
Wnt Signaling Rescues Amyloid Beta-Induced Gut Stem Cell Loss

 , , and
, , and {kind=link}
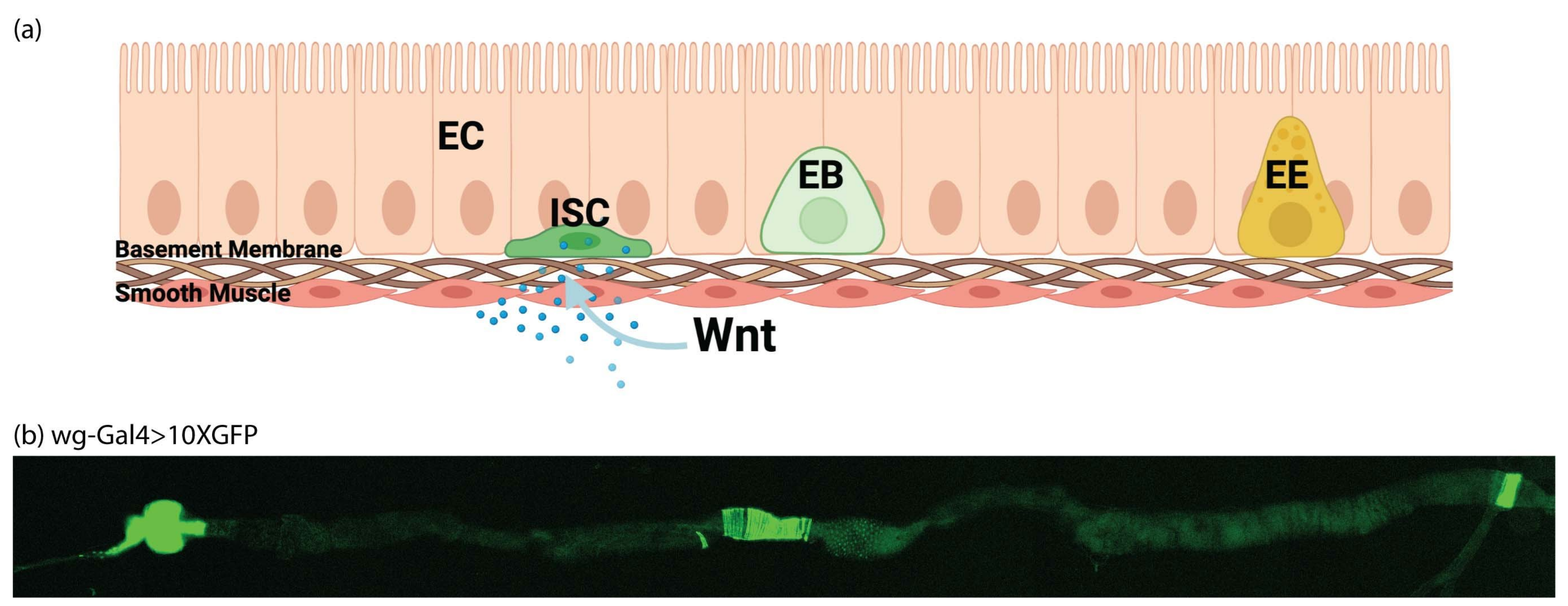{kind=link}
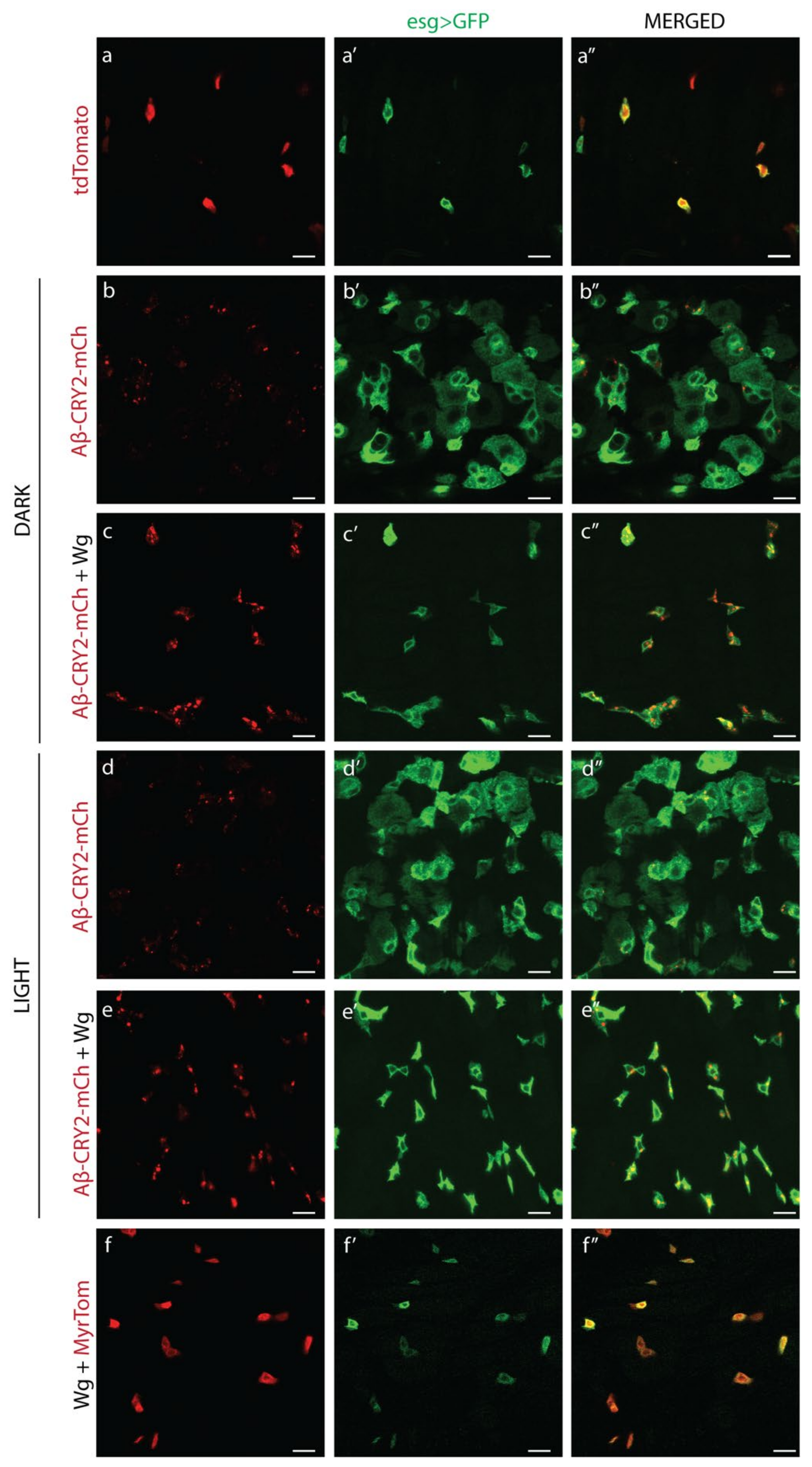{kind=link}
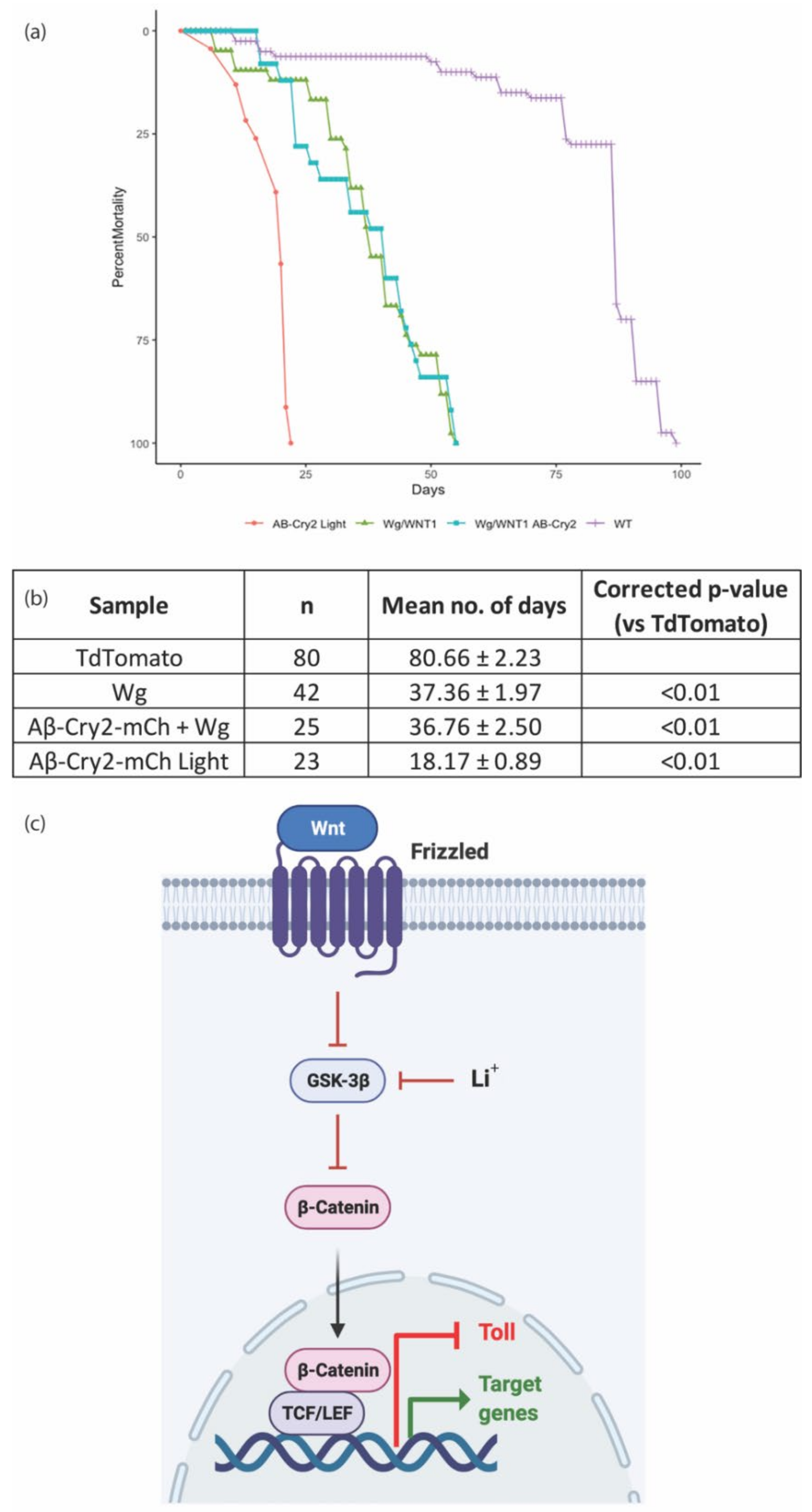{kind=link}
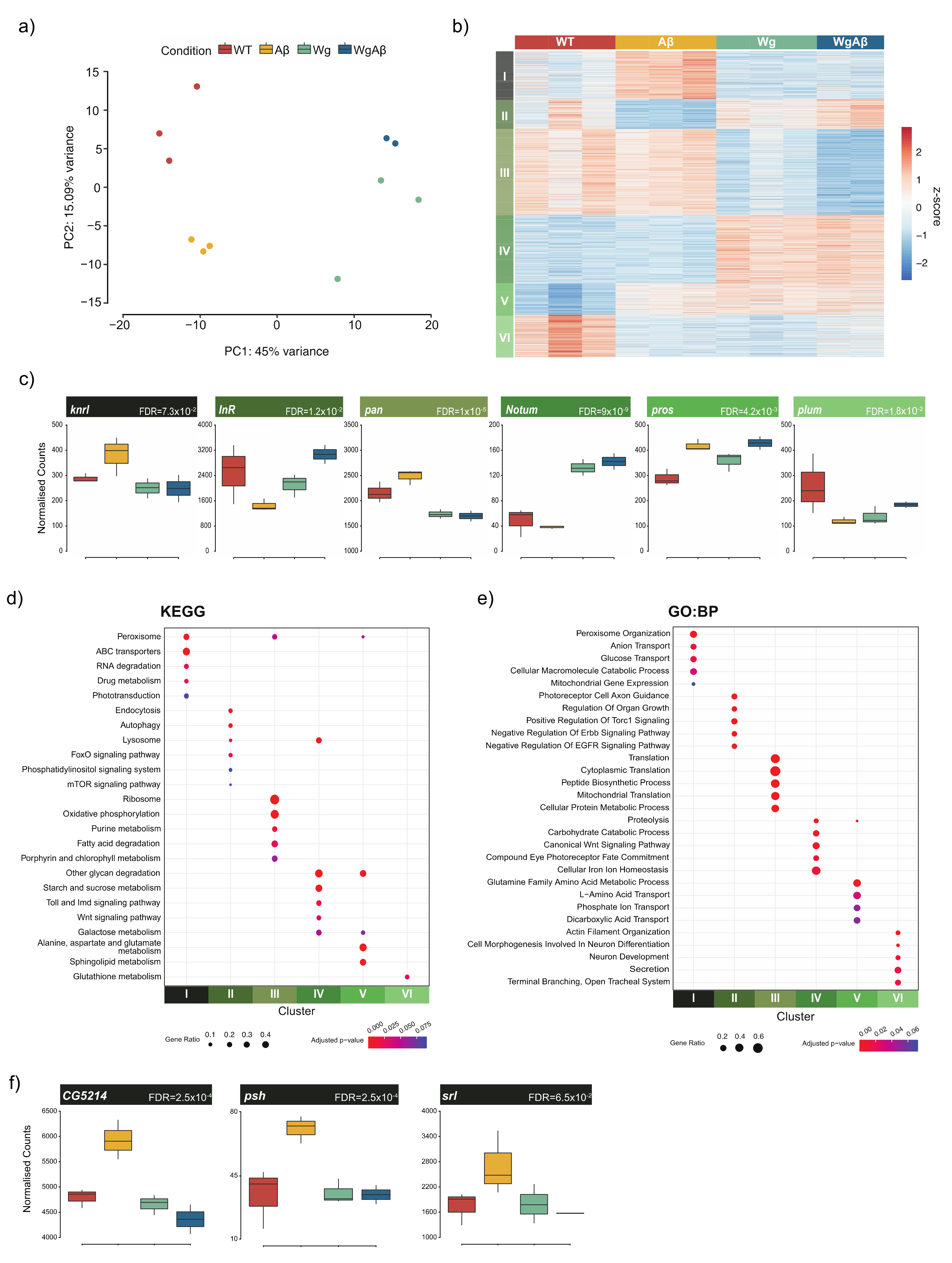{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Crosses and Expression of UAS Construct
- Elav-Gal4: BDSC 458 [39]
- Repo-QF2: BDSC 66477 [40]
- QUAS-6XGFP: BDSC 52263 [41]
- UAS-Aβ-CRY2-mCherry: [11]
- UAS-mCD8.RFP: BDSC 32220 [42]
- UAS-wg: BDSC 5918 [43]
- UAS-Td-Tomato: BDSC 36328 (Joost Schulte and Katharine Sepp)
- Tl(CRISPaint.T2A-GAL4)wg: BDSC #83627 [44]
- UAS-10XGFP: BDSC 32185 [42]
- Esg-Gal4, UAS-GFP; tub-Gal80ts, UAS-dCas9.VPR: BDSC 67069 [45]
- UAS-myr::tdTomato: BDSC 32222 [42]
- UAS-Toll-Cry2-mCherry: [46]
- Arm-Gal4: BDSC 1560 [47]
- ArmGal4; tub-GAL80ts: BDSC 86327 [38]
- TRiP.HMC03615 attP40 Persephone RNAi [48]
- TRiP.HM05191 attP2 Dif RNAi [48]
- UAS-Dsh [49].
