Abstract
The gonadotropins, follicle-stimulating hormone (FSH) and luteinizing hormone (LH), are secreted from the pituitary and bind to the FSH receptor (FSHR) and LH receptor (LHR) to regulate gonadal development in vertebrates. Previously, using fshr-knockout (KO) medaka (Oryzias latipes), we demonstrated that FSH regulates ovarian development by elevating estrogen levels. However, the lhr-KO phenotype in medaka is poorly characterized. Here, we generated lhr-KO medaka using the transcription activator-like effector nuclease (TALEN) technique. We analyzed its phenotype and that of fshr-KO, lhr;fshr double-heterozygotes (double-hetero), and double-KO fish. All genetically male medaka displayed normal testes and were fertile, whereas fshr-KO and double-KO genetically female fish displayed small ovaries containing many early pre-vitellogenic oocytes and were infertile. Although lhr-KO genetically female fish had normal ovaries with full-grown oocytes, ovulation did not occur. Levels of 17α,20β-dihydroxy-4-pregnen-3-one, which is required for meiotic maturation of oocytes and sperm maturation in teleost fish, were significantly decreased in all KO female medaka ovaries except for double-heteros. Further, 17β-estradiol levels in fshr-KO and double-KO ovaries were significantly lower than those in double-heteros. These findings indicate that LH is necessary for oocyte maturation and FSH is necessary for follicle development, but that neither are essential for spermatogenesis in medaka.
1. Introduction
Gonadotropin-releasing hormone (GnRH) released from the hypothalamus stimulates the pituitary gland to secrete follicle-stimulating hormone (FSH) and luteinizing hormone (LH), two gonadotropins responsible for gonadal development [1,2]. FSH and LH are heterodimers of a common α subunit and different β subunits [3,4]. After secretion from the pituitary gland, these hormones bind to specific receptors, FSH receptor (FSHR) and LH receptor (LHR), in gonadal somatic cells to regulate secretion of sex steroid hormones, such as testosterone, estrogen, and progesterone [5,6,7].
The relationships between sexual maturation and FSHR or LHR functions in vertebrates have been intensively studied. In mammals, FSHR is expressed exclusively in Sertoli cells of the testes and granulosa cells of the ovary [8,9]. Fshr-knockout (KO) male mice display small testes and partial failure of spermatogenesis; however, they are fertile [8]. Fshr-KO female mice, however, display thin uteri and small ovaries and are infertile because folliculogenesis does not occur before antral follicle formation [8,10]. In fish, Fshr is expressed not only in Sertoli cells and granulosa cells, but also in Leydig cells of the testes and theca cells of the vitellogenic ovarian follicle [11,12,13,14]. Using fshr-KO medaka (Oryzias latipes), we have recently demonstrated that FSH regulates ovarian development and maintenance, mainly by elevating estrogen levels [15]. fshr-KO male medaka show normal testes and are fertile, whereas female medaka display small ovaries and are infertile. These results indicate that the FSH signal via FSHR/Fshr is necessary for ovarian development but is not important for spermatogenesis in mammals and fish.
In mammals, LHR is expressed in Leydig cells of the testes and in theca cells, large follicles, and the corpora lutea of the ovary [16,17]. In Lhr-KO male mice, postnatal testes development is blocked, and spermatogenesis is arrested at the round spermatid stage [18,19]. Lhr-KO female mice display underdeveloped external genitalia and uteri postnatally [18,19]. Thus, both Lhr-KO sexes are infertile [18,19,20]. In fish, Lhr is expressed in Sertoli cells and Leydig cells of the testes and in follicle layers including granulosa cells of the ovary in several fish species, including zebrafish (Danio rerio) [14,21,22,23]. In a previous study using lhβ-KO or lhr-KO male zebrafish, the fertilization rate, sperm motility, and histological structure of the testis were not affected [24]. However, female homozygous lhβ-KO fish are infertile, whereas homozygous lhr-KO fish are fertile. Therefore, the effects of LHR/Lhr on gonadal development are different between mammals and fish. So et al. [25] reported that FSH specifically activates Fshr, whereas LH can stimulate both Fshr and Lhr in zebrafish, supporting the notion that LH acts through the Fshr in the absence of Lhr, leading to normal fertility in female lhr-KO zebrafish. The complex relationship between FSH and LH signaling pathways in fish has complicated elucidation of the roles of these gonadotropic hormones in sexual development. By generating KOs of the gonadotropin receptors in medaka, which are phylogenetically distant from Zebrafish, and analyzing their phenotypes, we can better understand FSH and LH signaling in sexual development and reproduction.
Medaka has desirable features for a model vertebrate, such as easy breeding, short generation time, and small genome, and has been used to study vertebrate reproduction [26]. In addition, transgenic and gene editing methods, using transcription activator-like effector nuclease (TALEN) or clustered regularly interspaced short palindromic repeats (CRISPR)/CRISPR-associated protein 9 (Cas9) technologies, are established [27,28], and the medaka sex-determining gene, DMY/dmrt1bY, which is located on the Y chromosome, has been identified [29,30,31]. Therefore, medaka is an excellent vertebrate model organism for the molecular genetic analysis of various biological phenomena, including embryonic development and sexual maturation.
In the present study, to reveal gonadotropin receptor function in the sexual development of medaka, we first generated lhr single-KO (lhr-KO) medaka using the TALEN technique and analyzed its phenotype and sex steroid hormone expression pattern. We then generated lhr;fshr double-heterozygotes (double-hetero), lhr heterozygote;fshr single-KO (fshr-KO), and lhr;fshr double-KO (double-KO) medaka to investigate the function of the two gonadotropin receptors in gonadal development and reproduction.
2. Materials and Methods
2.1. Ethics Statement
This study was performed using protocols approved by the Animal Care and Use Committee of Kumamoto University (A2020-014) and according to the ARRIVE guidelines.
2.2. Animals
The FLFII medaka line was used in this study. This stock allows the identification of genotypic sex by the appearance of leucophores at 2 days post-fertilization. Fish embryos were maintained in embryo-rearing medium (ERM: 17 mM NaCl, 0.4 mM KCl, 0.27 mM CaCl2·2H2O, 0.66 mM MgSO4, pH 7) at 26 °C under a 14 h light and 10 h dark cycle.
2.3. TALEN Preparation
TALEN experiments were performed as described in our recent report [27]. Briefly, the TALEN target sites in the lhr locus were searched for using the TALEN Targeter program (https://tale-nt.cac.cornell.edu/node/add/talen, accessed on 10 December 2021), and TAL repeats were assembled using the Golden Gate assembly system and the repeat variable di-residue (RVD) arrays assembly reaction, as described previously [32] but with slight modifications. Resulting plasmids that acquired RVD modules were identified by blue/white selection on IPTG/X-gal LB +ampicillin plates and correct assembly was determined by the size of insert generated by BspEI restriction enzyme digestion. Synthetic mRNAs were generated using ApaI-linearized TALEN expression vectors as templates and the mMessage mMachine T7 Ultra Kit (Ambion) and stored at −80 °C until use.
