

Multiplexed Digital Spatial Protein Profiling Reveals Distinct Phenotypes of Mononuclear Phagocytes in Livers with Advanced Fibrosis

 , , , and
, , , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Patients
2.2. RNA Extraction and Gene Expression Assay
2.3. Tissue Microarray (TMA) Construction
2.4. Digital Spatial Profiling (DSP) Assay
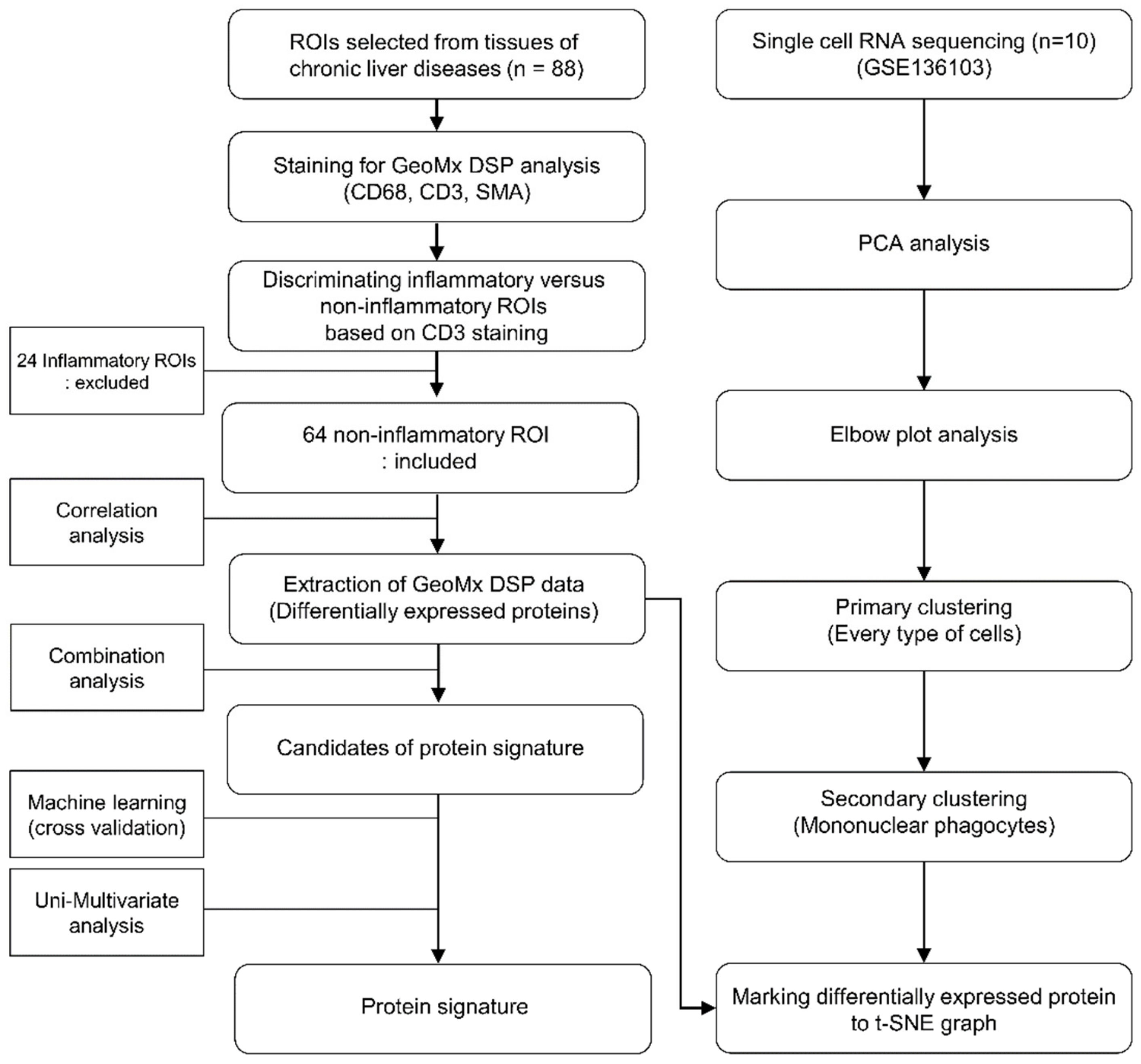
2.5. Selecting ROIs for Establishing the Protein Signatures
2.6. Analysis of DSP Data and Validation of the Protein Signatures
2.7. Single-Cell RNA Sequencing Analysis
2.8. Statistical Analysis
3. Results
3.1. Patient Characteristics
3.2. Multiplexed DSP of Protein Expression Level according to the Fibrosis Stage
3.3. Protein Signatures for the Advanced Fibrosis Derived from the DSP Analysis
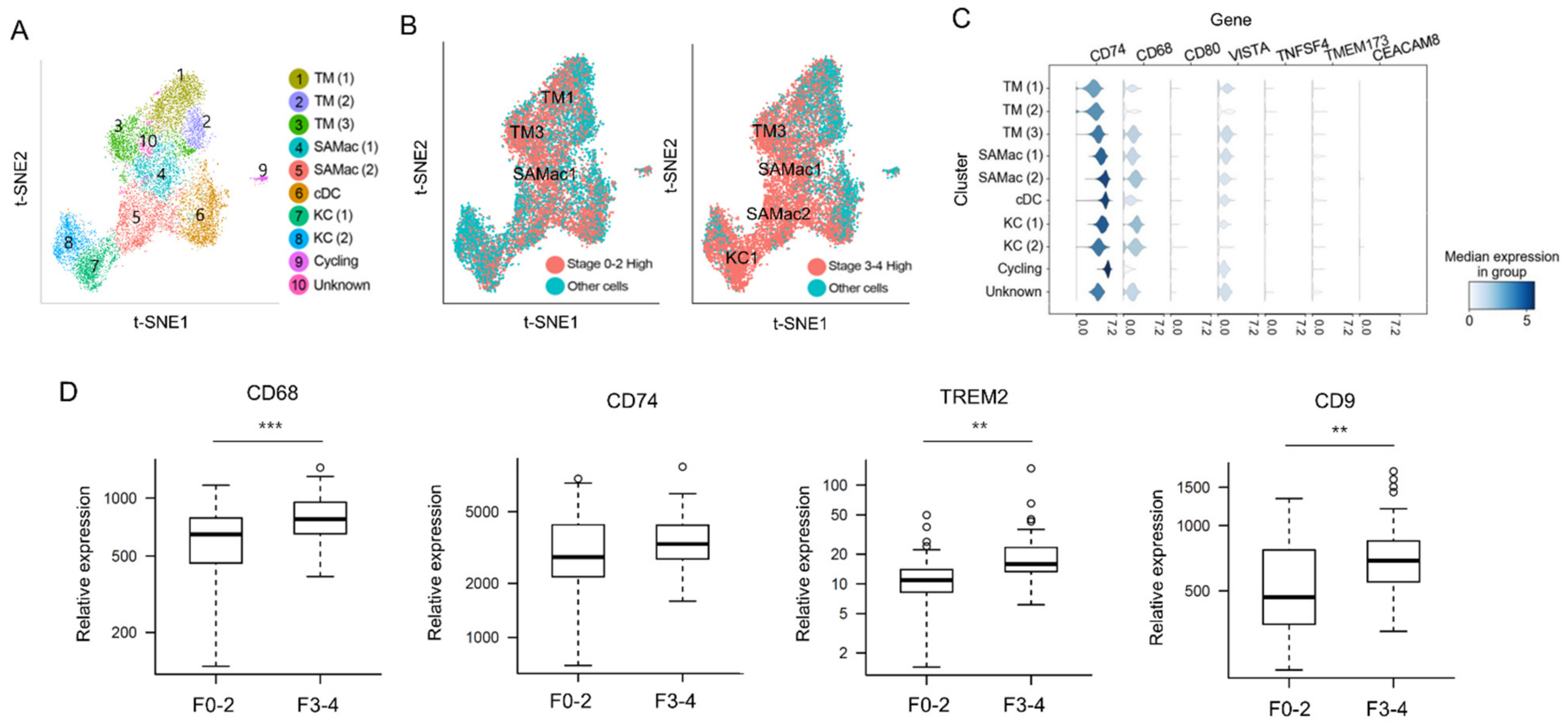
3.4. Predicting Related Immune Cells in Different Fibrosis Stage Using Single Cell RNA Sequencing Database
3.5. Validation of DSP Protein Analysis Using mRNA Expression Data from Snap-Frozen Livers Using NanoString nCounter MAX System
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Cheemerla, S.; Balakrishnan, M. Global Epidemiology of Chronic Liver Disease. Clin. Liver Dis. 2021, 17, 365–370. [Google Scholar] [CrossRef] [PubMed]
- KASL. Clinical practice guidelines for liver cirrhosis: Varices, hepatic encephalopathy, and related complications. Clin. Mol. Hepatol. 2020, 26, 83–127. [Google Scholar] [CrossRef] [PubMed]
- Odagiri, N.; Matsubara, T.; Sato-Matsubara, M.; Fujii, H.; Enomoto, M.; Kawada, N. Anti-fibrotic treatments for chronic liver diseases: The present and the future. Clin. Mol. Hepatol. 2021, 27, 413–424. [Google Scholar] [CrossRef] [PubMed]
- Sung, P.S.; Kim, C.M.; Cha, J.H.; Park, J.Y.; Yu, Y.S.; Wang, H.J.; Kim, J.K.; Bae, S.H. A Unique Immune-Related Gene Signature Represents Advanced Liver Fibrosis and Reveals Potential Therapeutic Targets. Biomedicines 2022, 10, 180. [Google Scholar] [CrossRef] [PubMed]
- Roehlen, N.; Crouchet, E.; Baumert, T.F. Liver Fibrosis: Mechanistic Concepts and Therapeutic Perspectives. Cells 2020, 9, 875. [Google Scholar] [CrossRef]
- Tacke, F.; Zimmermann, H.W. Macrophage heterogeneity in liver injury and fibrosis. J. Hepatol. 2014, 60, 1090–1096. [Google Scholar] [CrossRef]
