
RNA N6-Methyladenosine (m6A) Methyltransferase-like 3 Facilitates Tumorigenesis and Cisplatin Resistance of Arecoline-Exposed Oral Carcinoma
 , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Human Sample Collection
2.2. Cell Culture, Cell Transfection, and Lentiviral Transduction
2.3. Immunohistochemistry (IHC) Analysis
2.4. Luciferase Reporter Assay
2.5. RT-qPCR
2.6. Chromatin Immunoprecipitation (ChIP)–qPCR Assay
2.7. Western Blotting
2.8. Cell Proliferation Assay and Cell Migration Assay
2.9. ELISA
2.10. Mouse Tumorigenesis and Treatment
2.11. Statistical Analyses
3. Results
3.1. MTTL3 Is Elevated in Arecoline-Exposed OSCC Cells and Tissues
3.2. HIF-1α Regulates METTL3 Expression
3.3. HIF-1α is Enhanced in Arecoline-Exposed OSCC Cells and Tissues
3.4. METTL3-MYC- HIF-1α Forms a Positive Autoregulation Loop
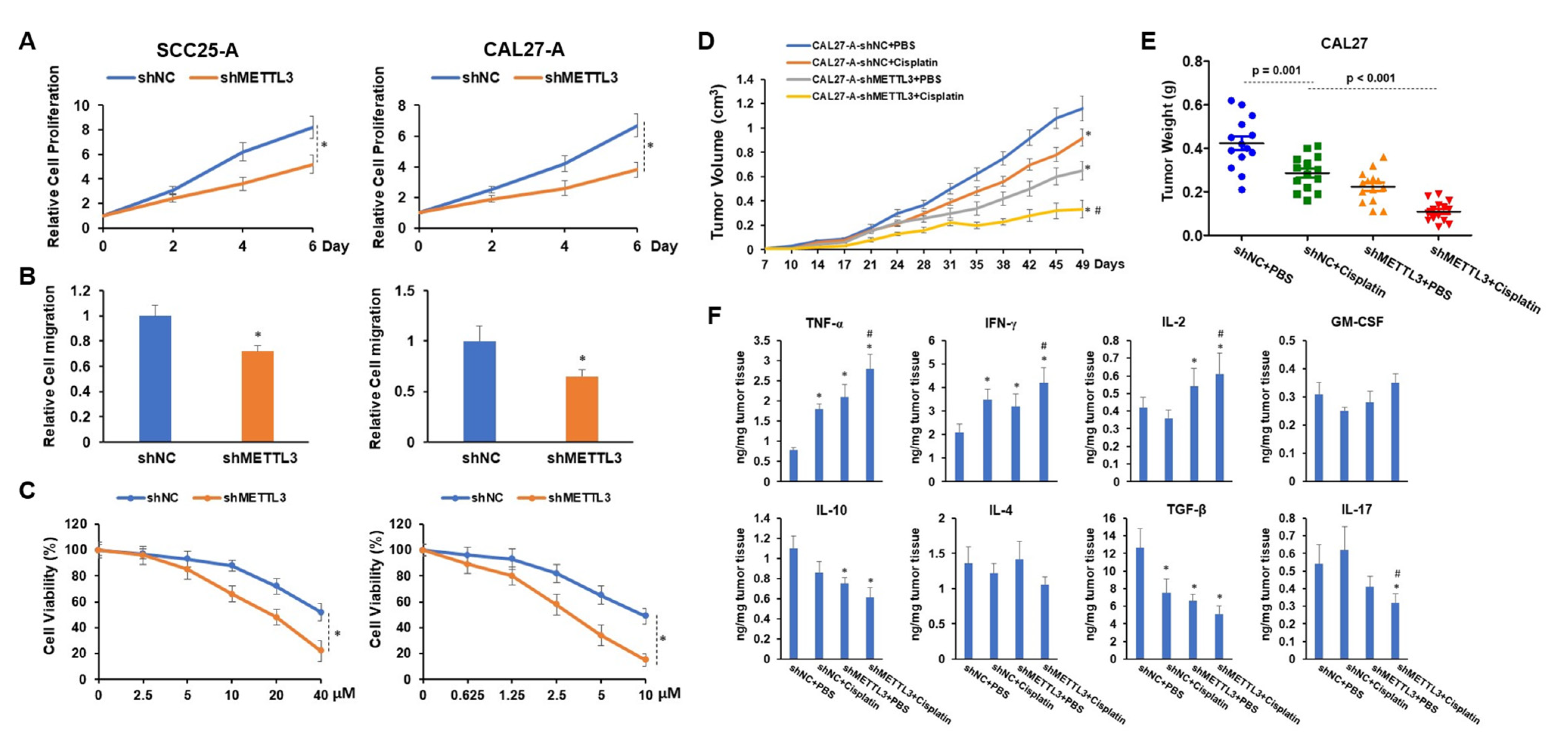
3.5. Knocking down METTL3 Sensitized Arecoline-Transformed OSCC Cells to Cisplatin-Induced Cytotoxicity
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Siegel, R.L.; Miller, K.D.; Fuchs, H.E.; Jemal, A. Cancer statistics, 2022. CA A Cancer J. Clin. 2022, 72, 7–33. [Google Scholar] [CrossRef] [PubMed]
- Ferlay, J.; Colombet, M.; Soerjomataram, I.; Parkin, D.M.; Pineros, M.; Znaor, A.; Bray, F. Cancer statistics for the year 2020: An overview. Int. J. Cancer 2021, 149, 778–789. [Google Scholar] [CrossRef] [PubMed]
- Johnson, D.E.; Burtness, B.; Leemans, C.R.; Lui, V.W.Y.; Bauman, J.E.; Grandis, J.R. Head and neck squamous cell carcinoma. Nat. Rev. Dis. Prim. 2020, 6, 92. [Google Scholar] [CrossRef] [PubMed]
- Walsh, T.; Macey, R.; Kerr, A.R.; Lingen, M.W.; Ogden, G.R.; Warnakulasuriya, S. Diagnostic tests for oral cancer and potentially malignant disorders in patients presenting with clinically evident lesions. Cochrane Database Syst. Rev. 2021, 7, CD010276. [Google Scholar]
- GBD 2019 Cancer Risk Factors Collaborators. The global burden of cancer attributable to risk factors, 2010–2019: A systematic analysis for the Global Burden of Disease Study 2019. Lancet 2022, 400, 563–591. [Google Scholar] [CrossRef]
- Nair, U.; Bartsch, H.; Nair, J. Alert for an epidemic of oral cancer due to use of the betel quid substitutes gutkha and pan masala: A review of agents and causative mechanisms. Mutagenesis 2004, 19, 251–262. [Google Scholar] [CrossRef]
- Su, S.Y.; Chen, W.T.; Chiang, C.J.; Yang, Y.W.; Lee, W.C. Oral cancer incidence rates from 1997 to 2016 among men in Taiwan: Association between birth cohort trends and betel nut consumption. Oral Oncol. 2020, 107, 104798. [Google Scholar] [CrossRef]
- Chen, P.H.; Mahmood, Q.; Mariottini, G.L.; Chiang, T.A.; Lee, K.W. Adverse Health Effects of Betel Quid and the Risk of Oral and Pharyngeal Cancers. BioMed Res. Int. 2017, 2017, 3904098. [Google Scholar] [CrossRef]