- (1)
- Elav-Gal4; Repo-QF2, QUAS-GFP × UAS-Aβ-CRY2-mCh
- (2)
- Elav-Gal4, UAS-RFP × UAS-wg
- (3)
- Elav-Gal4 × UAS-Td-Tomato
- (4)
- Tl(CRISPaint.T2A-GAL4)wg × UAS-10XGFP
- (5)
- w; UAS-TdTomato X esg-Gal4, UAS-GFP; tubP-GAL80ts
- (6)
- w; UAS-Aβ1−42-CRY2-mCh X esg-Gal4, UAS-GFP; tubP-GAL80ts
- (7)
- w; UAS-wg; UAS-Aβ1−42-CRY2-mCh X esg-Gal4, UAS-GFP; tub-GAL80ts
- (8)
- w; UAS-wg, UAS-myr-Tomato × esg-Gal4, UAS-GFP; tub-GAL80ts
- (9)
- w; UAS-wg × esg-Gal4, UAS-GFP; tub-GAL80ts
- (10)
- w; UAS-Toll-Cry2-mCh × esg-Gal4, UAS-GFP; tub-GAL80ts
- (11)
- w; UAS-Toll-Cry2-mCh × ArmGal4; tub-GAL80ts
- (12)
- w; UAS-Td-Tomato × esg-Gal4, UAS-GFP; tub-GAL80ts
- (13)
- w; UAS-Td-Tomato × ArmGal4; tub-GAL80ts
- (14)
- w; UAS-Persephone-RNAi × ArmGal4; tub-GAL80ts
- (15)
- w; UAS-Dif-RNAi × ArmGal4; tub-GAL80ts.
2.2. Light-Sheet Microscopy
2.2.1. Gut Preparations and Fluorescence Microscopy
2.2.2. RNA Preparation and RNA-Sequencing
2.2.3. RNA-Seq Analysis
2.2.4. Lifespan Studies
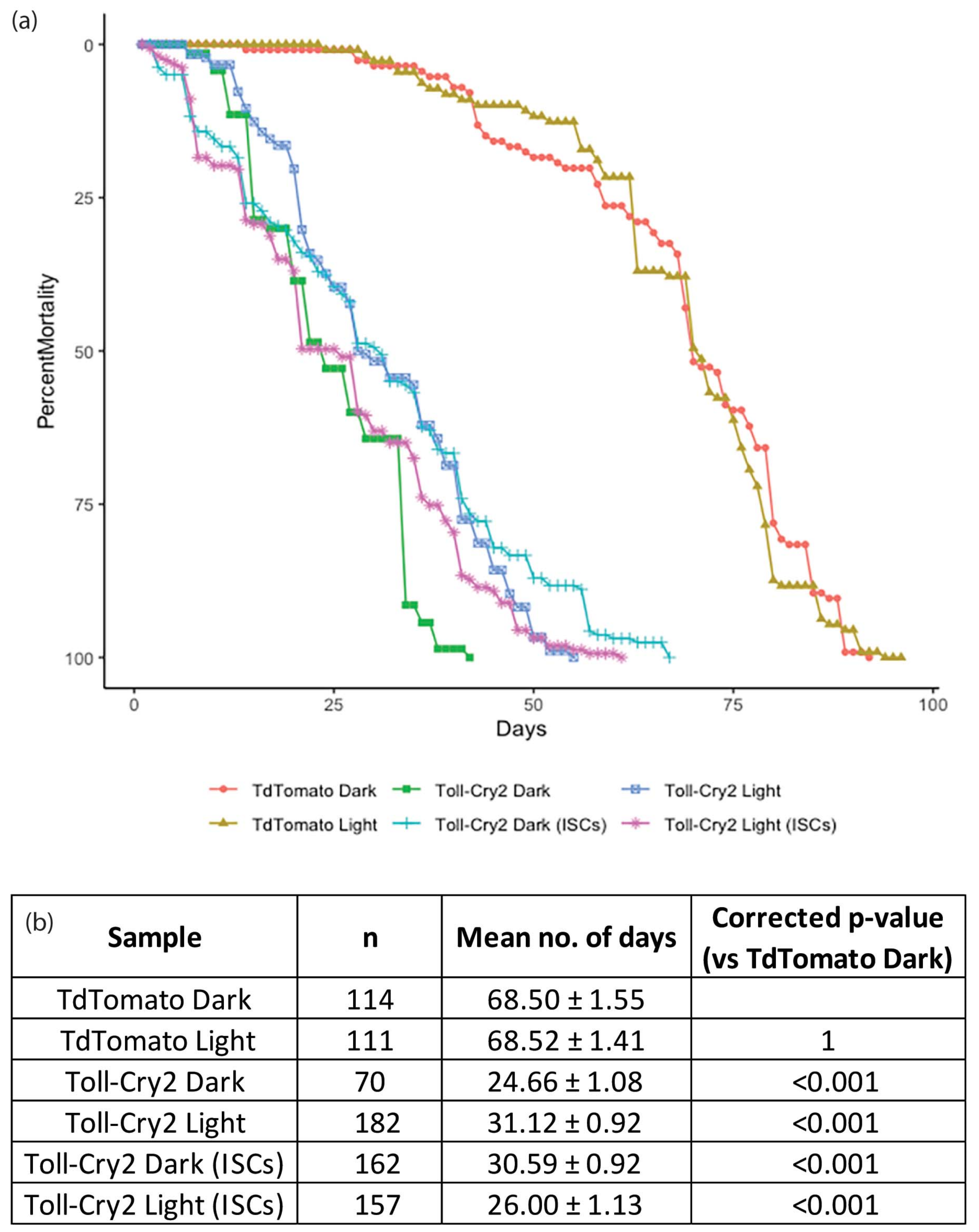
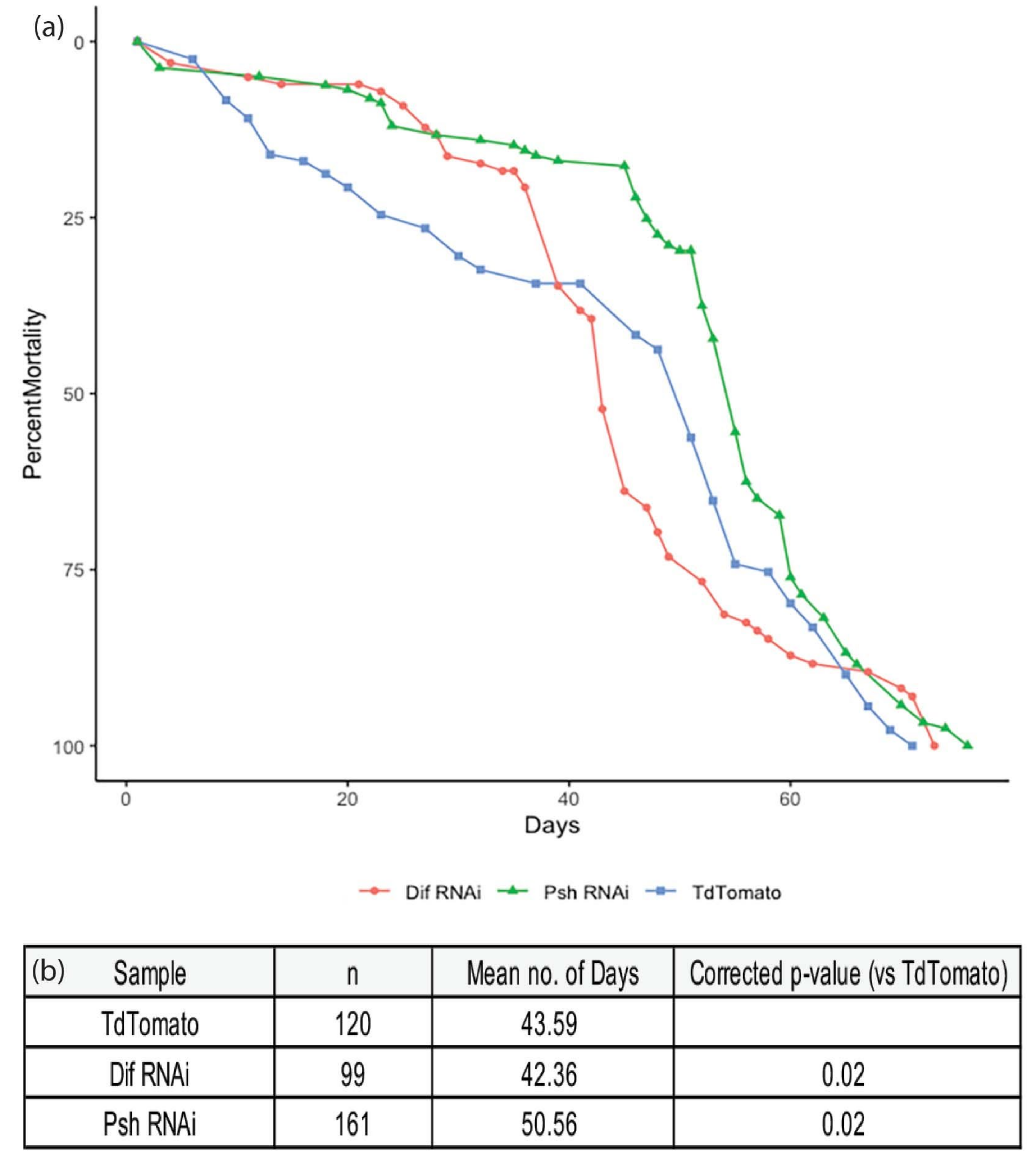
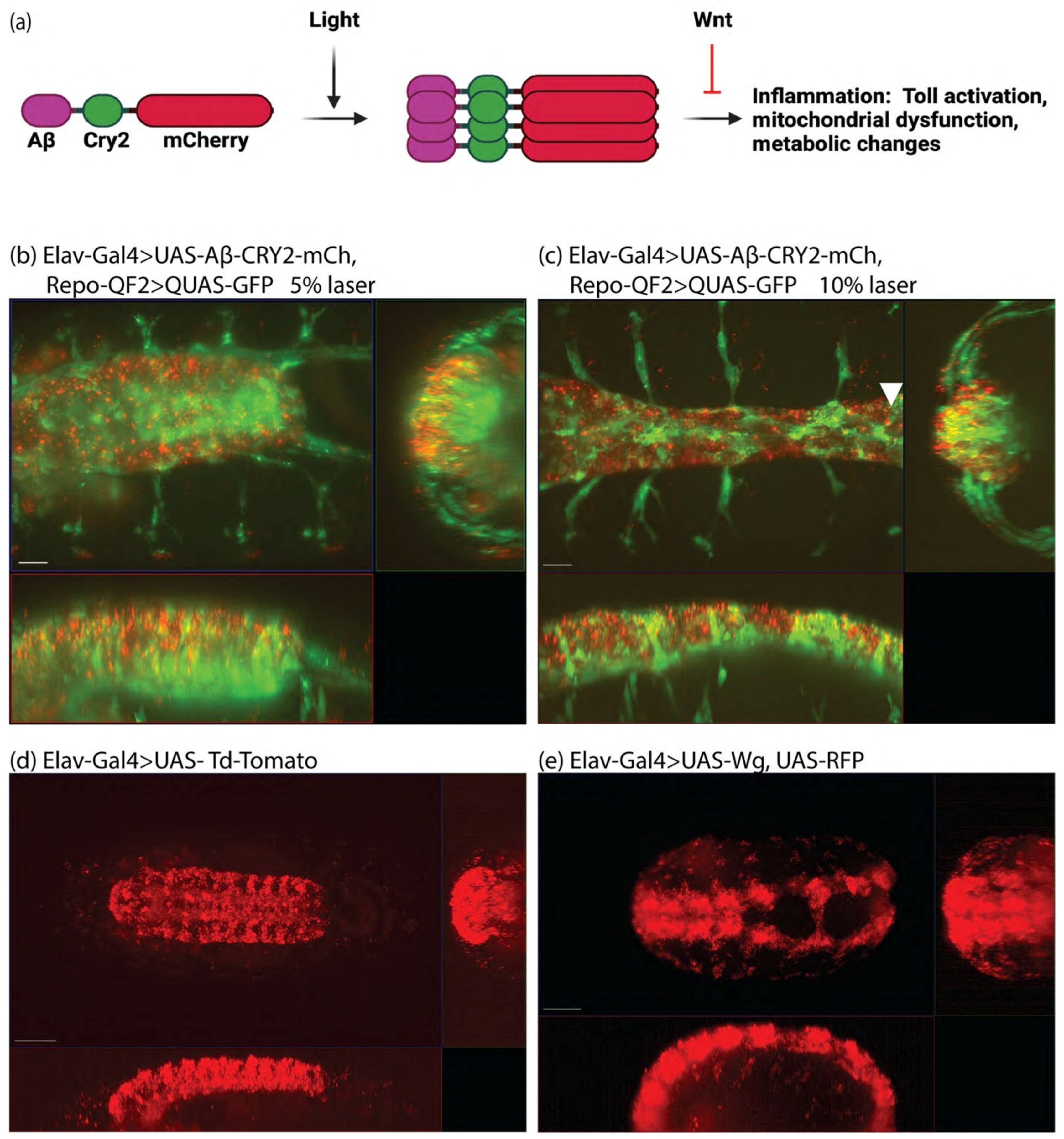
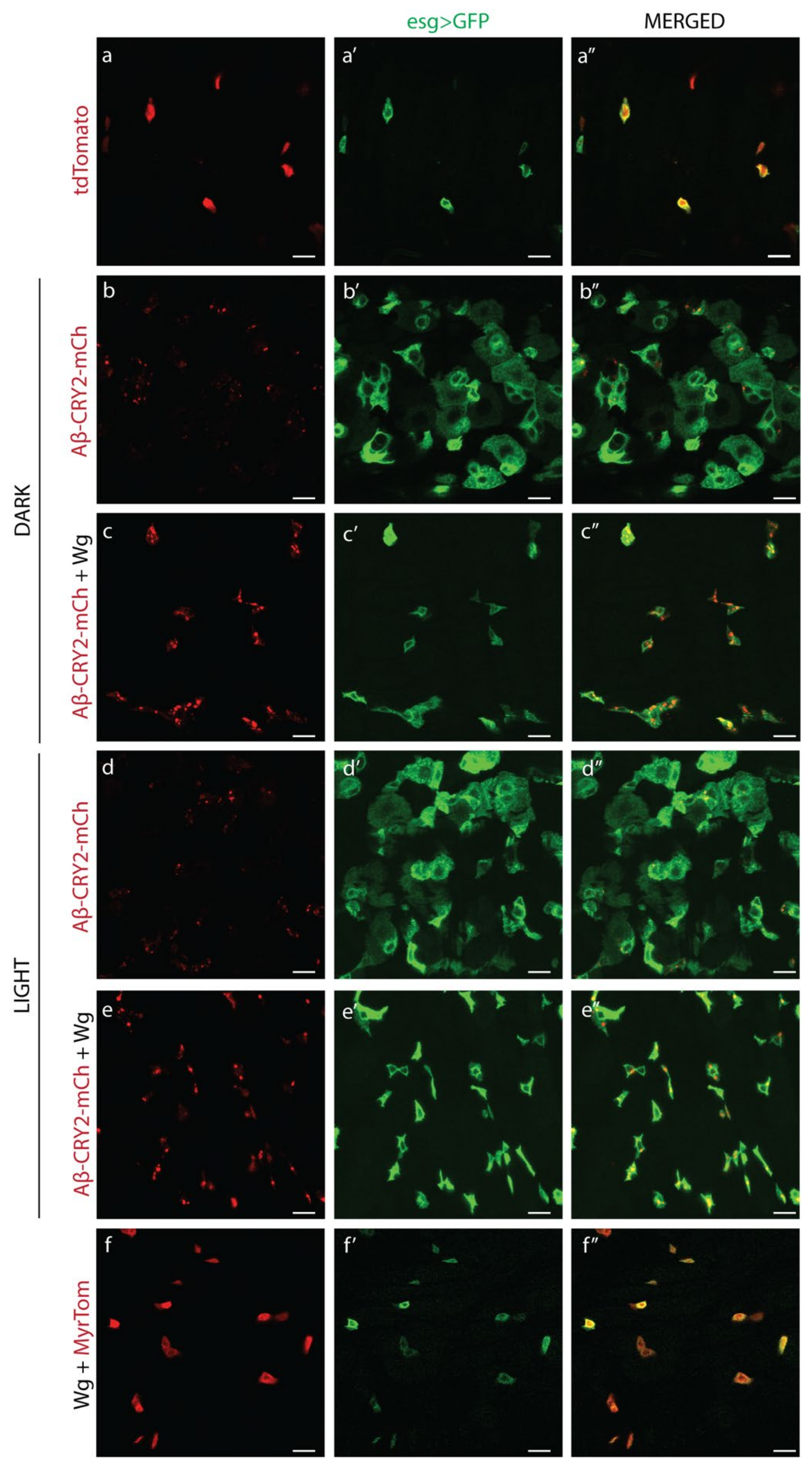
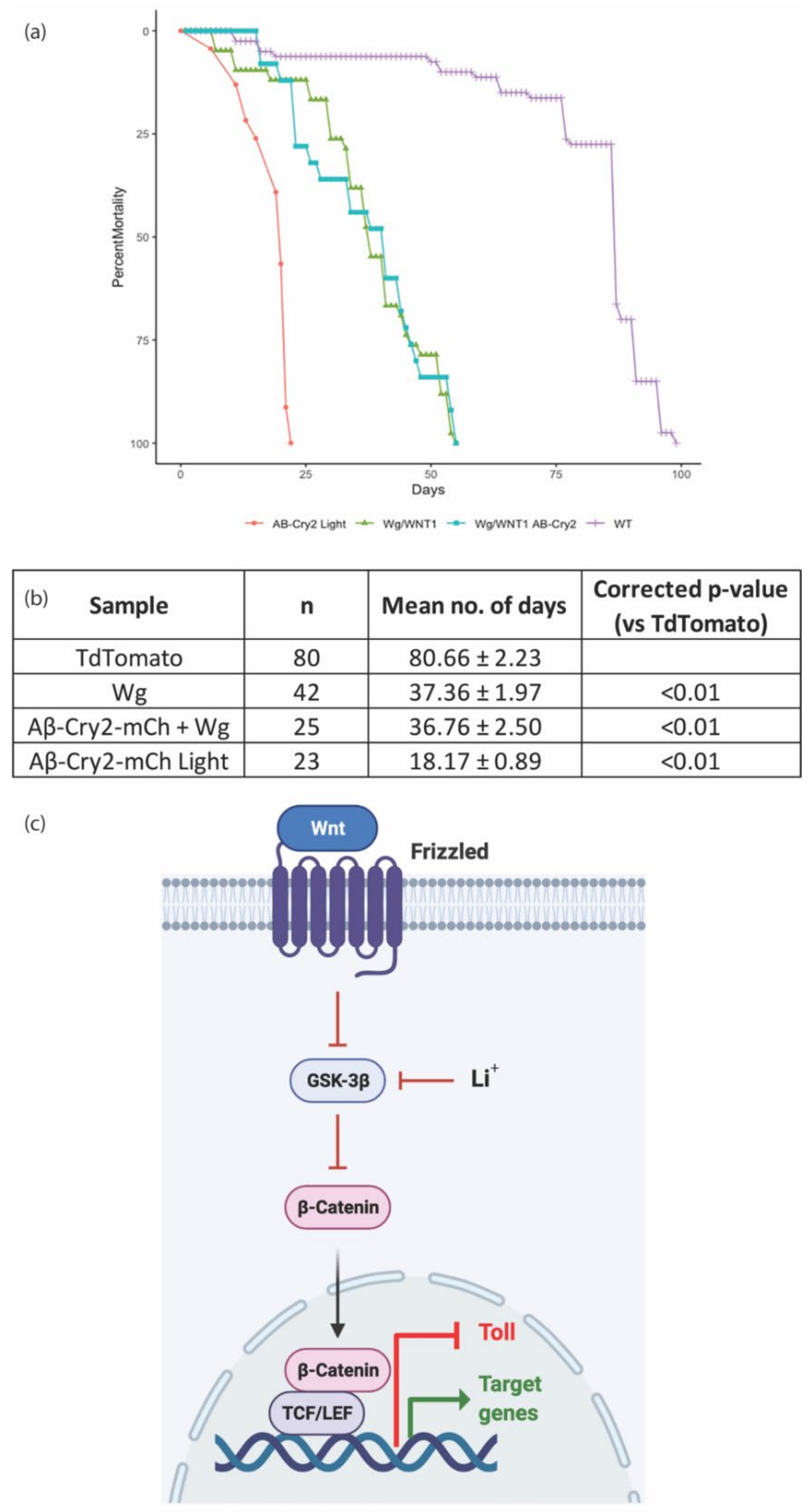
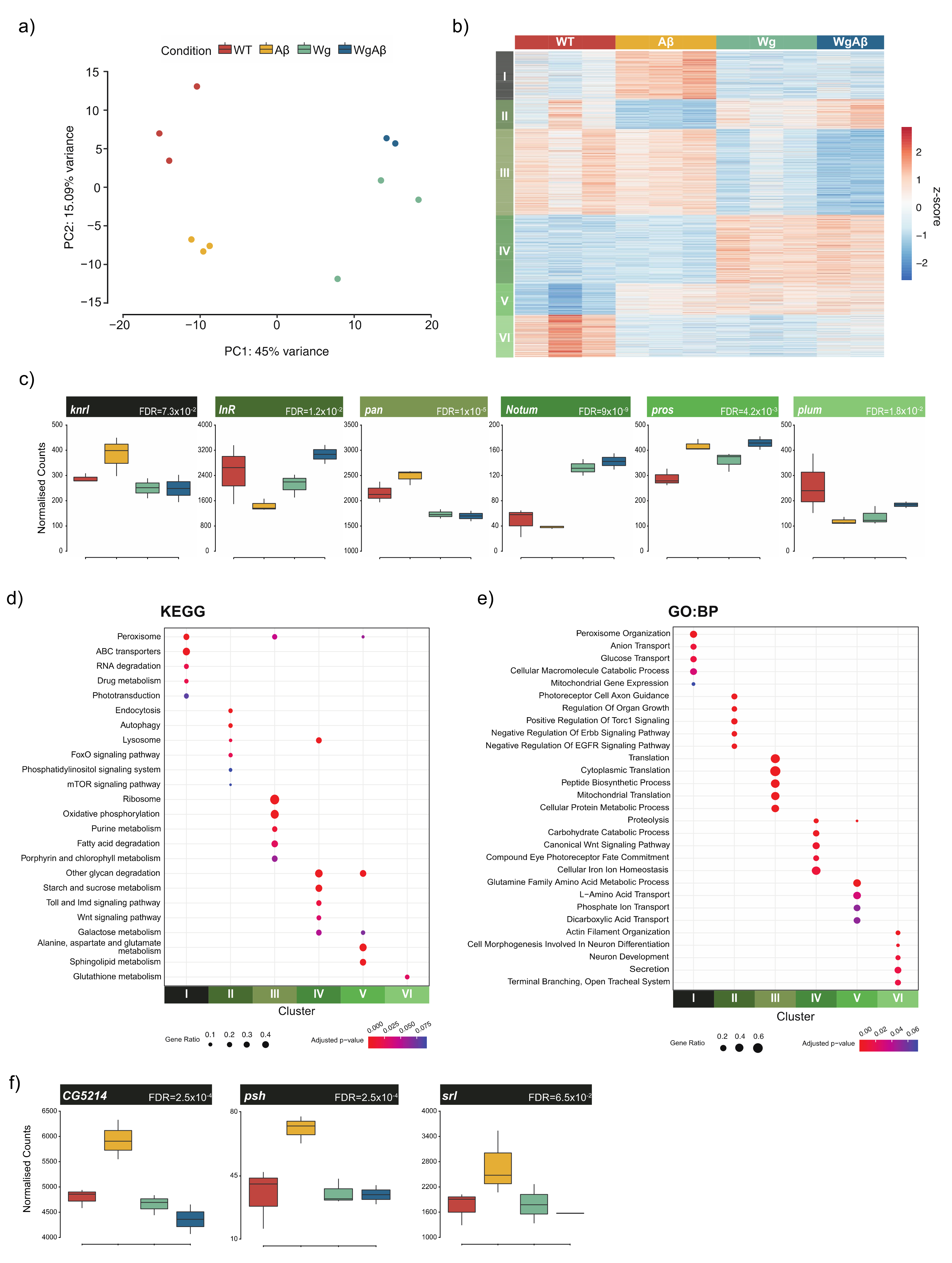
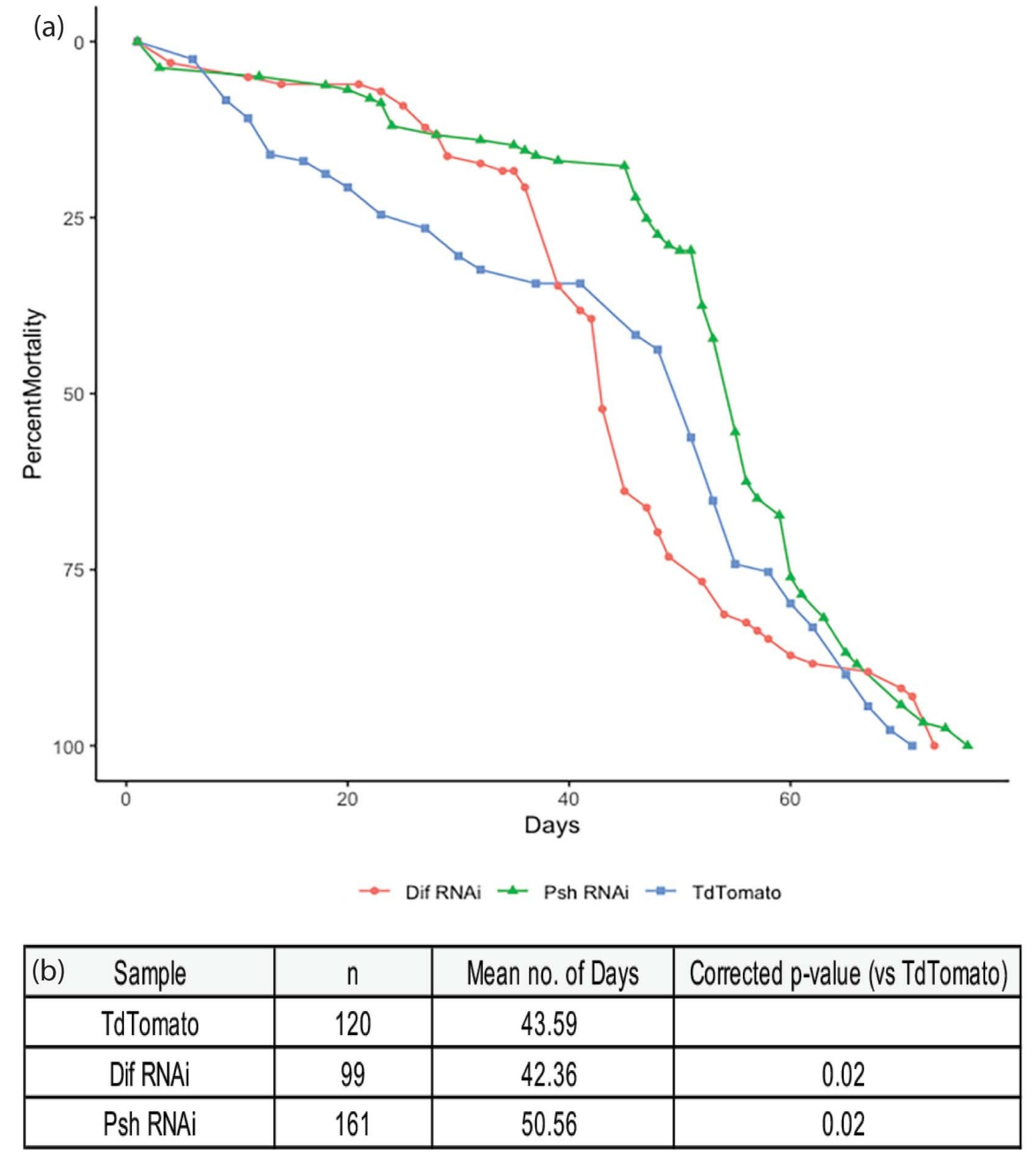
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- De-Paula, V.J.; Radanovic, M.; Diniz, B.S.; Forlenza, O.V. Alzheimer’s disease. Sub-Cell. Biochem. 2012, 65, 329–352. [Google Scholar]
- Kumar, A.; Singh, A.; Ekavali. A review on alzheimer’s disease pathophysiology and its management: An update. Pharmacol. Rep. PR 2015, 67, 195–203. [Google Scholar] [CrossRef] [PubMed]
- Anderson, R.M.; Hadjichrysanthou, C.; Evans, S.; Wong, M.M. Why do so many clinical trials of therapies for alzheimer’s disease fail? Lancet 2017, 390, 2327–2329. [Google Scholar] [CrossRef]
- Lalli, G.; Schott, J.M.; Hardy, J.; De Strooper, B. Aducanumab: A new phase in therapeutic development for alzheimer’s disease? EMBO Mol. Med. 2021, 13, e14781. [Google Scholar] [CrossRef] [PubMed]
- Sevigny, J.; Chiao, P.; Bussiere, T.; Weinreb, P.H.; Williams, L.; Maier, M.; Dunstan, R.; Salloway, S.; Chen, T.; Ling, Y.; et al. The antibody aducanumab reduces abeta plaques in alzheimer’s disease. Nature 2016, 537, 50–56. [Google Scholar] [CrossRef] [PubMed]
- Cummings, J. Lessons learned from alzheimer disease: Clinical trials with negative outcomes. Clin. Transl. Sci. 2018, 11, 147–152. [Google Scholar] [CrossRef] [PubMed]
- Cummings, J.; Lee, G.; Ritter, A.; Zhong, K. Alzheimer’s disease drug development pipeline: 2018. Alzheimer’s Dement. 2018, 4, 195–214. [Google Scholar] [CrossRef]
- Hardy, J.A.; Higgins, G.A. Alzheimer’s disease: The amyloid cascade hypothesis. Science 1992, 256, 184–185. [Google Scholar] [CrossRef]
- Driscoll, I.; Troncoso, J. Asymptomatic alzheimer’s disease: A prodrome or a state of resilience? Curr. Alzheimer Res. 2011, 8, 330–335. [Google Scholar] [CrossRef]
- Ferreira, S.T.; Klein, W.L. The abeta oligomer hypothesis for synapse failure and memory loss in alzheimer’s disease. Neurobiol. Learn. Mem. 2011, 96, 529–543. [Google Scholar] [CrossRef] [Green Version]
