
Chronic Intermittent Hypoxia Increases Cell Proliferation in Hepatocellular Carcinoma

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Housing
2.2. DEN-Induced Rat Model of HCC
2.3. Intermittent Hypoxia Exposure Procedure
2.4. Immunohistochemical and Immunofluorescence Analyses
2.5. Quantitative Real Time Polymerase Chain Reaction (RT-qPCR)
2.6. RNA-Seq
2.7. Statistical Analysis
3. Results
3.1. Chronic DEN Promotes Hypoxia in Liver
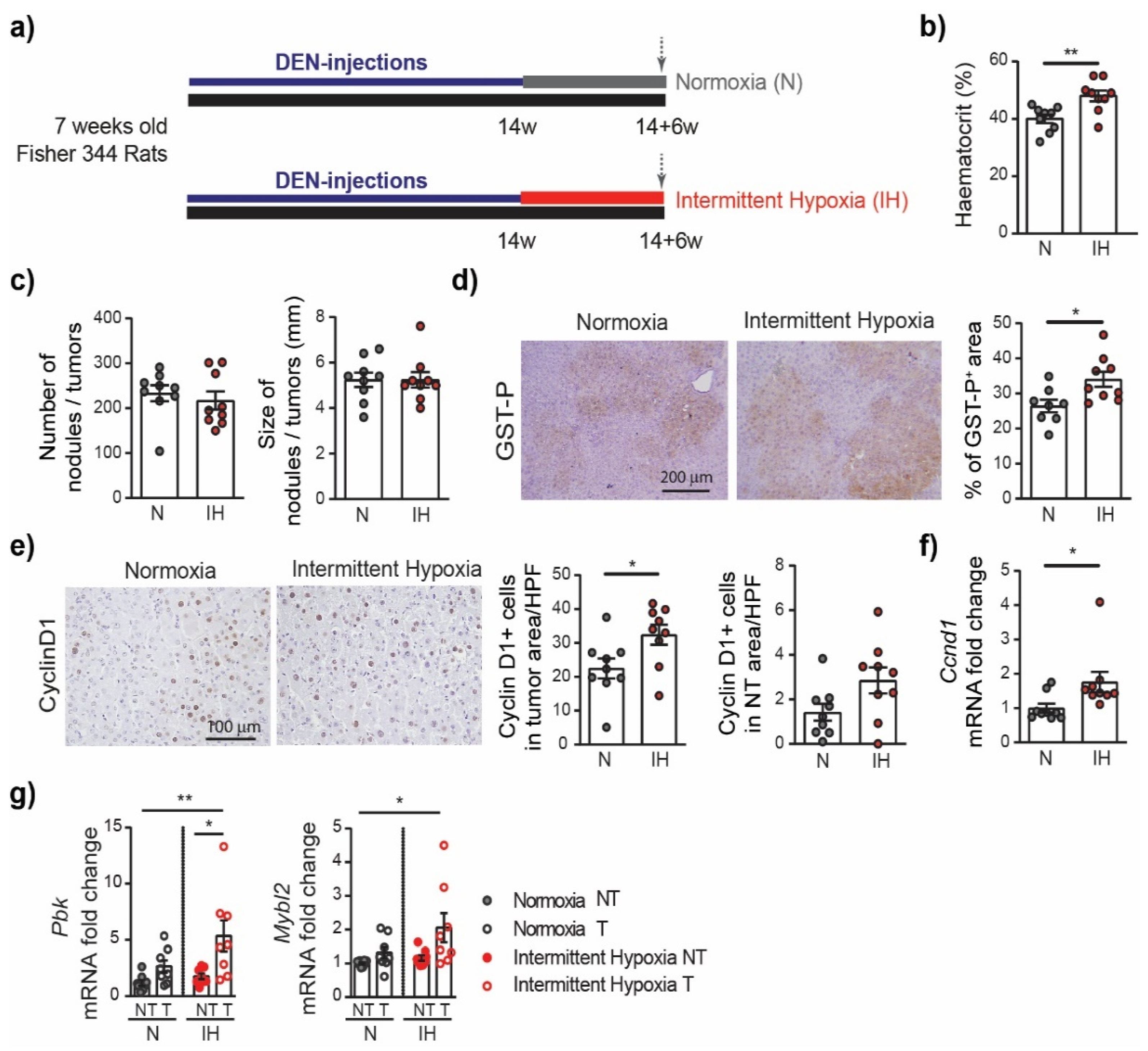
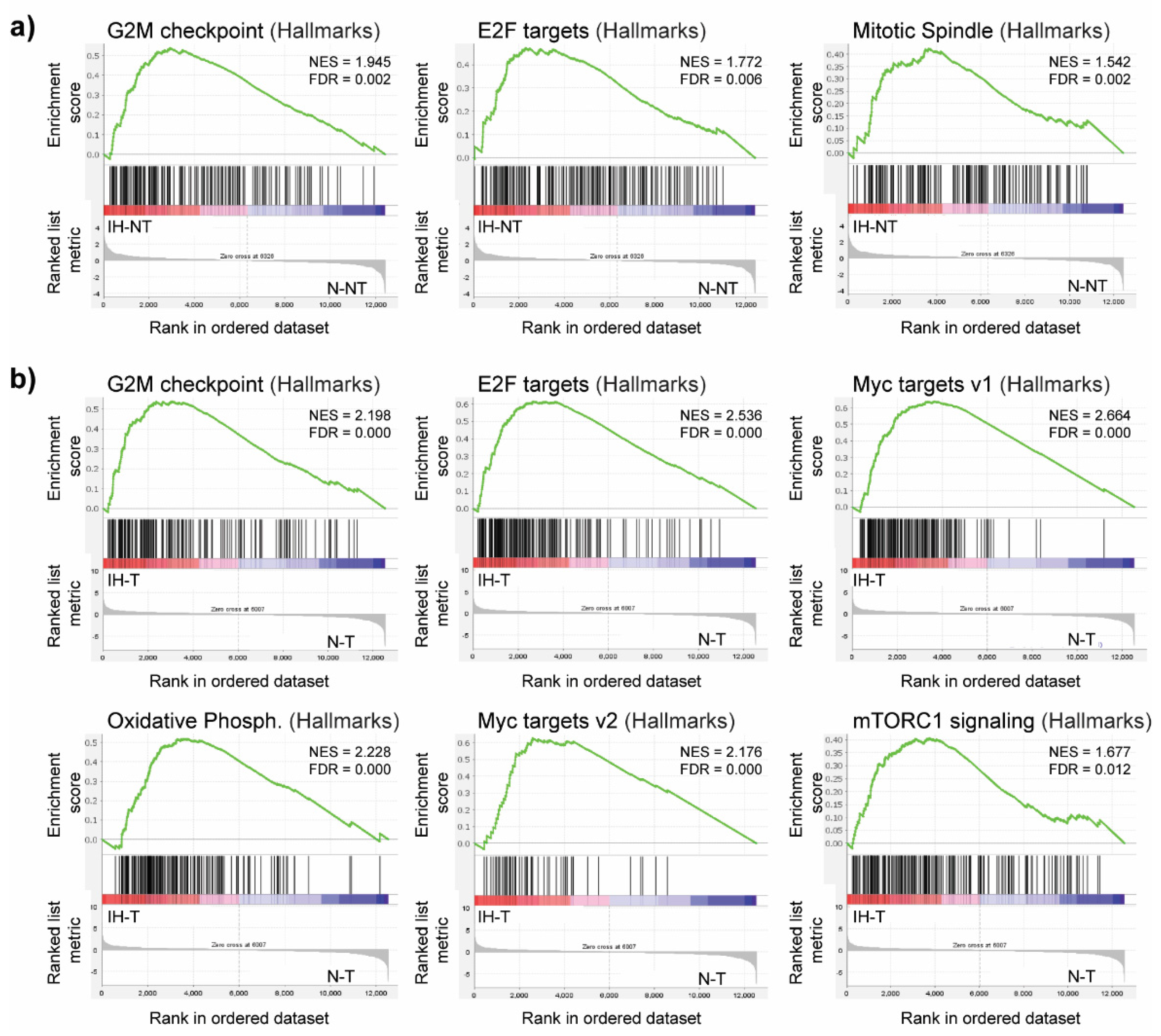
3.2. Intermittent Hypoxia Increases Cell Proliferation in Chronic DEN-Induced HCC
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dewan, N.A.; Nieto, F.J.; Somers, V.K. Intermittent hypoxemia and OSA: Implications for comorbidities. Chest 2015, 147, 266–274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nieto, F.J.; Peppard, P.E.; Young, T.; Finn, L.; Hla, K.M.; Farré, R. Sleep-disordered breathing and cancer mortality: Results from the Wisconsin Sleep Cohort Study. Am. J. Respir. Crit. Care Med. 2012, 186, 190–194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marshall, N.S.; Wong, K.K.H.; Cullen, S.R.J.; Knuiman, M.W.; Grunstein, R.R. Sleep Apnea and 20-Year Follow-Up for All-Cause Mortality, Stroke, and Cancer Incidence and Mortality in the Busselton Health Study Cohort. J. Clin. Sleep Med. 2014, 10, 355–362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gozal, D.; Almendros, I.; Phipps, A.I.; Campos-Rodriguez, F.; Martínez-García, M.A.; Farré, R. Sleep Apnoea Adverse Effects on Cancer: True, False, or Too Many Confounders? Int. J. Mol. Sci. 2020, 21, 8779. [Google Scholar] [CrossRef] [PubMed]
- Justeau, G.; Gervès-Pinquié, C.; Le Vaillant, M.; Trzepizur, W.; Meslier, N.; Goupil, F.; Pigeanne, T.; Launois, S.; Leclair-Visonneau, L.; Masson, P.; et al. Association Between Nocturnal Hypoxemia and Cancer Incidence in Patients Investigated for OSA: Data from a Large Multicenter French Cohort. Chest 2020, 158, 2610–2620. [Google Scholar] [CrossRef]
- Almendros, I.; Montserrat, J.M.; Ramírez, J.; Torres, M.; Duran-Cantolla, J.; Navajas, D.; Farré, R. Intermittent hypoxia enhances cancer progression in a mouse model of sleep apnoea. Eur. Respir. J. 2012, 39, 215–217. [Google Scholar] [CrossRef]