- Hernandez, B.Y.; Zhu, X.; Goodman, M.T.; Gatewood, R.; Mendiola, P.; Quinata, K.; Paulino, Y.C. Betel nut chewing, oral premalignant lesions, and the oral microbiome. PLoS ONE 2017, 12, e0172196. [Google Scholar] [CrossRef]
- Zhao, S.; Allis, C.D.; Wang, G.G. The language of chromatin modification in human cancers. Nat. Rev. Cancer 2021, 21, 413–430. [Google Scholar] [CrossRef]
- Zhou, Z.; Lv, J.; Yu, H.; Han, J.; Yang, X.; Feng, D.; Wu, Q.; Yuan, B.; Lu, Q.; Yang, H. Mechanism of RNA modification N6-methyladenosine in human cancer. Mol. Cancer 2020, 19, 104. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Xie, X.; Gu, Y.; Zhang, J.; Song, J.; Cheng, X.; Gao, Y.; Ai, Y. Fat mass and obesity-associated protein regulates tumorigenesis of arecoline-promoted human oral carcinoma. Cancer Med. 2021, 10, 6402–6415. [Google Scholar] [CrossRef] [PubMed]
- Tan, F.; Zhao, M.; Xiong, F.; Wang, Y.; Zhang, S.; Gong, Z.; Li, X.; He, Y.; Shi, L.; Wang, F.; et al. N6-methyladenosine-dependent signalling in cancer progression and insights into cancer therapies. J. Exp. Clin. Cancer Res. CR 2021, 40, 146. [Google Scholar] [CrossRef] [PubMed]
- Ai, Y.; Liu, S.; Luo, H.; Wu, S.; Wei, H.; Tang, Z.; Li, X.; Lv, X.; Zou, C. METTL3 Intensifies the Progress of Oral Squamous Cell Carcinoma via Modulating the m6A Amount of PRMT5 and PD-L1. J. Immunol. Res. 2021, 2021, 6149558. [Google Scholar] [CrossRef]
- Li, X.; Chen, W.; Gao, Y.; Song, J.; Gu, Y.; Zhang, J.; Cheng, X.; Ai, Y. Fat mass and obesity-associated protein regulates arecoline-exposed oral cancer immune response through programmed cell death-ligand 1. Cancer Sci. 2022, 113, 2962–2973. [Google Scholar] [CrossRef]
- Chuerduangphui, J.; Ekalaksananan, T.; Chaiyarit, P.; Patarapadungkit, N.; Chotiyano, A.; Kongyingyoes, B.; Promthet, S.; Pientong, C. Effects of arecoline on proliferation of oral squamous cell carcinoma cells by dysregulating c-Myc and miR-22, directly targeting oncostatin M. PLoS ONE 2018, 13, e0192009. [Google Scholar] [CrossRef]
- Doe, M.R.; Ascano, J.M.; Kaur, M.; Cole, M.D. Myc posttranscriptionally induces HIF1 protein and target gene expression in normal and cancer cells. Cancer Res. 2012, 72, 949–957. [Google Scholar] [CrossRef]
- Weili, Z.; Zhikun, L.; Jianmin, W.; Qingbao, T. Knockdown of USP28 enhances the radiosensitivity of esophageal cancer cells via the c-Myc/hypoxia-inducible factor-1 alpha pathway. J. Cell. Biochem. 2019, 120, 201–212. [Google Scholar] [CrossRef]
- Xiang, S.; Liang, X.; Yin, S.; Liu, J.; Xiang, Z. N6-methyladenosine methyltransferase METTL3 promotes colorectal cancer cell proliferation through enhancing MYC expression. Am. J. Transl. Res. 2020, 12, 1789–1806. [Google Scholar]
- Yuan, Y.; Du, Y.; Wang, L.; Liu, X. The M6A methyltransferase METTL3 promotes the development and progression of prostate carcinoma via mediating MYC methylation. J. Cancer 2020, 11, 3588–3595. [Google Scholar] [CrossRef]
