

Expression and Interaction Proteomics of GluA1- and GluA3-Subunit-Containing AMPARs Reveal Distinct Protein Composition

 , ,
, , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Antibodies
2.3. Preparation of Crude Synaptosomal Fractions
2.4. Immuno-Purifications/in-Gel Digestion/Data-Dependent Acquisition Analysis
2.5. Depletion Immuno-Purifications
2.6. Immuno-Purifications/Blue Native-PAGE/Data-Dependent Acquisition Analysis
2.7. Co-Purification from HEK293 Cells
2.8. BN-PAGE/Immunoblot Analysis
2.9. Quantitative Proteomics by in-Gel Digestion/Data-Independent Acquisition
2.10. Primary Neuronal Culture
2.11. Immunocytochemistry
2.12. STED Microscopy and Analysis
3. Results
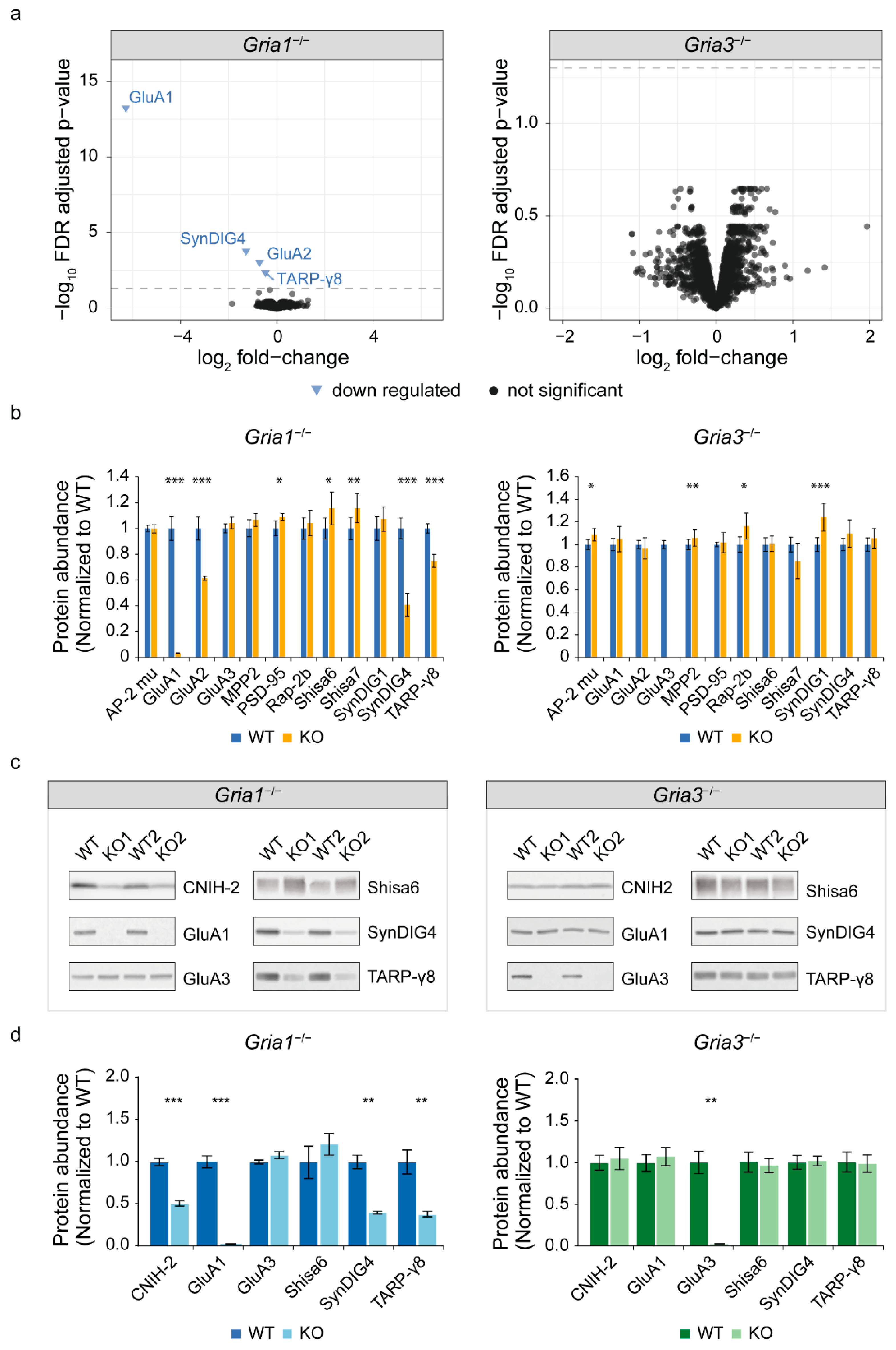
3.1. Expression Proteomics on Gria1- and Gria3 KO Synapses Reveals Differential Expression of Known AMPAR Interactors
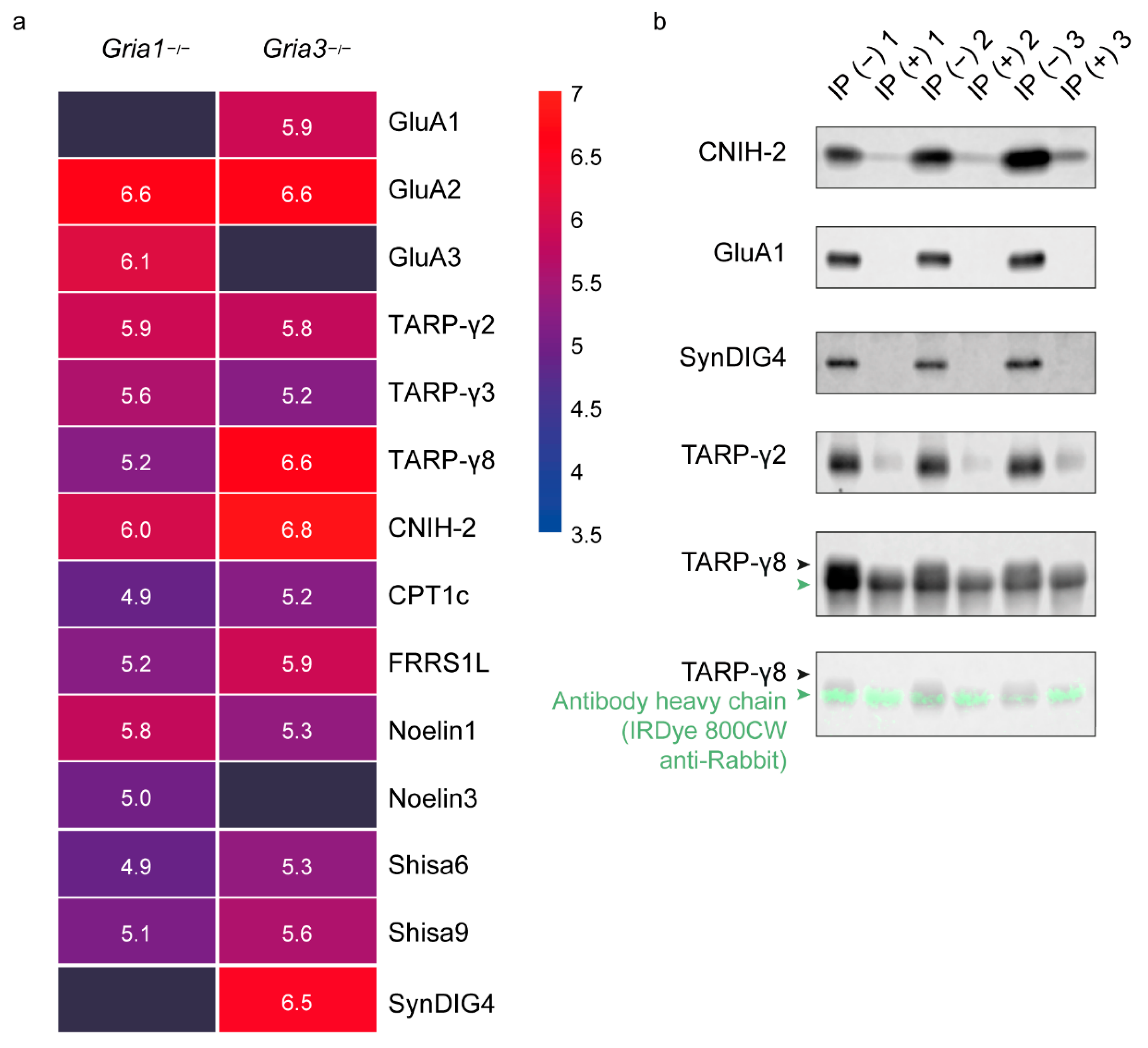
3.2. IP-MS of GluA2/3 from Gria1 and Gria3 KO Synapse Extracts Reveal Subunit-Specific Differential Interactors
3.3. Validation with Immunoblotting of GluA2/3 IP in the GluA1 Depleted Synapse Extract
3.4. Reversed IP Using Anti-SynDIG4 and Anti-TARP-γ8 Antibodies Reveal Their Co-Occurrence with the AMPAR
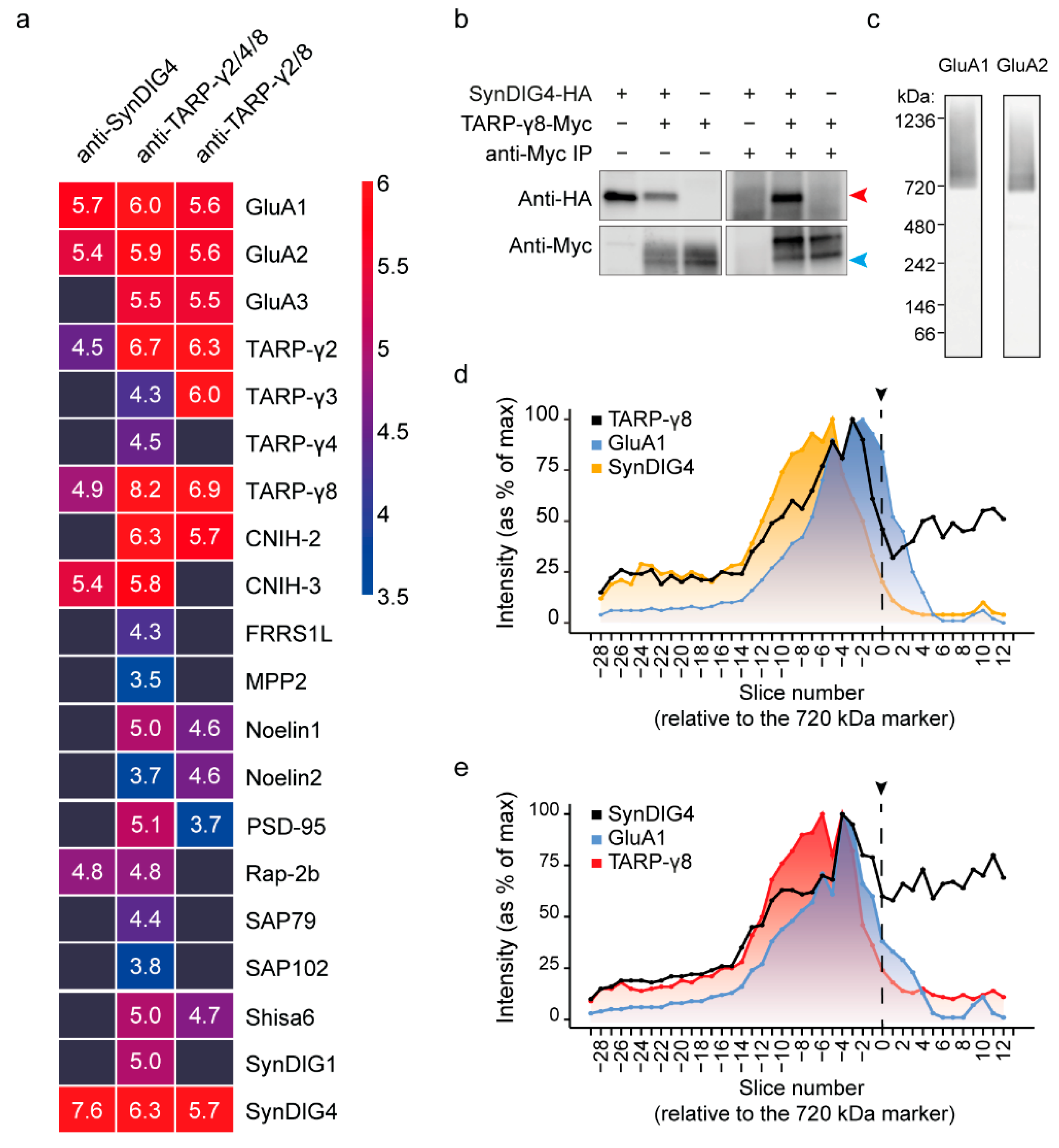
3.5. Combined IP-Blue Native Quantitative Proteomics Demonstrates the Presence of TARP-γ8 and SynDIG4 in an AMPAR Subcomplex
3.6. Super-Resolution Microscopy Shows Co-Localization of TARP-γ8 and SynDIG4 at the Synapse
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| Abbreviation | Full-Name | Gene Name |
| AP-2 complex subunit mu | Adaptor related protein complex 2 subunit mu 1 | Ap2m1 |
| CNIH-2 | Cornichon homolog 2 | Cnih2 |
| CNIH-3 | Cornichon homolog 3 | Cnih3 |
| CPT1c | Carnitine O-palmitoyltransferase 1C | Cpt1c |
| FRRS1L | Ferric chelate reductase 1 like | Frrs1l |
| GluA1 | Glutamate receptor ionotropic, AMPA 1 | Gria1 |
| GluA2 | Glutamate receptor ionotropic, AMPA 2 | Gria2 |
| GluA3 | Glutamate receptor ionotropic, AMPA 3 | Gria3 |
| GluA4 | Glutamate receptor ionotropic, AMPA 4 | Gria4 |
| GRIP-1 | Glutamate receptor-interacting protein 1 | Grip1 |
