
Prostaglandin 2α Promotes Autophagy and Mitochondrial Energy Production in Fish Hepatocytes

 ,
,  , ,
, , 

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Culture and Treatments
2.2. RNA Interference and Inhibitor Treatment
2.3. LD Staining
2.4. Immunofluorescence Staining
2.5. PG Analysis
2.6. Lysosomal and Mitochondrial Staining
2.7. Triglyceride (TG), Free Fatty Acid, and Glycerol Content
2.8. ATP Determination
2.9. Real-Time Quantitative Reverse Transcription-PCR (qRT-PCR)
2.10. Mitochondrial DNA (mtDNA) Content Assay
2.11. Statistical Analysis
3. Results
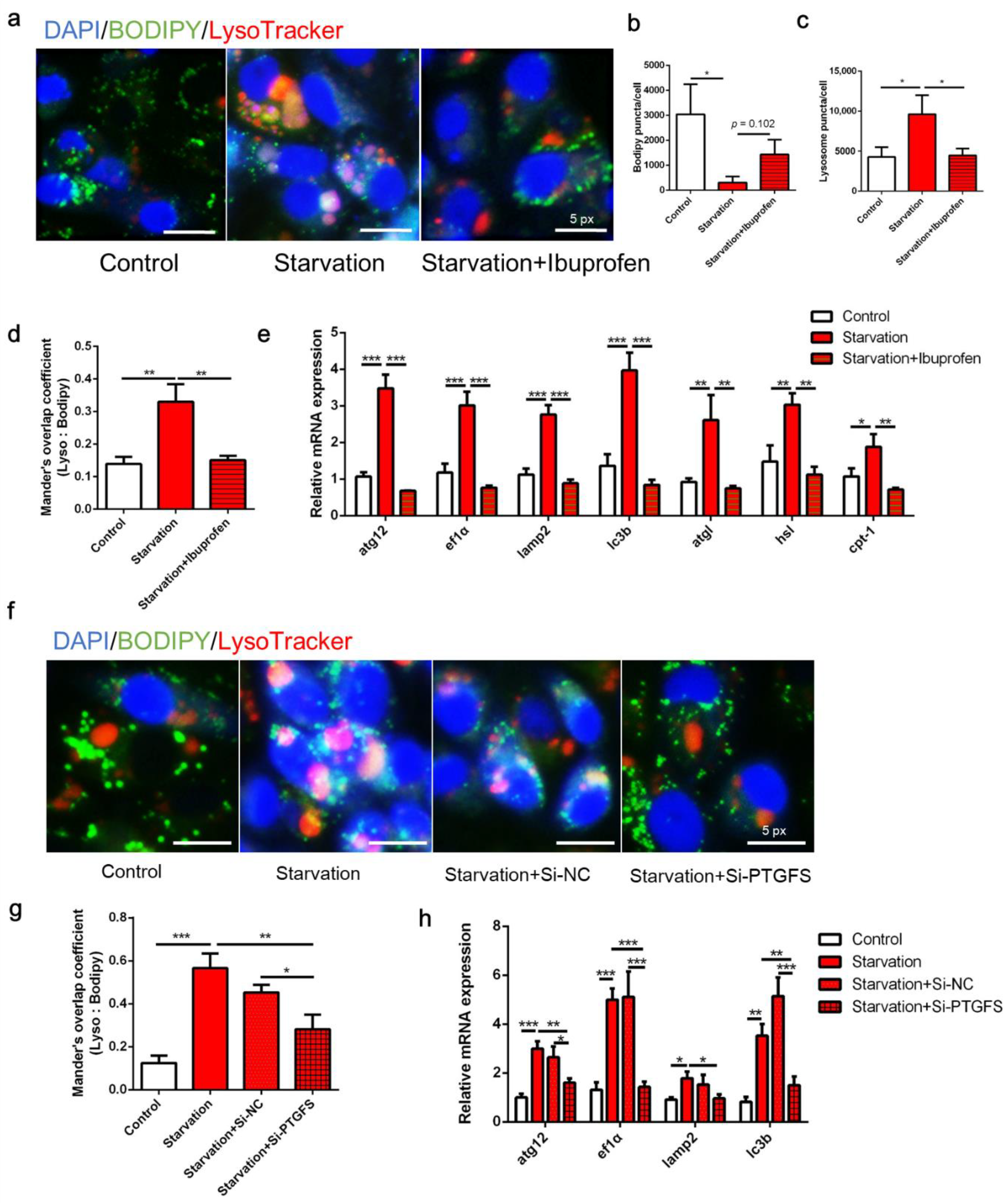
3.1. Serum Starvation Induces Autophagy, Lipid Depletion, and Mitochondrial Energy Production in ZFL Cells
3.2. Serum Starvation Induces COX Activity in ZFL Cells
3.3. COX Pathway Is Implicated in Serum Starvation-Induced Autophagy and LD Degradation in ZFL Cells
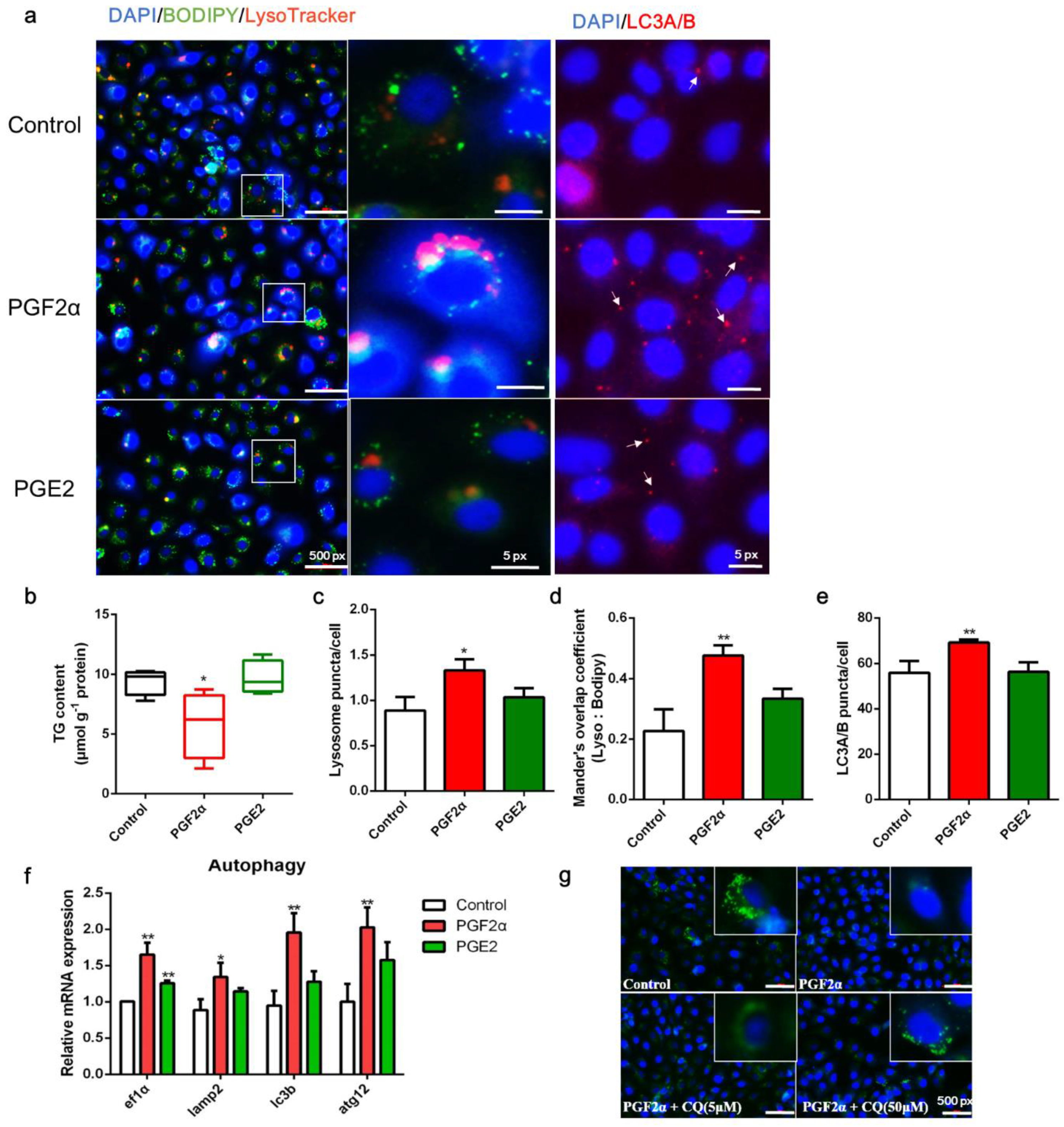
3.4. PGF2α Promotes Autophagy in ZFL Cells
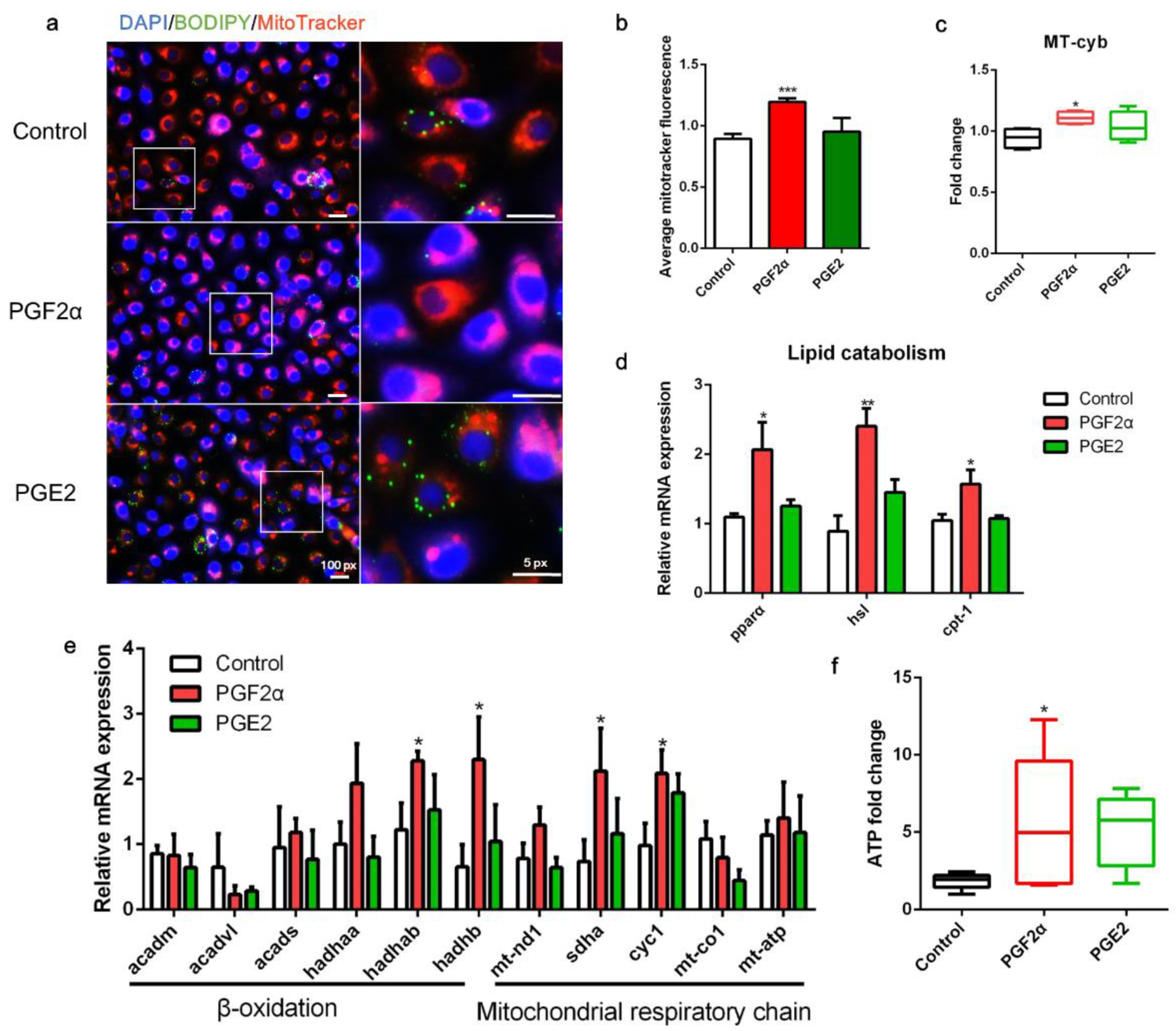
3.5. PGF2α Promotes Mitochondrial Energy Production in ZFL Cells
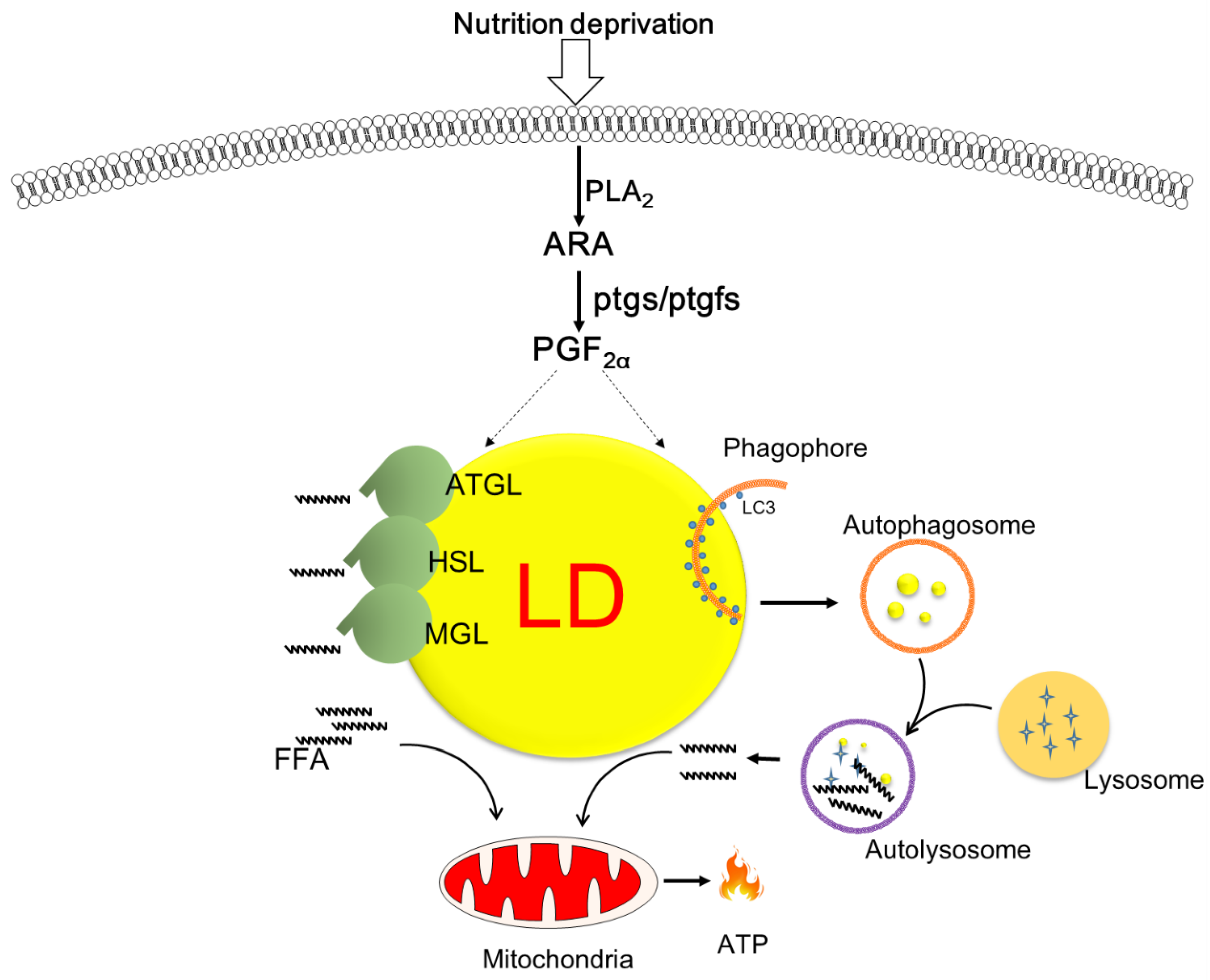
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Samuel, V.T.; Shulman, G.I. Nonalcoholic Fatty Liver Disease as a Nexus of Metabolic and Hepatic Diseases. Cell Metab. 2018, 27, 22–41. [Google Scholar] [CrossRef]
- Du, Z.Y.; Clouet, P.; Zheng, W.H.; Degrace, P.; Tian, L.X.; Liu, Y.J. Biochemical hepatic alterations and body lipid composition in the herbivorous grass carp (Ctenopharyngodon idella) fed high-fat diets. Brit. J. Nutr. 2006, 95, 905–915. [Google Scholar] [CrossRef]
- Tian, J.J.; Lu, R.H.; Ji, H.; Sun, J.; Li, C.; Liu, P.; Lei, C.X.; Chen, L.Q.; Du, Z.Y. Comparative analysis of the hepatopancreas transcriptome of grass carp (Ctenopharyngodon idellus) fed with lard oil and fish oil diets. Gene 2015, 565, 192–200. [Google Scholar] [CrossRef]
- Qian, Y.C.; Wang, X.; Ren, J.; Wang, J.; Limbu, S.M.; Li, R.X.; Zhou, W.H.; Qiao, F.; Zhang, M.L.; Du, Z.Y. Different effects of two dietary levels of tea polyphenols on the lipid deposition, immunity and antioxidant capacity of juvenile GIFT tilapia (Oreochromis niloticus) fed a high-fat diet. Aquaculture 2021, 542, 736896. [Google Scholar] [CrossRef]
