
Genome-Wide Association Analysis Reveals Trait-Linked Markers for Grain Nutrient and Agronomic Traits in Diverse Set of Chickpea Germplasm

 , ,
, ,
Abstract
:
1. Introduction
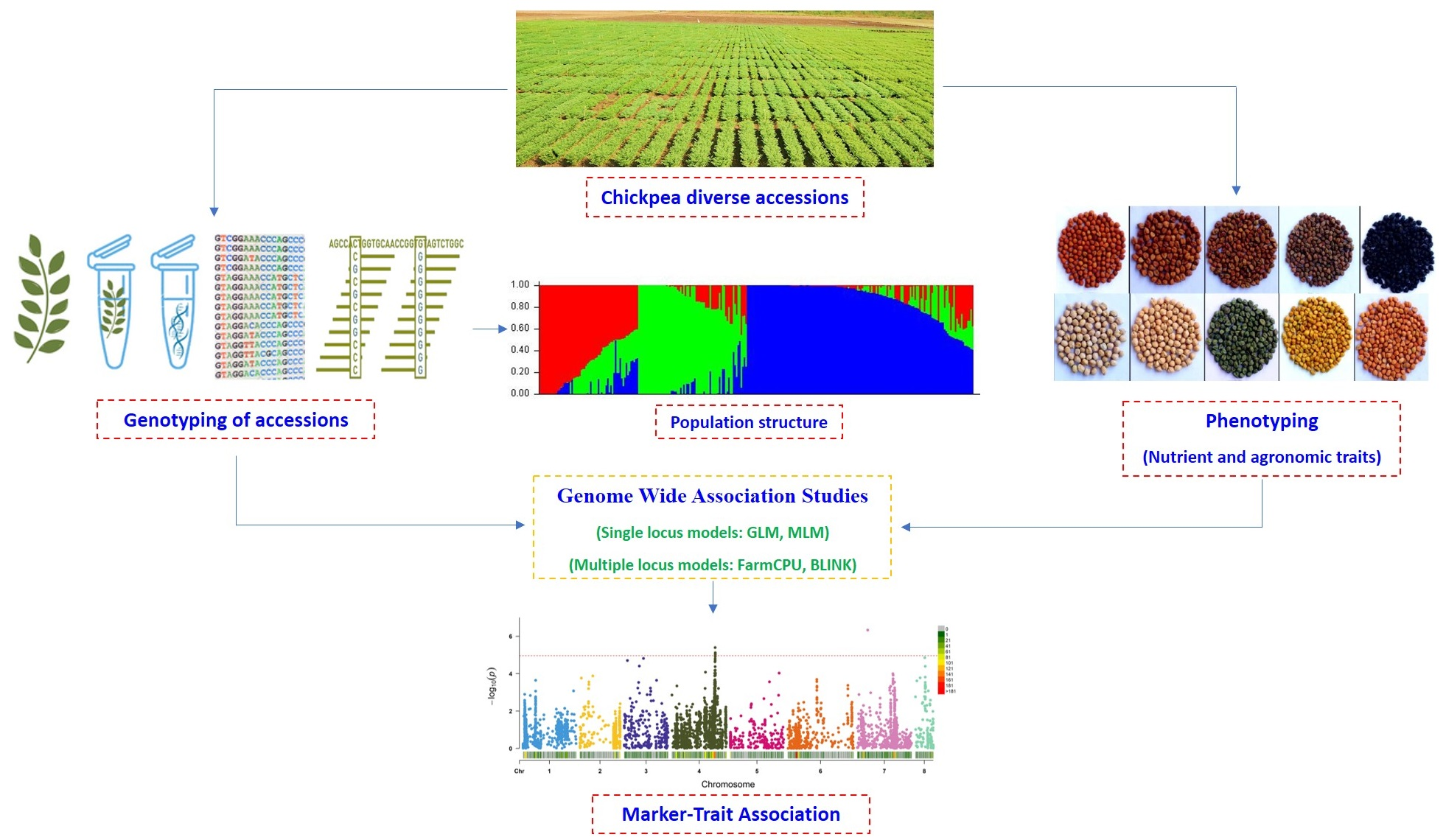
2. Materials and Methods
2.1. Plant Material
2.2. Estimation of Grain Protein, Fe and Zn
2.3. DNA Isolation and Quantification
2.4. SNP Genotyping and Filtering for GWAS
2.5. Analysis of Population Structure and Linkage Disequilibrium
2.6. Association Mapping–GWAS for Grain Nutrient and Agronomic Traits
2.7. In Silico SNP Annotation
3. Results
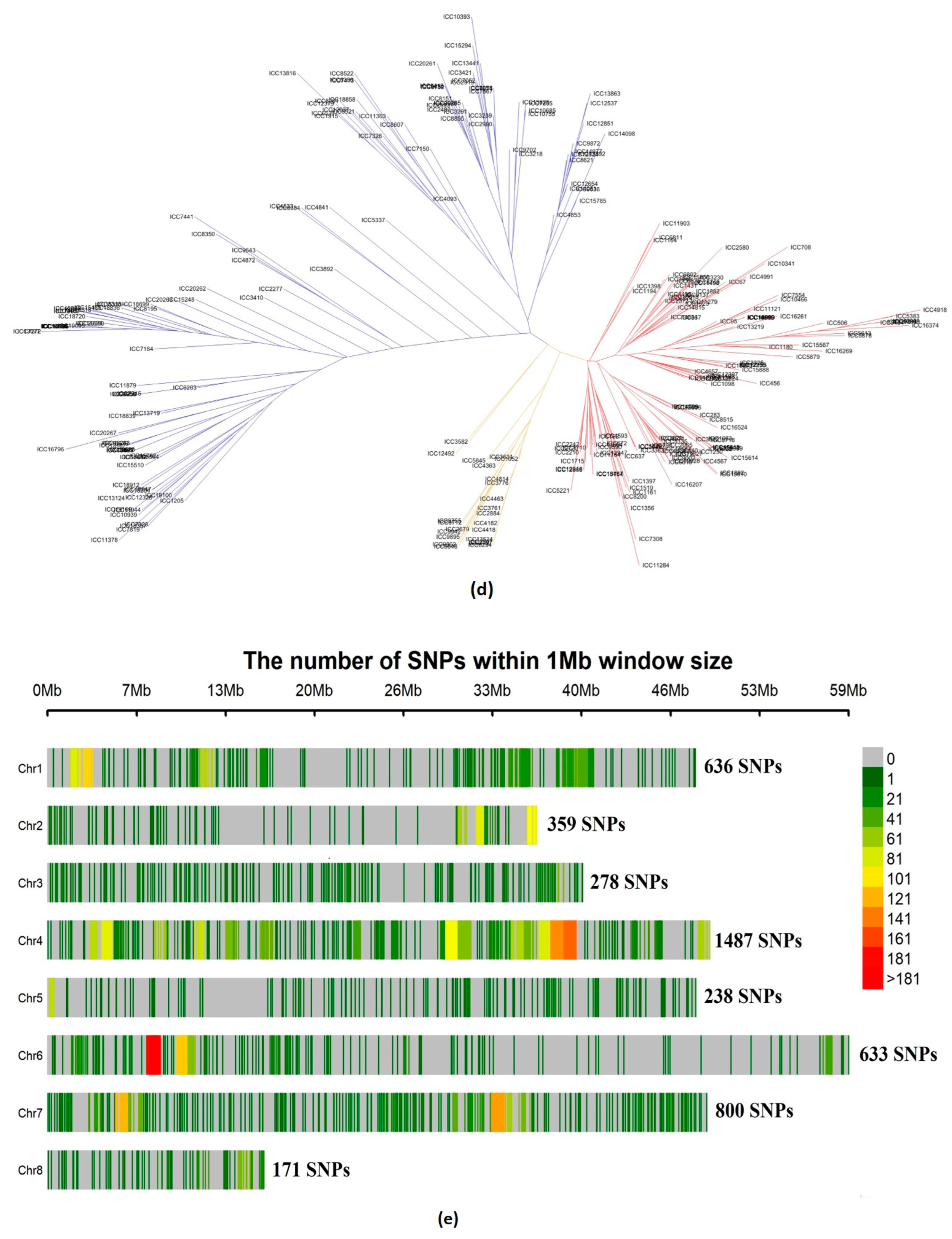
3.1. Characterization of the Population and the Genetic Relationships
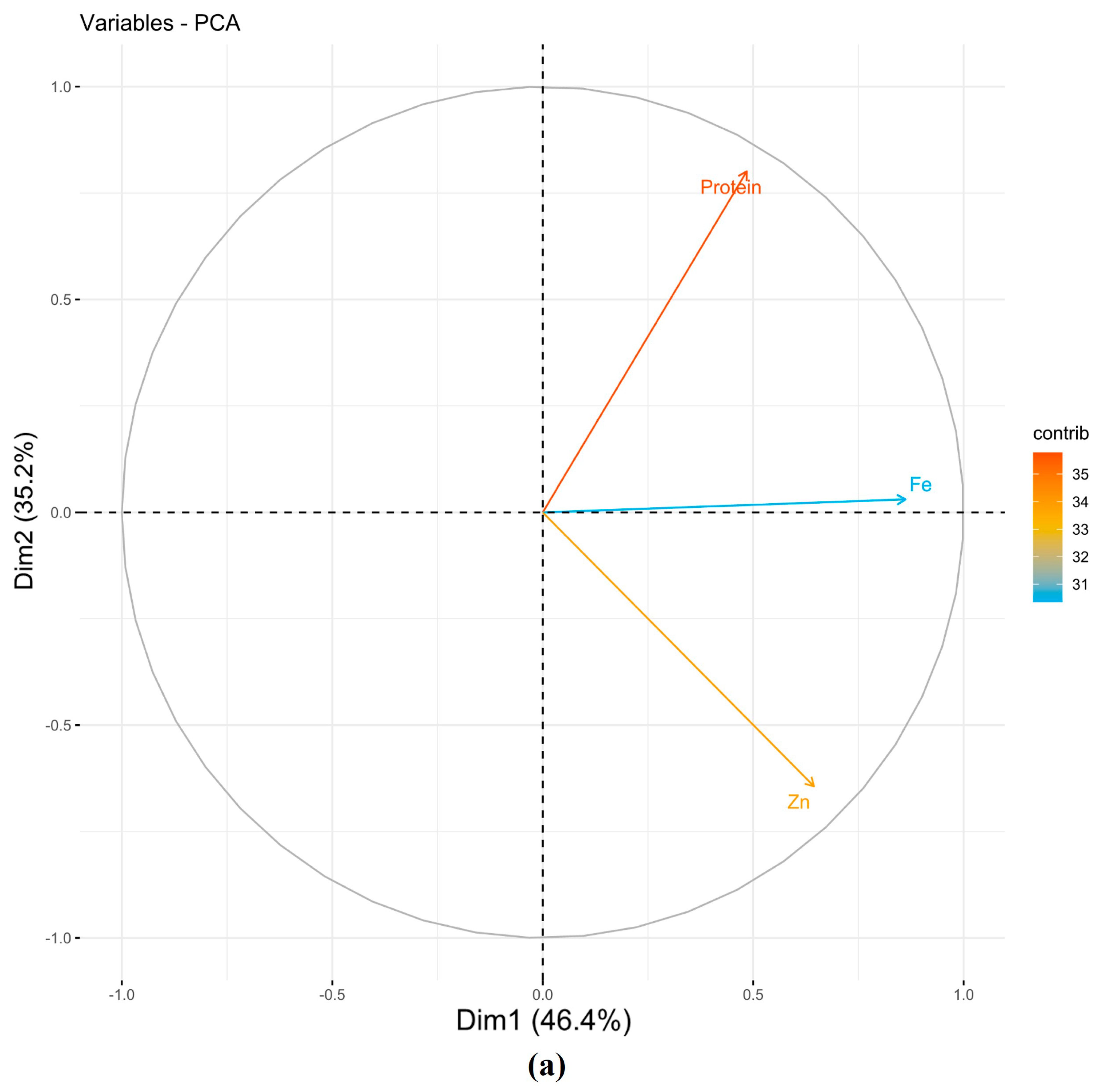
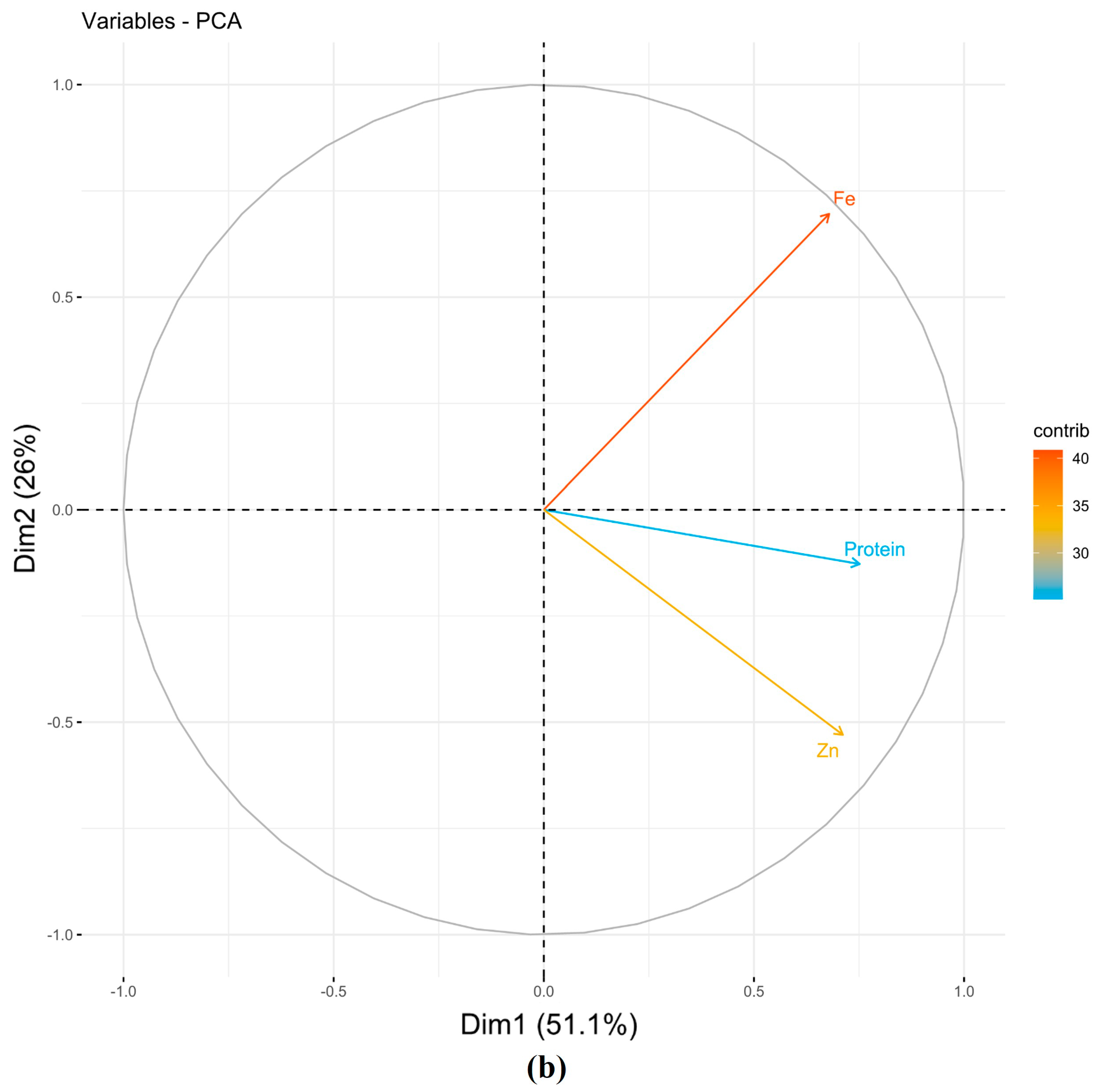
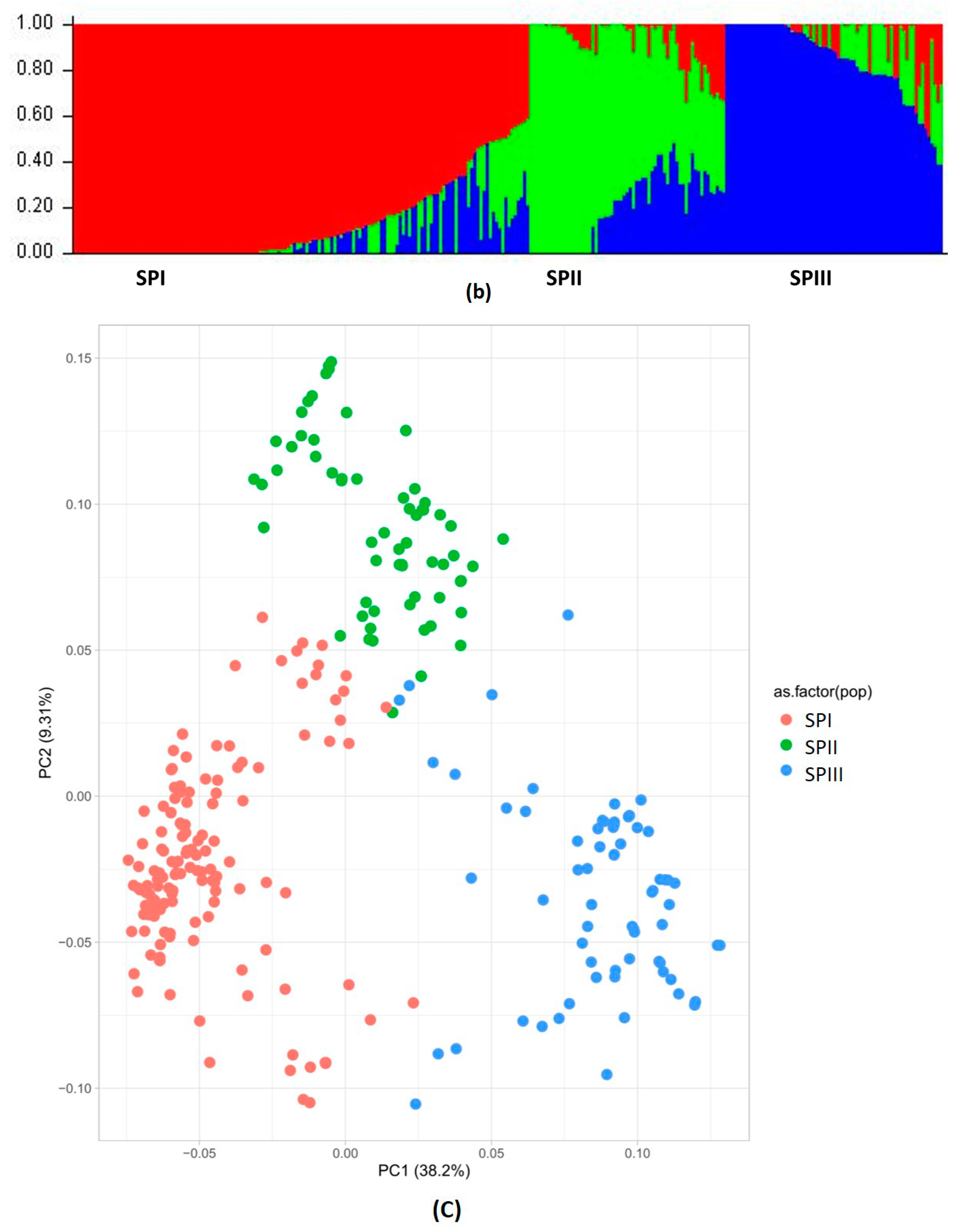
3.1.1. Principal Component Analysis (PCA)
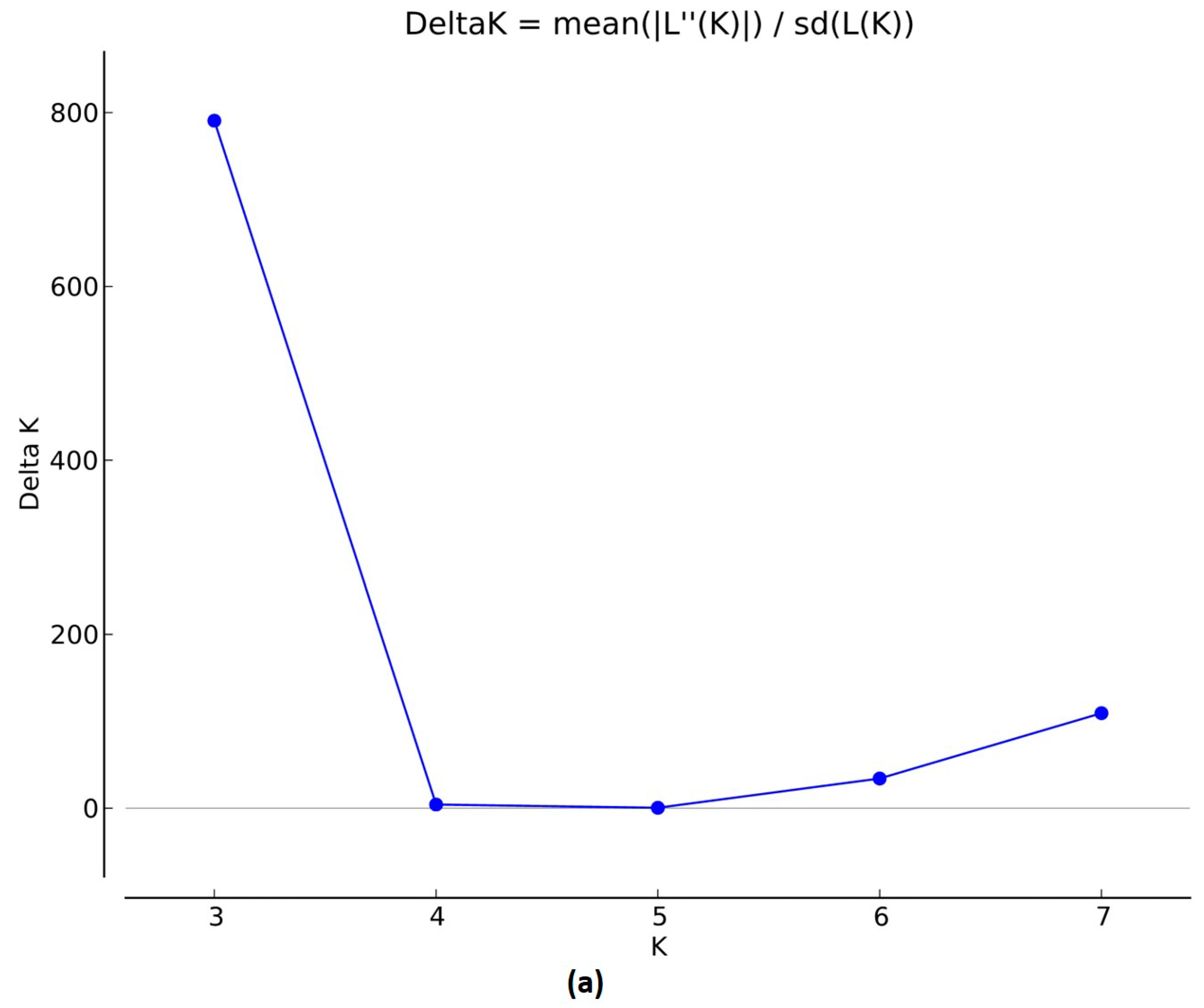
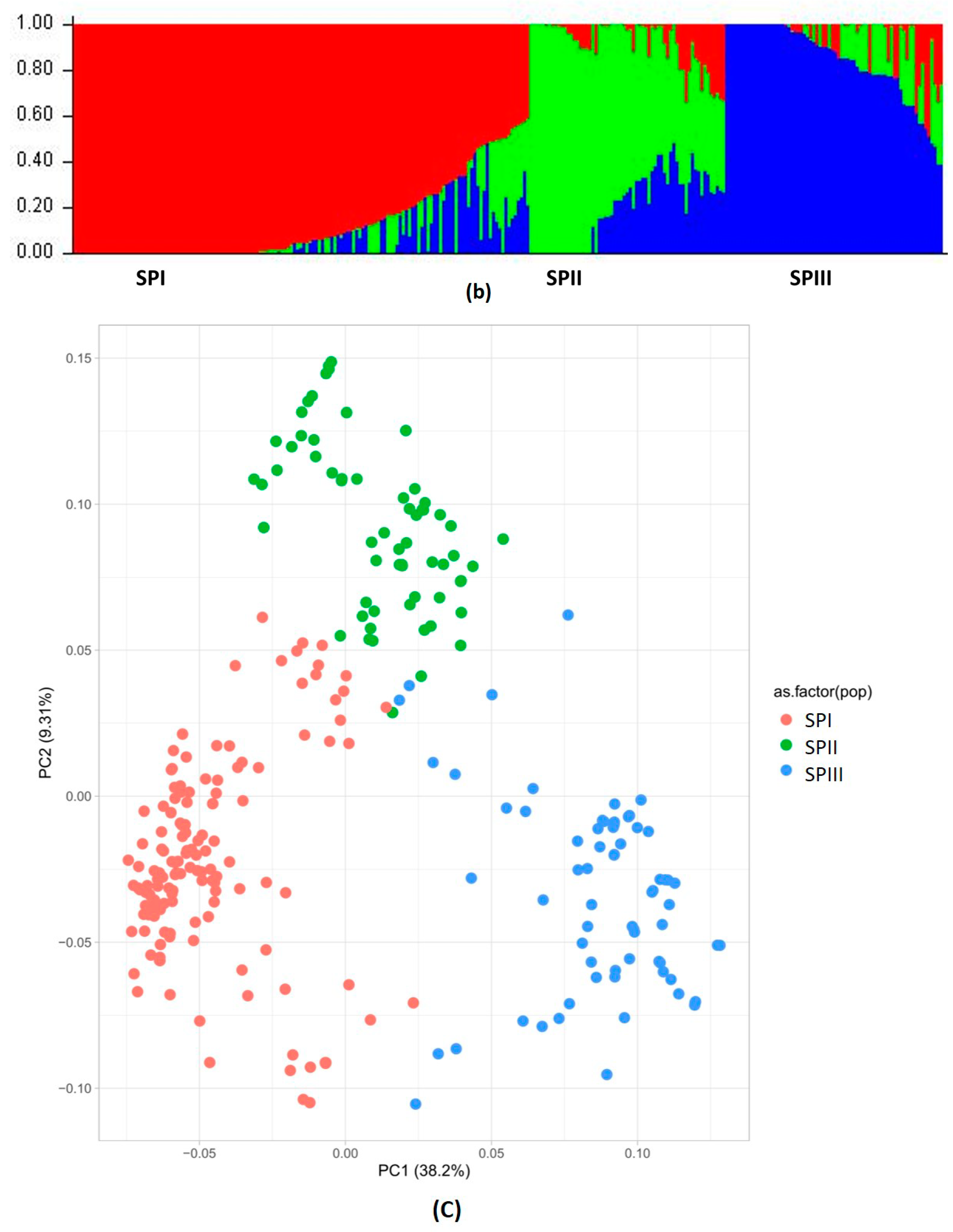
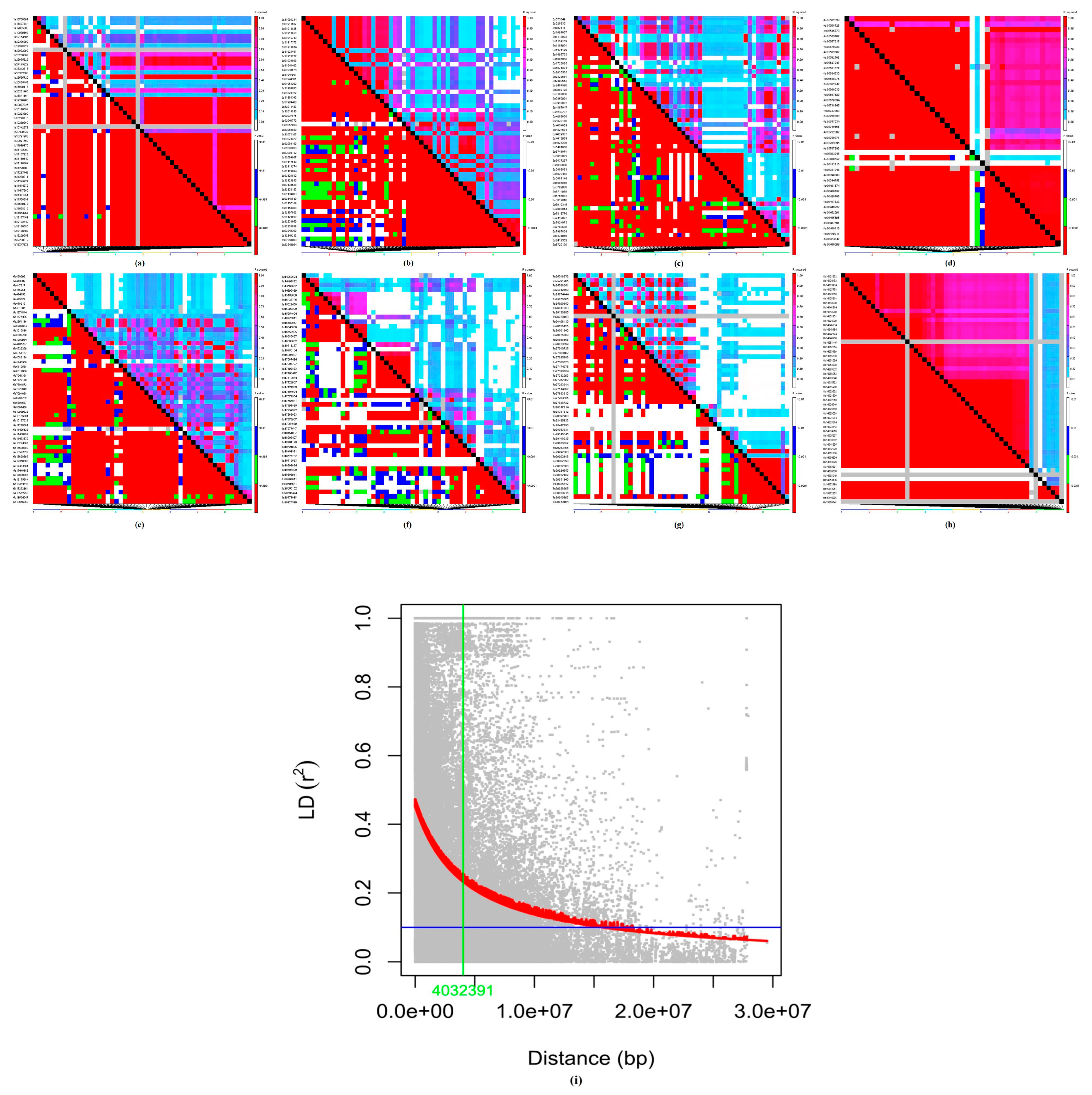
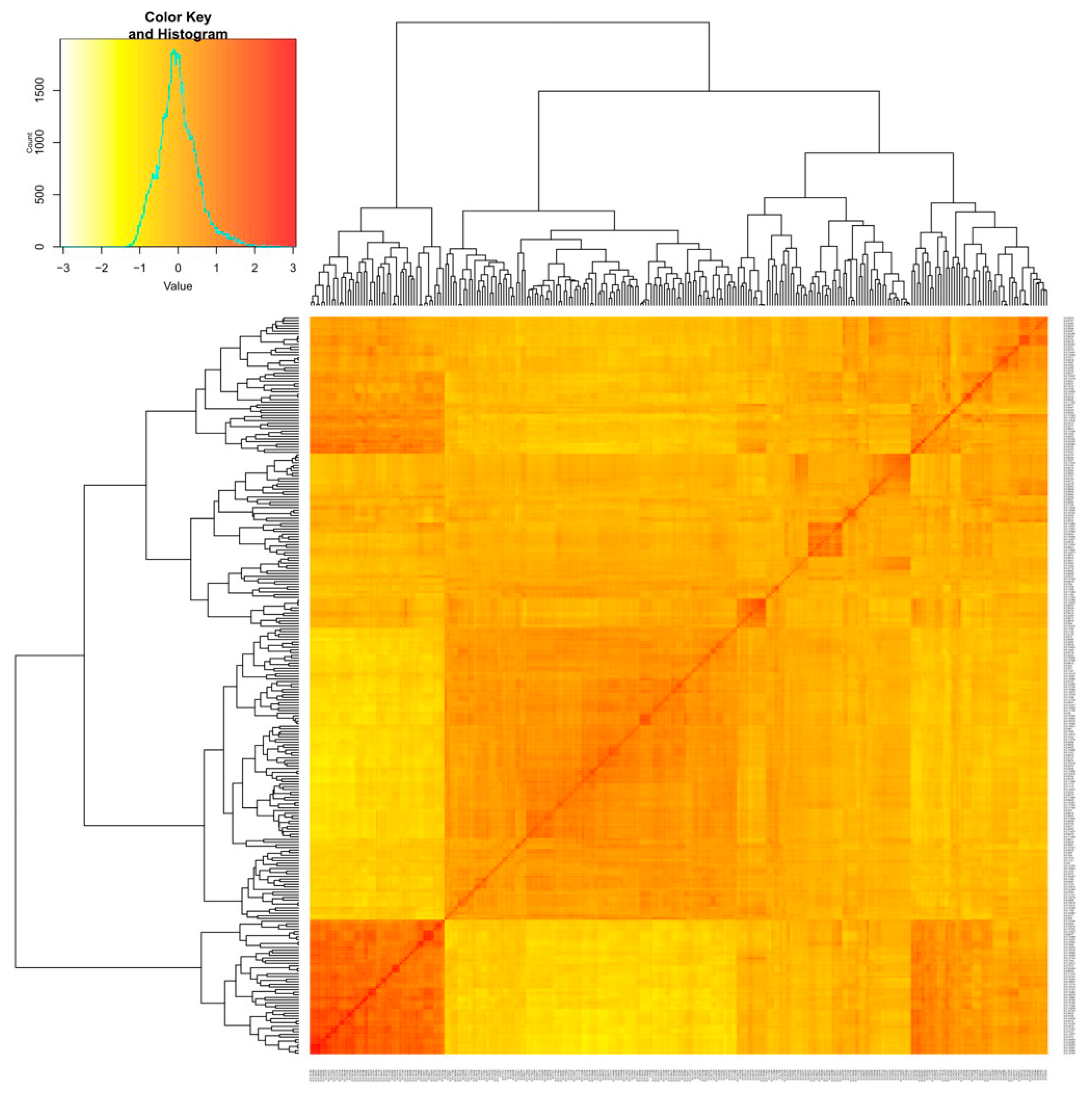
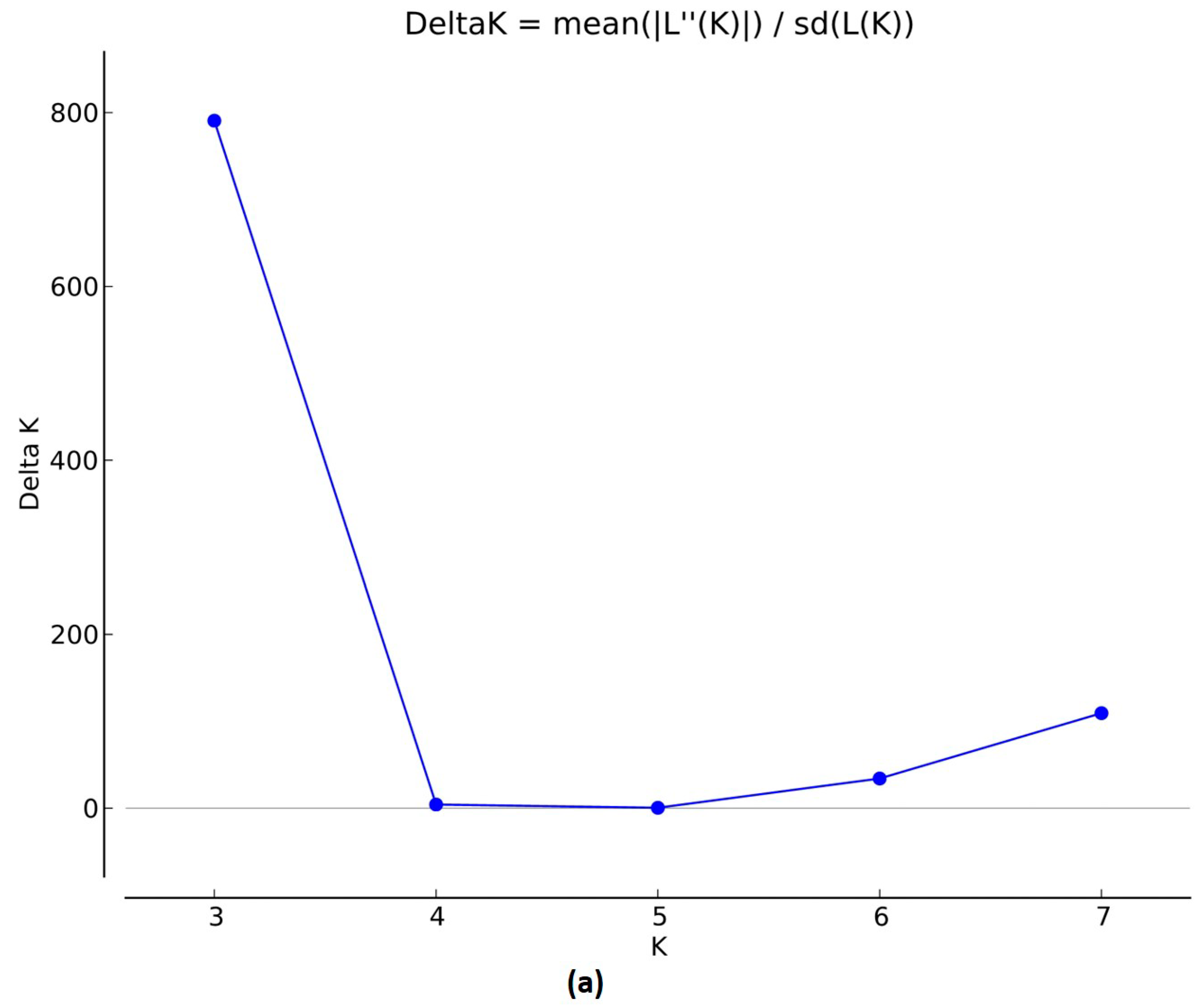
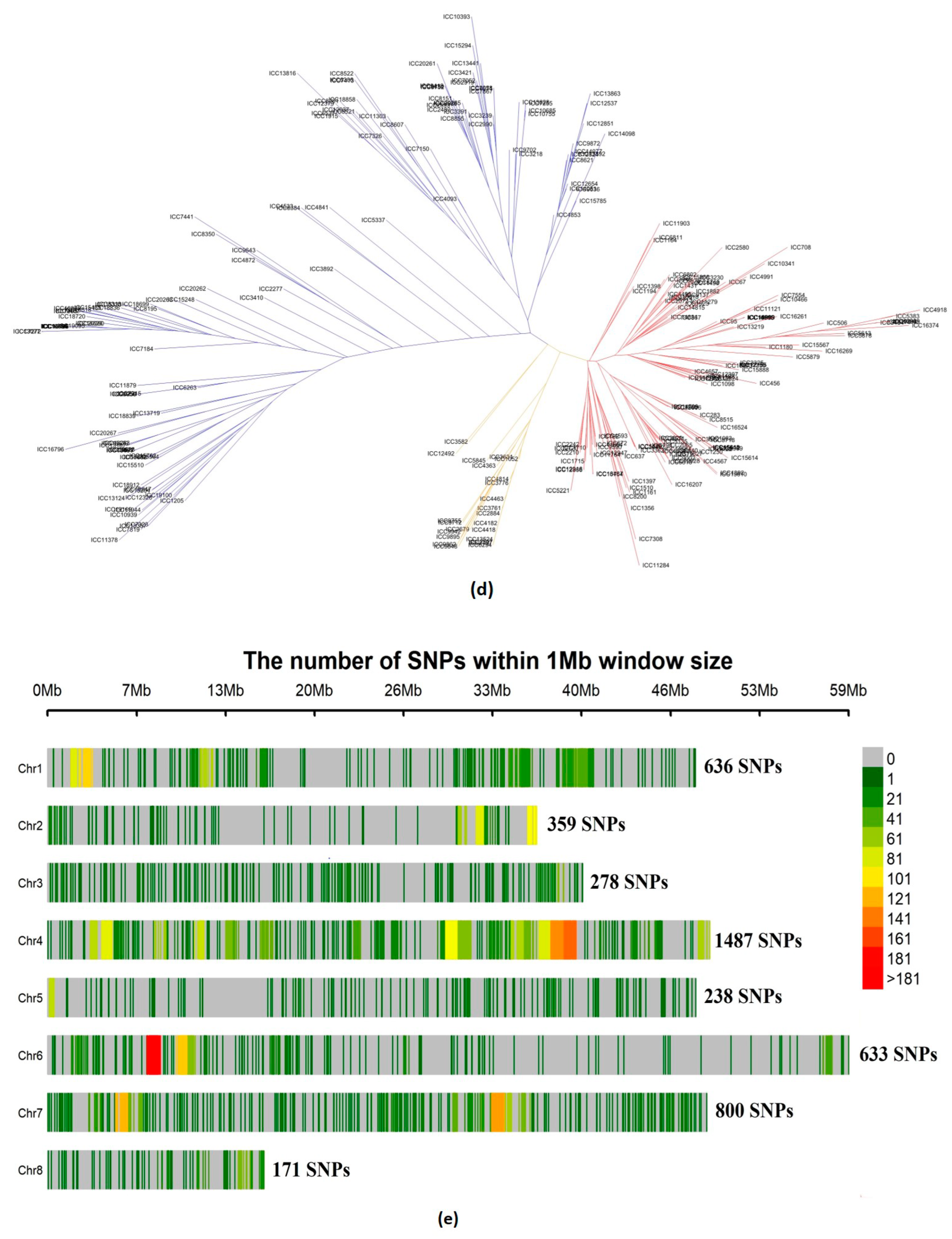
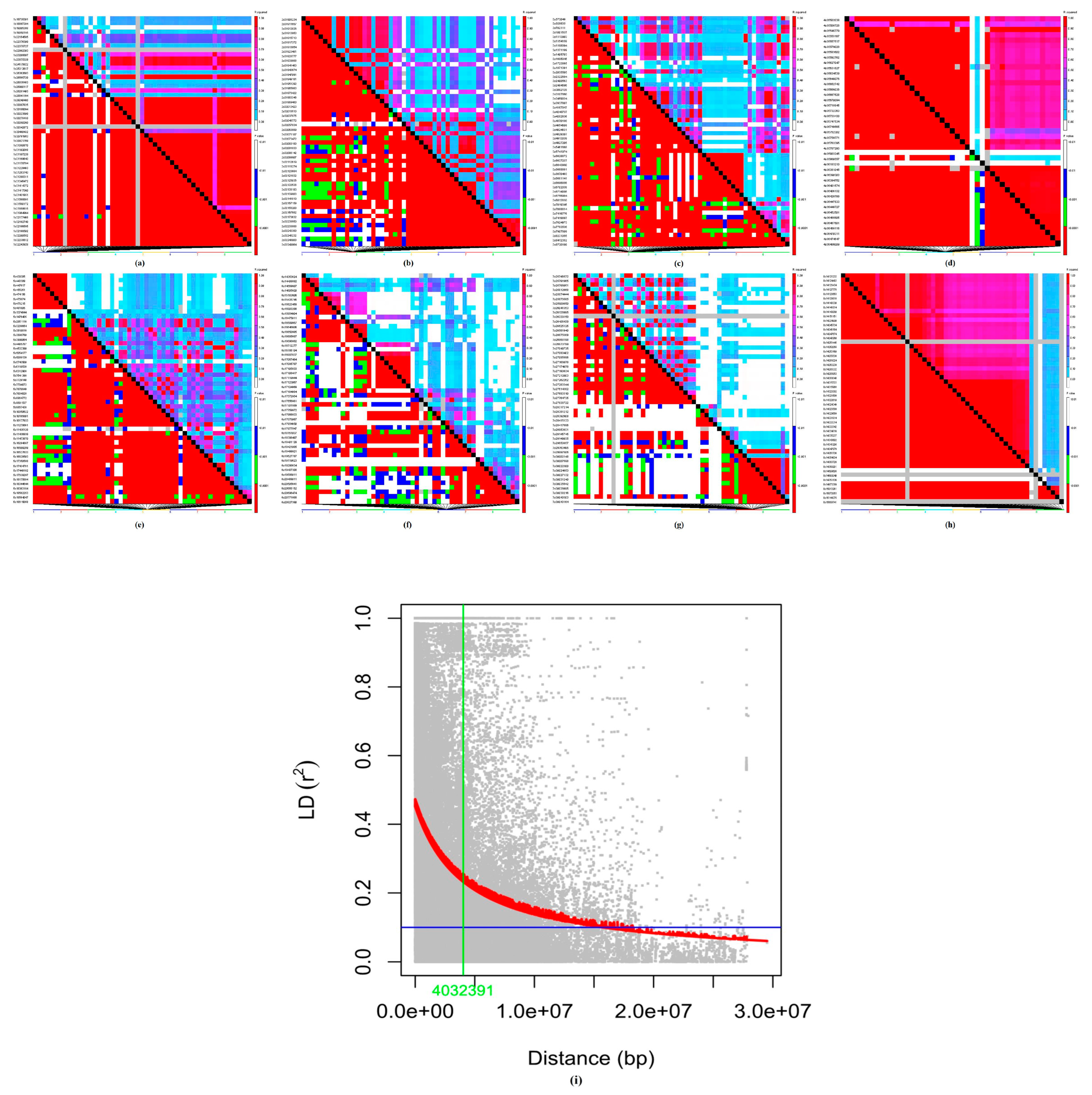
3.1.2. Population Structure, Kinship, and Linkage Disequilibrium
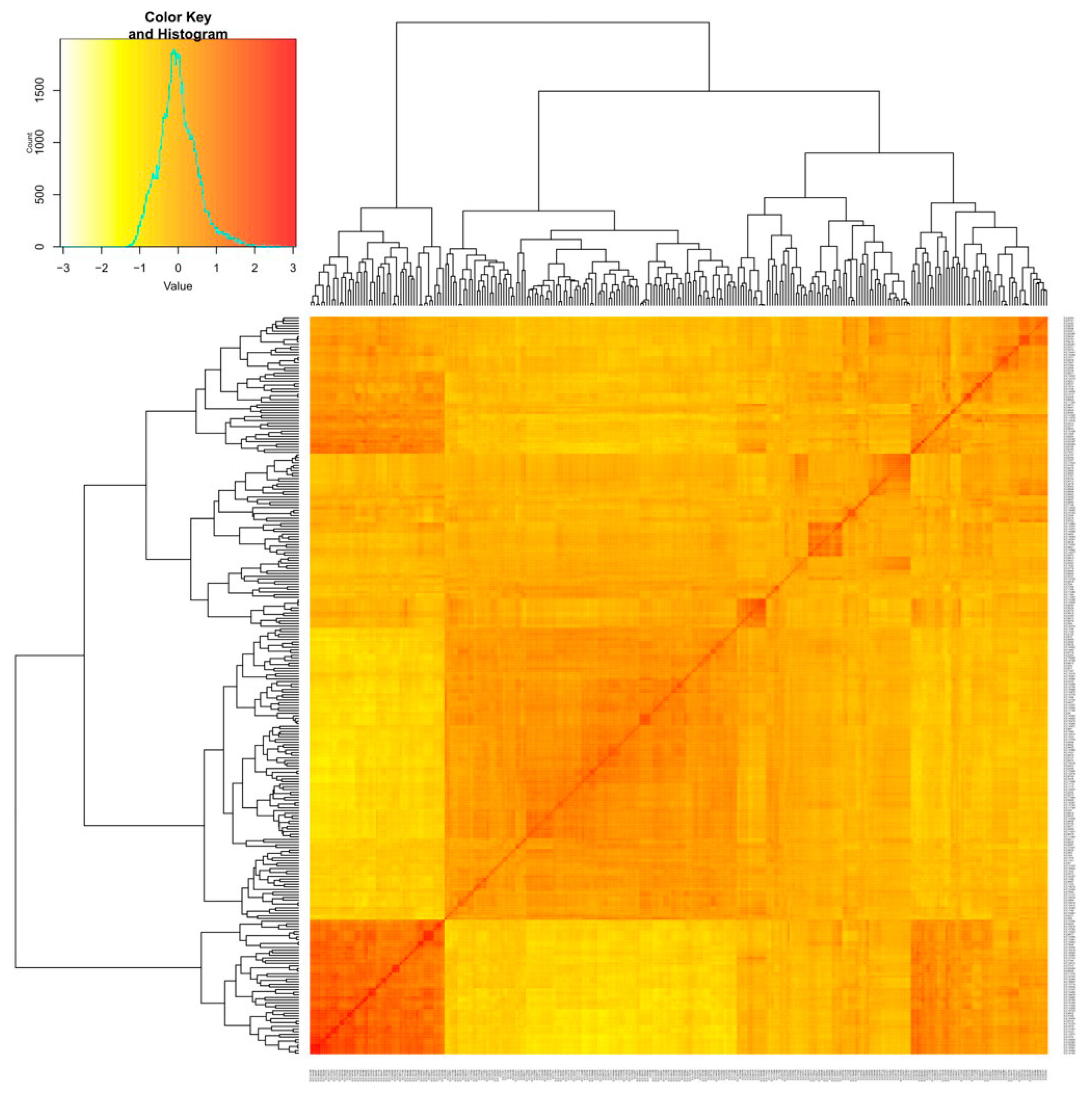
3.1.3. Relatedness between Chickpea Accessions
3.2. Genome-Wide Association Mapping for Grain Nutrient and Agronomic Traits
3.3. Annotation of Associated SNPs
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rapp, M.; Lein, V.; Lacoudre, F.; Lafferty, J.; Müller, E.; Vida, G.; Bozhanova, V.; Ibraliu, A.; Thorwarth, P.; Piepho, H.P.; et al. Simultaneous improvement of grain yield and protein content in durum wheat by different phenotypic indices and genomic selection. Theor. Appl. Genet. 2018, 131, 1315–1329. [Google Scholar] [CrossRef] [PubMed]
- Bailey, R.L.; West, J.K.P.; Black, R.E. The epidemiology of global micronutrient deficiencies. Ann. Nutr. Metab. 2015, 66, 22–33. [Google Scholar] [CrossRef] [PubMed]
- Crook, M.A. Zinc deficiency. Nutrition 2011, 27, 1085–1086. [Google Scholar] [CrossRef] [PubMed]
- Deshpande, J.D.; Joshi, M.M.; Giri, P.A. Zinc: The trace element of major importance in human nutrition and health. Int. J. Med. Sci. Public Health 2013, 2, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Palve, A.; Joshi, C.; Srivastava, R.K.; Rukhsar, R.K. Crop biofortifcation for iron (Fe), zinc (Zn) and vitamin A with transgenic approaches. Heliyon 2019, 5, e01914. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, A.D.; Ejeta, G. A new global agenda for nutrition and health: The importance of agriculture and food systems. Bull. World Health Organ. 2016, 94, 228–229. [Google Scholar] [CrossRef]
- Rehman, H.M.; Cooper, J.W.; Lam, H.M.; Yang, S.H. Legume biofortification is an underexploited strategy for combatting hidden hunger. Plant Cell Environ. 2019, 42, 52–70. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Pandey, G. Biofortification of pulses and legumes to enhance nutrition. Heliyon 2020, 6, 03682. [Google Scholar] [CrossRef]
