
Effect of Phytochrome Deficiency on Photosynthesis, Light-Related Genes Expression and Flavonoid Accumulation in Solanum lycopersicum under Red and Blue Light

 ,
,  ,
,  ,
,  ,
, 

Abstract
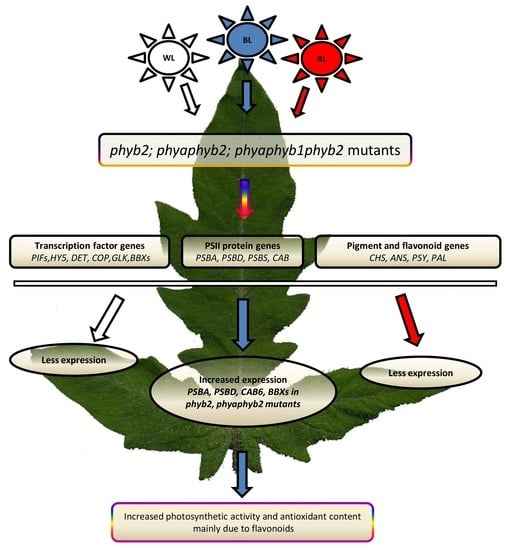
1. Introduction
2. Materials and Methods
2.1. Plant Materials and Experimental Design
2.2. Analysis of Low Molecular Weight Antioxidants
2.3. Measurements of CO2 Gas Exchange and Transpiration
2.4. Determination of Photochemical Activity
2.5. RNA Extraction and RT-PCR
2.6. Scanning Electron Microscopy
2.7. Statistics
3. Results
3.1. Low-Molecular Weight Antioxidant Capacity
3.2. Total Phenolics Content
3.3. Flavonoid Content
3.4. Measurements of CO2 Gas Exchange and Transpiration
3.5. Determination of Photochemical Activity
3.6. Gene Expression
3.7. Scanning Electron Microscopy
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Darko, E.; Heydarizadeh, P.; Schoefs, B.; Sabzalian, M.R. Photosynthesis under artificial light: The shift in primary and secondary metabolism. Philos. Trans. R. Soc. B 2014, 369, 20130243. [Google Scholar] [CrossRef] [PubMed]
- Bae, G.; Choi, G. Decoding of light signals by plant phytochromes and their interacting proteins. Annu. Rev. Plant Biol. 2008, 59, 281–311. [Google Scholar] [CrossRef] [PubMed]
- Lupi, A.C.D.; Lira, B.S.; Gramegna, G.; Trench, B.; Alves, F.R.R.; Demarco, D.; Peres, L.E.P.; Purgatto, E.; Freschi, L.; Rossi, M. Solanum lycopersicum GOLDEN 2-LIKE 2 transcription factor affects fruit quality in a light-and auxin-dependent manner. PLoS ONE 2019, 14, e0212224. [Google Scholar] [CrossRef]
- Song, Y.; Yang, C.; Gao, S.; Zhang, W.; Li, L.; Kuai, B. Age-triggered and dark-induced leaf senescence require the bHLH transcription factors PIF3, 4, and 5. Mol. Plant. 2014, 7, 1776–1787. [Google Scholar] [CrossRef]
- Waters, M.T.; Wang, P.; Korkaric, M.; Capper, R.G.; Saunders, N.J.; Langdale, J.A. GLK transcription factors coordinate expression of the photosynthetic apparatus in Arabidopsis. Plant Cell 2009, 21, 1109–1128. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; He, K.; Stolc, V.; Lee, H.; Figueroa, P.; Gao, Y.; Tongprasit, W.; Zhao, H.; Lee, I.; Deng, X.W. Analysis of transcription factor HY5 genomic binding sites revealed its hierarchical role in light regulation of development. Plant Cell 2007, 19, 731–749. [Google Scholar] [CrossRef]
- Kreslavski, V.D.; Los, D.A.; Schmitt, F.J.; Zharmukhamedov, S.K.; Kuznetsov, V.V.; Allakhverdiev, S.I. The impact of the phytochromes on photosynthetic processes. BBA-Bioenerg. 2018, 1859, 400–408. [Google Scholar] [CrossRef]
- Gavassi, M.A.; Monteiro, C.C.; Campos, M.L.; Melo, H.C.; Carvalho, R.F. Phytochromes are key regulators of abiotic stress responses in tomato. Sci. Hortic. 2017, 222, 126–135. [Google Scholar] [CrossRef]
- Kiełbowicz-Matuk, A. Involvement of plant C2H2-type zinc finger transcription factors in stress responses. Plant Sci. 2012, 185, 78–85. [Google Scholar] [CrossRef]