- Yoon, D.W.; So, D.; Min, S.; Kim, J.; Lee, M.; Khalmuratova, R.; Cho, C.-H.; Park, J.-W.; Shin, H.-W. Accelerated tumor growth under intermittent hypoxia is associated with hypoxia-inducible factor-1-dependent adaptive responses to hypoxia. Oncotarget 2017, 8, 61592–61603. [Google Scholar] [CrossRef] [Green Version]
- Kang, H.S.; Kwon, H.Y.; Kim, I.K.; Ban, W.H.; Kim, S.W.; Kang, H.H.; Yeo, C.D.; Lee, S.H. Intermittent hypoxia exacerbates tumor progression in a mouse model of lung cancer. Sci. Rep. 2020, 10, 1854. [Google Scholar] [CrossRef]
- Chen, A.; Sceneay, J.; Gödde, N.; Kinwel, T.; Ham, S.; Thompson, E.W.; Humbert, P.O.; Möller, A. Intermittent hypoxia induces a metastatic phenotype in breast cancer. Oncogene 2018, 37, 4214–4225. [Google Scholar] [CrossRef]
- Vilaseca, A.; Campillo, N.; Torres, M.; Musquera, M.; Gozal, D.; Montserrat, J.M.; Alcaraz, A.; Touijer, K.A.; Farré, R.; Almendros, I. Intermittent hypoxia increases kidney tumor vascularization in a murine model of sleep apnea. PLoS ONE 2017, 12, e0179444. [Google Scholar] [CrossRef] [Green Version]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Aron-Wisnewsky, J.; Minville, C.; Tordjman, J.; Lévy, P.; Bouillot, J.L.; Basdevant, A.; Bedossa, P.; Clément, K.; Pépin, J.L. Chronic intermittent hypoxia is a major trigger for non-alcoholic fatty liver disease in morbid obese. J. Hepatol. 2012, 56, 225–233. [Google Scholar] [CrossRef] [PubMed]
- Trzepizur, W.; Boursier, J.; Le Vaillant, M.; Ducluzeau, P.H.; Dubois, S.; Henni, S.; Abraham, P.; Aubé, C.; Calès, P.; Gagnadoux, F. Increased liver stiffness in patients with severe sleep apnoea and metabolic comorbidities. Eur. Respir. J. 2018, 51, 1800601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trzepizur, W.; Boursier, J.; Berréhare, A.; Le Vaillant, M.; Andriantsitohaina, R.; Ducluzeau, P.-H.; Dubois, S.; Henni, S.; Abraham, P.; Calès, P.; et al. Obstructive sleep apnoea severity and liver steatosis measured by magnetic resonance imaging. Eur. Respir. J. 2020, 55, 1901514. [Google Scholar] [CrossRef]