- Lim, C.H.; Kaur, P.; Teo, E.; Lam, V.Y.M.; Zhu, F.; Kibat, C.; Gruber, J.; Mathuru, A.S.; Tolwinski, N.S. Application of optogenetic amyloid-beta distinguishes between metabolic and physical damages in neurodegeneration. eLife 2020, 9, e52589. [Google Scholar] [CrossRef]
- Lim, W.K.; Kaur, P.; Huang, H.; Jo, R.S.; Ramamoorthy, A.; Ng, L.F.; Suresh, J.; Maisha, F.I.; Mathuru, A.S.; Tolwinski, N.S. Optogenetic approaches for understanding homeostatic and degenerative processes in drosophila. Cell. Mol. Life Sci. 2021, 78, 5865–5880. [Google Scholar] [CrossRef] [PubMed]
- Möglich, A.; Moffat, K. Engineered photoreceptors as novel optogenetic tools. Photochem. Photobiol. Sci. 2010, 9, 1286–1300. [Google Scholar] [CrossRef] [PubMed]
- Fenno, L.; Yizhar, O.; Deisseroth, K. The development and application of optogenetics. Annu. Rev. Neurosci. 2011, 34, 389–412. [Google Scholar] [CrossRef] [PubMed]
- Mas, P.; Devlin, P.F.; Panda, S.; Kay, S.A. Functional interaction of phytochrome b and cryptochrome 2. Nature 2000, 408, 207–211. [Google Scholar] [CrossRef]
- Clevers, H. Wnt/beta-catenin signaling in development and disease. Cell 2006, 127, 469–480. [Google Scholar] [CrossRef] [Green Version]
- Wodarz, A.; Nusse, R. Mechanisms of wnt signaling in development. Annu. Rev. Cell. Dev. Biol. 1998, 14, 59–88. [Google Scholar] [CrossRef] [Green Version]
- Hu, D.J.; Yun, J.; Elstrott, J.; Jasper, H. Non-canonical wnt signaling promotes directed migration of intestinal stem cells to sites of injury. Nat. Commun. 2021, 12, 7150. [Google Scholar] [CrossRef]
- Dunn, N.R.; Tolwinski, N.S. Ptk7 and mcc, unfancied components in non-canonical wnt signaling and cancer. Cancers 2016, 8, 68. [Google Scholar] [CrossRef] [Green Version]
- Tian, A.; Benchabane, H.; Wang, Z.; Ahmed, Y. Regulation of stem cell proliferation and cell fate specification by wingless/wnt signaling gradients enriched at adult intestinal compartment boundaries. PLoS Genet. 2016, 12, e1005822. [Google Scholar] [CrossRef] [Green Version]
- Tian, A.; Duwadi, D.; Benchabane, H.; Ahmed, Y. Essential long-range action of wingless/wnt in adult intestinal compartmentalization. PLoS Genet. 2019, 15, e1008111. [Google Scholar] [CrossRef] [PubMed]
- Ng, L.F.; Kaur, P.; Bunnag, N.; Suresh, J.; Sung, I.C.H.; Tan, Q.H.; Gruber, J.; Tolwinski, N.S. Wnt signaling in disease. Cells 2019, 8, 826. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suresh, J.; Khor, I.W.; Kaur, P.; Heng, H.L.; Torta, F.; Dawe, G.S.; Tai, E.S.; Tolwinski, N.S. Shared signaling pathways in alzheimer’s and metabolic disease may point to new treatment approaches. FEBS J. 2021, 288, 3855–3873. [Google Scholar] [CrossRef]
- Ali, A.; Ali, A.; Ahmad, W.; Ahmad, N.; Khan, S.; Nuruddin, S.M.; Husain, I. Deciphering the role of wnt signaling in metabolic syndrome–linked alzheimer’s disease. Mol. Neurobiol. 2020, 57, 302–314. [Google Scholar] [CrossRef] [PubMed]
