
Dermal Telocytes: A Different Viewpoint of Skin Repairing and Regeneration

 ,
, 
Abstract

{kind=link}
{kind=link}
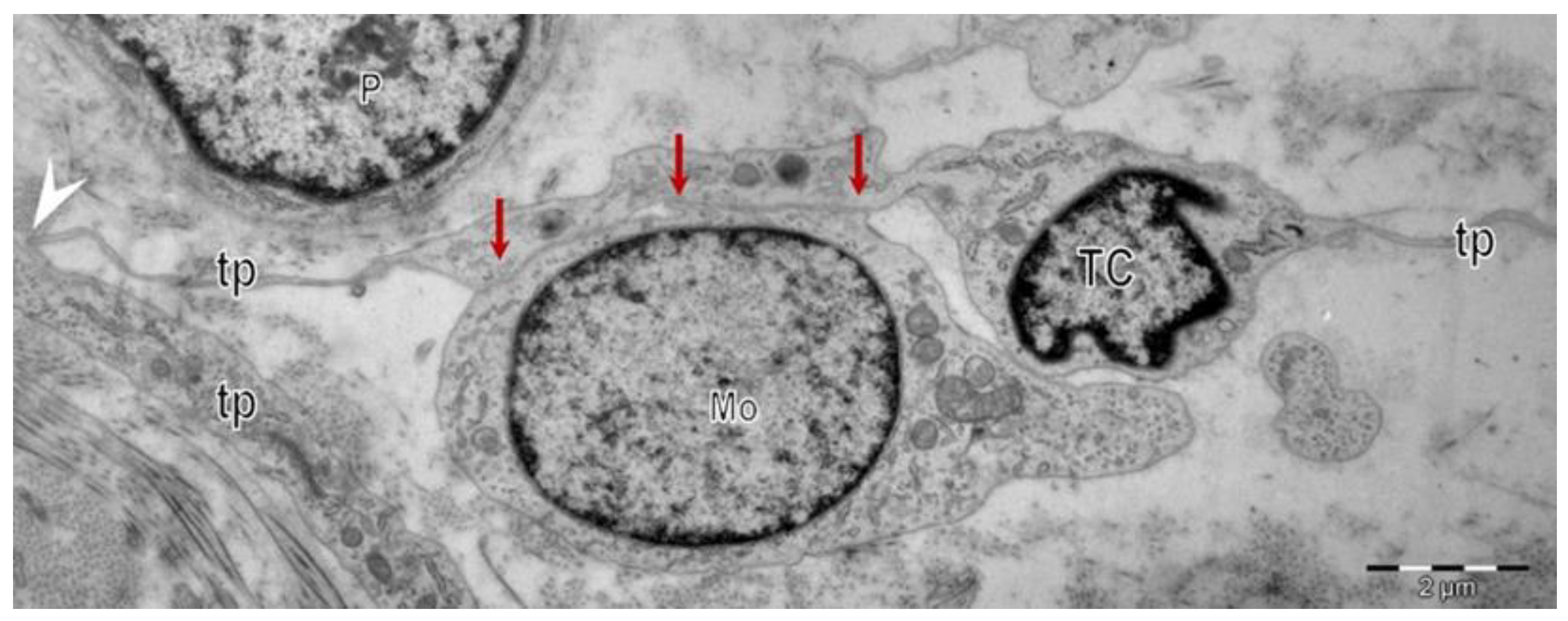{kind=link}
1. Introduction
2. Telocytes
3. Skin TCs
3.1. Location and Morphology of Skin TCs
3.2. Immunohistochemical Profile of Skin TCs
3.3. Skin TCs: Stromal Inter-/Intra- Relations
3.4. Presumptive Roles of Skin TCs
4. Skin TCs in Dermatologic Pathology
5. Skin TCs in Aging, Injury and Repair: Are TCs Key Players?
6. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Morrison, I.; Löken, L.S.; Olausson, H. The skin as a social organ. Exp. Brain Res. 2009, 204, 305–314. [Google Scholar] [CrossRef] [PubMed]
- Dai, C.; Shih, S.; Khachemoune, A. Skin substitutes for acute and chronic wound healing: An updated review. J. Dermatol. Treat. 2020, 31, 639–648. [Google Scholar] [CrossRef] [PubMed]
- Tottoli, E.M.; Dorati, R.; Genta, I.; Chiesa, E.; Pisani, S.; Conti, B. Skin Wound Healing Process and New Emerging Technologies for Skin Wound Care and Regeneration. Pharmaceutics 2020, 12, 735. [Google Scholar] [CrossRef]
- Yeganeh, P.M.; Tahmasebi, S.; Esmaeilzadeh, A. Cellular and biological factors involved in healing wounds and burns and treatment options in tissue engineering. Regen. Med. 2022, 17, 401–418. [Google Scholar] [CrossRef] [PubMed]
- Popescu, L.M.; Hinescu, M.E.; Ionescu, N.; Ciontea, S.M.; Cretoiu, D.; Ardeleanu, C. Interstitial cells of Cajal in pancreas. J. Cell. Mol. Med. 2005, 9, 169–190. [Google Scholar] [CrossRef]
- Yonghong, S.; Ruizhi, W.; Yue, Z.; Xuebing, B.; Tarique, I.; Chunhua, L.; Ping, Y.; Qiusheng, C. Telocytes in Different Organs of Vertebrates: Potential Essence Cells of the Meridian in Chinese Traditional Medicine. Microsc. Microanal. 2020, 26, 575–588. [Google Scholar] [CrossRef] [PubMed]
- Faussone Pellegrini, M.-S.; Popescu, L.M. Telocytes. Biomol. Concepts 2011, 2, 481–489. [Google Scholar] [CrossRef]
- Wang, F.; Song, Y.; Bei, Y.; Zhao, Y.; Xiao, J.; Yang, C. Telocytes in liver regeneration: Possible roles. J. Cell. Mol. Med. 2014, 18, 1720–1726. [Google Scholar] [CrossRef] [PubMed]
- Suciu, L.; Nicolescu, M.I.; Popescu, L.M. Cardiac telocytes: Serial dynamic images in cell culture. J. Cell. Mol. Med. 2010, 14, 2687–2692. [Google Scholar] [CrossRef]
- Gherghiceanu, M.; Manole, C.; Popescu, L. Telocytes in endocardium: Electron Microscope Evidence. J. Cell. Mol. Med. 2010, 14, 2330–2334. [Google Scholar] [CrossRef]
- Suciu, L.; Popescu, L.M.; Gherghiceanu, M.; Regalia, T.; Nicolescu, M.I.; Hinescu, M.E.; Faussone-Pellegrini, M.-S. Telocytes in Human Term Placenta: Morphology and Phenotype. Cells Tissues Organs 2010, 192, 325–339. [Google Scholar] [CrossRef]