- Zhang, J.; Bai, R.; Li, M.; Ye, H.; Wu, C.; Wang, C.; Li, S.; Tan, L.; Mai, D.; Li, G.; et al. Excessive miR-25-3p maturation via N(6)-methyladenosine stimulated by cigarette smoke promotes pancreatic cancer progression. Nat. Commun. 2019, 10, 1858. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Chen, C.; Ding, Q.; Zhao, Y.; Wang, Z.; Chen, J.; Jiang, Z.; Zhang, Y.; Xu, G.; Zhang, J.; et al. METTL3-mediated m(6)A modification of HDGF mRNA promotes gastric cancer progression and has prognostic significance. Gut 2020, 69, 1193–1205. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Song, S.; Meng, Q.; Wang, L.; Li, X.; Xie, S.; Chen, Y.; Jiang, X.; Wang, C.; Lu, Y.; et al. miR24-2 accelerates progression of liver cancer cells by activating Pim1 through tri-methylation of Histone H3 on the ninth lysine. J. Cell. Mol. Med. 2020, 24, 2772–2790. [Google Scholar] [CrossRef] [PubMed]
- Chamorro-Jorganes, A.; Sweaad, W.K.; Katare, R.; Besnier, M.; Anwar, M.; Beazley-Long, N.; Sala-Newby, G.; Ruiz-Polo, I.; Chandrasekera, D.; Ritchie, A.A.; et al. METTL3 Regulates Angiogenesis by Modulating let-7e-5p and miRNA-18a-5p Expression in Endothelial Cells. Arterioscler. Thromb. Vasc. Biol. 2021, 41, e325–e337. [Google Scholar] [CrossRef]
- Zhang, M.; Bai, M.; Wang, L.; Lu, N.; Wang, J.; Yan, R.; Cui, M.; Yan, H.; Zhang, L. Targeting SNHG3/miR-186-5p reverses the increased m6A level caused by platinum treatment through regulating METTL3 in esophageal cancer. Cancer Cell Int. 2021, 21, 114. [Google Scholar] [CrossRef]
- Yang, Z.; Quan, Y.; Chen, Y.; Huang, Y.; Huang, R.; Yu, W.; Wu, D.; Ye, M.; Min, Z.; Yu, B. Knockdown of RNA N6-methyladenosine methyltransferase METTL3 represses Warburg effect in colorectal cancer via regulating HIF-1alpha. Signal Transduct. Target. Ther. 2021, 6, 89. [Google Scholar] [CrossRef]
- Semenza, G.L. Defining the role of hypoxia-inducible factor 1 in cancer biology and therapeutics. Oncogene 2010, 29, 625–634. [Google Scholar] [CrossRef] [PubMed]
- Eckert, A.W.; Kappler, M.; Grosse, I.; Wickenhauser, C.; Seliger, B. Current Understanding of the HIF-1-Dependent Metabolism in Oral Squamous Cell Carcinoma. Int. J. Mol. Sci. 2020, 21, 6083. [Google Scholar] [CrossRef]
- Rashid, M.; Zadeh, L.R.; Baradaran, B.; Molavi, O.; Ghesmati, Z.; Sabzichi, M.; Ramezani, F. Up-down regulation of HIF-1alpha in cancer progression. Gene 2021, 798, 145796. [Google Scholar] [CrossRef]
- Li, Y.C.; Cheng, A.J.; Lee, L.Y.; Huang, Y.C.; Chang, J.T. Multifaceted Mechanisms of Areca Nuts in Oral Carcinogenesis: The Molecular Pathology from Precancerous Condition to Malignant Transformation. J. Cancer 2019, 10, 4054–4062. [Google Scholar] [CrossRef]