- Gomez Perdiguero, E.; Klapproth, K.; Schulz, C.; Busch, K.; Azzoni, E.; Crozet, L.; Garner, H.; Trouillet, C.; de Bruijn, M.F.; Geissmann, F.; et al. Tissue-resident macrophages originate from yolk-sac-derived erythro-myeloid progenitors. Nature 2015, 518, 547–551. [Google Scholar] [CrossRef]
- Krenkel, O.; Tacke, F. Liver macrophages in tissue homeostasis and disease. Nat. Rev. Immunol. 2017, 17, 306–321. [Google Scholar] [CrossRef]
- Ju, C.; Tacke, F. Hepatic macrophages in homeostasis and liver diseases: From pathogenesis to novel therapeutic strategies. Cell. Mol. Immunol. 2016, 13, 316–327. [Google Scholar] [CrossRef]
- Sung, P.S. Crosstalk between tumor-associated macrophages and neighboring cells in hepatocellular carcinoma. Clin. Mol. Hepatol. 2021, 28, 333–350. [Google Scholar] [CrossRef]
- Fallowfield, J.A.; Mizuno, M.; Kendall, T.J.; Constandinou, C.M.; Benyon, R.C.; Duffield, J.S.; Iredale, J.P. Scar-Associated Macrophages Are a Major Source of Hepatic Matrix Metalloproteinase-13 and Facilitate the Resolution of Murine Hepatic Fibrosis. J. Immunol. 2007, 178, 5288–5295. [Google Scholar] [CrossRef]
- Ramachandran, P.; Dobie, R.; Wilson-Kanamori, J.R.; Dora, E.F.; Henderson, B.E.P.; Luu, N.T.; Portman, J.R.; Matchett, K.P.; Brice, M.; Marwick, J.A.; et al. Resolving the fibrotic niche of human liver cirrhosis at single-cell level. Nature 2019, 575, 512–518. [Google Scholar] [CrossRef]
- Aizarani, N.; Saviano, A.; Sagar; Mailly, L.; Durand, S.; Herman, J.S.; Pessaux, P.; Baumert, T.F.; Grün, D. A human liver cell atlas reveals heterogeneity and epithelial progenitors. Nature 2019, 572, 199–204. [Google Scholar] [CrossRef]
- MacParland, S.A.; Liu, J.C.; Ma, X.Z.; Innes, B.T.; Bartczak, A.M.; Gage, B.K.; Manuel, J.; Khuu, N.; Echeverri, J.; Linares, I.; et al. Single cell RNA sequencing of human liver reveals distinct intrahepatic macrophage populations. Nat. Commun. 2018, 9, 4383. [Google Scholar] [CrossRef]
- Krenkel, O.; Hundertmark, J.; Ritz, T.P.; Weiskirchen, R.; Tacke, F. Single Cell RNA Sequencing Identifies Subsets of Hepatic Stellate Cells and Myofibroblasts in Liver Fibrosis. Cells 2019, 8, 503. [Google Scholar] [CrossRef]