| MPP2 | MAGUK p55 subfamily member 2 | Mpp2 |
| PICK1 | Protein interacting with C kinase 1 | Pick1 |
| PSD-95 | Postsynaptic density protein- 95 | Dlg4 |
| Rap-2b | Ras-related protein Rap-2b | Rap2b |
| SAP97 | Synapse-associated protein 97 | Dlg1 |
| Shisa6 | Shisa family member 6 | Shisa6 |
| Shisa7 | Shisa family member 7 | Shisa7 |
| Shisa9 | Shisa family members 9 | Shisa9 |
| SynDIG1 | synapse differentiation-induced gene 1 | Prrt2 |
| SynDIG4 | Synapse differentiation-induced protein 4 | Prrt1 |
| TARP-γ2 | Transmembrane AMPAR regulatory protein gamma-2 | Cacng2 |
| TARP-γ3 | Transmembrane AMPAR regulatory protein gamma-3 | Cacng3 |
| TARP-γ4 | Transmembrane AMPAR regulatory protein gamma-4 | Cacng4 |
| TARP-γ8 | Transmembrane AMPAR regulatory protein gamma-8 | Cacng8 |
References
- Nair, D.; Hosy, E.; Petersen, J.D.; Constals, A.; Giannone, G.; Choquet, D.; Sibarita, J.B. Super-Resolution Imaging Reveals That AMPA Receptors Inside Synapses Are Dynamically Organized in Nanodomains Regulated by PSD95. J. Neurosci. 2013, 33, 13204–13224. [Google Scholar] [CrossRef] [PubMed]
- MacGillavry, H.D.; Song, Y.; Raghavachari, S.; Blanpied, T.A. Nanoscale scaffolding domains within the postsynaptic density concentrate synaptic AMPA receptors. Neuron 2013, 78, 615–622. [Google Scholar] [CrossRef] [PubMed]
- Shepherd, J.D.; Huganir, R.L. The cell biology of synaptic plasticity: AMPA receptor trafficking. Annu. Rev. Cell Dev. Biol. 2007, 23, 613–643. [Google Scholar] [CrossRef] [PubMed]
- Wenthold, R.J.; Petralia, R.S.; Blahos, J., II; Niedzielski, A.S. Evidence for multiple AMPA receptor complexes in hippocampal CA1/CA2 neurons. J. Neurosci. 1996, 16, 1982–1989. [Google Scholar] [CrossRef]
- Zhao, Y.; Chen, S.; Swensen, A.C.; Qian, W.-J.; Gouaux, E. Architecture and subunit arrangement of native AMPA receptors elucidated by cryo-EM. Science 2019, 364, 355–362. [Google Scholar] [CrossRef]
- Morise, J.; Suzuki, K.G.; Kitagawa, A.; Wakazono, Y.; Takamiya, K.; Tsunoyama, T.A.; Nemoto, Y.L.; Takematsu, H.; Kusumi, A.; Oka, S. AMPA receptors in the synapse turnover by monomer diffusion. Nat. Commun. 2019, 10, 5245. [Google Scholar] [CrossRef]