- Popescu, L.M.; Faussone-Pellegrini, M.-S. TELOCYTES - a case of serendipity: The winding way from Interstitial Cells of Cajal (ICC), via Interstitial Cajal-Like Cells (ICLC) to TELOCYTES. J. Cell. Mol. Med. 2010, 14, 729–740. [Google Scholar] [CrossRef] [PubMed]
- Gherghiceanu, M.; Popescu, L.M. Interstitial Cajal-like cells (ICLC) in human resting mammary gland stroma. Transmission electron microscope (TEM) identification. J. Cell. Mol. Med. 2005, 9, 893–910. [Google Scholar] [CrossRef] [PubMed]
- Hinescu, M.E.; Popescu, L.M. Interstitial Cajal-like cells (ICLC) in human atrial myocardium. J. Cell. Mol. Med. 2005, 9, 972–975. [Google Scholar] [CrossRef] [PubMed]
- Hinescu, M.E.; Ardeleanu, C.; Gherghiceanu, M.; Popescu, L.M. Interstitial Cajal-like cells in human gallbladder. Histochem. J. 2007, 38, 275–284. [Google Scholar] [CrossRef]
- Hinescu, M.E.; Popescu, L.M.; Gherghiceanu, M.; Faussone-Pellegrini, M.-S. Interstitial Cajal-like cells in rat mesentery: An ultrastructural and immunohistochemical approach. J. Cell. Mol. Med. 2007, 12, 260–270. [Google Scholar] [CrossRef]
- Gherghiceanu, M.; Hinescu, M.E.; Andrei, F.; Mandache, E.; Macarie, C.E.; Faussone-Pellegrini, M.-S.; Popescu, L.M. Interstitial Cajal-like cells (ICLC) in myocardial sleeves of human pulmonary veins. J. Cell. Mol. Med. 2008, 12, 1777–1781. [Google Scholar] [CrossRef] [PubMed]
- Gherghiceanu, M.; Hinescu, M.E.; Popescu, L.M. Myocardial interstitial Cajal-like cells (ICLC) in caveolin-1 KO mice. J. Cell. Mol. Med. 2009, 13, 202–206. [Google Scholar] [CrossRef] [PubMed]
- Suciu, L.; Popescu, L.M.; Regalia, T.; Ardelean, A.; Manole, C.G. Epicardium: Interstitial Cajal-like cells (ICLC) highlighted by immunofluorescence. J. Cell. Mol. Med. 2009, 13, 771–777. [Google Scholar] [CrossRef] [PubMed]
- Aleksandrovych, V.; Pasternak, A.; Basta, P.; Sajewicz, M.; Walocha, J.A.; Gil, K. Telocytes: Facts, speculations and myths (Review article). Folia Med. Crac. 2017, 57, 5–22. [Google Scholar]
- Suciu, L.; Popescu, L.M.; Gherghiceanu, M. Human placenta: De visu demonstration of interstitial Cajal-like cells. J. Cell. Mol. Med. 2007, 11, 590–597. [Google Scholar] [CrossRef] [PubMed][Green Version]
- McCarthy, N.; Kraiczy, J.; Shivdasani, R.A. Cellular and molecular architecture of the intestinal stem cell niche. Nature 2020, 22, 1033–1041. [Google Scholar] [CrossRef] [PubMed]
- Ceafalan, L.; Gherghiceanu, M.; Popescu, L.M.; Simionescu, O. Telocytes in human skin--are they involved in skin regeneration? J. Cell. Mol. Med. 2012, 16, 1405–1420. [Google Scholar] [CrossRef] [PubMed]
- Horch, R.E.; Kneser, U.; Polykandriotis, E.; Schmidt, V.J.; Sun, J.; Arkudas, A. Tissue Engineering and Regenerative Medicine -Where Do We Stand? J. Cell. Mol. Med. 2012, 16, 1157–1165. [Google Scholar] [CrossRef]