- Kurma, K.; Manches, O.; Chuffart, F.; Sturm, N.; Gharzeddine, K.; Zhang, J.; Mercey-Ressejac, M.; Rousseaux, S.; Millet, A.; Lerat, H.; et al. DEN-Induced Rat Model Reproduces Key Features of Human Hepatocellular Carcinoma. Cancers 2021, 13, 4981. [Google Scholar] [CrossRef]
- Jilkova, Z.M.; Kuyucu, A.Z.; Kurma, K.; Ahmad Pour, S.T.; Roth, G.S.; Abbadessa, G.; Yu, Y.; Schwartz, B.; Sturm, N.; Marche, P.N.; et al. Combination of AKT inhibitor ARQ 092 and sorafenib potentiates inhibition of tumor progression in cirrhotic rat model of hepatocellular carcinoma. Oncotarget 2018, 9, 11145–11158. [Google Scholar] [CrossRef]
- Roth, G.S.; Macek Jilkova, Z.; Zeybek Kuyucu, A.; Kurma, K.; Ahmad Pour, S.T.; Abbadessa, G.; Yu, Y.; Busser, B.; Marche, P.N.; Leroy, V.; et al. Efficacy of AKT Inhibitor ARQ 092 Compared with Sorafenib in a Cirrhotic Rat Model with Hepatocellular Carcinoma. Mol. Cancer Ther. 2017, 16, 2157–2165. [Google Scholar] [CrossRef] [Green Version]
- Arnaud, C.; Beguin, P.C.; Lantuejoul, S.; Pepin, J.L.; Guillermet, C.; Pelli, G.; Burger, F.; Buatois, V.; Ribuot, C.; Baguet, J.P.; et al. The inflammatory preatherosclerotic remodeling induced by intermittent hypoxia is attenuated by RANTES/CCL5 inhibition. Am. J. Respir. Crit. Care Med. 2011, 184, 724–731. [Google Scholar] [CrossRef] [Green Version]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast universal RNA-seq aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef]
- Anders, S.; Pyl, P.T.; Huber, W. HTSeq—A Python framework to work with high-throughput sequencing data. Bioinformatics 2015, 31, 166–169. [Google Scholar] [CrossRef]
- Varet, H.; Brillet-Guéguen, L.; Coppée, J.Y.; Dillies, M.A. SARTools: A DESeq2- and EdgeR-Based R Pipeline for Comprehensive Differential Analysis of RNA-Seq Data. PLoS ONE 2016, 11, e0157022. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anders, S.; Huber, W. Differential expression analysis for sequence count data. Genome Biol. 2010, 11, R106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [Green Version]