- Jha, A.B.; Warkentin, T.D. Biofortification of pulse crops: Status and future perspectives. Plants 2020, 9, 73. [Google Scholar] [CrossRef] [Green Version]
- ICAR-Directorate of Knowledge Management in Agriculture, New Delhi. 2020. Available online: https://icar.org.in/content/pm-dedicates-17-biofortified-varieties-8-crops-nation-0 (accessed on 3 February 2022).
- Diapari, M.; Sindhu, A.; Bett, K.; Deokar, A.; Warkentin, T.D.; Tar’an, B. Genetic diversity and association mapping of iron and zinc concentrations in chickpea (Cicer arietinum L.). Genome 2014, 57, 459–468. [Google Scholar] [CrossRef] [Green Version]
- Jha, U.C.; Shil, S. Association analysis of yield contributing traits of chickpea genotypes under high temperature condition. Trends Biosci. 2015, 8, 2335–2341. [Google Scholar]
- Saha, S.; Chakraborty, D.; Sehgal, V.K.; Pal, M. Potential impact of rising atmospheric CO2 on quality of grains in chickpea (Cicer arietinum L.). Food Chem. 2015, 187, 431–436. [Google Scholar] [CrossRef]
- Gaur, P.M.; Singh, M.K.; Samineni, S.; Sajja, S.B.; Jukanti, A.K.; Kamatam, S.; Varshney, R.K. Inheritance of protein content and its relationships with seed size, grain yield and other traits in chickpea. Euphytica 2016, 209, 253–260. [Google Scholar] [CrossRef] [Green Version]
- Samineni, S.; Mahendrakar, M.D.; Hotti, A.; Chand, U.; Rathore, A.; Gaur, P.M. Impact of heat and drought stresses on grain nutrient content in chickpea: Genome-wide marker-trait associations for protein, Fe and Zn. Environ. Exp. Bot. 2022, 194, 104688. [Google Scholar] [CrossRef]
- Rajasekhar, S.; Mohammad, L.A.; Mahendrakar, M.D.; Chand, U.; Jagarlamudi, V.R.; Kondamudi, K.P.; Nandigam, S.R.; Vemula, A.; Samineni, S. Genetic Variation for Grain Protein, Fe and Zn Content traits in Chickpea Reference Set. J. Food Compos. Anal. 2022, 114, 104774. [Google Scholar] [CrossRef]
- Bajaj, D.; Srivastava, R.; Nath, M.; Tripathi, S.; Bharadwaj, C.; Upadhyaya, H.D.; Tyagi, A.K.; Parida, S.K. EcoTILLING-based association mapping efficiently delineates functionally relevant natural allelic variants of candidate genes governing agronomic traits in chickpea. Front. Plant Sci. 2016, 7, 450. [Google Scholar] [CrossRef] [Green Version]
- Upadhyaya, H.D.; Bajaj, D.; Das, S.; Saxena, M.S.; Badoni, S.; Kumar, V.; Tripathi, S.; Gowda, C.L.L.; Sharma, S.; Tyagi, A.K. A genome-scale integrated approach aids in genetic dissection of complex flowering time trait in chickpea. Plant Mol. Biol. 2015, 89, 403–420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Upadhyaya, H.D.; Bajaj, D.; Srivastava, R.; Daware, A.; Basu, U.; Tripathi, S.; Bharadwaj, C.; Tyagi, A.K.; Parida, S.K. Genetic dissection of plant growth habit in chickpea. Funct. Integr. Genom. 2017, 17, 711–723. [Google Scholar] [CrossRef] [Green Version]