- Lira, B.S.; Oliveira, M.J.; Shiose, L.; Wu, R.T.A.; Rosado, D.; Lupi, A.C.D.; Freschi, L.; Rossi, M. Light and ripening-regulated BBX protein-encoding genes in Solanum lycopersicum. Sci. Rep. 2020, 10, 19235. [Google Scholar] [CrossRef]
- Song, Z.; Bian, Y.; Liu, J.; Sun, Y.; Xu, D. B-box proteins: Pivotal players in light-mediated development in plants. J. Integr. Plant Biol. 2020, 62, 1293–1309. [Google Scholar] [CrossRef] [PubMed]
- Xu, D.; Jiang, Y.; Li, J.; Lin, F.; Holm, M.; Deng, X.W. BBX21, an Arabidopsis B-box protein, directly activates HY5 and is targeted by COP1 for 26S proteasome-mediated degradation. Proc. Natl. Acad. Sci. USA 2016, 113, 7655–7660. [Google Scholar] [CrossRef] [PubMed]
- Gangappa, S.N.; Crocco, C.D.; Johansson, H.; Datta, S.; Hettiarachchi, C.; Holm, M.; Botto, J.F. The Arabidopsis B-box protein BBX25 interacts with HY5, negatively regulating BBX22 expression to suppress seedling photomorphogenesis. Plant Cell 2013, 25, 1243–1257. [Google Scholar] [CrossRef] [PubMed]
- Lin, F.; Jiang, Y.; Li, J.; Yan, T.; Fan, L.; Liang, J.; Chen, Z.J.; Xu, D.; Deng, X.W. B-BOX DOMAIN PROTEIN28 negatively regulates photomorphogenesis by repressing the activity of transcription factor HY5 and undergoes COP1-mediated degradation. Plant Cell 2018, 30, 2006–2019. [Google Scholar] [CrossRef]
- Zhang, X.; Lin, R. Light signaling differentially regulates the expression of group IV of the B-box zinc finger family. Plant Signal. Behav. 2017, 12, e1365213. [Google Scholar] [CrossRef]
- Kreslavski, V.D.; Strokina, V.V.; Pashkovskiy, P.P.; Balakhnina, T.I.; Voloshin, R.A.; Alwasel, S.; Kosobryukhov, A.A.; Allakhverdiev, S.I. Deficiencies in phytochromes A and B and cryptochrome 1 affect the resistance of the photosynthetic apparatus to high-intensity light in Solanum lycopersicum. J. Photochem. Photobiol. B 2020, 210, 111976. [Google Scholar] [CrossRef]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Singleton, V.L.; Rossi, J.A. Colorimetry of total phenolics with phosphomolybdic-phosphotungstic acid reagents. Am. J. Enol. Viticult. 1965, 16, 144–158. [Google Scholar]
- Kim, D.O.; Jeong, S.W.; Lee, C.Y. Antioxidant capacity of phenolic phytochemicals from various cultivars of plums. Food Chem. 2003, 81, 321–326. [Google Scholar] [CrossRef]
- Pashkovskiy, P.P.; Vankova, R.; Zlobin, I.E.; Dobrev, P.; Kartashov, A.V.; Ivanova, A.I.; Ivanov, V.P.; Marchenko, S.I.; Nartov, D.I.; Ivanov, Y.V.; et al. Hormonal responses to short-term and long-term water deficit in native Scots pine and Norway spruce trees. Environ. Exp. Bot. 2022, 195, 104789. [Google Scholar] [CrossRef]
- Stetsenko, L.A.; Pashkovsky, P.P.; Voloshin, R.A.; Kreslavski, V.D.; Kuznetsov, V.V.; Allakhverdiev, S.I. Role of anthocyanin and carotenoids in the adaptation of the photosynthetic apparatus of purple-and green-leaved cultivars of sweet basil (Ocimum basilicum) to high-intensity light. Photosynthetica 2020, 58, 890–901. [Google Scholar] [CrossRef]