- Chen, Y.; Sun, Z.; Liang, Z.; Xie, Y.; Tan, X.; Su, J.; Luo, Q.; Zhu, J.; Liu, Q.; Wang, A. Addition of l-carnitine to formulated feed improved growth performance, antioxidant status and lipid metabolism of juvenile largemouth bass, Micropterus salmoides. Aquaculture 2020, 518, 734434. [Google Scholar] [CrossRef]
- Tian, J.J.; Jin, Y.Q.; Yu, E.M.; Sun, J.H.; Xia, Y.; Zhang, K.; Li, Z.F.; Gong, W.B.; Wang, G.J.; Xie, J. Farnesoid X receptor is an effective target for modulating lipid accumulation in grass carp, Ctenopharyngodon idella. Aquaculture 2021, 534, 736248. [Google Scholar] [CrossRef]
- Ma, C.; Zhang, Q.; Greten, T.F. Nonalcoholic fatty liver disease promotes hepatocellular carcinoma through direct and indirect effects on hepatocytes. FEBS J. 2018, 285, 752–762. [Google Scholar] [CrossRef]
- Schulze, R.J.; Sathyanarayan, A.; Mashek, D.G. Breaking fat: The regulation and mechanisms of lipophagy. BBA Mol. Cell Biol. Lipids 2017, 1862, 1178–1187. [Google Scholar] [CrossRef]
- Han, S.L.; Qian, Y.C.; Limbu, S.M.; Wang, J.; Chen, L.Q.; Zhang, M.L.; Du, Z.Y. Lipolysis and lipophagy play individual and interactive roles in regulating triacylglycerol and cholesterol homeostasis and mitochondrial form in zebrafish. BBA Mol. Cell Biol. Lipids 2021, 1866, 158988. [Google Scholar] [CrossRef]
- Zimmermann, R.; Strauss, J.G.; Haemmerle, G.; Schoiswohl, G.; Birner-Gruenberger, R.; Riederer, M.; Lass, A.; Neuberger, G.; Eisenhaber, F.; Hermetter, A.; et al. Fat Mobilization in Adipose Tissue Is Promoted by Adipose Triglyceride Lipase. Science 2004, 306, 1383. [Google Scholar] [CrossRef]
- Sun, J.; Ji, H.; Li, X.X.; Shi, X.C.; Du, Z.Y.; Chen, L.Q. Lipolytic enzymes involving lipolysis in Teleost: Synteny, structure, tis-sue distribution, and expression in grass carp (Ctenopharyngodon idella). Comp. Biochem. Phys. B 2016, 198, 110–118. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Xiao, P.Z.; Chang, Z.G.; Ji, H.; Du, Z.Y.; Chen, L.Q. Forkhead box O1 in grass carp Ctenopharyngodon idella: Molecular characterization, gene structure, tissue distribution and mRNA expression in insulin-inhibited adipocyte lipolysis. Comp. Biochem. Phys. A 2017, 204, 76–84. [Google Scholar] [CrossRef]