- Thudi, M.; Upadhyaya, H.D.; Rathore, A.; Gaur, P.M.; Krishnamurthy, L.; Roorkiwal, M.; Nayak, S.N.; Chaturvedi, S.K.; Basu, P.S.; Gangarao, N.V.; et al. Genetic dissection of drought and heat tolerance in chickpea through genome-wide and candidate gene-based association mapping approaches. PLoS ONE 2014, 9, e96758. [Google Scholar] [CrossRef] [Green Version]
- Jha, U.C.; Jha, R.; Thakro, V.; Kumar, A.; Gupta, S.; Nayyar, H.; Basu, P.; Parida, S.K.; Singh, N.P. Discerning molecular diversity and association mapping for phenological, physiological and yield traits under high temperature stress in chickpea (Cicer arietinum L.). J. Genet. 2021, 100, 4. [Google Scholar] [CrossRef]
- Kujur, A.; Bajaj, D.; Upadhyaya, H.D.; Das, S.; Ranjan, R.; Shree, T.; Saxena, M.S.; Badoni, S.; Kumar, V.; Tripathi, S.; et al. Employing genome-wide SNP discovery and genotyping strategy to extrapolate the natural allelic diversity and domestication patterns in chickpea. Front. Plant Sci. 2015, 6, 162. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Ruperao, P.; Batley, J.; Edwards, D.; Davidson, J.; Hobson, K.; Sutton, T. Genome analysis identified novel candidate genes for ascochyta blight resistance in chickpea using whole genome re-sequencing data. Front. Plant Sci. 2017, 8, 359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khazaei, H.; Podder, R.; Caron, C.T.; Kundu, S.S.; Diapari, M.; Vandenberg, A.; Bett, K.E. Marker–trait association analysis of iron and zinc concentration in lentil (Lens culinaris Medik.) seeds. Plant Genome 2017, 10, plantgenome2017-02. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katuuramu, D.N.; Hart, J.P.; Porch, T.G.; Grusak, M.A.; Glahn, R.P.; Cichy, K.A. Genome-wide association analysis of nutritional composition-related traits and iron bioavailability in cooked dry beans (Phaseolus vulgaris L.). Mol. Breed. 2018, 38, 44. [Google Scholar] [CrossRef]
- Diapari, M.; Sindhu, A.; Warkentin, T.D.; Bett, K.; Tar’an, B. Population structure and marker-trait association studies of iron, zinc and selenium concentrations in seed of field pea (Pisum sativum L.). Mol. Breed. 2015, 35, 30. [Google Scholar] [CrossRef]
- Jadhav, A.A.; Rayate, S.J.; Mhase, L.B.; Thudi, M.; Chitikineni, A.; Harer, P.N.; Jadhav, A.S.; Varshney, R.K.; Kulwal, P.L. Marker-trait association study for protein content in chickpea (Cicer arietinum L.). J. Genet. 2015, 94, 279–286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Upadhyaya, H.D.; Bajaj, D.; Das, S.; Kumar, V.; Gowda, C.L.L.; Sharma, S.; Tyagi, A.K.; Parida, S.K. Genetic dissection of seed-iron and zinc concentrations in chickpea. Sci. Rep. 2016, 6, 24050. [Google Scholar] [CrossRef] [Green Version]
- Upadhyaya, H.D.; Bajaj, D.; Narnoliya, L.; Das, S.; Kumar, V.; Gowda, C.L.L.; Sharma, S.; Tyagi, A.K.; Parida, S.K. Genome-wide scans for delineation of candidate genes regulating seed-protein content in chickpea. Front. Plant Sci. 2016, 7, 302. [Google Scholar] [CrossRef] [Green Version]