- Nusse, R.; Clevers, H. Wnt/beta-catenin signaling, disease, and emerging therapeutic modalities. Cell 2017, 169, 985–999. [Google Scholar] [CrossRef]
- Clevers, H.; Nusse, R. Wnt/beta-catenin signaling and disease. Cell 2012, 149, 1192–1205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cadigan, K.M.; Nusse, R. Wnt signaling: A common theme in animal development. Genes Dev. 1997, 11, 3286–3305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schlessinger, K.; Hall, A.; Tolwinski, N. Wnt signaling pathways meet rho gtpases. Genes Dev. 2009, 23, 265–277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tolwinski, N.S.; Wieschaus, E. A nuclear escort for beta-catenin. Nat. Cell. Biol. 2004, 6, 579–580. [Google Scholar] [CrossRef] [PubMed]
- Kaplan, N.A.; Colosimo, P.F.; Liu, X.; Tolwinski, N.S. Complex interactions between gsk3 and apkc in drosophila embryonic epithelial morphogenesis. PLoS One 2011, 6, e18616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martinez, M.; Inestrosa, N.C. The transcriptional landscape of alzheimer’s disease and its association with wnt signaling pathway. Neurosci. Biobehav. Rev. 2021, 128, 454–466. [Google Scholar] [CrossRef]
- Inestrosa, N.C.; Varela-Nallar, L. Wnt signaling in the nervous system and in alzheimer’s disease. J. Mol. Cell Biol 2014, 6, 64–74. [Google Scholar] [CrossRef]
- Yang, Y.; Zhang, Z. Microglia and wnt pathways: Prospects for inflammation in alzheimer’s disease. Front. Aging Neurosci. 2020, 12, 110. [Google Scholar] [CrossRef]
- Jia, L.; Piña-Crespo, J.; Li, Y. Restoring wnt/β-catenin signaling is a promising therapeutic strategy for alzheimer’s disease. Mol. Brain 2019, 12, 1–11. [Google Scholar] [CrossRef]
- Teo, E.; Ravi, S.; Barardo, D.; Kim, H.S.; Fong, S.; Cazenave-Gassiot, A.; Tan, T.Y.; Ching, J.; Kovalik, J.P.; Wenk, M.R.; et al. Metabolic stress is a primary pathogenic event in transgenic caenorhabditis elegans expressing pan-neuronal human amyloid beta. eLife 2019, 8, e50069. [Google Scholar] [CrossRef] [PubMed]
- Kaur, P.; Saunders, T.E.; Tolwinski, N.S. Coupling optogenetics and light-sheet microscopy, a method to study wnt signaling during embryogenesis. Sci. Rep. 2017, 7, 16636. [Google Scholar] [CrossRef] [PubMed]
- Brand, A.H.; Perrimon, N. Targeted gene expression as a means of altering cell fates and generating dominant phenotypes. Development 1993, 118, 401–415. [Google Scholar] [CrossRef]
- McGuire, S.E.; Le, P.T.; Osborn, A.J.; Matsumoto, K.; Davis, R.L. Spatiotemporal rescue of memory dysfunction in drosophila. Science 2003, 302, 1765–1768. [Google Scholar] [CrossRef] [Green Version]
- Lin, D.M.; Goodman, C.S. Ectopic and increased expression of fasciclin ii alters motoneuron growth cone guidance. Neuron 1994, 13, 507–523. [Google Scholar] [CrossRef]
- Potter, C.J.; Tasic, B.; Russler, E.V.; Liang, L.; Luo, L. The q system: A repressible binary system for transgene expression, lineage tracing, and mosaic analysis. Cell 2010, 141, 536–548. [Google Scholar] [CrossRef] [Green Version]