- Sun, J.; Yang, Z.; Shi, X.C.; Ji, H.; Du, Z.Y.; Chen, L.Q. G0S2a1 (G0/G1 switch gene 2a1) is downregulated by TNF-α in grass carp (Ctenopharyngodon idellus) hepatocytes through PPARα inhibition. Gene 2018, 641, 1–7. [Google Scholar] [CrossRef]
- Singh, R.; Cuervo, A.M. Autophagy in the Cellular Energetic Balance. Cell Metab. 2011, 13, 495–504. [Google Scholar] [CrossRef]
- Singh, R.; Kaushik, S.; Wang, Y.; Xiang, Y.; Novak, I.; Komatsu, M.; Tanaka, K.; Cuervo, A.M.; Czaja, M.J. Autophagy regulates lipid metabolism. Nature 2009, 458, 1131–1135. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Han, S.L.; Li, L.Y.; Lu, D.L.; Limbu, S.M.; Li, D.L.; Zhang, M.L.; Du, Z.Y. Lipophagy is essential for lipid me-tabolism in fish. Sci. Bull. 2018, 63, 879–882. [Google Scholar] [CrossRef]
- Sathyanarayan, A.; Mashek, M.T.; Mashek, D.G. ATGL Promotes Autophagy/Lipophagy via SIRT1 to Control Hepatic Lipid Droplet Catabolism. Cell Rep. 2017, 19, 1–9. [Google Scholar] [CrossRef]
- Bartlett, K.; Eaton, S. Mitochondrial β-oxidation. Eur. J. Biochem. 2004, 271, 462–469. [Google Scholar] [CrossRef]
- Akram, M. Citric Acid Cycle and Role of its Intermediates in Metabolism. Cell Biochem. Biophys. 2014, 68, 475–478. [Google Scholar] [CrossRef] [PubMed]
- Tian, J.J.; Fu, B.; Yu, E.M.; Li, Y.P.; Xia, Y.; Li, Z.F.; Zhang, K.; Gong, W.B.; Yu, D.G.; Wang, G.J.; et al. Feeding Faba Beans (Vicia faba L.) Reduces Myocyte Metabolic Activity in Grass Carp (Ctenopharyngodon idellus). Front. Physiol. 2020, 11, 391. [Google Scholar] [CrossRef]
- Martin, S.A.; Brash, A.R.; Murphy, R.C. The discovery and early structural studies of arachidonic acid. J. Lipid Res. 2016, 57, 1126–1132. [Google Scholar] [CrossRef] [PubMed]
- Harizi, H.; Corcuff, J.-B.; Gualde, N. Arachidonic-acid-derived eicosanoids: Roles in biology and immunopathology. Trends Mol. Med. 2008, 14, 461–469. [Google Scholar] [CrossRef]