- Lu, W.; Shi, Y.; Jackson, A.C.; Bjorgan, K.; During, M.J.; Sprengel, R.; Seeburg, P.H.; Nicoll, R.A. Subunit composition of synaptic AMPA receptors revealed by a single-cell genetic approach. Neuron 2009, 62, 254–268. [Google Scholar] [CrossRef]
- Pandya, N.J.; Koopmans, F.; Slotman, J.A.; Paliukhovich, I.; Houtsmuller, A.B.; Smit, A.B.; Li, K.W. Correlation profiling of brain sub-cellular proteomes reveals co-assembly of synaptic proteins and subcellular distribution. Sci. Rep. 2017, 7, 12107. [Google Scholar] [CrossRef]
- Schwenk, J.; Baehrens, D.; Haupt, A.; Bildl, W.; Boudkkazi, S.; Roeper, J.; Fakler, B.; Schulte, U. Regional diversity and developmental dynamics of the AMPA-receptor proteome in the mammalian brain. Neuron 2014, 84, 41–54. [Google Scholar] [CrossRef]
- Diering, G.H.; Huganir, R.L. The AMPA Receptor Code of Synaptic Plasticity. Neuron 2018, 100, 314–329. [Google Scholar] [CrossRef]
- Andrásfalvy, B.K.; Smith, M.A.; Borchardt, T.; Sprengel, R.; Magee, J.C. Impaired regulation of synaptic strength in hippocampal neurons from GluR1-deficient mice. J. Physiol. 2003, 552, 35–45. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; Liu, A.; Xia, S.; Leung, C.; Qi, J.; Meng, Y.; Xie, W.; Park, P.; Collingridge, G.L.; Jia, Z. The C-terminal tails of endogenous GluA1 and GluA2 differentially contribute to hippocampal synaptic plasticity and learning. Nat. Neurosci. 2018, 21, 50–62. [Google Scholar] [CrossRef]
- Shi, S.; Hayashi, Y.; Esteban, J.A.; Malinow, R. Subunit-specific rules governing AMPA receptor trafficking to synapses in hippocampal pyramidal neurons. Cell 2001, 105, 331–343. [Google Scholar] [CrossRef]
- Renner, M.C.; Albers, E.H.; Gutierrez-Castellanos, N.; Reinders, N.R.; van Huijstee, A.N.; Xiong, H.; Lodder, T.R.; Kessels, H.W. Synaptic plasticity through activation of GluA3-containing AMPA-receptors. eLife 2017, 6, e25462. [Google Scholar] [CrossRef]
- Reinders, N.R.; Pao, Y.; Renner, M.C.; da Silva-Matos, C.M.; Lodder, T.R.; Malinow, R.; Kessels, H.W. Amyloid-β effects on synapses and memory require AMPA receptor subunit GluA3. Proc. Natl. Acad. Sci. USA 2016, 113, E6526–E6534. [Google Scholar] [CrossRef] [PubMed]