- Schreiber, U. Pulse-amplitude-modulation (PAM) fluorometry and saturation pulse method: An overview. In Chlorophyll a Fluorescence: A Signature of Photosynthesis, 1st ed.; Papageorgiou, G.C., Govindjee, Eds.; Springer: Dordrecht, The Netherlands, 2004; pp. 279–319. [Google Scholar] [CrossRef]
- Rosado, D.; Gramegna, G.; Cruz, A.; Lira, B.S.; Freschi, L.; de Setta, N.; Rossi, M. Phytochrome Interacting Factors (PIFs) in Solanum lycopersicum: Diversity, evolutionary history and expression profiling during different developmental processes. PLoS ONE 2016, 11, e0165929. [Google Scholar] [CrossRef]
- Xu, D. COP1 and BBXs-HY5-mediated light signal transduction in plants. New Phytol. 2020, 228, 1748–1753. [Google Scholar] [CrossRef]
- Liu, C.C.; Chi, C.; Jin, L.J.; Zhu, J.; Yu, J.Q.; Zhou, Y.H. The bZip transcription factor HY5 mediates CRY1a-induced anthocyanin biosynthesis in tomato. Plant Cell Environ. 2018, 41, 1762–1775. [Google Scholar] [CrossRef]
- Pashkovskiy, P.; Kreslavski, V.; Khudyakova, A.; Ashikhmin, A.; Bolshakov, M.; Kozhevnikova, A.; Kosobryukhov, A.; Kuznetsov, V.V.; Allakhverdiev, S.I. Effect of high-intensity light on the photosynthetic activity, pigment content and expression of light-dependent genes of photomorphogenetic Solanum lycopersicum hp mutants. Plant Physiol. Biochem. 2021, 167, 91–100. [Google Scholar] [CrossRef]
- Pashkovskiy, P.; Ryazansky, S.; Kartashov, A.; Voloshin, R.; Khudyakova, A.; Kosobryukhov, A.A.; Kreslavski, V.D.; Kuznetsov, V.V.; Allakhverdiev, S.I. Effect of red light on photosynthetic acclimation and the gene expression of certain light signaling components involved in the microRNA biogenesis in the extremophile Eutrema salsugineum. J. Biotechnol. 2021, 325, 35–42. [Google Scholar] [CrossRef]
- Zhao, X.; Heng, Y.; Wang, X.; Deng, X.W.; Xu, D. A positive feedback loop of BBX11-BBX21-HY5 promotes photomorphogenic development in Arabidopsis. Plant Commun. 2020, 1, 100045. [Google Scholar] [CrossRef]
- Bu, X.; Wang, X.; Yan, J.; Zhang, Y.; Zhou, S.; Sun, X.; Yang, Y.; Ahammed, G.J.; Liu, Y.; Qi, M.; et al. Genome-wide characterization of B-box gene family and its roles in responses to light quality and cold stress in tomato. Front. Plant Sci. 2021, 12, 698525. [Google Scholar] [CrossRef]
- Heng, Y.; Lin, F.; Jiang, Y.; Ding, M.; Yan, T.; Lan, H.; Zhou, H.; Zhao, X.; Xu, D.; Deng, X.W. B-box containing proteins BBX30 and BBX31, acting downstream of HY5, negatively regulate photomorphogenesis in Arabidopsis. Plant Physiol. 2019, 180, 497–508. [Google Scholar] [CrossRef]
- McDowell, E.T.; Kapteyn, J.; Schmidt, A.; Li, C.; Kang, J.H.; Descour, A.; Shi, F.; Larson, M.; Schilmiller, A.; An, L.; et al. Comparative functional genomic analysis of Solanum glandular trichome types. Plant Physiol. 2010, 155, 524–539. [Google Scholar] [CrossRef]