- Funk, C.D. Prostaglandins and Leukotrienes: Advances in Eicosanoid Biology. Science 2001, 294, 1871. [Google Scholar] [CrossRef]
- Yin, H.; Zhou, Y.; Zhu, M.; Hou, S.; Li, Z.; Zhong, H.; Lu, J.; Meng, T.; Wang, J.; Xia, L.; et al. Role of mitochondria in programmed cell death mediated by arachidonic acid-derived eicosanoids. Mitochondrion 2013, 13, 209–224. [Google Scholar] [CrossRef]
- Tian, J.J.; Lei, C.X.; Ji, H.; Chen, L.Q.; Du, Z.Y. Dietary Arachidonic Acid Has a Time-Dependent Differential Impact on Adipo-genesis Modulated via COX and LOX Pathways in Grass Carp Ctenopharyngodon idellus. Lipids 2016, 51, 1325–1338. [Google Scholar] [CrossRef]
- Tian, J.J.; Lei, C.X.; Ji, H.; Jin, A. Role of cyclooxygenase-mediated metabolites in lipid metabolism and expression of some immune-related genes in juvenile grass carp (Ctenopharyngodon idellus) fed arachidonic acid. Fish Physiol. Biochem. 2017, 43, 703–717. [Google Scholar] [CrossRef]
- Tian, J.J.; Lei, C.X.; Ji, H.; Kaneko, G.; Zhou, J.S.; Yu, H.B.; Li, Y.; Yu, E.M.; Xie, J. Comparative analysis of effects of dietary ar-achidonic acid and EPA on growth, tissue fatty acid composition, antioxidant response and lipid metabolism in juvenile grass carp, Ctenopharyngodon idellus. Brit. J. Nutr. 2017, 118, 411–422. [Google Scholar] [CrossRef] [PubMed]
- Chatzipanteli, K.; Rudolph, S.; Axelrod, L. Coordinate Control of Lipolysis by Prostaglandin E2 and Prostacyclin in Rat Adipose Tissue. Diabetes 1992, 41, 927. [Google Scholar] [CrossRef]
- Forman, B.M.; Tontonoz, P.; Chen, J.; Brun, R.P.; Spiegelman, B.M.; Evans, R.M. 15-Deoxy-Δ12,14-Prostaglandin J2 is a ligand for the adipocyte determination factor PPARγ. Cell 1995, 83, 803–812. [Google Scholar] [CrossRef]
- Fain, J.N.; Leffler, C.W.; Bahouth, S.W.; Rice, A.M.; Rivkees, S.A. Regulation of leptin release and lipolysis by PGE2 in rat adipose tissue. Prostaglandins Other Lipid Mediat. 2000, 62, 343–350. [Google Scholar] [CrossRef]
- Henkel, J.; Frede, K.; Schanze, N.; Vogel, H.; Schürmann, A.; Spruss, A.; Bergheim, I.; Püschel, G.P. Stimulation of fat accumulation in hepatocytes by PGE2-dependent repression of hepatic lipolysis, β-oxidation and VLDL-synthesis. Lab. Investig. 2012, 92, 1597–1606. [Google Scholar] [CrossRef]