- Ramachandran, P.; Matchett, K.P.; Dobie, R.; Wilson-Kanamori, J.R.; Henderson, N.C. Single-cell technologies in hepatology: New insights into liver biology and disease pathogenesis. Nat. Rev. Gastroenterol. Hepatol. 2020, 17, 457–472. [Google Scholar] [CrossRef]
- Merritt, C.R.; Ong, G.T.; Church, S.E.; Barker, K.; Danaher, P.; Geiss, G.; Hoang, M.; Jung, J.; Liang, Y.; McKay-Fleisch, J.; et al. Multiplex digital spatial profiling of proteins and RNA in fixed tissue. Nat. Biotechnol. 2020, 38, 586–599. [Google Scholar] [CrossRef]
- Nerurkar, S.N.; Goh, D.; Cheung, C.C.L.; Nga, P.Q.Y.; Lim, J.C.T.; Yeong, J.P.S. Transcriptional Spatial Profiling of Cancer Tissues in the Era of Immunotherapy: The Potential and Promise. Cancers 2020, 12, 2572. [Google Scholar] [CrossRef]
- Vathiotis, I.A.; Yang, Z.; Reeves, J.; Toki, M.; Aung, T.N.; Wong, P.F.; Kluger, H.; Syrigos, K.N.; Warren, S.; Rimm, D.L. Models that combine transcriptomic with spatial protein information exceed the predictive value for either single modality. NPJ Precis. Oncol. 2021, 5, 45. [Google Scholar] [CrossRef]
- EASL. Clinical Practice Guidelines on non-invasive tests for evaluation of liver disease severity and prognosis—2021 update. J. Hepatol. 2021, 75, 659–689. [Google Scholar] [CrossRef]
- Dobie, R.; Wilson-Kanamori, J.R.; Henderson, B.E.P.; Smith, J.R.; Matchett, K.P.; Portman, J.R.; Wallenborg, K.; Picelli, S.; Zagorska, A.; Pendem, S.V.; et al. Single-Cell Transcriptomics Uncovers Zonation of Function in the Mesenchyme during Liver Fibrosis. Cell Rep. 2019, 29, 1832–1847. [Google Scholar] [CrossRef] [PubMed]
- Tamburini, B.A.J.; Finlon, J.M.; Gillen, A.E.; Kriss, M.S.; Riemondy, K.A.; Fu, R.; Schuyler, R.P.; Hesselberth, J.R.; Rosen, H.R.; Burchill, M.A. Chronic Liver Disease in Humans Causes Expansion and Differentiation of Liver Lymphatic Endothelial Cells. Front. Immunol. 2019, 10, 1036. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Bansal, M.B. Role of Kupffer Cells in Driving Hepatic Inflammation and Fibrosis in HIV Infection. Front. Immunol. 2020, 11, 1086. [Google Scholar] [CrossRef] [PubMed]
- Sung, P.S.; Park, D.J.; Roh, P.R.; Mun, K.D.; Cho, S.W.; Lee, G.W.; Jung, E.S.; Lee, S.H.; Jang, J.W.; Bae, S.H.; et al. Intrahepatic inflammatory IgA(+)PD-L1(high) monocytes in hepatocellular carcinoma development and immunotherapy. J. Immunother. Cancer 2022, 10, e003618. [Google Scholar] [CrossRef] [PubMed]