2.4. Microinjection
Microinjection of medaka embryos was performed at the one-cell stage using a Nanoject II (Drummond Scientific Co., Broomall, PA, USA). TALEN mRNAs were simultaneously injected into embryos. After injection, embryos were maintained in ERM at 26 °C in an MIR-153 incubator (SANYO Electric Co., Ltd., Hokkaido, Japan).
2.5. Genotyping
The genetic sex of each adult fish was decided by genomic PCR as described previously [33]. PCR was performed using primers specific for dmy (DDBJ accession no. AB071534). The conditions were as follows: preheating at 95 °C for 10 min, 40 cycles of PCR at 94 °C for 30 s, 59 °C for 30 s, 72 °C for 1 min, and a final extension at 72 °C for 5 min.
Genotyping of lhr-KO, double-hetero, fshr-KO, and double-KO medaka was performed by genomic PCR. PCR was performed using specific primers for lhr (DDBJ accession no. CP020635; 5′-GCTGAATATCAACAGTCTGAGG-3′ and 5′-CCGTAATTTTCTTAATATCTGTG-3′) and for fshr [15]. The conditions were as follows: preheating at 95 °C for 10 min, 40 cycles of PCR at 94 °C for 30 s, 59 °C for 30 s, 72 °C for 1 min, and final extension at 72 °C for 5 min.
2.6. Fertility Assessment
The fertility of adult fish was investigated by natural mating with fertile wild-type partners. Fertility assessment was conducted with three mating pairs for each genotype, and individuals that failed to produce fertilized embryos were considered infertile.
2.7. Determination of Gonadosomatic Index (GSI)
The adult gonads of double-hetero, lhr-KO, fshr-KO, and double-KO medaka at approximately 6 months of age were weighed, and the GSI was calculated as follows: GSI (%) = gonad weight (g) × 100/body weight (g).
2.8. Measurement of Steroids
Steroid hormones were extracted from gonads of adult individuals in diethyl ether as described previously [34]. The levels of 17α,20β-dihydroxy-4-pregnen-3-one (DHP) in males and females, 11-ketotestosterone (11-KT) in males and 17β-estradiol (E2) in females, were measured using time-resolved fluoroimmunoassays according to the method of Yamada et al. [35]. DHP, 11-KT and E2 were purchased from Sigma Chemicals (St. Louis, MO, USA), and antibodies against E2 and DHP were purchased from Cosmo-Bio Co., Ltd. (Tokyo, Japan).
2.9. Histological Analysis of Gonads
The ovaries and testes of adult fish at approximately 6 months of age were fixed in Bouin’s solution at 4 °C overnight. After fixation, each sample was dehydrated, embedded in paraffin, and sectioned serially at a thickness of 5 μm. Then, the sections were stained with hematoxylin and eosin [33]. The number of oocytes per ovary was counted, and the developmental stage of oocytes was determined by size and basic morphological characteristics [36].
2.10. Statistics
Experimental results were tested for homogeneity of variance using Levene’s test. Data were analyzed by one-way analysis of variance and then tested with Tukey’s multiple comparison test using SPSS statistics 20 (IBM Corp. Armonk, NY, USA).
3. Results
3.1. Establishment of lhr-KO Lines
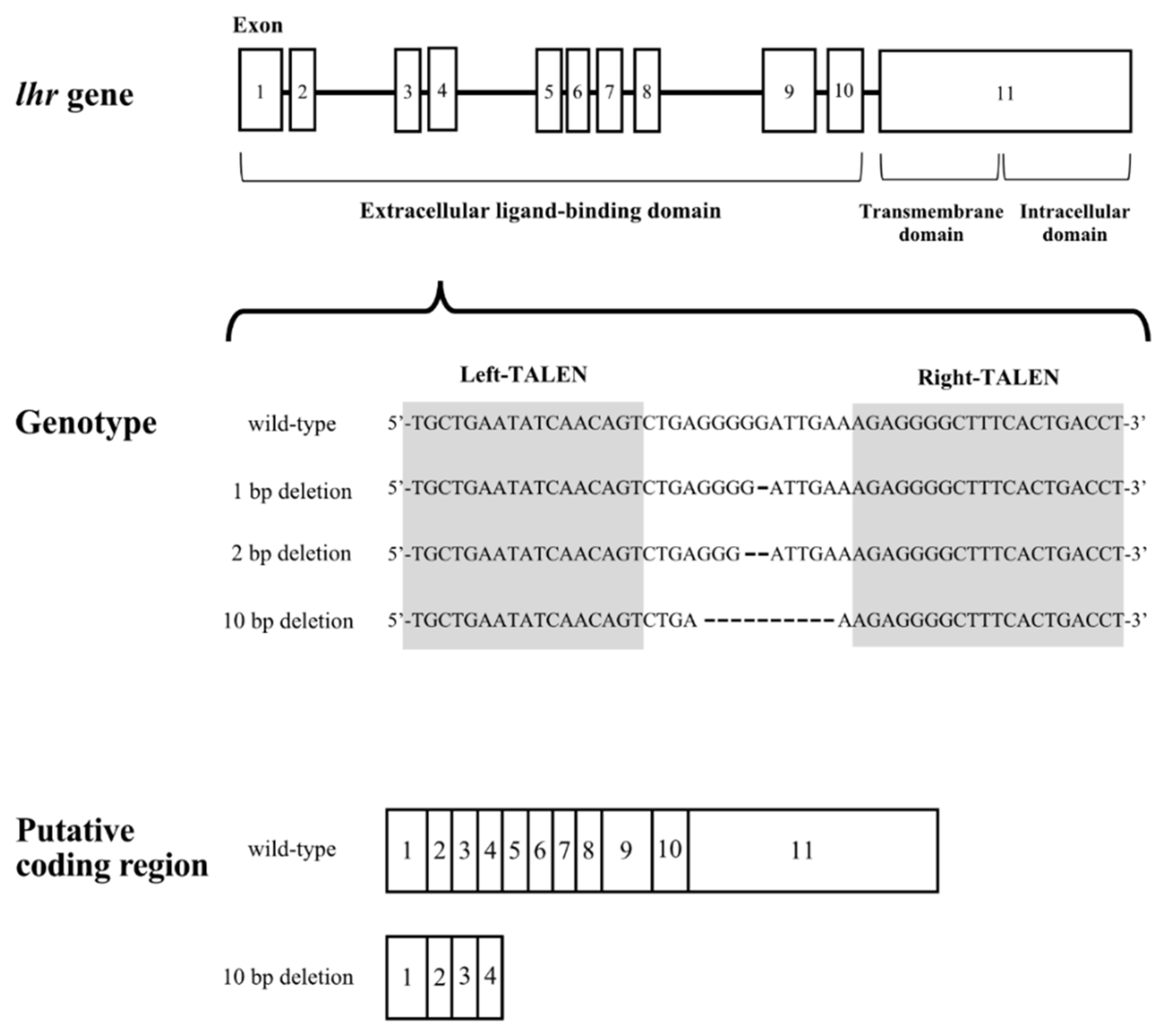
The target sequences of the TALEN for lhr and results of gene modification are shown in Figure 1. We obtained medaka with 1, 2, and 10-nucleotide deletions in the lhr gene. In the 10-nucleotide deletion, the 133rd leucine residue was replaced with a stop codon by a frameshift. No transmembrane domain or intracellular domain can be synthesized by this transcript because the TALEN was targeted to exon 4, which encodes part of the extracellular ligand-binding domain of Lhr. We mated lhr 10-nucleotide deletion heterozygous XY male and XX female medaka to obtain the F1 generation and successfully generated lhr-KO (lhr−/−) medaka in the F2 generation.