- Sab, S.; Lokesha, R.; Mannur, D.M. Genome-Wide SNP Discovery and Mapping QTLs for Seed Iron and Zinc Concentrations in Chickpea (Cicer arietinum L.). Front. Nutr. 2020, 7, 559120. [Google Scholar] [CrossRef]
- Roorkiwal, M.; Bharadwaj, C.; Barmukh, R.; Dixit, G.P.; Thudi, M.; Gaur, P.M.; Chaturvedi, S.K.; Fikre, A.; Hamwieh, A.; Kumar, S.; et al. Integrating genomics for chickpea improvement: Achievements and opportunities. Theor. Appl. Genet. 2020, 133, 1703–1720. [Google Scholar] [CrossRef] [Green Version]
- Pritchard, J.K.; Stephens, M.; Donnelly, P. Inference of population structure using multilocus genotype data. Genetics 2000, 155, 945–959. [Google Scholar] [CrossRef] [PubMed]
- Weir, B.S.; Cockerham, C.C. Estimating F-statistics for the analysis of population structure. Evolution 1984, 38, 1358. [Google Scholar] [PubMed]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the number of clusters of individuals using the software structure: A simulation study. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perrier, X.; Flori, A. Methods of data analysis. In Genetic Diversity of Cultivated Tropical Plants; CRC Press: Boca Raton, FL, USA, 2003; pp. 47–80. [Google Scholar]
- Lipka, A.E.; Tian, F.; Wang, Q.; Peiffer, J.; Li, M.; Bradbury, P.J.; Gore, M.A.; Buckler, E.S.; Zhang, Z. GAPIT: Genome association and prediction integrated tool. Bioinformatics 2012, 28, 2397–2399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, X.; Huang, M.; Fan, B.; Buckler, E.S.; Zhang, Z. Iterative Usage of Fixed and Random Effect Models for Powerful and Efficient Genome-Wide Association Studies. PLoS Genet. 2016, 12, e1005767. [Google Scholar] [CrossRef] [PubMed]
- Huang, M.; Liu, X.; Zhou, Y.; Summers, R.M.; Zhang, Z. BLINK: A package for the next level of genome-wide association studies with both individuals and markers in the millions. Gigascience 2019, 8, giy154. [Google Scholar] [CrossRef]
- Suwarno, W.B.; Pixley, K.V.; Palacios-Rojas, N.; Kaeppler, S.M.; Babu, R. Genome-wide association analysis reveals new targets for carotenoid biofortification in maize. Theor. Appl. Genet. 2015, 128, 851–864. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gowda, M.; Das, B.; Makumbi, D.; Babu, R.; Semagn, K.; Mahuku, G.; Olsen, M.S.; Bright, J.M.; Beyene, Y.; Prasanna, B.M. Genome-wide association and genomic prediction of resistance to maize lethal necrosis disease in tropical maize germplasm. Theor. Appl. Genet. 2015, 128, 1957–1968. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Longmei, N.; Gill, G.K.; Zaidi, P.H.; Kumar, R.; Nair, S.K.; Hindu, V.; Vinayan, M.T.; Vikal, Y. Genome wide association mapping for heat tolerance in sub-tropical maize. BMC Genom. 2021, 22, 154. [Google Scholar] [CrossRef]
- Varshney, R.K.; Song, C.; Saxena, R.K.; Azam, S.; Yu, S.; Sharpe, A.G.; Cannon, S.; Baek, J.; Rosen, B.D.; Tar’an, B.; et al. Draft genome sequence of chickpea (Cicer arietinum) provides a resource for trait improvement. Nat. Biotechnol. 2013, 31, 240–246. [Google Scholar] [CrossRef] [Green Version]