- Jophlin, L.L.; Cao, S.; Shah, V.H. The Transcriptome of Hepatic Fibrosis Revealed by Single-Cell RNA Sequencing. Hepatology 2020, 71, 1865–1867. [Google Scholar] [CrossRef]
- Beljaars, L.; Schippers, M.; Reker-Smit, C.; Martinez, F.O.; Helming, L.; Poelstra, K.; Melgert, B.N. Hepatic Localization of Macrophage Phenotypes during Fibrogenesis and Resolution of Fibrosis in Mice and Humans. Front. Immunol. 2014, 5, 430. [Google Scholar] [CrossRef]
- Chistiakov, D.A.; Killingsworth, M.C.; Myasoedova, V.A.; Orekhov, A.N.; Bobryshev, Y.V. CD68/macrosialin: Not just a histochemical marker. Lab. Investig. 2017, 97, 4–13. [Google Scholar] [CrossRef]
- Bernsmeier, C.; van der Merwe, S.; Périanin, A. Innate immune cells in cirrhosis. J. Hepatol. 2020, 73, 186–201. [Google Scholar] [CrossRef]
- Sun, G.; Jin, H.; Zhang, C.; Meng, H.; Zhao, X.; Wei, D.; Ou, X.; Wang, Q.; Li, S.; Wang, T.; et al. OX40 Regulates Both Innate and Adaptive Immunity and Promotes Nonalcoholic Steatohepatitis. Cell Rep. 2018, 25, 3786–3799. [Google Scholar] [CrossRef]
- Leicht, D.T.; Balan, V.; Kaplun, A.; Singh-Gupta, V.; Kaplun, L.; Dobson, M.; Tzivion, G. Raf kinases: Function, regulation and role in human cancer. Biochim. Biophys. Acta 2007, 1773, 1196–1212. [Google Scholar] [CrossRef]
- Takahashi, M.; Li, Y.; Dillon, T.J.; Kariya, Y.; Stork, P.J.S. Phosphorylation of the C-Raf N Region Promotes Raf Dimerization. Mol. Cell. Biol. 2017, 37, e00132-00117. [Google Scholar] [CrossRef]
- Qin, S.-Y.; Lu, D.-H.; Guo, X.-Y.; Luo, W.; Hu, B.-L.; Huang, X.-L.; Chen, M.; Wang, J.-X.; Ma, S.-J.; Yang, X.-W.; et al. A deleterious role for Th9/IL-9 in hepatic fibrogenesis. Sci. Rep. 2016, 6, 18694. [Google Scholar] [CrossRef]
- Guo, X.; Cen, Y.; Wang, J.; Jiang, H. CXCL10-induced IL-9 promotes liver fibrosis via Raf/MEK/ERK signaling pathway. Biomed. Pharmacother. 2018, 105, 282–289. [Google Scholar] [CrossRef]
- Pokatayev, V.; Yang, K.; Tu, X.; Dobbs, N.; Wu, J.; Kalb, R.G.; Yan, N. Homeostatic regulation of STING protein at the resting state by stabilizer TOLLIP. Nat. Immunol. 2020, 21, 158–167. [Google Scholar] [CrossRef]
- Burdette, D.L.; Vance, R.E. STING and the innate immune response to nucleic acids in the cytosol. Nat. Immunol. 2013, 14, 19–26. [Google Scholar] [CrossRef]