- Abdollahi Nejat, M.; Klaassen, R.V.; Spijker, S.; Smit, A.B. Auxiliary subunits of the AMPA receptor: The Shisa family of proteins. Curr. Opin. Pharmacol. 2021, 58, 52–61. [Google Scholar] [CrossRef]
- Schwenk, J.; Harmel, N.; Brechet, A.; Zolles, G.; Berkefeld, H.; Müller, C.S.; Bildl, W.; Baehrens, D.; Hüber, B.; Kulik, A.; et al. High-Resolution Proteomics Unravel Architecture and Molecular Diversity of Native AMPA Receptor Complexes. Neuron 2012, 74, 621–633. [Google Scholar] [CrossRef]
- Rao-Ruiz, P.; Rotaru, D.C.; van der Loo, R.J.; Mansvelder, H.D.; Stiedl, O.; Smit, A.B.; Spijker, S. Retrieval-specific endocytosis of GluA2-AMPARs underlies adaptive reconsolidation of contextual fear. Nat. Neurosci. 2011, 14, 1302–1308. [Google Scholar] [CrossRef]
- Bissen, D.; Foss, F.; Acker-Palmer, A. AMPA receptors and their minions: Auxiliary proteins in AMPA receptor trafficking. Cell. Mol. Life Sci. 2019, 76, 2133–2169. [Google Scholar] [CrossRef]
- Schwenk, J.; Harmel, N.; Zolles, G.; Bildl, W.; Kulik, A.; Heimrich, B.; Chisaka, O.; Jonas, P.; Schulte, U.; Fakler, B.; et al. Functional proteomics identify cornichon proteins as auxiliary subunits of AMPA receptors. Science 2009, 323, 1313. [Google Scholar] [CrossRef]
- Vandenberghe, W.; Nicoll, R.; Bredt, D.S. Stargazin is an AMPA receptor auxiliary subunit. Proc. Natl. Acad. Sci. USA 2005, 102, 485–490. [Google Scholar] [CrossRef] [PubMed]
- Bats, C.; Groc, L.; Choquet, D. The Interaction between Stargazin and PSD-95 Regulates AMPA Receptor Surface Trafficking. Neuron 2007, 53, 719–734. [Google Scholar] [CrossRef] [PubMed]
- Constals, A.; Penn, A.C.; Compans, B.; Toulme, E.; Phillipat, A.; Marais, S.; Retailleau, N.; Hafner, A.S.; Coussen, F.; Hosy, E.; et al. Glutamate-Induced AMPA Receptor Desensitization Increases Their Mobility and Modulates Short-Term Plasticity through Unbinding from Stargazin. Neuron 2015, 85, 787–803. [Google Scholar] [CrossRef] [PubMed]