- Deokar, A.A.; Ramsay, L.; Sharpe, A.G.; Diapari, M.; Sindhu, A.; Bett, K.; Warkentin, T.D.; Tar’an, B. Genome wide SNP identification in chickpea for use in development of a high density genetic map and improvement of chickpea reference genome assembly. BMC Genom. 2014, 15, 708. [Google Scholar] [CrossRef] [Green Version]
- Basu, U.; Srivastava, R.; Bajaj, D.; Thakro, V.; Daware, A.; Malik, N.; Upadhyaya, H.D.; Parida, S.K. Genome-wide generation and genotyping of informative SNPs to scan molecular signatures for seed yield in chickpea. Sci. Rep. 2018, 8, 13240. [Google Scholar] [CrossRef] [PubMed]
- Roorkiwal, M.; Jain, A.; Kale, S.M.; Doddamani, D.; Chitikineni, A.; Thudi, M.; Varshney, R.K. Development and evaluation of high-density Axiom® Cicer SNP Array for high-resolution genetic mapping and breeding applications in chickpea. Plant Biotechnol. J. 2018, 16, 890–901. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sorkheh, K.; Malysheva-Otto, L.V.; Wirthensohn, M.G.; Tarkesh-Esfahani, S.; Martínez-Gómez, P. Linkage disequilibrium, genetic association mapping and gene localization in crop plants. Genet. Mol. Biol. 2008, 31, 805–814. [Google Scholar] [CrossRef]
- Gali, K.K.; Sackville, A.; Tafesse, E.G.; Lachagari, V.; McPhee, K.; Hybl, M.; Miki´c, A.; Smýkal, P.; McGee, R.; Burstin, J.; et al. Genome-wide association mapping for agronomic and seed quality traits of field pea (Pisum sativum L.). Front. Plant Sci. 2019, 10, 1538. [Google Scholar] [CrossRef] [PubMed]
- Sokolkova, A.; Bulyntsev, S.V.; Chang, P.L.; Carrasquilla-Garcia, N.; Igolkina, A.A.; Noujdina, N.V.; von Wettberg, E.; Vishnyakova, M.A.; Cook, D.R.; Nuzhdin, S.V.; et al. Genomic Analysis of Vavilov’s Historic Chickpea Landraces Reveals Footprints of Environmental and Human Selection. Int. J. Mol. Sci. 2020, 21, 3952. [Google Scholar] [CrossRef]
- Huang, X.; Han, B. Natural variations and genome-wide association studies in crop plants. Annu. Rev. Plant Biol. 2014, 65, 531–551. [Google Scholar] [CrossRef]
- Niu, S.; Song, Q.; Koiwa, H.; Qiao, D.; Zhao, D.; Chen, Z.; Liu, X.; Wen, X. Genetic diversity, linkage disequilibrium, and population structure analysis of the tea plant (Camellia sinensis) from an origin center, Guizhou plateau, using genome-wide SNPs developed by genotyping-by-sequencing. BMC Plant Biol. 2019, 19, 328. [Google Scholar] [CrossRef]
- Basu, U.; Narnoliya, L.; Srivastava, R.; Sharma, A.; Bajaj, D.; Daware, A.; Thakro, V.; Malik, N.; Upadhyaya, H.D.; Tripathi, S.; et al. CLAVATA signaling pathway genes modulating flowering time and flower number in chickpea. Theor. Appl. Genet. 2019, 132, 2017–2038. [Google Scholar] [CrossRef]
- Sodedji, F.A.K.; Ryu, D.; Choi, J.; Agbahoungba, S.; Assogbadjo, A.E.; N’Guetta, S.P.A.; Jung, J.H.; Nho, C.W.; Kim, H.Y. Genetic Diversity and Association Analysis for Carotenoid Content among Sprouts of Cowpea (Vigna unguiculata L. Walp). Int. J. Mol. Sci. 2022, 23, 3696. [Google Scholar] [CrossRef]
