
- Migunova, V.D.; Tomashevich, N.S.; Konrat, A.N.; Lychagina, S.V.; Dubyaga, V.M.; D’Addabbo, T.; Sasanelli, N.; Asaturova, A.M. Selection of bacterial strains for control of root-knot disease caused by Meloidogyne incognita. Microorganisms 2021, 9, 1698. [Google Scholar] [CrossRef]
- Wang, J.Y.; Guo, C.; Zhao, P.; Yu, F.Y.; Su, Y.; Qu, J.P.; Wang, J.L.; Lin, R.S.; Wang, B.; Gao, Z.; et al. Biocontrol potential of Bacillus altitudinis AMCC1040 against root-knot nematode disease of ginger and its impact on rhizosphere microbial community. Biol. Control 2021, 158, 104598. [Google Scholar] [CrossRef]
- Jardim, I.N.; Oliveira, D.F.; Silva, G.H.; Campos, V.P.; de Souza, P.E. (E)-cinnamaldehyde from the essential oil of Cinnamomum cassia controls Meloidogyne incognita in soybean plants. J. Pest Sci. 2018, 91, 479–487. [Google Scholar] [CrossRef]
- Boyko, O.O.; Brygadyrenko, V.V. Nematicidal activity of aqueous tinctures of plants against larvae of the nematode Strongyloides papillosus. Trop. Biomed. 2021, 38, 85–93. [Google Scholar] [PubMed]
- Rinaldi, L.K.; Miamoto, A.; Calandrelli, A.; Silva, M.T.R.E.; Chidichima, L.P.S.; Pereira, C.B.; Dias-Arieira, C.R. Control of Meloidogyne javanica and induction of resistance-associated enzymes in soybean by extracts of Ascophyllum nodosum. J. Appl. Phycol. 2021, 33, 2655–2666. [Google Scholar] [CrossRef]
- Sun, X.W.; Zhang, R.; Ding, M.J.; Liu, Y.X.; Li, L. Biocontrol of the root-knot nematode Meloidogyne incognita by a nematicidal bacterium Pseudomonas simiae MB751 with cyclic dipeptide. Pest Manag. Sci. 2021, 77, 4365–4374. [Google Scholar] [CrossRef] [PubMed]
- Bashir, A.; Sun, Y.; Yu, X.; Sun, X.W.; Li, L. Nematicidal effects of 2-methyl-aconitate isomerase from the phytopathogen Pseudomonas syringae MB03 on the model nematode Caenorhabditis elegans. J. Invertebr. Pathol. 2021, 185, 107669. [Google Scholar] [CrossRef] [PubMed]
- Cheng, W.L.; Yang, J.Y.; Nie, Q.Y.; Huang, D.; Yu, C.; Zheng, L.Y.; Cai, M.M.; Thomashow, L.S.; Weller, D.M.; Yu, Z.N.; et al. Volatile organic compounds from Paenibacillus polymyxa KM2501-1 control Meloidogyne incognita by multiple strategies. Sci. Rep. 2017, 7, 16213. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Recillas, A.; Arroyo-Herrera, A.L.; Araujo-Leon, J.A.; Nunez, E.; Andrade, R.O. Spasmolytic and antibacterial activity of two Citrus sinensis Osbeck varieties cultivated in Mexico. Evid. Based Complementary Altern. Med. 2017, 2017, 3960837. [Google Scholar]
- Cheng, W.L.; Chen, Z.; Zeng, L.; Yang, X.; Huang, D.; Zhai, Y.L.; Cai, M.M.; Zheng, L.Y.; Thomashow, L.S.; Weller, D.M.; et al. Control of Meloidogyne incognita in three-dimensional model systems and pot experiments by the attract-and-kill effect of furfural acetone. Plant Dis. 2021, 105, 2169–2176. [Google Scholar] [CrossRef]
- Cheng, W.L.; Yang, X.; Zeng, L.; Huang, D.; Cai, M.M.; Zheng, L.Y.; Yu, Z.N.; Zhang, J.B. Evaluation of multiple impacts of furfural acetone on nematodes in vitro and control efficiency against root-knot nematodes in pots and fields. Antibiotics 2020, 9, 605. [Google Scholar] [CrossRef]
- Joint FAO/WHO Expert Committee on Food Additives. Evaluation of certain food additives: Eighty-sixth report of the Joint FAO/WHO Expert Committee on Food Additives. WHO Tech. Rep. Ser. 2019, 1014, 1–170. [Google Scholar]
- Pazdernik, N.; Schedl, T. Introduction to Germ Cell Development in Caenorhabditis elegans. Adv. Exp. Med. Biol. 2013, 757, 1–16. [Google Scholar]
- Zhou, D.; Yang, J.; Li, H.; Lu, Q.; Liu, Y.D.; Lin, K.F. Ecotoxicological evaluation of low-concentration bisphenol A exposure on the soil nematode Caenorhabditis elegans and intrinsic mechanisms of stress response in vivo. Environ. Toxicol. Chem. 2016, 35, 2041–2047. [Google Scholar] [CrossRef] [PubMed]