- Hänzelmann, S.; Castelo, R.; Guinney, J. GSVA: Gene set variation analysis for microarray and RNA-Seq data. BMC Bioinform. 2013, 14, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hugo, W.; Zaretsky, J.M.; Sun, L.; Song, C.; Moreno, B.H.; Hu-Lieskovan, S.; Berent-Maoz, B.; Pang, J.; Chmielowski, B.; Cherry, G.; et al. Genomic and Transcriptomic Features of Response to Anti-PD-1 Therapy in Metastatic Melanoma. Cell 2016, 165, 35–44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, B.; Tang, B.; Gao, J.; Li, J.; Kong, L.; Qin, L. A hypoxia-related signature for clinically predicting diagnosis, prognosis and immune microenvironment of hepatocellular carcinoma patients. J. Transl. Med. 2020, 18, 342. [Google Scholar] [CrossRef]
- Cai, J.; Hu, M.; Chen, Z.; Ling, Z. The roles and mechanisms of hypoxia in liver fibrosis. J. Transl. Med. 2021, 19, 186. [Google Scholar] [CrossRef]
- McGuire, M.; Bradford, A. Chronic intermittent hypoxia increases haematocrit and causes right ventricular hypertrophy in the rat. Respir. Physiol. 1999, 117, 53–58. [Google Scholar] [CrossRef]
- Song, J.; Sundar, K.M.; Hoidal, J.; Prchal, J.T. Hematological Changes in Chronic Sustained Hypoxia and Chronic Intermittent Hypoxia in a Mouse Model. Blood 2019, 134, 3525. [Google Scholar] [CrossRef]
- Dhanasekaran, R.; Deutzmann, A.; Mahauad-Fernandez, W.D.; Hansen, A.S.; Gouw, A.M.; Felsher, D.W. The MYC oncogene—The grand orchestrator of cancer growth and immune evasion. Nat. Rev. Clin. Oncol. 2022, 19, 23–36. [Google Scholar] [CrossRef]
- Sundaram, S.S.; Halbower, A.; Pan, Z.; Robbins, K.; Capocelli, K.E.; Klawitter, J.; Shearn, C.T.; Sokol, R.J. Nocturnal hypoxia-induced oxidative stress promotes progression of pediatric non-alcoholic fatty liver disease. J. Hepatol. 2016, 65, 560–569. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benotti, P.; Wood, G.C.; Argyropoulos, G.; Pack, A.; Keenan, B.T.; Gao, X.; Gerhard, G.; Still, C. The impact of obstructive sleep apnea on nonalcoholic fatty liver disease in patients with severe obesity. Obesity 2016, 24, 871–877. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Llovet, J.M.; Kelley, R.K.; Villanueva, A.; Singal, A.G.; Pikarsky, E.; Roayaie, S.; Lencioni, R.; Koike, K.; Zucman-Rossi, J.; Finn, R.S. Hepatocellular carcinoma. Nat. Rev. Dis. Primers 2021, 7, 6. [Google Scholar] [CrossRef] [PubMed]