- Chen, C.; Yang, R.X.; Xu, H.G. STING and liver disease. J. Gastroenterol. 2021, 56, 704–712. [Google Scholar] [CrossRef]
- Luo, X.; Li, H.; Ma, L.; Zhou, J.; Guo, X.; Woo, S.L.; Pei, Y.; Knight, L.R.; Deveau, M.; Chen, Y.; et al. Expression of STING Is Increased in Liver Tissues From Patients With NAFLD and Promotes Macrophage-Mediated Hepatic Inflammation and Fibrosis in Mice. Gastroenterology 2018, 155, 1971–1984. [Google Scholar] [CrossRef]
- Iracheta-Vellve, A.; Petrasek, J.; Gyongyosi, B.; Satishchandran, A.; Lowe, P.; Kodys, K.; Catalano, D.; Calenda, C.D.; Kurt-Jones, E.A.; Fitzgerald, K.A.; et al. Endoplasmic Reticulum Stress-induced Hepatocellular Death Pathways Mediate Liver Injury and Fibrosis via Stimulator of Interferon Genes. J. Biol. Chem. 2016, 291, 26794–26805. [Google Scholar] [CrossRef]
- Li, Y.; He, M.; Wang, Z.; Duan, Z.; Guo, Z.; Wang, Z.; Gong, R.; Chu, T.; Cai, J.; Gao, B. STING signaling activation inhibits HBV replication and attenuates the severity of liver injury and HBV-induced fibrosis. Cell. Mol. Immunol. 2022, 19, 92–107. [Google Scholar] [CrossRef]
- Zhao, L.; Yu, G.; Han, Q.; Cui, C.; Zhang, B. TIM-3: An emerging target in the liver diseases. Scand. J. Immunol. 2020, 91, e12825. [Google Scholar] [CrossRef]
- Shi, Y.; Wu, W.; Yang, Y.; Yang, Q.; Song, G.; Wu, Y.; Wei, L.; Chen, Z. Decreased Tim-3 expression is associated with functional abnormalities of monocytes in decompensated cirrhosis without overt bacterial infection. J. Hepatol. 2015, 63, 60–67. [Google Scholar] [CrossRef] [PubMed]
- Han, G.; Chen, G.; Shen, B.; Li, Y. Tim-3: An activation marker and activation limiter of innate immune cells. Front. Immunol. 2013, 4, 449. [Google Scholar] [CrossRef] [PubMed]
- Sierro, F.; Evrard, M.; Rizzetto, S.; Melino, M.; Mitchell, A.J.; Florido, M.; Beattie, L.; Walters, S.B.; Tay, S.S.; Lu, B.; et al. A Liver Capsular Network of Monocyte-Derived Macrophages Restricts Hepatic Dissemination of Intraperitoneal Bacteria by Neutrophil Recruitment. Immunity 2017, 47, 374–388. [Google Scholar] [CrossRef] [PubMed]
- Remmerie, A.; Martens, L.; Thoné, T.; Castoldi, A.; Seurinck, R.; Pavie, B.; Roels, J.; Vanneste, B.; De Prijck, S.; Vanhockerhout, M.; et al. Osteopontin Expression Identifies a Subset of Recruited Macrophages Distinct from Kupffer Cells in the Fatty Liver. Immunity 2020, 53, 641–657. [Google Scholar] [CrossRef]