- Milstein, A.D.; Zhou, W.; Karimzadegan, S.; Bredt, D.S.; Nicoll, R.A. TARP Subtypes Differentially and Dose-Dependently Control Synaptic AMPA Receptor Gating. Neuron 2007, 55, 905–918. [Google Scholar] [CrossRef] [PubMed]
- Boudkkazi, S.; Brechet, A.; Schwenk, J.; Fakler, B. Cornichon2 Dictates the Time Course of Excitatory Transmission at Individual Hippocampal Synapses. Neuron 2014, 82, 848–858. [Google Scholar] [CrossRef]
- Klaassen, R.V.; Stroeder, J.; Coussen, F.; Hafner, A.S.; Petersen, J.D.; Renancio, C.; Schmitz, L.J.; Normand, E.; Lodder, J.C.; Rotaru, D.C.; et al. Shisa6 traps AMPARs at postsynaptic sites and prevents their desensitization during high frequency synaptic stimulation. Nat. Commun. 2016, 7, 10682. [Google Scholar] [CrossRef]
- Von Engelhardt, J.; Mack, V.; Sprengel, R.; Kavenstock, N.; Li, K.W.; Stern-Bach, Y.; Smit, A.B.; Seeburg, P.H.; Monyer, H. CKAMP44: A brain-specific protein attenuating short-term synaptic plasticity in the dentate gyrus. Science 2010, 327, 1518–1522. [Google Scholar] [CrossRef]
- Schmitz, L.J.; Klaassen, R.V.; Ruiperez-Alonso, M.; Zamri, A.E.; Stroeder, J.; Rao-Ruiz, P.; Lodder, J.C.; van der Loo, R.J.; Mansvelder, H.D.; Smit, A.B.; et al. The AMPA receptor-associated protein Shisa7 regulates hippocampal synaptic function and contextual memory. eLife 2017, 6, e24192. [Google Scholar] [CrossRef]
- Brechet, A.; Buchert, R.; Schwenk, J.; Boudkkazi, S.; Zolles, G.; Siquier-Pernet, K.; Schaber, I.; Bildl, W.; Saadi, A.; Bole-Feysot, C.; et al. AMPA-receptor specific biogenesis complexes control synaptic transmission and intellectual ability. Nat. Commun. 2017, 8, 15910. [Google Scholar] [CrossRef]
- Schwenk, J.; Boudkkazi, S.; Kocylowski, M.K.; Brechet, A.; Zolles, G.; Bus, T.; Costa, K.; Kollewe, A.; Jordan, J.; Bank, J.; et al. An ER Assembly Line of AMPA-Receptors Controls Excitatory Neurotransmission and Its Plasticity. Neuron 2019, 104, 680–692.e9. [Google Scholar] [CrossRef]
- Gonzalez-Lozano, M.A.; Koopmans, F.; Sullivan, P.F.; Protze, J.; Krause, G.; Verhage, M.; Li, K.W.; Liu, F.; Smit, A.B. Stitching the synapse: Cross-linking mass spectrometry into resolving synaptic protein interactions. Sci. Adv. 2020, 6, eaax5783. [Google Scholar] [CrossRef] [PubMed]