- Shearin, H.K.; Macdonald, I.S.; Spector, L.P.; Stowers, R.S. Hexameric gfp and mcherry reporters for the drosophila gal4, q, and lexa transcription systems. Genetics 2014, 196, 951–960. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pfeiffer, B.D.; Ngo, T.T.; Hibbard, K.L.; Murphy, C.; Jenett, A.; Truman, J.W.; Rubin, G.M. Refinement of tools for targeted gene expression in drosophila. Genetics 2010, 186, 735–755. [Google Scholar] [CrossRef] [Green Version]
- Hays, R.; Gibori, G.B.; Bejsovec, A. Wingless signaling generates pattern through two distinct mechanisms. Development 1997, 124, 3727–3736. [Google Scholar] [CrossRef] [PubMed]
- Bosch, J.A.; Colbeth, R.; Zirin, J.; Perrimon, N. Gene knock-ins in drosophila using homology-independent insertion of universal donor plasmids. Genetics 2020, 214, 75–89. [Google Scholar] [CrossRef]
- Micchelli, C.A.; Perrimon, N. Evidence that stem cells reside in the adult drosophila midgut epithelium. Nature 2006, 439, 475–479. [Google Scholar] [CrossRef]
- Bunnag, N.; Tan, Q.H.; Kaur, P.; Ramamoorthy, A.; Sung, I.C.H.; Lusk, J.; Tolwinski, N.S. An optogenetic method to study signal transduction in intestinal stem cell homeostasis. J. Mol. Biol 2020, 432, 3159–3176. [Google Scholar] [CrossRef]
- White, P.; Vincent, J.-P. Uncoupling cadherin-based adhesion from wingless signalling in drosophila. Nature 1996, 383, 627–630. [Google Scholar]
- Perkins, L.A.; Holderbaum, L.; Tao, R.; Hu, Y.; Sopko, R.; McCall, K.; Yang-Zhou, D.; Flockhart, I.; Binari, R.; Shim, H.-S. The transgenic rnai project at harvard medical school: Resources and validation. Genetics 2015, 201, 843–852. [Google Scholar] [CrossRef]
- Kaur, P.; Lam, V.Y.M.; Mannava, A.G.; Suresh, J.; Jenny, A.; Tolwinski, N.S. Membrane targeting of disheveled can bypass the need for arrow/lrp5. Sci. Rep. 2017, 7, 6934. [Google Scholar] [CrossRef]
- Kaur, P.; Jin, H.J.; Lusk, J.B.; Tolwinski, N.S. Modeling the role of wnt signaling in human and drosophila stem cells. Genes 2018, 9, 201. [Google Scholar] [CrossRef] [Green Version]
- Kaur, P.; Kibat, C.; Teo, E.; Gruber, J.; Mathuru, A.; Tolwinski, N.S. Use of optogenetic amyloid-β to monitor protein aggregation in drosophila melanogaster, danio rerio and caenorhabditis elegans. Bio-Protocol 2020, 10, e3856. [Google Scholar] [CrossRef]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. Nih image to imagej: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef] [PubMed]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. Star: Ultrafast universal rna-seq aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Dewey, C.N. Rsem: Accurate transcript quantification from rna-seq data with or without a reference genome. BMC Bioinform. 2011, 12, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for rna-seq data with deseq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [Green Version]