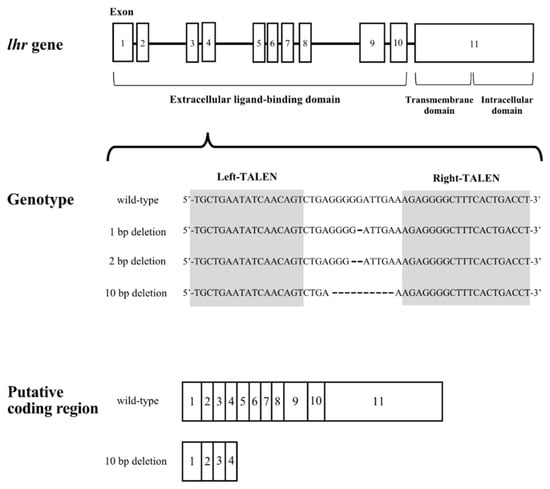
Figure 1.
Schematic diagram of the lhr gene structure. Gray arabic numerals indicate exon positions. Shaded sequence indicates the sequence targeted by TALEN in exon 4.
3.2. Phenotypic Analyses of lhr-KO, Double-Hetero, fshr-KO, and Double-KO Medaka
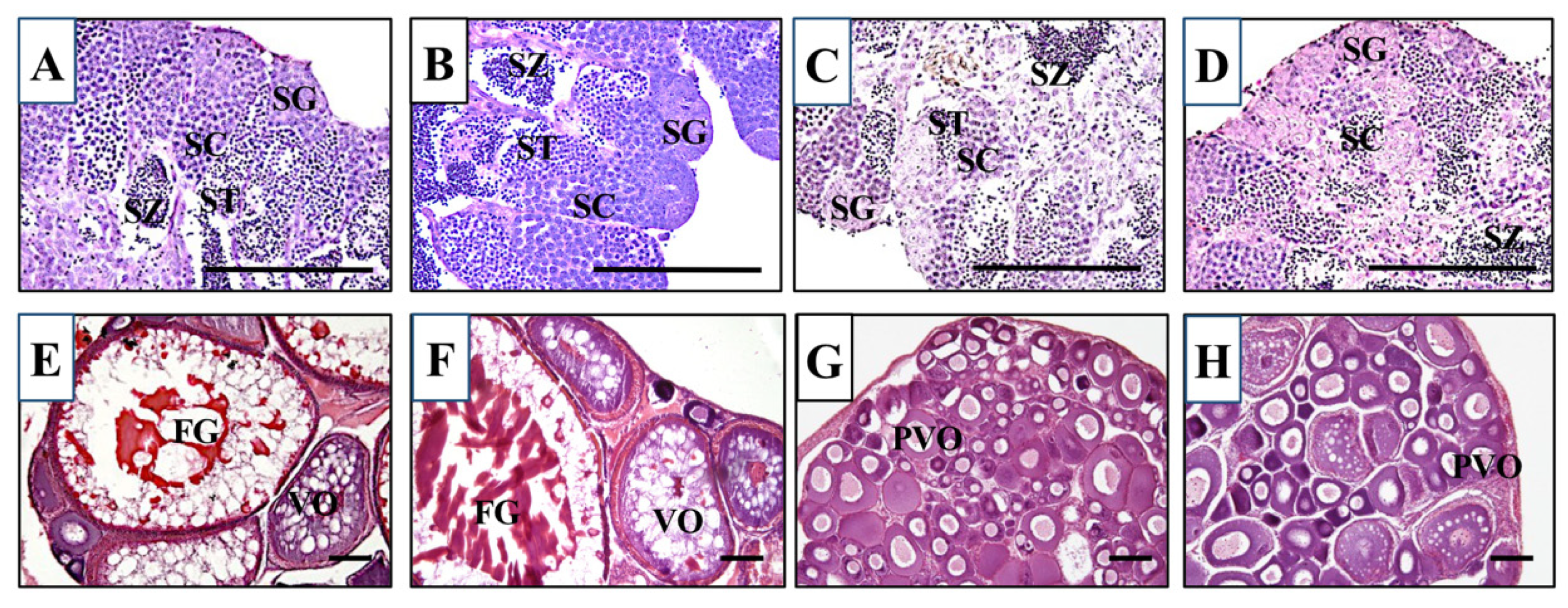
We first crossed an lhr 10-nucleotide deletion heterozygous XX female (lhr+/−;fshr+/+) with an fshr-KO XY male (lhr+/+;fshr−/−). Males and females of double-hetero (lhr+/−;fshr+/−) were selected by genotyping of lhr and fshr genes, and then they were crossed with each other. This cross generated lhr-KO (lhr−/−;fshr+/+), double-hetero (lhr+/−;fshr+/−), fshr-KO (lhr+/−;fshr−/−), and double-KO (lhr−/−;fshr−/−) medaka, which were genotyped using two-step genomic PCR and HRM analysis. For phenotypic analyses, all genotypically male medaka displayed normal testes containing from spermatocytes to spermatozoa, and no abnormal stage of spermatogenesis was observed (Figure 2A–D), similar to wild-type testes [15]. The double-hetero and lhr-KO female medaka also displayed normal ovaries, including full-grown oocytes (Figure 2E,F), similar to wild-type ovaries [15]. In fshr-KO and double-KO female fish ovaries, many immature oocytes were observed (Figure 2G,H).
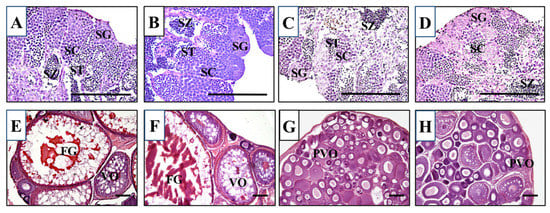
Figure 2.
Histological sections of double-hetero (A), lhr-KO (B), fshr-KO (C), and double-KO (D) male medaka testes and double-hetero (E), lhr-KO (F), fshr-KO (G), and double-KO (H) female medaka ovaries. FG: full-grown oocyte, PVO: pre-vitellogenic oocyte, SC: spermatocyte, SG: spermatogonium, ST: spermatid, SZ: spermatozoa, VO: vitellogenic oocyte. Bar, 100 µm.