- Anggono, V.; Huganir, R.L. Regulation of AMPA receptor trafficking and synaptic plasticity. Curr. Opin. Neurobiol. 2012, 22, 461–469. [Google Scholar] [CrossRef] [PubMed]
- Leonard, A.S.; Davare, M.A.; Horne, M.C.; Garner, C.C.; Hell, J.W. SAP97 is associated with the alpha-amino-3-hydroxy-5-methylisoxazole-4-propionic acid receptor GluR1 subunit. J. Biol. Chem. 1998, 273, 19518–19524. [Google Scholar] [CrossRef] [PubMed]
- Cai, C.; Coleman, S.K.; Niemi, K.; Keinänen, K. Selective binding of synapse-associated protein 97 to GluR-A alpha-amino-5-hydroxy-3-methyl-4-isoxazole propionate receptor subunit is determined by a novel sequence motif. J. Biol. Chem. 2002, 277, 31484–31490. [Google Scholar] [CrossRef] [PubMed]
- Dong, H.; O’Brien, R.J.; Fung, E.T.; Lanahan, A.A.; Worley, P.F.; Huganir, R.L. GRIP: A synaptic PDZ domain-containing protein that interacts with AMPA receptors. Nature 1997, 386, 279–284. [Google Scholar] [CrossRef]
- Xia, J.; Zhang, X.; Staudinger, J.; Huganir, R.L. Clustering of AMPA receptors by the synaptic PDZ domain-containing protein PICK1. Neuron 1999, 22, 179–187. [Google Scholar] [CrossRef]
- Osten, P.; Khatri, L.; Perez, J.L.; Köhr, G.; Giese, G.; Daly, C.; Schulz, T.W.; Wensky, A.; Lee, L.M.; Ziff, E.B. Mutagenesis reveals a role for ABP/GRIP binding to GluR2 in synaptic surface accumulation of the AMPA receptor. Neuron 2000, 27, 313–325. [Google Scholar] [CrossRef]
- Setou, M.; Seog, D.H.; Tanaka, Y.; Kanai, Y.; Takei, Y.; Kawagishi, M.; Hirokawa, N. Glutamate-receptor-interacting protein GRIP1 directly steers kinesin to dendrites. Nature 2002, 417, 83–87. [Google Scholar] [CrossRef]
- Chung, H.J.; Xia, J.; Scannevin, R.H.; Zhang, X.; Huganir, R.L. Phosphorylation of the AMPA receptor subunit GluR2 differentially regulates its interaction with PDZ domain-containing proteins. J. Neurosci. 2000, 20, 7258–7267. [Google Scholar] [CrossRef]