- Wang, J.; Kubes, P. A Reservoir of Mature Cavity Macrophages that Can Rapidly Invade Visceral Organs to Affect Tissue Repair. Cell 2016, 165, 668–678. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Fibrosis Stage 0–2 (N = 31) | Fibrosis Stage 3–4 (N = 33) | p Value ** | |
|---|---|---|---|
| Sex | 0.322 # | ||
| Male | 20 (64.5) | 26 (78.8) | |
| Female | 11 (35.5) | 7 (21.2) | |
| Mean age (±SD) | 54.2 (±11.3) | 50.9 (±8.2) | 0.186 $ |
| Etiology | 0.586 ^ | ||
| CHB | 21 (67.7) | 25 (75.8) | |
| CHC | 2 (6.5) | 0 (0.0) | |
| Alcohol | 3 (9.7) | 2 (6.1) | |
| Others | 5 (16.1) | 6 (18.2) | |
| Diabetes | 6 (19.4) | 11 (33.3) | 0.326 # |
| BMI | 0.085 # | ||
| ≤25 kg/m2 | 25 (80.7) | 19 (57.6) | |
| >25 kg/m2 | 6 (19.4) | 14 (42.4) | |
| ALT | 1.000 # | ||
| ≥31 (F), ≥41 (M) IU/L | 12 (38.7) | 13 (39.4) | |
| AST | 1.000 # | ||
| ≥31 (F), ≥37 (M) IU/L | 19 (61.3) | 20 (60.6) | |
| GGT | (−5) | (−1) | 0.640 # |
| ≥50 (IU/L) | 18 (58.1) | 25 (78.1) | |
| Albumin | 0.644 # | ||
| <4.0 g/dL | 14 (45.2) | 12 (36.4) | |
| Platelets | 0.543 # | ||
| <150 × 109/L | 9 (29.0) | 13 (39.4) | |
| Cholesterol (mmol/L) | (−2) | 0.942 # | |
| ≥200 mg/dL | 5 (16.1) | 7 (21.2) | |
| Antiviral treatment | 3 (9.7) | 8 (24.2) | 0.226 # |
| Fibrosis | |||
| Stage 0 | 7 (22.6) | 0 (0.0) | |
| Stage 1 | 17 (54.8) | 0 (0.0) | |
| Stage 2 | 7 (22.6) | 0 (0.0) | |
| Stage 3 | 0 (0.0) | 15 (45.5) | |
| Stage 4 | 0 (0.0) | 18 (54.6) |
| SEQ | Protein | N | Coef | Logistic Regression p-Value | Wilcoxon Test p-Value | Fold Change | F0–2 (N = 31) | F3–4 (N = 33) |
|---|---|---|---|---|---|---|---|---|
| 1 | CD68 | 64 | 0.00650 | 2.78 × 10−2 | 1.96 × 10−2 | 1.50 | 118.62 | 178.04 |
| 2 | HLA.DR | 64 | 0.02140 | 4.62 × 10−2 | 3.83 × 10−2 | 1.38 | 35.41 | 48.82 |
| 3 | Phospho.c.RAF | 64 | −1.06091 | 4.96 × 10−2 | 4.43 × 10−2 | −1.27 | 1.23 | 0.97 |
| 4 | Cleaved.Caspase.9 | 64 | −0.10881 | 3.22 × 10−2 | 2.59 × 10−2 | −1.28 | 14.62 | 11.46 |
| 5 | CD127 | 64 | −0.48734 | 2.09 × 10−2 | 1.57 × 10−2 | −1.29 | 3.77 | 2.93 |
| 6 | ARG1 | 64 | −0.00693 | 4.21 × 10−2 | 3.52 × 10−2 | −1.31 | 200.13 | 152.53 |