- Tian, J.J.; Zhang, J.M.; Yu, E.M.; Sun, J.H.; Xia, Y.; Zhang, K.; Li, Z.F.; Gong, W.B.; Wang, G.J.; Xie, J. Identification and analysis of lipid droplet-related proteome in the adipose tissue of grass carp (Ctenopharyngodon idella) under fed and starved condi-tions. Comp. Biochem. Phys. D 2020, 36, 100710. [Google Scholar] [CrossRef]
- Choi, J.; Jo, M.; Lee, E.; Choi, D. ERK1/2 is involved in luteal cell autophagy regulation during corpus luteum regression via an mTOR-independent pathway. Mol. Hum. Reprod. 2014, 20, 972–980. [Google Scholar] [CrossRef]
- Wen, X.; Liu, L.; Li, S.; Lin, P.; Jin, Y. Prostaglandin F2α Induces Goat Corpus Luteum Regression via Endoplasmic Reticulum Stress and Autophagy. Front. Physiol. 2020, 11, 868. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT–PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef] [PubMed]
- Bento, C.F.; Renna, M.; Ghislat, G.; Puri, C.; Ashkenazi, A.; Vicinanza, M.; Menzies, F.M.; Rubinsztein, D.C. Mammalian Autoph-agy: How Does It Work? Annu. Rev. Biochem. 2016, 85, 685–713. [Google Scholar] [CrossRef]
- Yang, Z.; Klionsky, D.J. Mammalian autophagy: Core molecular machinery and signaling regulation. Curr. Opin. Cell Biol. 2010, 22, 124–131. [Google Scholar] [CrossRef] [PubMed]
- Alers, S.; Löffler, A.F.; Wesselborg, S.; Stork, B. Role of AMPK-mTOR-Ulk1/2 in the regulation of autophagy: Cross talk, shortcuts, and feedbacks. Mol. Cell Biol. 2012, 32, 2–11. [Google Scholar] [CrossRef]
- Wang, C.W. Lipid droplets, lipophagy, and beyond. BBA Mol. Cell Biol. Lipids 2016, 1861, 793–805. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Han, S.L.; Lu, D.L.; Li, L.Y.; Limbu, S.M.; Li, D.L.; Zhang, M.L.; Du, Z.Y. Inhibited Lipophagy Suppresses Lipid Me-tabolism in Zebrafish Liver Cells. Front. Physiol. 2019, 10, 1077. [Google Scholar] [CrossRef] [PubMed]
- Pirianov, G.; Colston, K.W. Interaction of vitamin D analogs with signaling pathways leading to active cell death in breast cancer cells. Steroids 2001, 66, 309–318. [Google Scholar] [CrossRef]