- Matt, L.; Kirk, L.M.; Chenaux, G.; Speca, D.J.; Puhger, K.R.; Pride, M.C.; Qneibi, M.; Haham, T.; Plambeck, K.E.; Stern-Bach, Y.; et al. SynDIG4/Prrt1 Is Required for Excitatory Synapse Development and Plasticity Underlying Cognitive Function. Cell Rep. 2018, 22, 2246–2253. [Google Scholar] [CrossRef]
- Kim, C.H.; Takamiya, K.; Petralia, R.S.; Sattler, R.; Yu, S.; Zhou, W.; Kalb, R.; Wenthold, R.; Huganir, R. Persistent hippocampal CA1 LTP in mice lacking the C-terminal PDZ ligand of GluR1. Nat. Neurosci. 2005, 8, 985–987. [Google Scholar] [CrossRef] [PubMed]
- Specht, C.G.; Schoepfer, R. Deletion of the alpha-synuclein locus in a subpopulation of C57BL/6J inbred mice. BMC Neurosci. 2001, 2, 11. [Google Scholar] [CrossRef] [PubMed]
- Chen, N.; Pandya, N.J.; Koopmans, F.; Castelo-Székelv, V.; van der Schors, R.C.; Smit, A.B.; Li, K.W. Interaction proteomics reveals brain region-specific AMPA receptor complexes. J. Proteome Res. 2014, 13, 5695–5706. [Google Scholar] [CrossRef]
- Van der Spek, S.J.; Koopmans, F.; Paliukhovich, I.; Ramsden, S.L.; Harvey, K.; Harvey, R.J.; Smit, A.B.; Li, K.W. Glycine Receptor Complex Analysis Using Immunoprecipitation-Blue Native Gel Electrophoresis-Mass Spectrometry. Proteomics 2020, 20, e1900403. [Google Scholar] [CrossRef] [PubMed]
- Koopmans, F.; Pandya, N.J.; Franke, S.K.; Phillippens, I.H.; Paliukhovich, I.; Li, K.W.; Smit, A.B. Comparative Hippocampal Synaptic Proteomes of Rodents and Primates: Differences in Neuroplasticity-Related Proteins. Front. Mol. Neurosci. 2018, 11, 364. [Google Scholar] [CrossRef] [PubMed]
- Rosato, M.; Stringer, S.; Gebuis, T.; Paliukhovich, I.; Li, K.W.; Posthuma, D.; Sullivan, P.F.; Smit, A.B.; van Kesteren, R.E. Combined cellomics and proteomics analysis reveals shared neuronal morphology and molecular pathway phenotypes for multiple schizophrenia risk genes. Mol. Psychiatry 2021, 26, 784–799. [Google Scholar] [CrossRef]
- Van der Spek, S.J.; Gonzalez-Lozano, M.A.; Koopmans, F.; Miedema, S.S.; Paliukhovich, I.; Smit, A.B.; Li, K.W. Age-Dependent Hippocampal Proteomics in the APP/PS1 Alzheimer Mouse Model: A Comparative Analysis with Classical SWATH/DIA and directDIA Approaches. Cells 2021, 10, 1588. [Google Scholar] [CrossRef]