3.3. Gonad Development and Fertility in KO Medaka
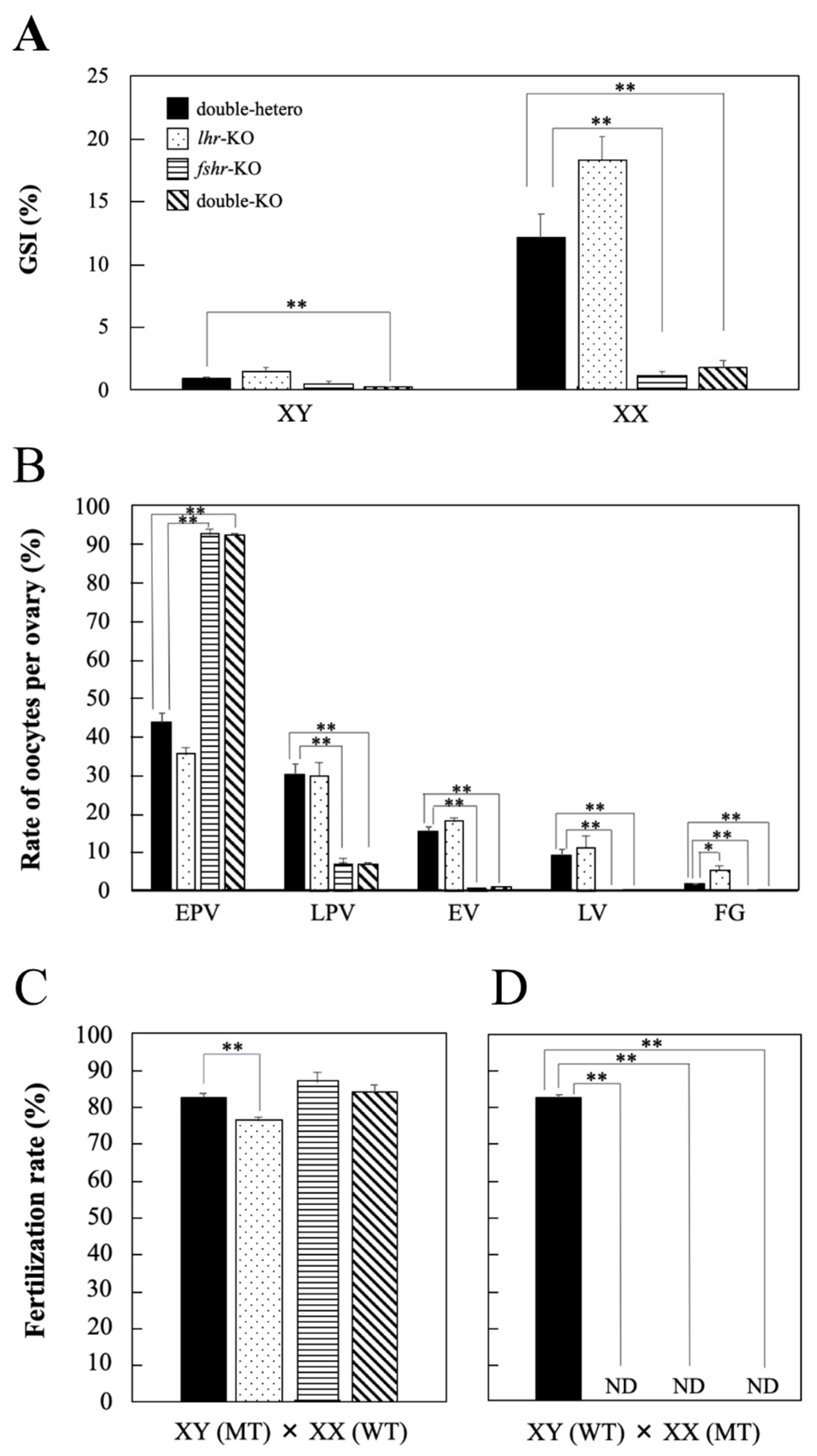
No significant differences in GSI values were observed in lhr-KO and fshr-KO males compared with double-hetero male medaka; however, that in double-KO males was significantly decreased (p < 0.01) (Figure 3A). In fshr-KO and double-KO female fish, GSI values were significantly lower (p < 0.01) than in double-hetero female medaka (Figure 3A). However, there was no difference in GSI between double-hetero and lhr-KO females.
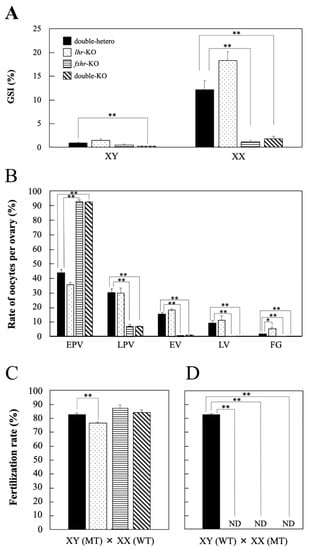
Figure 3.
GSI value (%) in gonads of double-hetero, lhr-KO, fshr-KO, and double-KO adult male (XY) and female (XX) medaka (A). The ratio of different stage oocytes in double-hetero, lhr-KO, fshr-KO, and double-KO female medaka ovaries (B). EPV: early pre-vitellogenic oocyte, LPV: late pre-vitellogenic oocyte, EV: early vitellogenic oocyte, LV: late vitellogenic oocyte, FG: full-grown oocyte. Fertilization rate (%) in double-hetero, lhr-KO, fshr-KO, and double-KO male (C) and female (D) medaka mated with fertile wild-type partners. MT: mutant type, WT: wild-type. ND: not detected. Individuals that failed to produce fertilized embryos after 3 trials were considered ND. Vertical bar indicates ± SEM (n = 3). * p < 0.05, ** p < 0.01.
The number of oocytes per ovary was counted to investigate the developmental stage of ovaries in each genotype (Figure 3B). There were many full-grown oocytes in lhr-KO females compared with double-hetero females. Ovary histology was similar between fshr-KO and double-KO females, and there were many early pre-vitellogenic oocytes compared with double-hetero fish (Figure 3B).
The fertilization rate of lhr-KO males was slightly decreased compared with that of double-hetero male medaka (p < 0.05) (Figure 3C). The fshr-KO and double-KO male medaka had the same level of fertility as double-hetero males. However, when lhr-KO, fshr-KO, or a double-KO female was crossed with a wild-type male, fertilized eggs were not obtained (Figure 3D).