- Xiao, J.; Bei, Y. Decoding Telocytes. Adv. Exp. Med. Biol. 2016, 913, 23–39. [Google Scholar] [CrossRef]
- Díaz-Flores, L.; Gutiérrez, R.; García, M.; González-Gómez, M.; Rodríguez-Rodriguez, R.; Hernández-León, N.; Díaz-Flores, L.; Carrasco, J. Cd34+ Stromal Cells/Telocytes in Normal and Pathological Skin. Int. J. Mol. Sci. 2021, 22, 7342. [Google Scholar] [CrossRef]
- Rosa, I.; Romano, E.; Fioretto, B.S.; Guasti, D.; Ibba-Manneschi, L.; Matucci-Cerinic, M.; Manetti, M. Scleroderma-like Impairment in the Network of Telocytes/CD34+ Stromal Cells in the Experimental Mouse Model of Bleomycin-Induced Dermal Fibrosis. Int. J. Mol. Sci. 2021, 22, 12407. [Google Scholar] [CrossRef] [PubMed]
- Manole, C.G.; Simionescu, O. The Cutaneous Telocytes. Telocytes 2016, 913, 303–323. [Google Scholar] [CrossRef]
- Eyden, B. Translational Medicine: The myofibroblast: Phenotypic characterization as a prerequisite to understanding its functions in translational medicine. J. Cell. Mol. Med. 2007, 12, 22–37. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Xiao, L.; Zhang, R.; Jin, H.; Shi, H. Ultrastructural and immunohistochemical characteristics of telocytes in human scalp tissue. Sci. Rep. 2020, 10, 1–7. [Google Scholar] [CrossRef]
- Manole, C.; Gherghiceanu, M.; Simionescu, O. Telocyte dynamics in psoriasis. J. Cell. Mol. Med. 2015, 19, 1504–1519. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.-E.; Fei, L.; Yin, W.-C.; Coquenlorge, S.; Rao-Bhatia, A.; Zhang, X.; Shi, S.S.W.; Lee, J.H.; Hahn, N.A.; Rizvi, W.; et al. Single cell and genetic analyses reveal conserved populations and signaling mechanisms of gastrointestinal stromal niches. Nat. Commun. 2020, 11, 334. [Google Scholar] [CrossRef]
- Mustafa, F.E.-Z.A.; Abdel-maksoud, F.M.; Hassan, A.H.S.; Mokhtar, D.M. Melatonin Induces a Stimulatory Action on the Scrotal Skin Components of Soay Ram in the Non-Breeding Season. Sci. Rep. 2020, 10, 10154. [Google Scholar] [CrossRef] [PubMed]
- Milia, A.F.; Ruffo, M.; Manetti, M.; Rosa, I.; Conte, D.; Fazi, M.; Messerini, L.; Ibba-Manneschi, L. Telocytes in Crohn’s disease. J. Cell. Mol. Med. 2013, 17, 1525–1536. [Google Scholar] [CrossRef] [PubMed]
- Manetti, M.; Rosa, I.; Messerini, L.; Ibba-Manneschi, L. Telocytes are reduced during fibrotic remodelling of the colonic wall in ulcerative colitis. J. Cell. Mol. Med. 2015, 19, 62–73. [Google Scholar] [CrossRef] [PubMed]
- Kang, Y.; Zhu, Z.; Zheng, Y.; Wan, W.; Manole, C.G.; Zhang, Q. Skin telocytes versus fibroblasts: Two distinct dermal cell populations. J. Cell. Mol. Med. 2015, 19, 2530–2539. [Google Scholar] [CrossRef]