| 7 | Beta.2.microglobulin | 64 | −0.11174 | 2.84 × 10−2 | 2.32 × 10−2 | −1.31 | 12.67 | 9.63 |
| 8 | X4.1BB | 64 | −1.28366 | 1.77 × 10−2 | 1.22 × 10−2 | −1.35 | 1.41 | 1.04 |
| 9 | LAG3 | 64 | −0.94575 | 4.71 × 10−2 | 4.05 × 10−2 | −1.37 | 1.17 | 0.85 |
| 10 | B7.H3 | 64 | −0.07086 | 3.41 × 10−2 | 2.50 × 10−2 | −1.46 | 19.21 | 13.17 |
| 11 | VISTA | 64 | −0.41498 | 2.02 × 10−2 | 1.27 × 10−2 | −1.50 | 3.58 | 2.39 |
| 12 | Tim.3 | 64 | −0.50608 | 1.91 × 10−3 | 3.18 × 10−4 | −1.51 | 5.56 | 3.68 |
| 13 | pan.RAS | 64 | −1.75375 | 1.46 × 10−3 | 6.02 × 10−4 | −1.70 | 1.69 | 1.00 |
| 14 | OX40L | 64 | −1.13933 | 8.75 × 10−3 | 1.76 × 10−3 | −2.60 | 2.69 | 1.03 |
| 15 | STING | 64 | −0.09585 | 8.76 × 10−3 | 4.27 × 10−3 | −2.65 | 19.19 | 7.26 |
| SEQ | Protein | p_Value | AUROC | Sensitivity | Specificity | Accuracy | PPV | NPV |
|---|---|---|---|---|---|---|---|---|
| 1 | CD68_HLA.DR_OX40L_Phospho.c.RAF_STING_Tim.3 | 1.16 × 10−6 | 0.873 | 81.82 | 93.55 | 87.50 | 93.10 | 82.86 |
| 2 | ARG1_B7.H3_CD127_CD68_HLA.DR_OX40L_pan.RAS_STING_Tim.3 | 5.19 × 10−7 | 0.894 | 87.88 | 83.87 | 85.94 | 85.29 | 86.67 |
| 3 | B7.H3_CD68_HLA.DR_OX40L_Phospho.c.RAF_STING_VISTA | 5.19 × 10−7 | 0.874 | 87.88 | 83.87 | 85.94 | 85.29 | 86.67 |
| 4 | Beta.2.microglobulin_CD127_CD68_HLA.DR_OX40L_Tim.3 | 5.06 × 10−7 | 0.878 | 84.85 | 87.10 | 85.94 | 87.50 | 84.38 |
| 5 | CD68_HLA.DR_OX40L_pan.RAS_STING | 5.06 × 10−7 | 0.870 | 84.85 | 87.10 | 85.94 | 87.50 | 84.38 |
| Univariable Logistic Regression | |||||
|---|---|---|---|---|---|
| Variable | n | Coefficient | Se (Coefficient) | Odds Ratio (95% CI) | p-Value |
| CD68_HLA-DR_OX40L_Phospho-c-RAF_STING_Tim-3 (low vs. high) | 64 | 4.178 | 0.859 | 65.25 (12.11–351.49) | 1.16 × 10−6 |
| Age (≤55 years vs. >55 years) | 64 | −0.945 | 0.543 | 0.39 (0.13–1.13) | 0.082 |
| Sex (male vs. female) | 64 | −0.714 | 0.568 | 0.49 (0.16–1.49) | 0.208 |
| Etiology | |||||
| CHB (absent vs. present) | 64 | 0.397 | 0.559 | 1.49 (0.50–4.45) | 0.477 |
| CHC (absent vs. present) | 64 | −16.695 | 1696.734 | 0.00 (0.00–Inf) | 0.992 |
| Alcohol (absent vs. present) | 64 | −0.507 | 0.949 | 0.60 (0.09–3.87) | 0.593 |
| Others (absent vs. present) | 64 | 0.145 | 0.665 | 1.16 (0.31–4.25) | 0.828 |
| BMI (≤25 kg/m2 vs. >25 kg/m2) | 64 | 1.122 | 0.575 | 3.07 (0.99–9.48) | 0.051 |