- Alallaq, S.; Ranjan, A.; Brunoni, F.; Novák, O.; Lakehal, A.; Bellini, C. Red light controls adventitious root regeneration by modulating hormone homeostasis in Picea abies seedlings. Front. Plant Sci. 2020, 11, 586140. [Google Scholar] [CrossRef]
- Bantis, F.; Panteris, E.; Dangitsis, C.; Carrera, E.; Koukounaras, A. Blue light promotes vascular reconnection, while red light boosts the physiological response and quality of grafted watermelon seedlings. Sci. Rep. 2021, 11, 21754. [Google Scholar] [CrossRef]
- Pashkovskiy, P.; Kreslavski, V.D.; Ivanov, Y.; Ivanova, A.; Kartashov, A.; Shmarev, A.; Strokina, V.; Kuznetsov, V.V.; Allakhverdiev, S.I. Influence of Light of Different Spectral Compositions on the Growth, Photosynthesis, and Expression of Light-Dependent Genes of Scots Pine Seedlings. Cells 2021, 10, 3284. [Google Scholar] [CrossRef]
- Nagaoka, S.; Takano, T. Salt tolerance-related protein STO binds to a Myb transcription factor homologue and confers salt tolerance in Arabidopsis. J. Exp. Bot. 2003, 54, 2231–2237. [Google Scholar] [CrossRef]
- Kreslavski, V.D.; Khudyakova, A.Y.; Strokina, V.V.; Shirshikova, G.N.; Pashkovskiy, P.P.; Balakhnina, T.I.; Kosobryukhov, A.A.; Kuznetsov, V.V.; Allakhverdiev, S.I. Impact of high irradiance and UV-B on the photosynthetic activity, pro-/antioxidant balance and expression of light-activated genes in Arabidopsis thaliana hy4 mutants grown under blue light. Plant Physiol. Biochem. 2021, 167, 153–162. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variant | Mutants | Pn, µmol CO2 m−2 s−1 | Tr, mmol H2O m−2 s−1 | TEAC, µmol Trolox/g FW | GAE, mg/g FW | Flavonoids, mg catechin/g FW |
|---|---|---|---|---|---|---|
| WFL | wt | 15.8 ± 1.0 a | 0.32 ± 0.02 b | 9.17 ± 0.12 c | 0.83 ± 0.02 c | 0.98 ± 0.02 c |
| phyb2 | 10.6 ± 0.6 b | 0.12 ± 0.02 d | 18.43 ±1.28 a | 1.76 ± 0.08 a | 1.63 ± 0.10 a | |
| phyaphyb2 | 8.8 ± 0.4 c | 0.45 ± 0.02 a | 14.97 ± 1.16 b | 1.18 ± 0.10 b | 1.32 ± 0.05 b | |
| phyaphyb1phyb2 | 6.5 ± 0.3 d | 0.22 ± 0.03 c | 11.08 ± 0.55 c | 0.98 ± 0.06 bc | 1.14 ± 0.05 bc | |
| BL | wt | 11.7 ± 1.0 a | 0.48 ± 0.02 c | 19.00 ± 0.75 c | 1.95 ± 0.20 c | 1.99 ± 0.09 c |
| phyb2 | 11.2 ± 0.4 a | 1.19 ± 0.03 a | 50.50 ± 4.24 a | 4.95 ± 0.20 a | 5.30 ± 0.26 a | |
| phyaphyb2 | 7.5 ± 0.3 b | 0.73 ± 0.04 b | 37.37 ± 3.22 b | 4.13 ± 0.25 b | 4.25 ± 0.30 b | |
| phyaphyb1phyb2 | 3.1 ± 0.2 c | 0.38 ± 0.01 d | 15.77 ± 1.74 c | 1.27 ± 0.03 d | 1.82 ± 0.18 c | |
| RL | wt | 14 ± 0.7 a | 0.19 ± 0.02 c | 9.80 ± 0.46 b | 0.98 ± 0.03 ab | 0.98 ± 0.08 b |
| phyb2 | 9.8 ± 0.6 b | 0.48 ± 0.02 a | 7.73 ± 1.28 b | 0.69 ± 0.16 b | 0.79 ± 0.12 b | |
| phyaphyb2 | 6.7 ± 0.2 c | 0.38 ± 0.02 b | 13.43 ± 1.39 a | 1.30 ± 0.14 a | 1.49 ± 0.23 a | |
| phyaphyb1phyb2 | 3.8 ± 0.2 d | 0.16 ± 0.02 c | 10.3 ± 0.35 ab | 0.63 ± 0.01 b | 0.81 ± 0.04 b |
| Variant | Mutants | Y(II) | NPQ | Y(NO) | Y(NPQ) | Fv/Fm |