- Clark, J.D.; Lin, L.L.; Kriz, R.W.; Ramesha, C.S.; Sultzman, L.A.; Lin, A.Y.; Milona, N.; Knopf, J.L. A novel arachidonic acid-selective cytosolic PLA2 contains a Ca2+-dependent translocation domain with homology to PKC and GAP. Cell 1991, 65, 1043–1051. [Google Scholar] [CrossRef]
- Auguste, L.J.; Angus, L.; Stein, T.A.; Wise, L. Starvation and mucosal prostaglandin-E2 in gastric stress ulceration. Crit. Care Med. 1988, 16, 610–611. [Google Scholar] [CrossRef] [PubMed]
- Fujimori, K.; Aritake, K.; Urade, Y. Enhancement of prostaglandin D2 production through cyclooxygenase-2 and lipocalin-type prostaglandin D synthase by upstream stimulatory factor 1 in human brain-derived TE671 cells under serum starvation. Gene 2008, 426, 72–80. [Google Scholar] [CrossRef]
- Gartung, A.; Zhao, J.; Chen, S.; Mottillo, E.; VanHecke, G.C.; Ahn, Y.-H.; Maddipati, K.R.; Sorokin, A.; Granneman, J.; Lee, M.-J. Characterization of Eicosanoids Produced by Adipocyte Lipolysis: Implication of cyclooxygenase-2 in adipose inflamma-tion. J. Biol. Chem. 2016, 291, 16001–16010. [Google Scholar] [CrossRef] [PubMed]
- Takayuki, T.; Chika, F.; Akane, H.; Katsueki, O. Recent Advances in the Understanding of Teleost Medaka Ovulation: The Roles of Proteases and Prostaglandins. Zool. Sci. 2013, 30, 239–247. [Google Scholar] [CrossRef]
- Karpe, F.; Wejde, J.; Änggård, E. Dietary Arachidonic Acid Protects Mice against the Fatty Liver Induced by a High Fat Diet and by Ethanol. Acta Pharmacol. Toxicol. 1984, 55, 95–99. [Google Scholar] [CrossRef] [PubMed]
- Goheen, S.C.; Larkin, E.C.; Manix, M.; Rao, G.A. Dietary arachidonic acid reduces fatty liver, increases diet consumption and weight gain in ethanol-fed rats. Lipids 1980, 15, 328–336. [Google Scholar] [CrossRef] [PubMed]
- Tian, J.J.; Ji, H.; Oku, H.; Zhou, J.S. Effects of dietary arachidonic acid (ARA) on lipid metabolism and health status of juvenile grass carp, Ctenopharyngodon idellus. Aquaculture 2014, 430, 57–65. [Google Scholar] [CrossRef]