| Diabetes (absent vs. present) | 64 | 0.734 | 0.586 | 2.08 (0.66–6.57) | 0.210 |
| ALT (<31(F), <41(M) IU/L vs. ≥31(F), ≥41(M) IU/L) | 64 | 0.029 | 0.513 | 1.03 (0.38–2.81) | 0.955 |
| AST (<31(F), <37(M) IU/L vs. ≥31(F), ≥37(M) IU/L) | 64 | −0.029 | 0.513 | 0.97 (0.36–2.65) | 0.955 |
| GGT (<50 IU/L vs.≥50 IU/L) | 58 | 0.462 | 0.603 | 1.59 (0.49–5.17) | 0.443 |
| Albumin (<4.0 g/dL vs.≥4.0 g/dL) | 64 | 0.366 | 0.511 | 1.44 (0.53–3.92) | 0.475 |
| Platelets (<150 × 109/L vs. ≥150 × 109/L) | 64 | −0.463 | 0.532 | 0.63 (0.22–1.79) | 0.384 |
| Cholesterol (<200 mg/dL vs. ≥200 mg/dL) | 62 | 0.256 | 0.650 | 1.29 (0.36–4.62) | 0.693 |
| Multivariable Logistic Regression | |||||
| Variable | Coefficient | Odds Ratio (95% CI) | p-Value | ||
| CD68_HLA-DR_OX40L_Phospho-c-RAF_STING_Tim-3 (Low vs. High) | 4.646 | 104.13 (14.29–758.66) | 4.54 × 10−6 | ||
| Age (≤55 years vs. >55 years) | −0.558 | 0.57 (0.10–3.29) | 0.532 | ||
| BMI (≤25 kg/m2 vs. >25 kg/m2) | 2.097 | 8.15 (1.28–51.93) | 0.026 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, J.; Kim, C.M.; Cha, J.H.; Park, J.Y.; Yu, Y.S.; Wang, H.J.; Sung, P.S.; Jung, E.S.; Bae, S.H. Multiplexed Digital Spatial Protein Profiling Reveals Distinct Phenotypes of Mononuclear Phagocytes in Livers with Advanced Fibrosis. Cells 2022, 11, 3387. https://doi.org/10.3390/cells11213387
Lee J, Kim CM, Cha JH, Park JY, Yu YS, Wang HJ, Sung PS, Jung ES, Bae SH. Multiplexed Digital Spatial Protein Profiling Reveals Distinct Phenotypes of Mononuclear Phagocytes in Livers with Advanced Fibrosis. Cells. 2022; 11(21):3387. https://doi.org/10.3390/cells11213387
Chicago/Turabian StyleLee, Jaejun, Chang Min Kim, Jung Hoon Cha, Jin Young Park, Yun Suk Yu, Hee Jung Wang, Pil Soo Sung, Eun Sun Jung, and Si Hyun Bae. 2022. "Multiplexed Digital Spatial Protein Profiling Reveals Distinct Phenotypes of Mononuclear Phagocytes in Livers with Advanced Fibrosis" Cells 11, no. 21: 3387. https://doi.org/10.3390/cells11213387
APA StyleLee, J., Kim, C. M., Cha, J. H., Park, J. Y., Yu, Y. S., Wang, H. J., Sung, P. S., Jung, E. S., & Bae, S. H. (2022). Multiplexed Digital Spatial Protein Profiling Reveals Distinct Phenotypes of Mononuclear Phagocytes in Livers with Advanced Fibrosis. Cells, 11(21), 3387. https://doi.org/10.3390/cells11213387