- Valdisser, P.A.M.R.; Pereira, W.J.; Filho, J.E.A.; Müller, B.S.F.; Coelho, G.R.C.; De Menezes, I.P.P.; Vianna, J.P.G.; Zucchi, M.I.; Lanna, A.C.; Coelho, A.S.G.; et al. In-depth genome characterization of a Brazilian common bean core collection using DArTseq high-density SNP genotyping. BMC Genom. 2017, 18, 423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farahani, S.; Maleki, M.; Mehrabi, R.; Kanouni, H.; Scheben, A.; Batley, J.; Talebi, R. Whole genome diversity, population structure, and linkage disequilibrium analysis of chickpea (Cicer arietinum L.) genotypes using genome-wide DArTseq-based SNP markers. Genes 2019, 10, 676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saxena, M.S.; Bajaj, D.; Kujur, A.; Das, S.; Badoni, S.; Kumar, V.; Singh, M.; Bansal, K.C.; Tyagi, A.K.; Parida, S.K. Natural allelic diversity, genetic structure and linkage disequilibrium pattern in wild chickpea. PLoS ONE 2014, 9, e107484. [Google Scholar]
- Thudi, M.; Khan, A.W.; Kumar, V.; Gaur, P.M.; Katta, A.V.S.K.; Garg, V.; Roorkiwal, M.; Samineni, S.; Varshney, R.K. Whole genome resequencing reveals genome wide variations among parental lines of mapping populations in chickpea (Cicer arietinum). BMC Plant Biol. 2016, 16, 10. [Google Scholar] [CrossRef] [Green Version]
- Upadhyaya, H.D.; Dwivedi, S.L.; Baum, M.; Varshney, R.K.; Udupa, S.M.; Gowda, C.L.; Hoisington, D.; Singh, S. Genetic structure, diversity, and allelic richness in composite collection and reference set in chickpea (Cicer arietinum L.). BMC Plant Biol. 2008, 8, 106. [Google Scholar] [CrossRef] [Green Version]
- De Giovanni, C.; Pavan, S.; Taranto, F.; Di Rienzo, V.; Miazzi, M.M.; Marcotrigiano, A.R.; Mangini, G.; Montemurro, C.; Ricciardi, L.; Lotti, C. Genetic variation of a global germplasm collection of chickpea (Cicer arietinum L.) including Italian accessions at risk of genetic erosion. Physiol. Mol. Biol. Plants 2017, 23, 197–205. [Google Scholar] [CrossRef]
- Hinrichs, A.L.; Larkin, E.K.; Suarez, B.K. Population stratification and patterns of linkage disequilibrium. Genet. Epidemiol. 2009, 33, S88–S92. [Google Scholar] [CrossRef] [Green Version]
- Wang, R.; Gangola, M.P.; Irvine, C.; Gaur, P.M.; Baga, M.; Chibbar, R.N. Co-localization of genomic regions associated with seed morphology and composition in a desi chickpea (Cicer arietinum L.) population varying in seed protein concentration. Theor. Appl. Genet. 2019, 132, 1263–1281. [Google Scholar] [CrossRef]
- Karaca, N.; Ates, D.; Nemli, S.; Ozkuru, E.; Yilmaz, H.; Yagmur, B.; Kartal, C.; Tosun, M.; Ozdestan, O.; Otles, S.; et al. Identification of SNP Markers Associated with Iron and Zinc Concentrations in Cicer Seeds. Curr. Genom. 2020, 21, 212–223. [Google Scholar] [CrossRef]