- Luo, Z.; Tan, X.Y.; Li, X.D.; Yin, G.J. Effect of dietary arachidonic acid levels on growth performance, hepatic fatty acid profile, intermediary metabolism and antioxidant responses for juvenile Synechogobius hasta. Aquacult. Nutr. 2012, 18, 340–348. [Google Scholar] [CrossRef]
- Fountoulaki, E.; Alexis, M.N.; Nengas, I.; Venou, B. Effects of dietary arachidonic acid (20:4n-6), on growth, body composition, and tissue fatty acid profile of gilthead bream fingerlings (Sparus aurata L.). Aquaculture 2003, 225, 309–323. [Google Scholar] [CrossRef]
- Wakai, E.; Aritake, K.; Urade, Y.; Fujimori, K. Prostaglandin D2 enhances lipid accumulation through suppression of lipolysis via DP2 (CRTH2) receptors in adipocytes. Biochem. Bioph. Res. Commun. 2017, 490, 393–399. [Google Scholar] [CrossRef] [PubMed]
- Dowaidar, M.; Gestin, M.; Cerrato, C.P.; Jafferali, M.H.; Margus, H.; Kivistik, P.A.; Ezzat, K.; Hallberg, E.; Pooga, M.; Hällbrink, M.; et al. Role of autophagy in cell-penetrating peptide transfection model. Sci. Rep. 2017, 7, 12635. [Google Scholar] [CrossRef]
- Breyer, R.M.; Bagdassarian, C.K.; Myers, S.A.; Breyer, M.D. Prostanoid Receptors: Subtypes and Signaling. Annu. Rev. Pharmacol. 2001, 41, 661–690. [Google Scholar] [CrossRef]
- Chaves, V.E.; Frasson, D.; Kawashita, N.H. Several agents and pathways regulate lipolysis in adipocytes. Biochimie 2011, 93, 1631–1640. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Geng, F.; Cheng, X.; Guo, Q.; Zhong, Y.; Cloughesy, T.F.; Yong, W.H.; Chakravarti, A.; Guo, D. Lipid Droplets Maintain Energy Homeostasis and Glioblastoma Growth via Autophagic Release of Stored Fatty Acids. iScience 2020, 23, 101569. [Google Scholar] [CrossRef]
- Tian, J.J.; Lei, C.X.; Ji, H.; Zhou, J.S.; Yu, H.B.; Li, Y.; Yu, E.M.; Xie, J. Dietary arachidonic acid decreases the expression of tran-scripts related to adipocyte development and chronic inflammation in the adipose tissue of juvenile grass carp, Ctenopha-ryngodon idella. Comp. Biochem. Phys. D 2019, 30, 122–132. [Google Scholar] [CrossRef]
- Landar, A.; Shiva, S.; Levonen, A.L.; Oh, J.Y.; Zaragoza, C.; Johnson, M.S.; Darley-Usmar, V.M. Induction of the permeability transition and cytochrome c release by 15-deoxy-Δ12,14-prostaglandin J2 in mitochondria. Biochem. J. 2006, 394, 185–195. [Google Scholar] [CrossRef]