- Vannucchi, M.G.; Faussone-Pellegrini, M.S. The Telocyte Subtypes. Adv. Exp. Med. Biol. 2016, 913, 115–126. [Google Scholar] [CrossRef] [PubMed]
- Rosa, I.; Taverna, C.; Novelli, L.; Marini, M.; Ibba-Manneschi, L.; Manetti, M. Telocytes constitute a widespread interstitial meshwork in the lamina propria and underlying striated muscle of human tongue. Sci. Rep. 2019, 9, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Vannucchi, M.; Traini, C.; Guasti, D.; Giulio, D.P.; Faussone-Pellegrini, M. Telocytes subtypes in human urinary bladder. J. Cell. Mol. Med. 2014, 18, 2000–2008. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Zeng, J.; Huang, Y.; Gong, M.; Ye, Y.; Zhao, H.; Chen, Z.; Zhang, H. Telocytes and their structural relationships with surrounding cell types in the skin of silky fowl by immunohistochemistrical, transmission electron microscopical and morphometric analysis. Poult. Sci. 2021, 100, 101367. [Google Scholar] [CrossRef] [PubMed]
- Jarrar, A.; Chumakova, A.; Hitomi, M.; Lathia, J. Enrichment and Interrogation of Cancer. Cancer Stem Cells 2016, 59–98. [Google Scholar] [CrossRef]
- Alexeev, V.; Yoon, K. Distinctive Role of the cKit Receptor Tyrosine Kinase Signaling in Mammalian Melanocytes. J. Investig. Dermatol. 2006, 126, 1102–1110. [Google Scholar] [CrossRef]
- Manetti, M.; Guiducci, S.; Ruffo, M.; Rosa, I.; Faussone-Pellegrini, M.S.; Matucci-Cerinic, M.; Ibba-Manneschi, L. Evidence for Progressive Reduction and Loss of Telocytes in the Dermal Cellular Network of Systemic Sclerosis. J. Cell. Mol. Med. 2013, 17, 482–496. [Google Scholar] [CrossRef] [PubMed]
- Matei, A.E.; Distler, J.H.W. Response to: ‘In Search for the Ideal Anatomical Composition of Vascularised Human Skin Equivalents for Systemic Sclerosis Translational Research: Should We Recruit the Telocytes?’ By Manetti and Matucci-Cerinic. Ann. Rheum. Dis. 2021, 80, e150. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Song, D.; Wei, C.; Chen, C.; Yang, Y.; Deng, X.; Gu, J. Telocytes inhibited inflammatory factor expression and enhanced cell migration in LPS-induced skin wound healing models in vitro and in vivo. J. Transl. Med. 2020, 18, 60. [Google Scholar] [CrossRef] [PubMed]
- Romano, E.; Rosa, I.; Fioretto, B.S.; Lucattelli, E.; Innocenti, M.; Ibba-Manneschi, L.; Matucci-Cerinic, M.; Manetti, M. A Two-Step Immunomagnetic Microbead-Based Method for the Isolation of Human Primary Skin Telocytes/CD34+ Stromal Cells. Int. J. Mol. Sci. 2020, 21, 5877. [Google Scholar] [CrossRef]
- Rosa, I.; Marini, M.; Manetti, M. Telocytes: An Emerging Component of Stem Cell Niche Microenvironment. J. Histochem. Cytochem. 2021, 69, 795–818. [Google Scholar] [CrossRef]