- Herguedas, B.; Watson, J.F.; Ho, H.; Cais, O.; García-Nafría, J.; Greger, I.H. Architecture of the heteromeric GluA1/2 AMPA receptor in complex with the auxiliary subunit TARP γ8. Science 2019, 364, eaav9011. [Google Scholar] [CrossRef]
- Lu, J.; Helton, T.D.; Blanpied, T.A.; Rácz, B.; Newpher, T.M.; Weinberg, R.J.; Ehlers, M.D. Postsynaptic positioning of endocytic zones and AMPA receptor cycling by physical coupling of dynamin-3 to Homer. Neuron 2007, 55, 874–889. [Google Scholar] [CrossRef]
- Oláh, J.; Vincze, O.; Virók, D.; Simon, D.; Bozsó, Z.; Tőkési, D.; Horváth, I.; Hlavanda, E.; Kovács, J.; Magyar, A.; et al. Interactions of Pathological Hallmark Proteins: Tubulin Polymerization Promoting Protein/P25, Β-Amyloid, And A-Synuclein*. J. Biol. Chem. 2011, 286, 34088–34100. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
van der Spek, S.J.F.; Pandya, N.J.; Koopmans, F.; Paliukhovich, I.; van der Schors, R.C.; Otten, M.; Smit, A.B.; Li, K.W. Expression and Interaction Proteomics of GluA1- and GluA3-Subunit-Containing AMPARs Reveal Distinct Protein Composition. Cells 2022, 11, 3648. https://doi.org/10.3390/cells11223648
van der Spek SJF, Pandya NJ, Koopmans F, Paliukhovich I, van der Schors RC, Otten M, Smit AB, Li KW. Expression and Interaction Proteomics of GluA1- and GluA3-Subunit-Containing AMPARs Reveal Distinct Protein Composition. Cells. 2022; 11(22):3648. https://doi.org/10.3390/cells11223648
Chicago/Turabian Stylevan der Spek, Sophie J. F., Nikhil J. Pandya, Frank Koopmans, Iryna Paliukhovich, Roel C. van der Schors, Mylene Otten, August B. Smit, and Ka Wan Li. 2022. "Expression and Interaction Proteomics of GluA1- and GluA3-Subunit-Containing AMPARs Reveal Distinct Protein Composition" Cells 11, no. 22: 3648. https://doi.org/10.3390/cells11223648
APA Stylevan der Spek, S. J. F., Pandya, N. J., Koopmans, F., Paliukhovich, I., van der Schors, R. C., Otten, M., Smit, A. B., & Li, K. W. (2022). Expression and Interaction Proteomics of GluA1- and GluA3-Subunit-Containing AMPARs Reveal Distinct Protein Composition. Cells, 11(22), 3648. https://doi.org/10.3390/cells11223648