- Zong, Y.F.; Gao, J.L.; Feng, H.Y.; Cheng, B.J.; Zhang, X. Toxicity of 7-ketocholesterol on lethality, growth, reproduction, and germline apoptosis in the nematode Caenorhabditis elegans. J. Toxicol. Environ. Health A 2014, 77, 716–723. [Google Scholar] [CrossRef] [PubMed]
- Guha, S.; Calarco, S.; Gachet, M.S.; Gertsch, J. Juniperonic acid biosynthesis is essential in Caenorhabditis elegans lacking Δ6 desaturase (fat-3) and generates new ω-3 endocannabinoids. Cells-Basel 2020, 9, 2127. [Google Scholar] [CrossRef] [PubMed]
- Corsi, A.K.; Wightman, B.; Chalfie, M. A transparent window into biology: A primer on Caenorhabditis elegans. Genetics 2015, 200, 387–407. [Google Scholar] [CrossRef] [Green Version]
- Schumacher, B.; Schertel, C.; Wittenburg, N.; Tuck, S.; Mitani, S.; Gartner, A.; Conradt, B.; Shaham, S. C. elegans ced-13 can promote apoptosis and is induced in response to DNA damage. Cell Death Differ. 2005, 12, 153–161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gebel, J.; Tuppi, M.; Sanger, N.; Schumacher, B.; Dotsch, V. DNA damaged induced cell death in oocytes. Molecules 2020, 25, 5714. [Google Scholar] [CrossRef] [PubMed]
- Tran, A.T.; Chapman, E.M.; Flamand, M.N.; Yu, B.; Krempel, S.J.; Duchaine, T.F.; Eroglu, M.; Derry, W.B. MiR-35 buffers apoptosis thresholds in the C. elegans germline by antagonizing both MAPK and core apoptosis pathways. Cell Death Differ. 2019, 26, 2637–2651. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhai, Y.L.; Shao, Z.Z.; Cai, M.M.; Zheng, L.Y.; Li, G.Y.; Huang, D.; Cheng, W.L.; Thomashow, L.S.; Weller, D.M.; Yu, Z.N.; et al. Multiple modes of nematode control by volatiles of Pseudomonas putida 1A00316 from antarctic soil against Meloidogyne incognita. Front. Microbiol. 2018, 9, 253. [Google Scholar] [CrossRef] [Green Version]
- Freckman, D.H.; Virginia, R.A. Extraction of nematodes from dry valley antarctic soils. Polar Biol. 1993, 13, 483–487. [Google Scholar] [CrossRef]
- Manan, A.; Bazai, Z.A.; Fan, J.; Yu, H.F.; Li, L. The Nif3-family protein YqfO03 from Pseudomonas syringae MB03 has multiple nematicidal activities against Caenorhabditis elegans and Meloidogyne incognita. Int. J. Mol. Sci. 2018, 19, 3915. [Google Scholar] [CrossRef] [Green Version]
- Gartner, A.; Milstein, S.; Ahmed, S.; Hodgkin, J.; Hengartner, M.O. A conserved checkpoint pathway mediates DNA damage-induced apoptosis and cell cycle arrest in C. elegans. Mol. Cell 2000, 5, 435–443. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.C.; Zhao, Y.; Wu, L.J.; Tang, M.L.; Su, C.X.; Hei, T.K.; Yu, Z.L. Induction of germline cell cycle arrest and apoptosis by sodium arsenite in Caenorhabditis elegans. Chem. Res. Toxicol. 2007, 20, 181–186. [Google Scholar] [CrossRef]
- Lu, Q.; Bu, Y.Q.; Ma, L.Y.; Liu, R. Transgenerational reproductive and developmental toxicity of tebuconazole in Caenorhabditis elegans. J. Appl. Toxicol. 2020, 40, 578–591. [Google Scholar] [CrossRef] [PubMed]
- Hoogewijs, D.; Houthoofd, K.; Matthijssens, F.; Vandesompele, J.; Vanfleteren, J.R. Selection and validation of a set of reliable reference genes for quantitative sod gene expression analysis in C. elegans. BMC Mol. Biol. 2008, 9, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Gordon, K. Recent advances in the genetic, anatomical, and environmental regulation of the C. elegans germ line progenitor zone. J. Dev. Biol. 2020, 8, 14. [Google Scholar] [CrossRef] [PubMed]
- Min, H.; Kim, J.S.; Ahn, J.; Shim, Y.H. Gliadin intake causes disruption of the intestinal barrier and an increase in germ cell apoptosis in a Caenorhabditis elegans model. Nutrients 2019, 11, 2587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, N.; Chai, J.J.; Lee, E.S.; Gu, L.C.; Liu, Q.; He, J.Q.; Wu, J.W.; Kokel, D.; Li, H.L.; Hao, Q.; et al. Structure of the CED-4-CED-9 complex provides insights into programmed cell death in Caenorhabditis elegans. Nature 2005, 437, 831–837. [Google Scholar] [CrossRef]