- Soliman, S.A. Telocytes are major constituents of the angiogenic apparatus. Sci. Rep. 2021, 11, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Cismasiu, V.B.; Popescu, L.M. Telocytes transfer extracellular vesicles loaded with microRNAs to stem cells. J. Cell. Mol. Med. 2015, 19, 351–358. [Google Scholar] [CrossRef] [PubMed]
- Smythies, J.; Edelstein, L. Telocytes, Exosomes, Gap Junctions and the Cytoskeleton: The Makings of a Primitive Nervous System? Front. Cell. Neurosci. 2014, 7, 278. [Google Scholar] [CrossRef]
- Díaz-Flores, L.; Gutiérrez, R.; Goméz, M.G.; Sáez, F.J.; Madrid, J.F. Behaviour of telocytes during physiopathological activation. Semin. Cell Dev. Biol. 2016, 55, 50–61. [Google Scholar] [CrossRef] [PubMed]
- Kondo, A.; Kaestner, K.H. Emerging Diverse Roles of Telocytes. Development 2019, 146, dev175018. [Google Scholar] [CrossRef] [PubMed]
- Manetti, M.; Matucci-Cerinic, M. In search for the ideal anatomical composition of vascularised human skin equivalents for systemic sclerosis translational research: Should we recruit the telocytes? Ann Rheum Dis. 2021, 80, e149. [Google Scholar] [CrossRef] [PubMed]
- Manole, C.G.; Cismaşiu, V.; Gherghiceanu, M.; Popescu, L.M. Experimental acute myocardial infarction: Telocytes involvement in neo-angiogenesis. J. Cell. Mol. Med. 2011, 15, 2284–2296. [Google Scholar] [CrossRef] [PubMed]
- Fertig, E.T.; Gherghiceanu, M.; Popescu, L.M. Extracellular vesicles release by cardiac telocytes: Electron microscopy and electron tomography. J. Cell. Mol. Med. 2014, 18, 1938–1943. [Google Scholar] [CrossRef] [PubMed]
- Meng, X.; Zhu, Z.; Ahmed, N.; Ma, Q.; Wang, Q.; Deng, B.; Chen, Q.; Lu, Y.; Yang, P. Dermal Microvascular Units in Domestic Pigs (Sus scrofa domestica): Role as Transdermal Passive Immune Channels. Front. Vet. Sci. 2022, 9. [Google Scholar] [CrossRef]
- Moisan, F.; Oucherif, S.; Kaulanjan-Checkmodine, P.; Prey, S.; Rousseau, B.; Bonneu, M.; Claverol, S.; Gontier, E.; Lacomme, S.; Dousset, L.; et al. Critical Role of Aquaporin-1 and Telocytes in Infantile Hemangioma Response to Propranolol Beta Blockade. Proc. Natl. Acad. Sci. USA 2021, 118, e2018690118. [Google Scholar] [CrossRef]
- Bei, Y.; Wang, F.; Yang, C.; Xiao, J. Telocytes in regenerative medicine. J. Cell. Mol. Med. 2015, 19, 1441–1454. [Google Scholar] [CrossRef]
- Díaz-Flores, L.; Gutiérrez, R.; García, M.P.; Sáez, F.; Aparicio, F.; Madrid, J.F. Uptake and intracytoplasmic storage of pigmented particles by human CD34+ stromal cells/telocytes: Endocytic property of telocytes. J. Cell. Mol. Med. 2014, 18, 2478–2487. [Google Scholar] [CrossRef] [PubMed]