- Makovec, T. Cisplatin and beyond: Molecular mechanisms of action and drug resistance development in cancer chemotherapy. Radiol. Oncol. 2019, 53, 148–158. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.; Li, S.; Gao, L.; Zhi, K.; Ren, W. The Molecular Basis and Therapeutic Aspects of Cisplatin Resistance in Oral Squamous Cell Carcinoma. Front. Oncol. 2021, 11, 761379. [Google Scholar] [CrossRef] [PubMed]
- Khoo, X.H.; Paterson, I.C.; Goh, B.H.; Lee, W.L. Cisplatin-Resistance in Oral Squamous Cell Carcinoma: Regulation by Tumor Cell-Derived Extracellular Vesicles. Cancers 2019, 11, 1166. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Huang, C.M.; Shao, Z.; Zhao, X.P.; Wang, M.; Yan, T.L.; Zhou, X.C.; Jiang, E.H.; Liu, K.; Shang, Z.J. Autophagy Induced by Areca Nut Extract Contributes to Decreasing Cisplatin Toxicity in Oral Squamous Cell Carcinoma Cells: Roles of Reactive Oxygen Species/AMPK Signaling. Int. J. Mol. Sci. 2017, 18, 524. [Google Scholar] [CrossRef] [PubMed]
- Wen, Q.T.; Wang, T.; Yu, D.H.; Wang, Z.R.; Sun, Y.; Liang, C.W. Development of a mouse model of arecoline-induced oral mucosal fibrosis. Asian Pac. J. Trop. Med. 2017, 10, 1177–1184. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Relative METTL3 mRNA Levels | p-Value | |
|---|---|---|
| Gender | ||
| Female | 1.83 | 0.061 |
| Male | 2.06 | |
| Age | ||
| <65 | 2.01 | 0.83 |
| ≥65 | 2.05 | |
| Areca nut chewing habit | ||
| Yes | 1.75 | 0.041 |
| No | 2.18 | |
| Tumor Stage | ||
| 1 and 2 | 1.89 | 0.055 |
| 3 and 4 | 2.01 | |
| Nodal Stage | ||
| 0 | 1.83 | 0.073 |
| 1 | 1.95 | |
| 2 | 2.02 | |
| Recurrence | ||
| No | 1.24 | 0.034 |
| Local | 1.66 | |
| Distant | 2.03 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, C.; Kadigamuwa, C.; Wu, S.; Gao, Y.; Chen, W.; Gu, Y.; Wang, S.; Li, X. RNA N6-Methyladenosine (m6A) Methyltransferase-like 3 Facilitates Tumorigenesis and Cisplatin Resistance of Arecoline-Exposed Oral Carcinoma. Cells 2022, 11, 3605. https://doi.org/10.3390/cells11223605
Wang C, Kadigamuwa C, Wu S, Gao Y, Chen W, Gu Y, Wang S, Li X. RNA N6-Methyladenosine (m6A) Methyltransferase-like 3 Facilitates Tumorigenesis and Cisplatin Resistance of Arecoline-Exposed Oral Carcinoma. Cells. 2022; 11(22):3605. https://doi.org/10.3390/cells11223605
Chicago/Turabian StyleWang, Chuang, Chamila Kadigamuwa, Songlv Wu, Yijun Gao, Wuya Chen, Yangcong Gu, Shengli Wang, and Xia Li. 2022. "RNA N6-Methyladenosine (m6A) Methyltransferase-like 3 Facilitates Tumorigenesis and Cisplatin Resistance of Arecoline-Exposed Oral Carcinoma" Cells 11, no. 22: 3605. https://doi.org/10.3390/cells11223605
APA StyleWang, C., Kadigamuwa, C., Wu, S., Gao, Y., Chen, W., Gu, Y., Wang, S., & Li, X. (2022). RNA N6-Methyladenosine (m6A) Methyltransferase-like 3 Facilitates Tumorigenesis and Cisplatin Resistance of Arecoline-Exposed Oral Carcinoma. Cells, 11(22), 3605. https://doi.org/10.3390/cells11223605