- Dang, H.; Pomyen, Y.; Martin, S.P.; Dominguez, D.A.; Yim, S.Y.; Lee, J.S.; Budhu, A.; Shah, A.P.; Bodzin, A.S.; Wang, X.W. NELFE-Dependent MYC Signature Identifies a Unique Cancer Subtype in Hepatocellular Carcinoma. Sci. Rep. 2019, 9, 3369. [Google Scholar] [CrossRef] [PubMed]
- Carreras, A.; Kayali, F.; Zhang, J.; Hirotsu, C.; Wang, Y.; Gozal, D. Metabolic effects of intermittent hypoxia in mice: Steady versus high-frequency applied hypoxia daily during the rest period. Am. J. Physiol-Regul. Integr. Comp. Physiol. 2012, 303, R700–R709. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uchiyama, T.; Ota, H.; Ohbayashi, C.; Takasawa, S. Effects of Intermittent Hypoxia on Cytokine Expression Involved in Insulin Resistance. Int. J. Mol. Sci. 2021, 22, 12898. [Google Scholar] [CrossRef]
- Ota, H.; Fujita, Y.; Yamauchi, M.; Muro, S.; Kimura, H.; Takasawa, S. Relationship Between Intermittent Hypoxia and Type 2 Diabetes in Sleep Apnea Syndrome. Int. J. Mol. Sci. 2019, 20, 4756. [Google Scholar] [CrossRef] [Green Version]
- Wojan, F.; Stray-Gundersen, S.; Nagel, M.J.; Lalande, S. Short exposure to intermittent hypoxia increases erythropoietin levels in healthy individuals. J. Appl. Physiol. 2021, 130, 1955–1960. [Google Scholar] [CrossRef]
- Helm, J.; Drukewitz, S.; Poser, I.; Richter, S.; Friedemann, M.; William, D.; Mohr, H.; Nölting, S.; Robledo, M.; Bornstein, S.R.; et al. Treatment of Pheochromocytoma Cells with Recurrent Cycles of Hypoxia: A New Pseudohypoxic In Vitro Model. Cells 2022, 11, 560. [Google Scholar] [CrossRef]
- Uchiyama, T.; Ota, H.; Itaya-Hironaka, A.; Shobatake, R.; Yamauchi, A.; Sakuramoto-Tsuchida, S.; Makino, M.; Kimura, H.; Takeda, M.; Ohbayashi, C.; et al. Up-regulation of selenoprotein P and HIP/PAP mRNAs in hepatocytes by intermittent hypoxia via down-regulation of miR-203. Biochem. Biophys. Rep. 2017, 11, 130–137. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
| Normoxia-Treated Rats, n = 9 | Intermittent Hypoxia-Treated Rats, n = 9 | p Value | |
|---|---|---|---|
| Body weight (BW), g | 242 [182–283] | 242 [222–270] | 0.560 |
| Liver weight, g | 13.5 [7.0–15.1] | 13.9 [10.8–15.4] | 0.743 |