3.4. Steroid Hormone Levels in KO Medaka
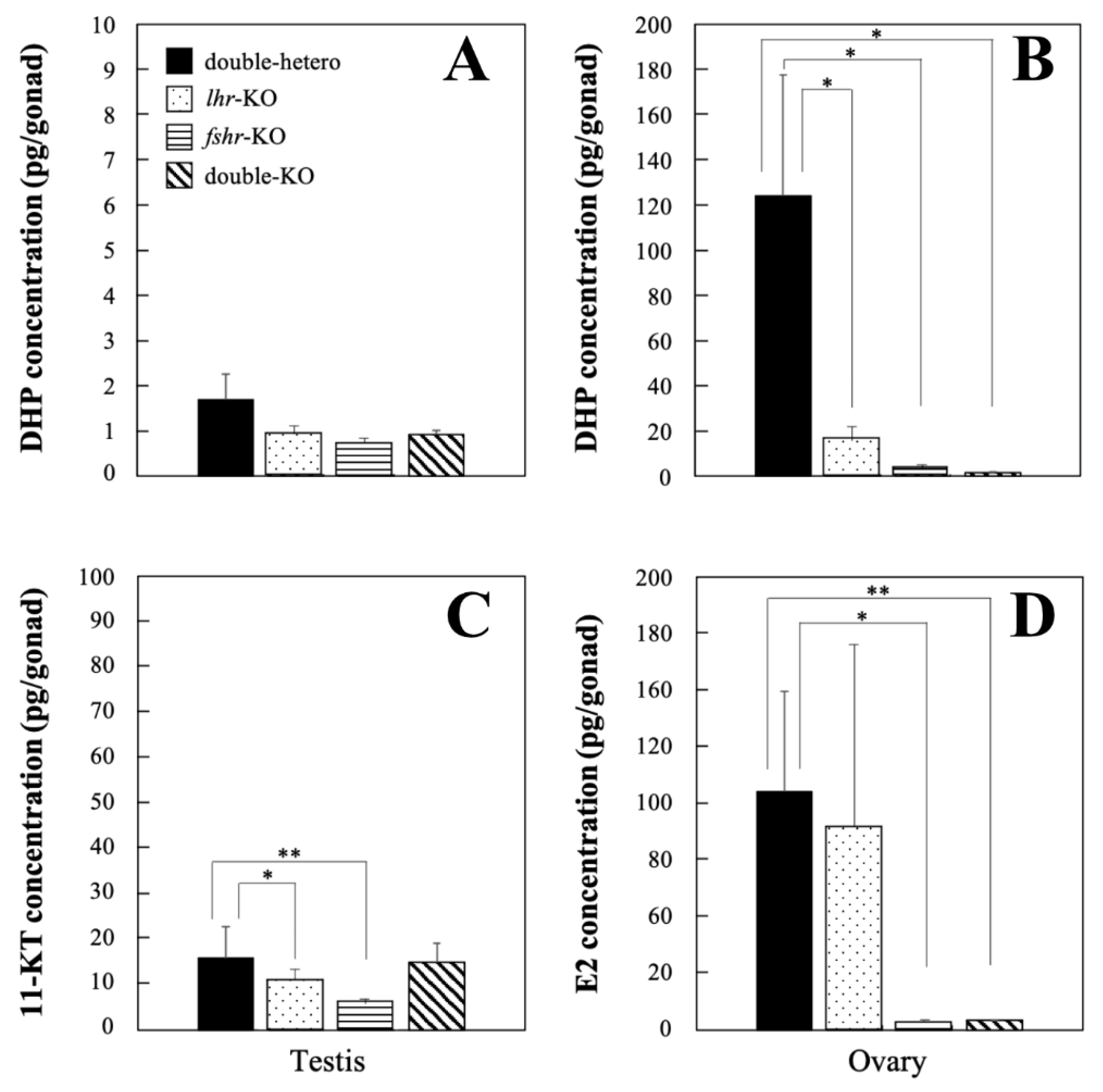
No significant difference in DHP levels was observed among lhr-KO, fshr-KO, double-KO, and double-hetero male testes at the adult stage (Figure 4A), whereas there was a significant decrease (p < 0.01) in lhr-KO, fshr-KO, and double-KO female ovaries compared with double-hetero female ovaries (Figure 4B). Levels of 11-KT in lhr-KO and fshr-KO testes were significantly lower (p < 0.05, p < 0.01) than those in double-hetero testes, while there was no difference between double-KO and double-hetero testes (Figure 4C). E2 levels were the same in lhr-KO and double-hetero ovaries, similar to those in wild-type ovaries [15], but were significantly lower in fshr-KO and double-KO ovaries (p < 0.05, p < 0.01) (Figure 4D).
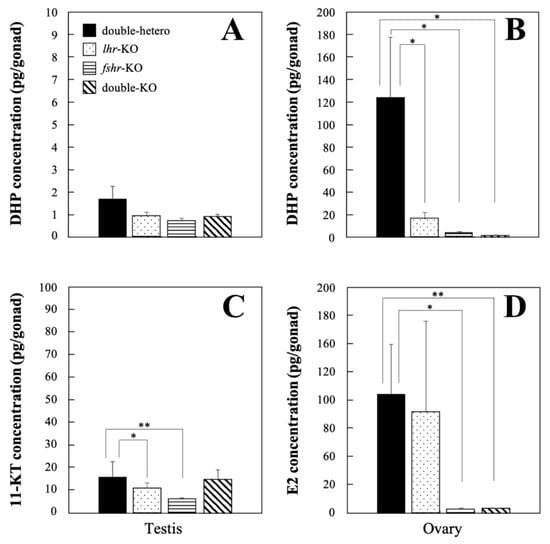
Figure 4.
DHP concentration in double-hetero, lhr-KO, fshr-KO, and double-KO male (A) and female (B) medaka. 11-KT concentration in male testes (C) and E2 concentration in female ovaries (D) in double-hetero, lhr-KO, fshr-KO, and double-KO medaka. Steroid hormones were extracted from gonads of adult medaka. Vertical bar indicates ± SEM (n = 3). * p < 0.05, ** p < 0.01.
4. Discussion
In this study, we generated lhr-KO, double-hetero, fshr-KO, and lhr;fshr double-KO medaka and demonstrated the involvement of Lhr and Fshr in gonad development and fertility of medaka. All genotypically male medaka were fertile and possessed normal testes containing all stages of spermatogenic germ cells. However, in fshr-KO and lhr;fshr double-KO ovaries, oogenesis halted at the previtellogenic oocyte stage. Furthermore, lhr-KO females had full-grown oocytes before maturation, but no ovulation occurred. Consequently, these KO females were infertile. These results strongly indicate that LH and FSH are not involved in male reproduction but play a crucial role in ovarian development and female reproduction in medaka. Furthermore, LH and FSH were shown to have distinct roles in female gonad development; FSH is necessary for the development of follicles, whereas LH is essential for oocyte maturation.
In lhr-KO female medaka, ovulation did not occur, although the oocytes developed up to the stage of full-grown follicles. Then, in lhr-KO females, levels of DHP, which induces oocyte maturation [37,38], were significantly lower than those in double-hetero female medaka (Figure 4B). It is widely accepted that, in fish, LH induces oocyte maturation by stimulating the production of maturation-inducing hormones (DHP in most fishes) in follicle layer cells [38,39]. Hence, DHP production is suggested to be suppressed in lhr-KO female medaka because LH signaling is not established, leading to the inhibition of ovulation. In zebrafish [39] and mammals [40], insulin-like growth factors (IGFs) act as important mediators of LH action on oocyte maturation. Therefore, it appears likely that LH induces the maturation via the IGF system in vertebrates.