- Chen, E.Y.; Tan, C.M.; Kou, Y.; Duan, Q.; Wang, Z.; Meirelles, G.V.; Clark, N.R.; Ma’ayan, A. Enrichr: Interactive and collaborative html5 gene list enrichment analysis tool. BMC Bioinform. 2013, 14, 128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuleshov, M.V.; Jones, M.R.; Rouillard, A.D.; Fernandez, N.F.; Duan, Q.; Wang, Z.; Koplev, S.; Jenkins, S.L.; Jagodnik, K.M.; Lachmann, A.; et al. Enrichr: A comprehensive gene set enrichment analysis web server 2016 update. Nucleic Acids Res. 2016, 44, W90–W97. [Google Scholar] [CrossRef] [Green Version]
- Xie, Z.; Bailey, A.; Kuleshov, M.V.; Clarke, D.J.B.; Evangelista, J.E.; Jenkins, S.L.; Lachmann, A.; Wojciechowicz, M.L.; Kropiwnicki, E.; Jagodnik, K.M.; et al. Gene set knowledge discovery with enrichr. Curr. Protoc. 2021, 1, e90. [Google Scholar] [CrossRef]
- Han, S.K.; Lee, D.; Lee, H.; Kim, D.; Son, H.G.; Yang, J.S.; Lee, S.V.; Kim, S. Oasis 2: Online application for survival analysis 2 with features for the analysis of maximal lifespan and healthspan in aging research. Oncotarget 2016, 7, 56147–56152. [Google Scholar] [CrossRef] [Green Version]
- Jasper, H. Intestinal stem cell aging: Origins and interventions. Annu. Rev. Physiol. 2020, 82, 203–226. [Google Scholar] [CrossRef] [Green Version]
- Zhang, P.; Edgar, B.A. Insect gut regeneration. Cold Spring Harb. Perspect. Biol. 2021, a040915. [Google Scholar] [CrossRef] [PubMed]
- Kou, J.; Kovacs, G.G.; Hoftberger, R.; Kulik, W.; Brodde, A.; Forss-Petter, S.; Honigschnabl, S.; Gleiss, A.; Brugger, B.; Wanders, R.; et al. Peroxisomal alterations in alzheimer’s disease. Acta Neuropathol. 2011, 122, 271–283. [Google Scholar] [CrossRef] [Green Version]
- Jo, D.S.; Park, N.Y.; Cho, D.H. Peroxisome quality control and dysregulated lipid metabolism in neurodegenerative diseases. Exp. Mol. Med. 2020, 52, 1486–1495. [Google Scholar] [CrossRef]
- Oddo, S. The role of mtor signaling in alzheimer disease. Front. Biosci. 2012, 4, 941–952. [Google Scholar] [CrossRef] [Green Version]
- Cai, Z.; Chen, G.; He, W.; Xiao, M.; Yan, L.J. Activation of mtor: A culprit of alzheimer’s disease? Neuropsychiatr. Dis. Treat. 2015, 11, 1015–1030. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hong, S.Y.; Ng, L.T.; Ng, L.F.; Inoue, T.; Tolwinski, N.S.; Hagen, T.; Gruber, J. The role of mitochondrial non-enzymatic protein acylation in ageing. PLoS ONE 2016, 11, e0168752. [Google Scholar] [CrossRef] [PubMed]
- Bonnay, F.; Veloso, A.; Steinmann, V.; Kocher, T.; Abdusselamoglu, M.D.; Bajaj, S.; Rivelles, E.; Landskron, L.; Esterbauer, H.; Zinzen, R.P.; et al. Oxidative metabolism drives immortalization of neural stem cells during tumorigenesis. Cell 2020, 182, 1490–1507.e1419. [Google Scholar] [CrossRef]
- Tiefenbock, S.K.; Baltzer, C.; Egli, N.A.; Frei, C. The drosophila pgc-1 homologue spargel coordinates mitochondrial activity to insulin signalling. EMBO J. 2010, 29, 171–183. [Google Scholar] [CrossRef] [Green Version]
- Mukherjee, S.; Duttaroy, A. Spargel/dpgc-1 is a new downstream effector in the insulin-tor signaling pathway in drosophila. Genetics 2013, 195, 433–441. [Google Scholar] [CrossRef] [Green Version]
- Ligoxygakis, P.; Pelte, N.; Hoffmann, J.A.; Reichhart, J.M. Activation of drosophila toll during fungal infection by a blood serine protease. Science 2002, 297, 114–116. [Google Scholar] [CrossRef] [PubMed]