| Liver weight/BW, % | 5.26 ± 0.29 | 5.39 ± 0.29 | 0.782 |
| Heart weight, mg | 754.8 ± 33.5 | 741.6 ± 35.2 | 0.789 |
| Spleen weight, mg | 1081 ± 67.9 | 987.3 ± 28.8 | 0.224 |
| Visceral adipose tissue, g | 1.68 ± 0.29 | 1.58 ± 0.08 | 0.754 |
| Glucose, mmol/L (before sacrifice) | 5.1 ± 0.1 | 4.4 ± 0.1 | <0.0001 |
| ALP, IU/L | 200.9 ± 7.2 | 213.3 ± 8.7 | 0.286 |
| ALT, IU/L | 156.8 ± 17.3 | 196.0 ± 17.5 | 0.131 |
| AST, IU/L | 329.6 ± 62.2 | 365.6 ± 56.7 | 0.675 |
| Total Bilirubin, µmol/L | 3.20 (0.1–14.4) | 3.4 [1.2–5.5] | >0.999 |
| Albumin, g/L | 37.2 ± 0.7 | 37.6 ± 0.4 | 0.605 |
| Cholesterol, mmol/L | 3.4 ± 0.2 | 3.2 ± 0.2 | 0.509 |
| Triglycerides, mmol/L | 0.63 ± 0.06 | 0.91 ± 0.14 | 0.096 |
| GGT, IU/L | 50.7 ± 4.8 | 55.8 ± 5.4 | 0.491 |
| Liver Triglycerides, mmol/L | 20.3 ± 1.5 | 21.3 ± 1.0 | 0.567 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Carreres, L.; Mercey-Ressejac, M.; Kurma, K.; Ghelfi, J.; Fournier, C.; Manches, O.; Chuffart, F.; Rousseaux, S.; Minoves, M.; Decaens, T.; et al. Chronic Intermittent Hypoxia Increases Cell Proliferation in Hepatocellular Carcinoma. Cells 2022, 11, 2051. https://doi.org/10.3390/cells11132051
Carreres L, Mercey-Ressejac M, Kurma K, Ghelfi J, Fournier C, Manches O, Chuffart F, Rousseaux S, Minoves M, Decaens T, et al. Chronic Intermittent Hypoxia Increases Cell Proliferation in Hepatocellular Carcinoma. Cells. 2022; 11(13):2051. https://doi.org/10.3390/cells11132051
Chicago/Turabian StyleCarreres, Lydie, Marion Mercey-Ressejac, Keerthi Kurma, Julien Ghelfi, Carole Fournier, Olivier Manches, Florent Chuffart, Sophie Rousseaux, Mélanie Minoves, Thomas Decaens, and et al. 2022. "Chronic Intermittent Hypoxia Increases Cell Proliferation in Hepatocellular Carcinoma" Cells 11, no. 13: 2051. https://doi.org/10.3390/cells11132051
APA StyleCarreres, L., Mercey-Ressejac, M., Kurma, K., Ghelfi, J., Fournier, C., Manches, O., Chuffart, F., Rousseaux, S., Minoves, M., Decaens, T., Lerat, H., & Macek Jilkova, Z. (2022). Chronic Intermittent Hypoxia Increases Cell Proliferation in Hepatocellular Carcinoma. Cells, 11(13), 2051. https://doi.org/10.3390/cells11132051