The ovarian development phenotype in the loss of Fshr function fish (fshr-KO and lhr;fshr double-KO) was similar to that observed in our previous study using Fshr-deficient medaka [15]. The present results show that folliculogenesis in fshr-KO and lhr;fshr double-KO females was halted at the previtellogenic stage (Figure 2G,H). In addition, E2 levels in these females were significantly lower than those in double-hetero females (Figure 4D). Similarly, in zebrafish lacking Fshr, all ovarian follicles arrested at the previtellogenic stage, indicating that FSH/Fshr is crucial for primary growth of oocytes [41]. These findings indicate that the disruption of folliculogenesis can be caused by a decrease in E2 level as a result of deficient FSH signaling. Indeed, in salmonid fish, FSH induces vitellogenesis through elevation of E2 levels [42]. Furthermore, a lack of FSH signaling in female mice causes severe ovarian underdevelopment resulting in chronic estrogen deficiency [10]. Therefore, the mechanism of ovarian development by regulating E2 production via FSH signaling is common in vertebrates. In Fshr-KO female mice, early loss of estrogen leads to obesity and skeletal abnormalities that intensify with age [10]. In this study, no obvious obesity and skeletal abnormalities were found in all genotypically female medaka, even though estrogen levels decreased in fshr-KO and lhr;fshr double-KO female fish (data not shown). Accordingly, roles of estrogen in mammals and teleosts may differ slightly, although further investigation is needed to confirm this.
A similar study has been reported for lhr and fshr KO zebrafish. Using single lhr and fshr KO and lhr;fshr double-KO zebrafish, FSH was shown to be essential for folliculogenesis and LH for oocyte maturation [24,41,43]. lhr-KO male and female zebrafish were fertile, whereas double-KO male and female fish were infertile [40], which is not consistent with the present study. So et al. reported that, in zebrafish, FSH specifically activates Fshr, whereas LH can stimulate both Fshr and Lhr [25], meaning that Fshr bind both ligands. This supports the notion that LH signaling acts through the Fshr in the absence of the Lhr, to maintain fertility in lhr-KO female zebrafish. In teleosts, the ligand selectivity of Fshr/Lhr is not highly specific [44]; in particular, teleost Fshrs appear to possess a broader but limited functional selectivity for both FSH and LH that might depend on the fish species [45]. However, the present results reveal that females, even lhr single-KO females, in which Fshr is active were infertile because ovulation was prevented. This result indicates that ligand selectivity of Fshr/Lhr is apparently exclusive in medaka, as in mammals.
Interestingly, lhr;fshr double-KO phenotypes are distinctly different between medaka and zebrafish. In double-KO zebrafish males, testes are underdeveloped, and the fish are infertile [41]; however, in medaka, double-KO males had histologically normal testes with all stages of spermatogenic germ cells, no marked differences in steroid hormone levels compared to double-hetero fish, and fertility was confirmed. In teleosts, both gonadotropins, LH and FSH, directly stimulate gonadal sex steroid hormone production by activating Leydig cells [44,46]. Specifically, 11-KT and DHP, stimulated by FSH, appear to play an important role in spermatogenesis in salmonids [47]. In fact, there are several reports that FSH can induce testicular growth and spermatogenesis in vitro and in vivo [11,48,49]. Based on these observations, the double-KO testis phenotype observed in zebrafish is reasonable. In wild-type male medaka, fshr and lhr are expressed in all testis stages, from immature to fully mature [50], and it is assumed that gonadotropins are involved. However, even with these functional defects, no phenotype is expressed. To clarify the role of gonadotropins in male medaka, further studies are needed.
5. Conclusions
Here, we report the phenotypic analysis of lhr- and lhr;fshr-KO medaka. The double-hetero, lhr-KO, fshr-KO, and double-KO male medaka displayed normal testes and fertility. lhr-KO female fish, however, showed low levels of DHP, no ovulation, and infertility, although normal ovaries with full-grown oocytes were observed. Moreover, fshr-KO and double-KO female fish displayed small ovaries with pre-vitellogenic oocytes and were infertile. We suggest that folliculogenesis was halted at the previtellogenic stage because of deficient FSH signaling that resulted in depleted levels of E2. These findings indicate that, in medaka, LH is necessary for oocyte maturation and FSH is necessary for the development of follicles, whereas these gonadotropins are not essential for male sexual development including spermatogenesis.
Author Contributions
T.K. obtained funding and designed the study. T.T., H.T., Y.Y., Y.K., T.H., K.M. and R.N. performed the experiments and collected the data. T.K., K.M. and R.N. wrote the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by JSPS KAKENHI Grant Number 19H03052 (to T.K.) from the Ministry of Education, Culture, Sports, Science and Technology of Japan.
Institutional Review Board Statement
The present study was performed using protocols approved by the Animal Care and Use Committee of Kumamoto University (A2020-014) and according to the ARRIVE guidelines.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data are contained within this article. Raw data are available on request from the corresponding author.
Acknowledgments
We thank Tomomi Iwashita and Aki Hokyo for technical assistance. We also thank Jeremy Allen for editing a draft of this manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Conn, P.P.M.; Crowley, W.F. Gonadotropin-releasing hormone and its analogs. Annu. Rev. Med. 1994, 45, 391–405. [Google Scholar] [CrossRef] [PubMed]
- Kaiser, U.B.; Conn, P.M.; Chin, W.W. Studies of gonadotropin-releasing hormone (GnRH) action using GnRH receptor-expressing pituitary cell lines. Endocr. Rev. 1997, 18, 46–70. [Google Scholar] [CrossRef] [PubMed]
- Pierce, J.G.; Parsons, T.F. Glycoprotein hormones: Structure and function. Ann. Rev. Biochem. 1981, 50, 465–495. [Google Scholar] [CrossRef] [PubMed]