- Shimray, P.W.; Bajaj, D.; Srivastava, R.; Daware, A.; Upadhyaya, H.D.; Kumar, R.; Bharadwaj, C.; Tyagi, A.K.; Parida, S.K. Identifying transcription factor genes associated with yield traits in chickpea. Plant Mol. Biol. Rep. 2017, 35, 562–574. [Google Scholar] [CrossRef]
- Verma, S.; Gupta, S.; Bandhiwal, N.; Kumar, T.; Bharadwaj, C.; Bhatia, S. High-density linkage map construction and mapping of seed trait QTLs in chickpea (Cicer arietinum L.) using Genotyping-by-Sequencing (GBS). Sci. Rep. 2015, 5, 17512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barmukh, R.; Soren, K.R.; Madugula, P.; Gangwar, P.; Shanmugavadivel, P.S.; Bharadwaj, C.; Konda, A.K.; Chaturvedi, S.K.; Bhandari, A.; Rajain, K.; et al. Construction of a high-density genetic map and QTL analysis for yield, yield components and agronomic traits in chickpea (Cicer arietinum L.). PLoS ONE 2021, 16, e0251669. [Google Scholar] [CrossRef] [PubMed]
- Varshney, R.K.; Thudi, M.; Roorkiwal, M.; He, W.; Upadhyaya, H.D.; Yang, W.; Bajaj, P.; Cubry, P.; Rathore, A.; Jian, J.; et al. Resequencing of 429 chickpea accessions from 45 countries provides insights into genome diversity, domestication and agronomic traits. Nat. Genet. 2019, 51, 857–864. [Google Scholar] [CrossRef] [Green Version]
- Pradhan, S.; Kant, C.; Verma, S.; Bhatia, S. Genome-wide analysis of the CCCH zinc finger family identifies tissue specific and stress responsive candidates in chickpea (Cicer arietinum L.). PLoS ONE 2017, 12, e0180469. [Google Scholar] [CrossRef] [PubMed]
- Rajkumar, M.S.; Garg, R.; Jain, M. Genome-wide discovery of DNA polymorphisms among chickpea cultivars with contrasting seed size/weight and their functional relevance. Sci. Rep. 2018, 8, 16795. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
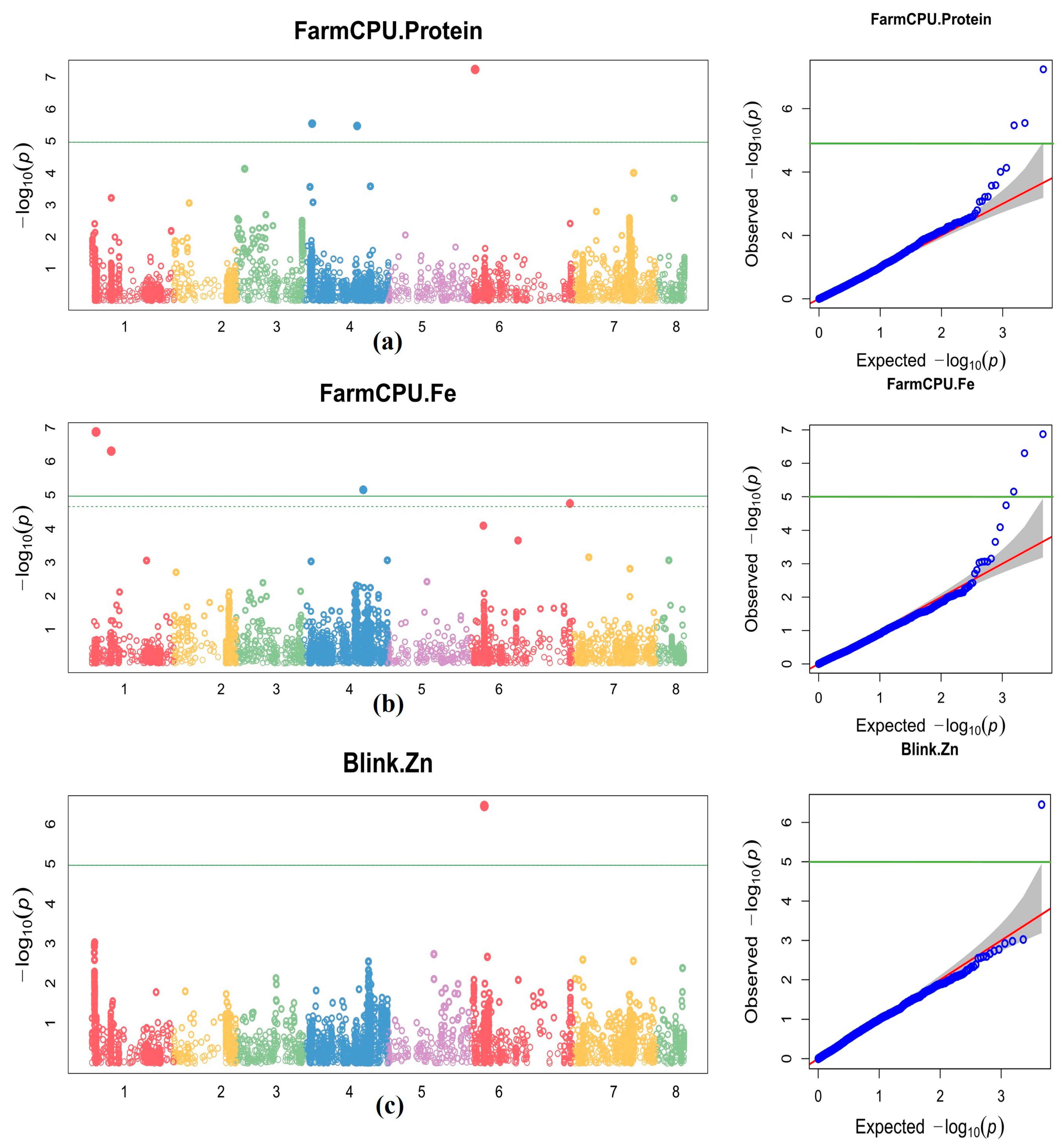{kind=link}
{kind=link}
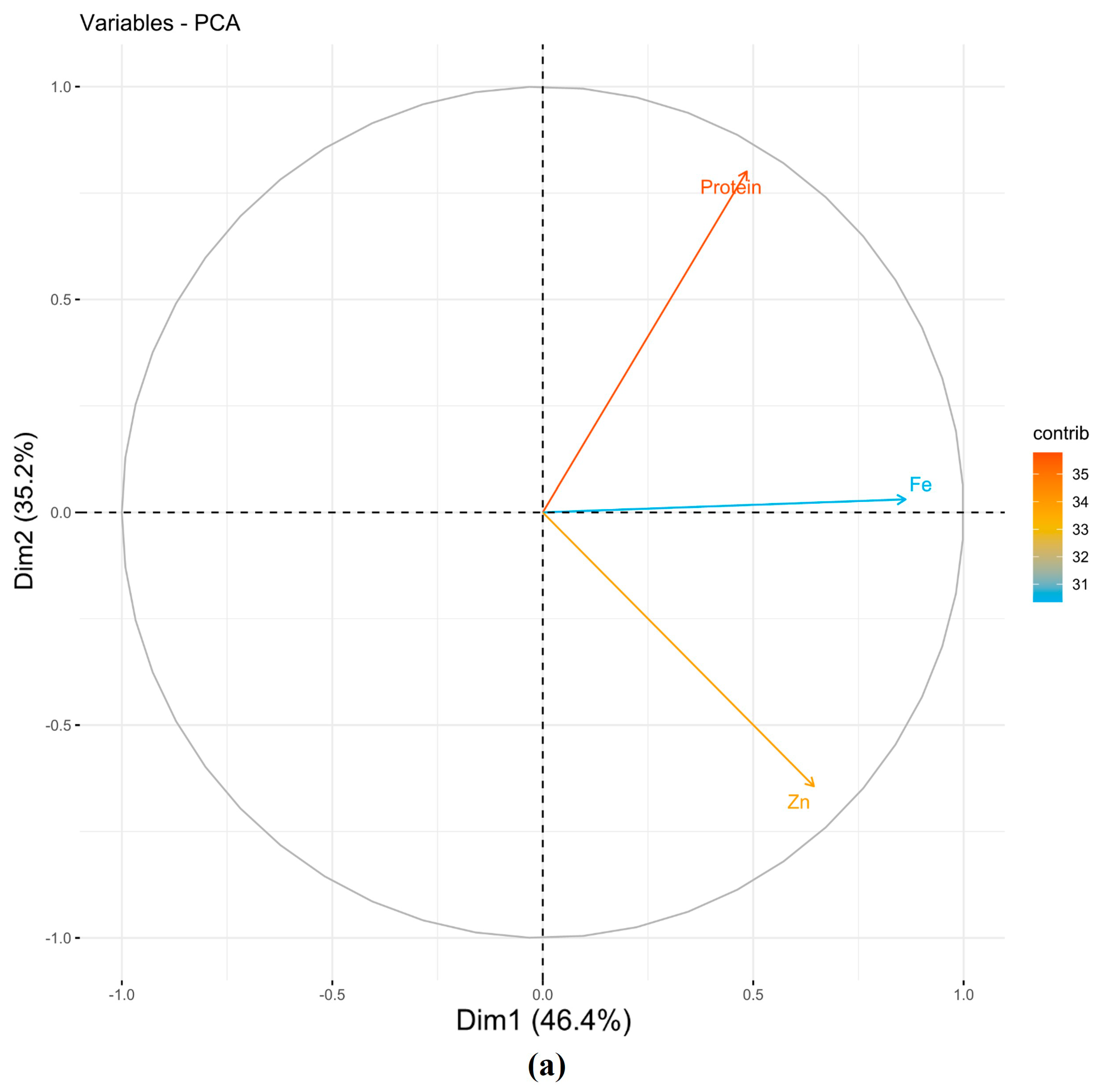
| PCs | Protein | Fe | Zn | Eigenvalues | Variance % | Cumulative Variance |
|---|---|---|---|---|---|---|
| NSI | ||||||
| PC1 | 0.41 | 0.73 | 0.55 | 1.39 | 46.44 | 46.44 |
| PC2 | −0.78 | −0.03 | 0.63 | 1.06 | 35.23 | 81.67 |
| PC3 | 0.47 | −0.68 | 0.56 | 0.55 | 18.33 | 100.00 |
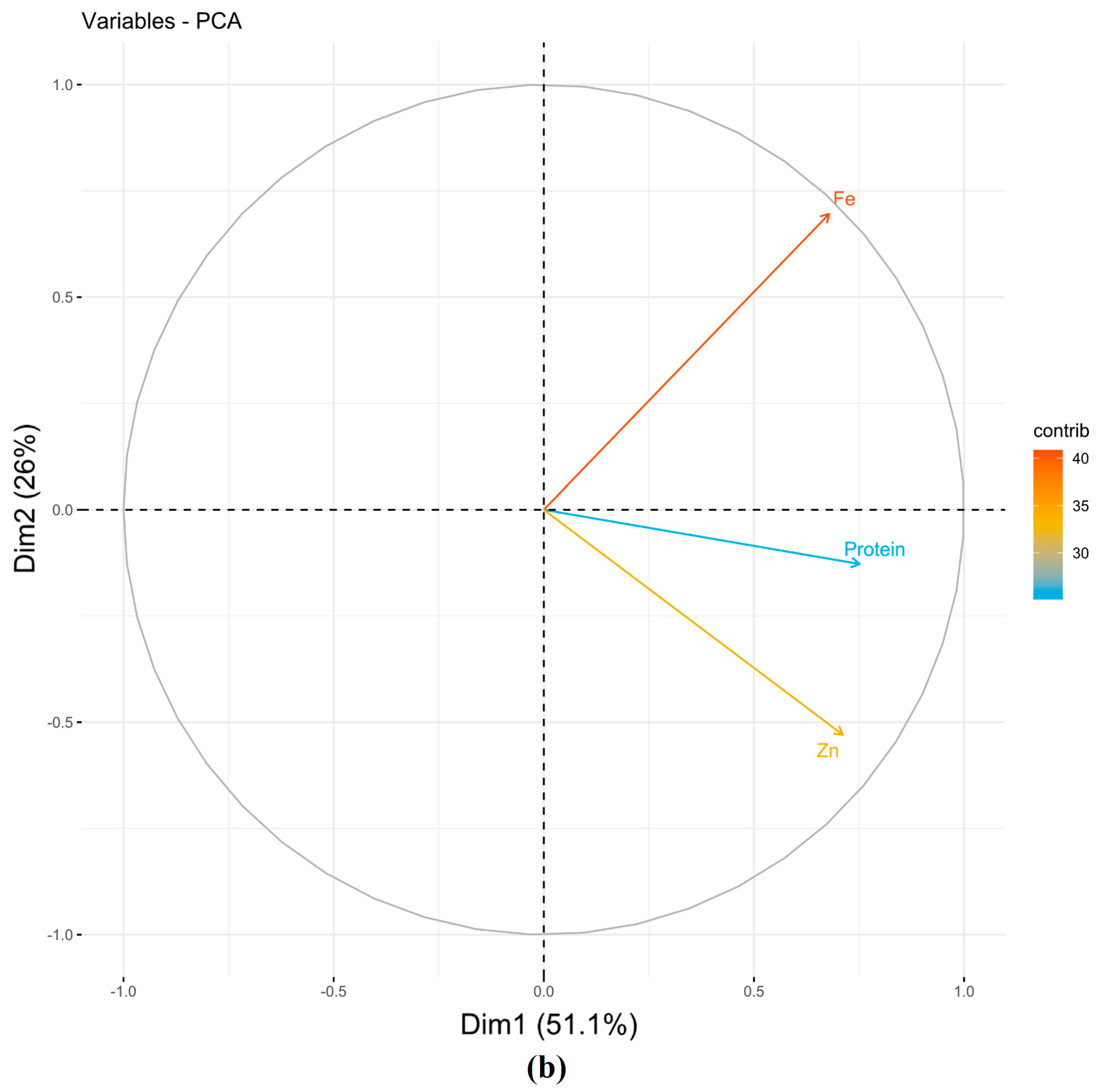
| NSII | ||||||
| PC1 | 0.61 | 0.55 | 0.57 | 1.53 | 51.09 | 51.09 |
| PC2 | −0.14 | 0.79 | −0.60 | 0.78 | 26.05 | 77.13 |
| PC3 | −0.78 | 0.28 | 0.56 | 0.69 | 22.87 | 100.00 |
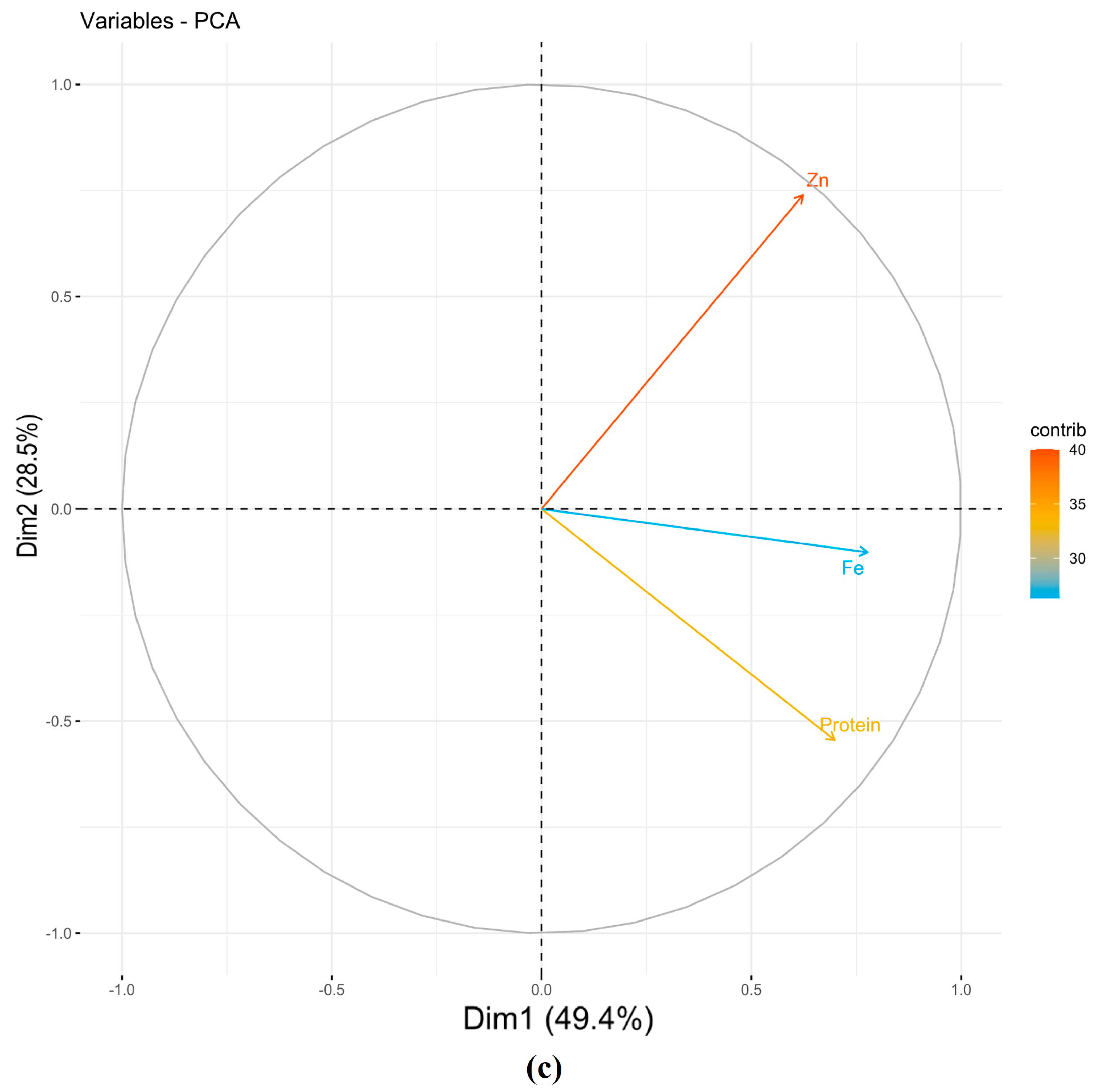
| Pooled seasons | ||||||
| PC1 | 0.57 | 0.64 | 0.51 | 1.48 | 49.37 | 49.37 |
| PC2 | −0.59 | −0.11 | 0.80 | 0.85 | 28.48 | 77.85 |
| PC3 | 0.57 | −0.76 | 0.31 | 0.66 | 22.15 | 100.00 |
| Population | Net Nucleotide Distance | Expected Heterozygosity | % of Membership | Mean Fixation Index (Fst) | |
|---|---|---|---|---|---|
| SPII | SPIII | ||||
| SPI | 0.44 | 0.29 | 0.23 | 0.29 | 0.63 |
| SPII | 0.19 | 0.19 | 0.49 | 0.64 | |
| SPIII | 0.17 | 0.22 | 0.61 | ||
| SNP | Model | Chromosome | Position | Allele 1 | Allele 2 | MAF | p-Value |
|---|---|---|---|---|---|---|---|
| Protein, NSI | |||||||