- Diaz-Flores, L.; Gutiérrez, R.; García, M.P.; González, M.; Madrid, J.F. Telocytes as a Source of Progenitor Cells in Regeneration and Repair Through Granulation Tissue. Curr. Stem Cell Res. Ther. 2016, 11, 395–403. [Google Scholar] [CrossRef]
- Edelstein, L.; Smythies, J. The Role of Telocytes in Morphogenetic Bioelectrical Signaling: Once More unto the Breach. Front. Mol. Neurosci. 2014, 7, 41. [Google Scholar] [CrossRef] [PubMed]
- Varga, I.; Polák, Š.; Kyselovič, J.; Kachlík, D.; Danišovič, Ľ.; Klein, M. Recently Discovered Interstitial Cell Population of Telocytes: Distinguishing Facts from Fiction Regarding Their Role in the Pathogenesis of Diverse Diseases Called “Telocytopathies”. Medicina 2019, 55, 56. [Google Scholar] [CrossRef]
- Aleksandrovych, V.; Gil, A.; Wrona, A. Sex steroid hormone receptors of telocytes—potential key role in leiomyoma development. Folia Med. Crac. 2020, 60, 81–95. [Google Scholar]
- Gherghiceanu, M.; Popescu, L.M. Cardiomyocyte precursors and telocytes in epicardial stem cell niche: Electron microscope images. J. Cell. Mol. Med. 2010, 14, 871–877. [Google Scholar] [CrossRef]
- Popescu, L.M.; Gherghiceanu, M.; Suciu, L.C.; Manole, C.G.; Hinescu, M.E. Telocytes and putative stem cells in the lungs: Electron microscopy, electron tomography and laser scanning microscopy. Cell Tissue Res. 2011, 345, 391–403. [Google Scholar] [CrossRef] [PubMed]
- Liao, Z.; Chen, Y.; Duan, C.; Zhu, K.; Huang, R.; Zhao, H.; Hintze, M.; Pu, Q.; Yuan, Z.; Lv, L.; et al. Cardiac telocytes inhibit cardiac microvascular endothelial cell apoptosis through exosomal miRNA-21-5p-targeted cdip1 silencing to improve angiogenesis following myocardial infarction. Theranostics 2021, 11, 268–291. [Google Scholar] [CrossRef]
- Boroujerdi, A.; Welser-Alves, J.V.; Milner, R. Matrix metalloproteinase-9 mediates post-hypoxic vascular pruning of cerebral blood vessels by degrading laminin and claudin-5. Angiogenesis 2015, 18, 255–264. [Google Scholar] [CrossRef] [PubMed]
- Vannucchi, M. The Telocytes: Ten Years after Their Introduction in the Scientific Literature. An Update on Their Morphology, Distribution, and Potential Roles in the Gut. Int. J. Mol. Sci. 2020, 21, 4478. [Google Scholar] [CrossRef] [PubMed]
- Bahar Halpern, K.; Massalha, H.; Zwick, R.K.; Moor, A.E.; Castillo-Azofeifa, D.; Rozenberg, M.; Farack, L.; Egozi, A.; Miller, D.R.; Averbukh, I.; et al. Lgr5+ Telocytes Are a Signaling Source at the Intestinal Villus Tip. Nat. Commun. 2020, 11, 1936. [Google Scholar] [CrossRef] [PubMed]
- Shoshkes-Carmel, M.; Wang, Y.J.; Wangensteen, K.J.; Tóth, B.; Kondo, A.; Massasa, E.E.; Itzkovitz, S.; Kaestner, K.H. Subepithelial Telocytes Are an Important Source of Wnts That Supports Intestinal Crypts. Nature 2018, 557, 242–246. [Google Scholar] [CrossRef] [PubMed]