- Ryan, R.J.; Charlesworth, M.C.; McCormick, D.J.; Milius, R.P.; Keutmann, H.T. The glycoprotein hormones: Recent studies of structure-function relationships. FASEB J. 1988, 2, 2661–2669. [Google Scholar] [CrossRef]
- Ulloa-Aguirre, A.; Midgley, A.R.; Beitins, I.Z.; Padmanabhan, V. Follicle-stimulating isohormones: Characterization and physiological relevance. Endocr. Rev. 1995, 16, 765–787. [Google Scholar] [CrossRef]
- Sprengel, R.; Braun, T.; Nikolics, K.; Segaloff, D.L.; Seeburg, P.H. The testicular receptor for follicle stimulating formone: Structure and functional expression of cloned cDNA. Mol. Endocrinol. 1990, 4, 525–530. [Google Scholar] [CrossRef] [Green Version]
- Kishi, H.; Kitahara, Y.; Imai, F.; Nakao, K.; Suwa, H. Expression of the gonadotropin receptors during follicular development. Reprod. Med. Biol. 2018, 17, 11–19. [Google Scholar] [CrossRef]
- Dierich, A.; Sairam, M.R.; Monaco, L.; Fimia, G.M.; Gansmuller, A.; LeMeur, M.; Sassone-Corsi, P. Impairing follicle-stimulating hormone (FSH) signaling in vivo: Targeted disruption of the FSH receptor leads to aberrant gametogenesis and hormonal imbalance. Proc. Natl. Acad. Sci. USA 1998, 95, 13612–13617. [Google Scholar] [CrossRef] [Green Version]
- Kempisty, B.; Ziółkowska, A.; Ciesiolka, S.; Piotrowska, H.; Antosik, P.; Bukowska, D.; Nowicki, M.; Brüssow, K.P.; Zabel, M. Association between the expression of LHR, FSHR and CYP19 genes, cellular distribution of encoded proteins and proliferation of porcine granulosa cells in real-time. J. Boil. Regul. Homeost. Agents 2014, 28, 419–431. [Google Scholar]
- Danilovich, N.; Babu, P.S.; Xing, W.; Gerdes, M.; Krishnamurthy, H.; Sairam, M.R. Estrogen Deficiency, Obesity, and Skeletal Abnormalities in Follicle-Stimulating Hormone Receptor Knockout (FORKO) Female Mice. Endocrinology 2000, 141, 4295–4308. [Google Scholar] [CrossRef]
- Ohta, T.; Miyake, H.; Miura, C.; Kamei, H.; Aida, K.; Miura, T. Follicle-stimulating hormone induces spermatogenesis mediated by androgen production in Japanese eel, Anguilla japonica. Biol. Reprod. 2007, 77, 970–977. [Google Scholar] [CrossRef] [Green Version]
- Andersson, E.; Nijenhuis, W.; Male, R.; Swanson, P.; Bogerd, J.; Taranger, G.L.; Schulz, R.W. Pharmacological characterization, localization and quantification of expression of gonadotropin receptors in Atlantic salmon (Salmo salar L.) ovaries. Gen. Comp. Endocrinol. 2009, 163, 329–339. [Google Scholar] [CrossRef] [PubMed]
- García-López, A.; Bogerd, J.; Granneman, J.C.M.; Van Dijk, W.; Trant, J.M.; Taranger, G.L.; Schulz, R.W. Leydig cells express follicle-stimulating hormone receptors in African catfish. Endocrinology 2009, 150, 357–365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- García-López, A.; de Jonge, H.; Nóbrega, R.H.; de Waal, P.P.; van Dijk, W.; Hemrika, W.; Taranger, G.L.; Bogerd, J.; Schulz, R.W. Studies in zebrafish reveal unusual cellular expression patterns of gonadotropin receptor messenger ribonucleic acids in the testis and unexpected functional differentiation of the gonadotropins. Endocrinology 2010, 151, 2349–2360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murozumi, N.; Nakashima, R.; Hirai, T.; Kamei, Y.; Ishikawa-Fujiwara, T.; Todo, T.; Kitano, T. Loss of follicle-stimulating hormone receptor function causes masculinization and suppression of ovarian development in genetically female medaka. Endocrinology 2014, 155, 3136–3145. [Google Scholar] [CrossRef] [Green Version]
- Zheng, M.; Shi, H.; Segaloff, D.L.; Van Voorhis, B.J. Expression and localization of luteinizing hormone receptor in the female mouse reproductive tract. Biol. Reprod. 2001, 64, 179–187. [Google Scholar] [CrossRef] [Green Version]
- Guo, J.-J.; Ma, X.; Wang, C.Q.; Ge, Y.-F.; Lian, Q.-Q.; Hardy, D.O.; Zhang, Y.-F.; Dong, Q.; Xu, Y.-F.; Ge, R.-S. Effects of luteinizing hormone and androgen on the development of rat progenitor Leydig cells in vitro and in vivo. Asian J. Androl. 2013, 15, 685–691. [Google Scholar] [CrossRef] [Green Version]
- Zhang, F.-P.; Poutanen, M.; Wilbertz, J.; Huhtaniemi, I. Normal prenatal but arrested postnatal sexual development of luteinizing hormone receptor knockout (LuRKO) mice. Mol. Endocrinol. 2001, 15, 172–183. [Google Scholar] [CrossRef]
- Lei, Z.M.; Mishra, S.; Zou, W.; Xu, B.; Foltz, M.; Li, X.; Rao, C.V. Targeted disruption of luteinizing hormone/human chorionic gonadotropin receptor gene. Mol. Endocrinol. 2001, 15, 184–200. [Google Scholar] [CrossRef]
- Ascoli, M.; Fanelli, F.; Segaloff, D.L. The lutropin/choriogonadotropin receptor, a 2002 perspective. Endocr. Rev. 2002, 23, 141–174. [Google Scholar] [CrossRef]
- Miwa, S.; Yan, L.; Swanson, P. Localization of two gonadotropin receptors in the salmon gonad by in vitro ligand autoradiography. Biol. Reprod. 1994, 50, 629–642. [Google Scholar] [CrossRef] [PubMed]
- Oba, Y.; Hirai, T.; Yoshiura, Y.; Kobayashi, T.; Nagahama, Y. Fish gonadotropin and thyrotropin receptors: The evolution of glycoprotein hormone receptors in vertebrates. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2001, 129, 441–448. [Google Scholar] [CrossRef]
- Ogiwara, K.; Fujimori, C.; Rajapakse, S.; Takahashi, T. Characterization of luteinizing hormone and luteinizing hormone receptor and their indispensable role in the ovulatory process of the medaka. PLoS ONE 2013, 8, e54482. [Google Scholar] [CrossRef] [PubMed]
- Chu, L.; Li, J.; Liu, Y.; Hu, W.; Cheng, C.H.K. Targeted gene disruption in zebrafish reveals noncanonical functions of LH signaling in reproduction. Mol. Endocrinol. 2014, 28, 1785–1795. [Google Scholar] [CrossRef] [Green Version]
- So, W.-K.; Kwok, H.-F.; Ge, W. Zebrafish gonadotropins and their receptors: II. Cloning and characterization of zebrafish follicle-stimulating hormone and luteinizing hormone subunits—their spatial-temporal expression patterns and receptor specificity. Biol. Reprod. 2005, 72, 1382–1396. [Google Scholar] [CrossRef] [Green Version]
- Ishikawa, Y. Medakafish as a model system for vertebrate developmental genetics. Bioessays 2000, 22, 487–495. [Google Scholar] [CrossRef]