- Kinney, J.W.; Bemiller, S.M.; Murtishaw, A.S.; Leisgang, A.M.; Salazar, A.M.; Lamb, B.T. Inflammation as a central mechanism in alzheimer’s disease. Alzheimer’s Dement. Transl. Res. Clin. Interv. 2018, 4, 575–590. [Google Scholar] [CrossRef] [PubMed]
- Issa, N.; Guillaumot, N.; Lauret, E.; Matt, N.; Schaeffer-Reiss, C.; Van Dorsselaer, A.; Reichhart, J.-M.; Veillard, F. The circulating protease persephone is an immune sensor for microbial proteolytic activities upstream of the drosophila toll pathway. Mol. Cell 2018, 69, 539–550.e536. [Google Scholar] [CrossRef] [Green Version]
- Ip, Y.T.; Reach, M.; Engstrom, Y.; Kadalayil, L.; Cai, H.; González-Crespo, S.; Tatei, K.; Levine, M. Dif, a dorsal-related gene that mediates an immune response in drosophila. Cell 1993, 75, 753–763. [Google Scholar] [CrossRef]
- Palomer, E.; Buechler, J.; Salinas, P.C. Wnt signaling deregulation in the aging and alzheimer’s brain. Front. Cell. Neurosci. 2019, 13, 227. [Google Scholar] [CrossRef]
- Sofola-Adesakin, O.; Castillo-Quan, J.I.; Rallis, C.; Tain, L.S.; Bjedov, I.; Rogers, I.; Li, L.; Martinez, P.; Khericha, M.; Cabecinha, M.; et al. Lithium suppresses abeta pathology by inhibiting translation in an adult drosophila model of alzheimer’s disease. Front. Aging Neurosci. 2014, 6, 190. [Google Scholar] [CrossRef]
- Teo, E.; Fong, S.; Tolwinski, N.; Gruber, J. Drug synergy as a strategy for compression of morbidity in a caenorhabditis elegans model of alzheimer’s disease. Geroscience 2020, 42, 849–856. [Google Scholar] [CrossRef]
- Castillo-Quan, J.I.; Tain, L.S.; Kinghorn, K.J.; Li, L.; Gronke, S.; Hinze, Y.; Blackwell, T.K.; Bjedov, I.; Partridge, L. A triple drug combination targeting components of the nutrient-sensing network maximizes longevity. Proc. Natl. Acad. Sci. USA 2019, 116, 20817–20819. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rallis, A.; Navarro, J.A.; Rass, M.; Hu, A.; Birman, S.; Schneuwly, S.; Therond, P.P. Hedgehog signaling modulates glial proteostasis and lifespan. Cell Rep. 2020, 30, 2627–2643.e5. [Google Scholar] [CrossRef] [Green Version]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kaur, P.; Chua, E.H.Z.; Lim, W.K.; Liu, J.; Harmston, N.; Tolwinski, N.S. Wnt Signaling Rescues Amyloid Beta-Induced Gut Stem Cell Loss. Cells 2022, 11, 281. https://doi.org/10.3390/cells11020281
Kaur P, Chua EHZ, Lim WK, Liu J, Harmston N, Tolwinski NS. Wnt Signaling Rescues Amyloid Beta-Induced Gut Stem Cell Loss. Cells. 2022; 11(2):281. https://doi.org/10.3390/cells11020281
Chicago/Turabian StyleKaur, Prameet, Ellora Hui Zhen Chua, Wen Kin Lim, Jiarui Liu, Nathan Harmston, and Nicholas S. Tolwinski. 2022. "Wnt Signaling Rescues Amyloid Beta-Induced Gut Stem Cell Loss" Cells 11, no. 2: 281. https://doi.org/10.3390/cells11020281
APA StyleKaur, P., Chua, E. H. Z., Lim, W. K., Liu, J., Harmston, N., & Tolwinski, N. S. (2022). Wnt Signaling Rescues Amyloid Beta-Induced Gut Stem Cell Loss. Cells, 11(2), 281. https://doi.org/10.3390/cells11020281