- Samuelson, L.C. Debate over the identity of an intestinal niche-cell population settled. Nature 2018, 558, 380–381. [Google Scholar] [CrossRef] [PubMed]
- Cucu, I.; Nicolescu, M.I.; Busnatu, Ș.-S.; Manole, C.G. Dynamic Involvement of Telocytes in Modulating Multiple Signaling Pathways in Cardiac Cytoarchitecture. Int. J. Mol. Sci. 2022, 23, 5769. [Google Scholar] [CrossRef] [PubMed]
- Mirancea, N.; Moroşanu, A.-M.; Mirancea, G.-V.; Juravle, F.D.; Mănoiu, V.S. Infrastructure of the telocytes from tumor stroma in the skin basal and squamous cell carcinomas. Romanian J. Morphol. Embryol. 2013, 54. [Google Scholar]
- Zhang, S.; Duan, E. Fighting against Skin Aging: The Way from Bench to Bedside. Cell Transplant. 2018, 27, 729–738. [Google Scholar] [CrossRef] [PubMed]
- Makrantonaki, E.; Bekou, V.; Zouboulis, C.C. Genetics and skin aging. Dermato Endocrinol. 2012, 4, 280–284. [Google Scholar] [CrossRef]
- Russell-Goldman, E.; Murphy, G.F. The Pathobiology of Skin Aging. Am. J. Pathol. 2020, 190, 1356–1369. [Google Scholar] [CrossRef] [PubMed]
- Glass, D.; Viñuela, A.; Davies, M.N.; Ramasamy, A.; Parts, L.; Knowles, D.; Brown, A.A.; Hedman, K.; Small, K.S.; Buil, A.; et al. Gene expression changes with age in skin, adipose tissue, blood and brain. Genome Biol. 2013, 14, R75. [Google Scholar] [CrossRef] [PubMed]
- Driskell, R.R.; Watt, F.M. Understanding fibroblast heterogeneity in the skin. Trends Cell Biol. 2015, 25, 92–99. [Google Scholar] [CrossRef] [PubMed]
- Rognoni, E.; Watt, F.M. Skin Cell Heterogeneity in Development, Wound Healing, and Cancer. Trends Cell Biol. 2018, 28, 709–722. [Google Scholar] [CrossRef] [PubMed]
- Martin, P.; Nunan, R. Cellular and molecular mechanisms of repair in acute and chronic wound healing. Br. J. Dermatol. 2015, 173, 370–378. [Google Scholar] [CrossRef] [PubMed]
- Zhao, P.; Sui, B.-D.; Liu, N.; Lv, Y.-J.; Zheng, C.-X.; Lu, Y.-B.; Huang, W.-T.; Zhou, C.-H.; Chen, J.; Pang, D.-L.; et al. Anti-aging pharmacology in cutaneous wound healing: Effects of metformin, resveratrol, and rapamycin by local application. Aging Cell 2017, 16, 1083–1093. [Google Scholar] [CrossRef] [PubMed]
- Rittié, L. Cellular mechanisms of skin repair in humans and other mammals. J. Cell Commun. Signal. 2016, 10, 103–120. [Google Scholar] [CrossRef] [PubMed]
- Eming, S.A.; Martin, P.; Tomic-Canic, M. Wound repair and regeneration: Mechanisms, signaling, and translation. Sci. Transl. Med. 2014, 6, 265sr6. [Google Scholar] [CrossRef] [PubMed]
- Komi, D.E.A.; Khomtchouk, K.; Santa Maria, P.L. A Review of the Contribution of Mast Cells in Wound Healing: Involved Molecular and Cellular Mechanisms. Clin. Rev. Allergy Immunol. 2020, 58, 298–312. [Google Scholar] [CrossRef] [PubMed]