|---|---|---|---|---|---|---|
| WFL | wt | 0.411 ± 0.016 b | 0.69 ± 0.013 b | 0.34 ± 0.006 a | 0.24 ± 0.009 b | 0.84 ± 0.002 a |
| phyb2 | 0.401 ± 0.023 b | 1.02 ± 0.111 a | 0.29 ± 0.022 b | 0.30 ± 0.018 a | 0.82 ± 0.009 b | |
| phyaphyb2 | 0.502 ± 0.013 a | 0.89 ± 0.025 ab | 0.25 ± 0.004 b | 0.22 ± 0.009 c | 0.84 ± 0.002 a | |
| phyaphyb1phyb2 | 0.396 ± 0.029 b | 0.83 ± 0.041 ab | 0.32 ± 0.008 a | 0.27 ± 0.021 b | 0.83 ± 0.002 b | |
| BL | wt | 0.473 ± 0.008 b | 1.10 ± 0.071 ab | 0.25 ± 0.009 b | 0.27 ± 0.018 b | 0.85 ± 0.011 a |
| phyb2 | 0.547 ± 0.014 a | 0.63 ± 0.058 c | 0.27 ± 0.015 b | 0.17 ± 0.011 c | 0.84 ± 0.004 a | |
| phyaphyb2 | 0.407 ± 0.009 c | 1.16 ± 0.062 a | 0.27 ± 0.011 b | 0.31 ± 0.009 a | 0.84 ± 0.004 a | |
| phyaphyb1phyb2 | 0.343 ± 0.007 d | 0.92 ± 0.024 b | 0.34 ± 0.011 a | 0.31 ± 0.007 a | 0.85 ± 0.005 a | |
| RL | wt | 0.374 ± 0.011 ab | 0.71 ± 0.071 b | 0.36 ± 0.017 a | 0.25 ± 0.040 b | 0.83 ± 0.002 b |
| phyb2 | 0.403 ± 0.007 a | 0.66 ± 0.012 c | 0.35 ± 0.003 a | 0.23 ± 0.003 b | 0.82 ± 0.002 a | |
| phyaphyb2 | 0.413 ± 0.041 a | 0.73 ± 0.004 b | 0.33 ± 0.024 b | 0.24 ± 0.016 b | 0.83 ± 0.003 b | |
| phyaphyb1phyb2 | 0.314 ± 0.014 b | 0.93 ± 0.071 a | 0.35 ± 0.024 a | 0.33 ± 0.038 a | 0.79 ± 0.003 c |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pashkovskiy, P.; Vereshchagin, M.; Kreslavski, V.; Ivanov, Y.; Kumachova, T.; Ryabchenko, A.; Voronkov, A.; Kosobryukhov, A.; Kuznetsov, V.; Allakhverdiev, S.I. Effect of Phytochrome Deficiency on Photosynthesis, Light-Related Genes Expression and Flavonoid Accumulation in Solanum lycopersicum under Red and Blue Light. Cells 2022, 11, 3437. https://doi.org/10.3390/cells11213437
Pashkovskiy P, Vereshchagin M, Kreslavski V, Ivanov Y, Kumachova T, Ryabchenko A, Voronkov A, Kosobryukhov A, Kuznetsov V, Allakhverdiev SI. Effect of Phytochrome Deficiency on Photosynthesis, Light-Related Genes Expression and Flavonoid Accumulation in Solanum lycopersicum under Red and Blue Light. Cells. 2022; 11(21):3437. https://doi.org/10.3390/cells11213437
Chicago/Turabian StylePashkovskiy, Pavel, Mikhail Vereshchagin, Vladimir Kreslavski, Yury Ivanov, Tamara Kumachova, Andrey Ryabchenko, Alexander Voronkov, Anatoliy Kosobryukhov, Vladimir Kuznetsov, and Suleyman I. Allakhverdiev. 2022. "Effect of Phytochrome Deficiency on Photosynthesis, Light-Related Genes Expression and Flavonoid Accumulation in Solanum lycopersicum under Red and Blue Light" Cells 11, no. 21: 3437. https://doi.org/10.3390/cells11213437
APA StylePashkovskiy, P., Vereshchagin, M., Kreslavski, V., Ivanov, Y., Kumachova, T., Ryabchenko, A., Voronkov, A., Kosobryukhov, A., Kuznetsov, V., & Allakhverdiev, S. I. (2022). Effect of Phytochrome Deficiency on Photosynthesis, Light-Related Genes Expression and Flavonoid Accumulation in Solanum lycopersicum under Red and Blue Light. Cells, 11(21), 3437. https://doi.org/10.3390/cells11213437