| S4_4477846 * | BLINK | 4 | 447,7846 | A | G | 0.49 | 2.52 × 10−7 |
| NSII | |||||||
| S4_4477846 | BLINK | 4 | 447,7846 | A | G | 0.49 | 2.42 × 10−9 |
| S6_2302393 | FarmCPU | 6 | 2,302,393 | T | C | 0.39 | 5.83 × 10−8 |
| S4_4477846 | FarmCPU | 4 | 447,7846 | A | G | 0.49 | 2.76 × 10−6 |
| S4_30653910 | FarmCPU | 4 | 30,653,910 | A | G | 0.31 | 3.35 × 10−6 |
| Pooled | |||||||
| S4_4477846 | BLINK | 4 | 4,477,846 | A | G | 0.49 | 4.90 × 10−9 |
| S6_2667543 | BLINK | 6 | 2,667,543 | C | A | 0.47 | 2.43 × 10−6 |
| Fe, NSI | |||||||
| S7_9379786 # | BLINK | 7 | 9,379,786 | T | C | 0.44 | 7.42 × 10−9 |
| NSII | |||||||
| S1_2772537 | FarmCPU | 1 | 2,772,537 | C | A | 0.31 | 2.89 × 10−7 |
| S4_34459338 * | BLINK | 4 | 34,459,338 | C | G | 0.34 | 5.09 × 10−7 |
| S1_11613376 * | FarmCPU | 1 | 11,613,376 | T | A | 0.42 | 6.96 × 10−7 |
| S1_11613376 | BLINK | 1 | 11,613,376 | T | A | 0.42 | 2.15 × 10−6 |
| S6_57720344 | FarmCPU | 6 | 57,720,344 | C | T | 0.08 | 5.56 × 10−6 |
| S4_34459338 | FarmCPU | 4 | 34,459,338 | C | G | 0.34 | 1.12 × 10−5 |
| Pooled | |||||||
| S7_9379786 | BLINK | 7 | 9,379,786 | T | C | 0.44 | 4.13 × 10−6 |
| S4_31996956 | BLINK | 4 | 31,996,956 | A | C | 0.14 | 4.52 × 10−6 |
| S1_11613376 | FarmCPU | 1 | 11,613,376 | T | A | 0.42 | 1.01 × 10−6 |
| S1_2772537 | FarmCPU | 1 | 2,772,537 | C | A | 0.31 | 2.34 × 10−6 |
| S7_9379786 | FarmCPU | 7 | 9,379,786 | T | C | 0.44 | 3.03 × 10−6 |
| Zn, NSI | |||||||
| S6_7891103 | BLINK | 6 | 7,891,103 | G | A | 0.29 | 3.52 × 10−7 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Srungarapu, R.; Mahendrakar, M.D.; Mohammad, L.A.; Chand, U.; Jagarlamudi, V.R.; Kondamudi, K.P.; Kudapa, H.; Samineni, S. Genome-Wide Association Analysis Reveals Trait-Linked Markers for Grain Nutrient and Agronomic Traits in Diverse Set of Chickpea Germplasm. Cells 2022, 11, 2457. https://doi.org/10.3390/cells11152457
Srungarapu R, Mahendrakar MD, Mohammad LA, Chand U, Jagarlamudi VR, Kondamudi KP, Kudapa H, Samineni S. Genome-Wide Association Analysis Reveals Trait-Linked Markers for Grain Nutrient and Agronomic Traits in Diverse Set of Chickpea Germplasm. Cells. 2022; 11(15):2457. https://doi.org/10.3390/cells11152457
Chicago/Turabian StyleSrungarapu, Rajasekhar, Mahesh Damodhar Mahendrakar, Lal Ahamed Mohammad, Uttam Chand, Venkata Ramana Jagarlamudi, Kiran Prakash Kondamudi, Himabindu Kudapa, and Srinivasan Samineni. 2022. "Genome-Wide Association Analysis Reveals Trait-Linked Markers for Grain Nutrient and Agronomic Traits in Diverse Set of Chickpea Germplasm" Cells 11, no. 15: 2457. https://doi.org/10.3390/cells11152457
APA StyleSrungarapu, R., Mahendrakar, M. D., Mohammad, L. A., Chand, U., Jagarlamudi, V. R., Kondamudi, K. P., Kudapa, H., & Samineni, S. (2022). Genome-Wide Association Analysis Reveals Trait-Linked Markers for Grain Nutrient and Agronomic Traits in Diverse Set of Chickpea Germplasm. Cells, 11(15), 2457. https://doi.org/10.3390/cells11152457