- Chester, D.; Brown, A.C. The role of biophysical properties of provisional matrix proteins in wound repair. Matrix Biol. 2017, 60–61, 124–140. [Google Scholar] [CrossRef] [PubMed]
- Landén, N.X.; Li, D.; Ståhle, M. Transition from inflammation to proliferation: A critical step during wound healing. Cell. Mol. Life Sci. 2016, 73, 3861–3885. [Google Scholar] [CrossRef]
- Diller, R.B.; Tabor, A.J. The Role of the Extracellular Matrix (ECM) in Wound Healing: A Review. Biomimetics 2022, 7, 87. [Google Scholar] [CrossRef]
- Mathew-Steiner, S.S.; Roy, S.; Sen, C.K. Collagen in Wound Healing. Bioengineering 2021, 8, 63. [Google Scholar] [CrossRef] [PubMed]
- Schultz, G.S.; Wysocki, A. Interactions between extracellular matrix and growth factors in wound healing. Wound Repair Regen. 2009, 17, 153–162. [Google Scholar] [CrossRef]
- Ganapathy, N.; Venkataraman, S.S.; Daniel, R.; Aravind, R.J.; Kumarakrishnan, V.B. Molecular biology of wound healing. J. Pharm. Bioallied Sci. 2012, 4, S334–S337. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.-M.; An, J. Cytokines, Inflammation, and Pain. Int. Anesthesiol. Clin. 2007, 45, 27–37. [Google Scholar] [CrossRef] [PubMed]
- Iavarone, C.; Catania, A.; Marinissen, M.J.; Visconti, R.; Acunzo, M.; Tarantino, C.; Carlomagno, M.S.; Bruni, C.B.; Gutkind, J.S.; Chiariello, M. The Platelet-derived Growth Factor Controls c-myc Expression through a JNK- and AP-1-dependent Signaling Pathway. J. Biol. Chem. 2003, 278, 50024–50030. [Google Scholar] [CrossRef] [PubMed]
- Krishnaswami, V.; Raju, N.S.; Alagarsamy, S.; Kandasamy, R. Novel Nanocarriers for the Treatment of Wound Healing. Curr. Pharm. Des. 2020, 26, 4591–4600. [Google Scholar] [CrossRef] [PubMed]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Manole, C.G.; Gherghiceanu, M.; Ceafalan, L.C.; Hinescu, M.E. Dermal Telocytes: A Different Viewpoint of Skin Repairing and Regeneration. Cells 2022, 11, 3903. https://doi.org/10.3390/cells11233903
Manole CG, Gherghiceanu M, Ceafalan LC, Hinescu ME. Dermal Telocytes: A Different Viewpoint of Skin Repairing and Regeneration. Cells. 2022; 11(23):3903. https://doi.org/10.3390/cells11233903
Chicago/Turabian StyleManole, Catalin G., Mihaela Gherghiceanu, Laura Cristina Ceafalan, and Mihail E. Hinescu. 2022. "Dermal Telocytes: A Different Viewpoint of Skin Repairing and Regeneration" Cells 11, no. 23: 3903. https://doi.org/10.3390/cells11233903
APA StyleManole, C. G., Gherghiceanu, M., Ceafalan, L. C., & Hinescu, M. E. (2022). Dermal Telocytes: A Different Viewpoint of Skin Repairing and Regeneration. Cells, 11(23), 3903. https://doi.org/10.3390/cells11233903