- Furukawa, F.; Hamasaki, S.; Hara, S.; Uchimura, T.; Shiraishi, E.; Osafune, N.; Takagi, H.; Yazawa, T.; Kamei, Y.; Kitano, T. Heat shock factor 1 protects germ cell proliferation during early ovarian differentiation in medaka. Sci. Rep. 2019, 9, 6927. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sawamura, R.; Osafune, N.; Murakami, T.; Furukawa, F.; Kitano, T. Generation of biallelic F0 mutants in medaka using the CRISPR/Cas9 system. Genes Cells 2017, 22, 756–763. [Google Scholar] [CrossRef] [Green Version]
- Matsuda, M.; Nagahama, Y.; Shinomiya, A.; Sato, T.; Matsuda, C.; Kobayashi, T.; Morrey, C.E.; Shibata, N.; Asakawa, S.; Shimizu, N.; et al. DMY is a Y-specific DM-domain gene required for male development in the medaka fish. Nature 2002, 417, 559–563. [Google Scholar] [CrossRef]
- Nanda, I.; Kondo, M.; Hornung, U.; Asakawa, S.; Winkler, C.; Shimizu, A.; Shan, Z.; Haaf, T.; Shimizu, N.; Shima, A.; et al. A duplicated copy of DMRT1 in the sex-determining region of the Y chromosome of the medaka, Oryzias latipes. Proc. Natl. Acad. Sci. USA 2002, 99, 11778–11783. [Google Scholar] [CrossRef] [Green Version]
- Matsuda, M.; Shinomiya, A.; Kinoshita, M.; Suzuki, A.; Kobayashi, T.; Paul-Prasanth, B.; Lau, E.-L.; Hamaguchi, S.; Sakaizumi, M.; Nagahama, Y. DMY gene induces male development in genetically female (XX) medaka fish. Proc. Natl. Acad. Sci. USA 2007, 104, 3865–3870. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cermak, T.; Doyle, E.L.; Christian, M.; Wang, L.; Zhang, Y.; Schmidt, C.; Baller, J.A.; Somia, N.V.; Bogdanove, A.J.; Voytas, D.F. Efficient design and assembly of custom TALEN and other TAL effector-based constructs for DNA targeting. Nucleic Acids Res. 2011, 39, e82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayashi, Y.; Kobira, H.; Yamaguchi, T.; Shiraishi, E.; Yazawa, T.; Hirai, T.; Kamei, Y.; Kitano, T. High temperature causes masculinization of genetically female medaka by elevation of cortisol. Mol. Reprod. Dev. 2010, 77, 679–686. [Google Scholar] [CrossRef] [PubMed]
- Kitano, T.; Takamune, K.; Kobayashi, T.; Nagahama, Y.; Abe, S.-I. Suppression of P450 aromatase gene expression in sex-reversed males produced by rearing genetically female larvae at a high water temperature during a period of sex differentiation in the Japanese flounder (Paralichthys olivaceus). J. Mol. Endocrinol. 1999, 23, 167–176. [Google Scholar] [CrossRef]
- Yamada, H.; Satoh, R.-I.; Ogoh, M.; Takaji, K.; Fujimoto, Y.; Hakuba, T.; Chiba, H.; Kambegawa, A.; Iwata, M. Circadian changes in serum concentrations of steroids in Japanese char Salvelinus leucomaenis at the stage of final maturation. Zool. Sci. 2002, 19, 891–898. [Google Scholar] [CrossRef]
- Iwamatsu, T. Stages of normal development in the medaka Oryzias latipes. Mech. Dev. 2004, 121, 605–618. [Google Scholar] [CrossRef]
- Kawai, T.; Yoshimura, A.; Oka, Y. Neurones in the preoptic area of the male goldfish are activated by a sex pheromone 17α,20β-dihydroxy-4-pregnen-3-one. J. Neuroendocrinol. 2015, 27, 123–130. [Google Scholar] [CrossRef]
- Nagahama, Y.; Yamashita, M. Regulation of oocyte maturation in fish. Dev. Growth Differ. 2008, 50, S195–S219. [Google Scholar] [CrossRef]
- Li, J.; Chu, L.; Sun, X.; Liu, Y.; Cheng, C.H. IGFs mediate the action of LH on oocyte maturation in zebrafish. Mol. Endocrinol. 2015, 29, 373–383. [Google Scholar] [CrossRef] [Green Version]
- Silva, J.R.V.; Figueiredo, J.R.; van den Hurk, R. Involvement of growth hormone (GH) and insulin-like growth factor (IGF) system in ovarian folliculogenesis. Theriogenology 2009, 71, 1193–1208. [Google Scholar] [CrossRef]
- Zhang, Z.; Lau, S.-W.; Zhang, L.; Ge, W. Disruption of zebrafish follicle-stimulating hormone receptor (fshr) but not luteinizing hormone receptor (lhcgr) gene by TALEN leads to failed follicle activation in females followed by sexual reversal to males. Endocrinology 2015, 156, 3747–3762. [Google Scholar] [CrossRef] [PubMed]
- Hara, A.; Hiramatsu, N.; Fujita, T. Vitellogenesis and choriogenesis in fishes. Fish. Sci. 2016, 82, 187–202. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Zhu, B.; Ge, W. Genetic analysis of zebrafish gonadotropin (FSH and LH) functions by TALEN-mediated gene disruption. Mol. Endocrinol. 2015, 29, 76–98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levavi-Sivan, B.; Bogerd, J.; Mañanós, E.; Gómez, A.; Lareyre, J. Perspectives on fish gonadotropins and their receptors. Gen. Comp. Endocrinol. 2010, 165, 412–437. [Google Scholar] [CrossRef] [PubMed]
- Aizen, J.; Kowalsman, N.; Kobayashi, M.; Hollander, L.; Sohn, Y.C.; Yoshizaki, G.; Niv, M.Y.; Levavi-Sivan, B. Experimental and computational study of inter- and intra- species specificity of gonadotropins for various gonadotropin receptors. Mol. Cell. Endocrinol. 2012, 364, 89–100. [Google Scholar] [CrossRef]
- Schulz, R.W.; de França, L.R.; Lareyre, J.J.; Le Gac, F.; Chiarini-Garcia, H.; Nobrega, R.H.; Miura, T. Spermatogenesis in fish. Gen. Comp. Endocrinol. 2010, 165, 390–411. [Google Scholar] [CrossRef]
- Planas, J. Maturation-associated changes in the response of the salmon testis to the steroidogenic actions of gonadotropins (GTH I and GTH II) in vitro. Biol. Reprod. 1995, 52, 697–704. [Google Scholar] [CrossRef] [Green Version]
- Kamei, H.; Kaneko, T.; Aida, K. In vivo gonadotropic effects of recombinant Japanese eel follicle-stimulating hormone. Aquaculture 2006, 261, 771–775. [Google Scholar] [CrossRef]
- Kazeto, Y.; Kohara, M.; Miura, T.; Miura, C.; Yamaguchi, S.; Trant, J.M.; Adachi, S.; Yamauchi, K. Japanese eel follicle-stimulating hormone (Fsh) and luteinizing hormone (Lh): Production of biologically active recombinant Fsh and Lh by Drosophila S2 cells and their differential actions on the reproductive biology. Biol. Reprod. 2008, 79, 938–9466. [Google Scholar] [CrossRef] [Green Version]
- Burow, S.; Mizrahi, N.; Maugars, G.; von Krogh, K.; Nourizadeh-Lillabadi, R.; Hollander-Cohen, L.; Shpilman, M.; Atre, I.; Weltzien, F.-A.; Levavi-Sivan, B. Characterization of gonadotropin receptors Fshr and Lhr in Japanese medaka, Oryzias latipes. Gen. Comp. Endocrinol. 2020, 285, 113276. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).