- Kang, Y.Y.; Xu, J.; Liu, Y.; Sun, J.; Sun, D.P.; Hu, Y.S.; Liu, Y.F. Crystal structure of the cell corpse engulfment protein CED-2 in Caenorhabditis elegans. Biochem. Biophys. Res. Commun. 2011, 410, 189–194. [Google Scholar] [CrossRef]
- Detrey, J.; Cognard, V.; Djian-Caporalino, C.; Marteu, N.; Doidy, J.; Pourtau, N.; Vriet, C.; Maurousset, L.; Bouchon, D.; Clause, J. Growth and root-knot nematode infection of tomato are influenced by mycorrhizal fungi and earthworms in an intercropping cultivation system with leeks. Appl. Soil Ecol. 2022, 169, 104181. [Google Scholar] [CrossRef]
- Blok, V.C.; Jones, J.T.; Phillips, M.S.; Trudgill, D.L. Parasitism genes and host range disparities in biotrophic nematodes: The conundrum of polyphagy versus specialization. Bioessays 2008, 30, 249–259. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Fan, H.Y.; Zhao, D.; Zhu, X.F.; Wang, Y.Y.; Liu, X.Y.; Liu, D.; Duan, Y.X.; Chen, L.J. Multifunctional efficacy of the nodule endophyte Pseudomonas fragi in stimulating tomato immune response against Meloidogyne incognita. Biol. Control 2021, 164, 104773. [Google Scholar] [CrossRef]
- Huang, W.K.; Sun, J.H.; Cui, J.K.; Wang, G.F.; Kong, L.A.; Peng, H.; Chen, S.L.; Peng, D.L. Efficacy evaluation of fungus Syncephalastrum racemosum and nematicide avermectin against the root-knot nematode Meloidogyne incognita on cucumber. PLoS ONE 2014, 9, e89717. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Estupinan-Lopez, L.; Campos, V.P.; Silva, A.P.; Barros, A.F.; Pedroso, M.P.; Silva, J.C.P.; Terra, W.C. Volatile organic compounds from cottonseed meal are toxic to Meloidogyne incognita. Trop. Plant Pathol. 2017, 42, 443–450. [Google Scholar] [CrossRef]
- Lehmann, S.; Herrmann, F.; Kleemann, K.; Spiegler, V.; Liebau, E.; Hensel, A. Extract and the quassinoid ailanthone from Ailanthus altissima inhibit nematode reproduction by damaging germ cells and rachis in the model organism Caenorhabditis elegans. Fitoterapia 2020, 146, 104651. [Google Scholar] [CrossRef]
- Liu, Y.; Zhang, W.P.; Wang, Y.; Liu, H.M.; Zhang, S.A.; Ji, X.X.; Qiao, K. Oxidative stress, intestinal damage, and cell apoptosis: Toxicity induced by fluopyram in Caenorhabditis elegans. Chemosphere 2022, 286, 131830. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Dose (mg/pot) | Nematodes in Root/g | M. incognita Reduction in Root (%) | Nematodes in Soil/5 g | M. incognita Reduction in Soil (%) |
|---|---|---|---|---|---|
| FAc 1 | 0 | 4374 ± 821 a 3 | 75 ± 20 a | ||
| FAc | 1 | 2326 ± 429 b | 46.8 | 38 ± 3 ab | 49.6 |
| FAc | 2 | 1775 ± 480 bc | 59.4 | 32 ± 14 b | 57.1 |
| Ave 2 | 1 | 921 ± 306 c | 78.9 | 30 ± 12 b | 59.5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cheng, W.; Yang, X.; Xue, H.; Huang, D.; Cai, M.; Huang, F.; Zheng, L.; Yu, Z.; Zhang, J. Reproductive Toxicity of Furfural Acetone in Meloidogyne incognita and Caenorhabditis elegans. Cells 2022, 11, 401. https://doi.org/10.3390/cells11030401
Cheng W, Yang X, Xue H, Huang D, Cai M, Huang F, Zheng L, Yu Z, Zhang J. Reproductive Toxicity of Furfural Acetone in Meloidogyne incognita and Caenorhabditis elegans. Cells. 2022; 11(3):401. https://doi.org/10.3390/cells11030401
Chicago/Turabian StyleCheng, Wanli, Xue Yang, Hua Xue, Dian Huang, Minmin Cai, Feng Huang, Longyu Zheng, Ziniu Yu, and Jibin Zhang. 2022. "Reproductive Toxicity of Furfural Acetone in Meloidogyne incognita and Caenorhabditis elegans" Cells 11, no. 3: 401. https://doi.org/10.3390/cells11030401
APA StyleCheng, W., Yang, X., Xue, H., Huang, D., Cai, M., Huang, F., Zheng, L., Yu, Z., & Zhang, J. (2022). Reproductive Toxicity of Furfural Acetone in Meloidogyne incognita and Caenorhabditis elegans. Cells, 11(3), 401. https://doi.org/10.3390/cells11030401