- Sanin, D.E.; Matsushita, M.; Geltink, R.I.K.; Grzes, K.M.; Bakker, N.V.T.; Corrado, M.; Kabat, A.M.; Buck, M.D.; Qiu, J.; Lawless, S.J.; et al. Mitochondrial Membrane Potential Regulates Nuclear Gene Expression in Macrophages Exposed to Prostaglandin E2. Immunity 2018, 49, 1021–1033. [Google Scholar] [CrossRef]
- Okabe, K.; Malchesky, P.S.; Nose, Y. Protective Effect of Prostaglandin I2 on Hepatic Mitochondrial Function of the Preserved Rat Liver. Tohoku J. Exp. Med. 1986, 150, 373–379. [Google Scholar] [CrossRef] [PubMed]
- Chiou, G.Y.; Fong, J.C. Synergistic effect of prostaglandin F2α and cyclic AMP on glucose transport in 3T3-L1 adipocytes. J. Cell. Biochem. 2005, 94, 627–634. [Google Scholar] [CrossRef] [PubMed]
- Lescat, L.; Véron, V.; Mourot, B.; Péron, S.; Chenais, N.; Dias, K.; Riera-Heredia, N.; Beaumatin, F.; Pinel, K.; Priault, M.; et al. Chaperone-Mediated Autophagy in the Light of Evolution: Insight from Fish. Mol. Biol. Evol. 2020, 37, 2887–2899. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tian, J.; Du, Y.; Yu, E.; Lei, C.; Xia, Y.; Jiang, P.; Li, H.; Zhang, K.; Li, Z.; Gong, W.; et al. Prostaglandin 2α Promotes Autophagy and Mitochondrial Energy Production in Fish Hepatocytes. Cells 2022, 11, 1870. https://doi.org/10.3390/cells11121870
Tian J, Du Y, Yu E, Lei C, Xia Y, Jiang P, Li H, Zhang K, Li Z, Gong W, et al. Prostaglandin 2α Promotes Autophagy and Mitochondrial Energy Production in Fish Hepatocytes. Cells. 2022; 11(12):1870. https://doi.org/10.3390/cells11121870
Chicago/Turabian StyleTian, Jingjing, Yihui Du, Ermeng Yu, Caixia Lei, Yun Xia, Peng Jiang, Hongyan Li, Kai Zhang, Zhifei Li, Wangbao Gong, and et al. 2022. "Prostaglandin 2α Promotes Autophagy and Mitochondrial Energy Production in Fish Hepatocytes" Cells 11, no. 12: 1870. https://doi.org/10.3390/cells11121870
APA StyleTian, J., Du, Y., Yu, E., Lei, C., Xia, Y., Jiang, P., Li, H., Zhang, K., Li, Z., Gong, W., Xie, J., & Wang, G. (2022). Prostaglandin 2α Promotes Autophagy and Mitochondrial Energy Production in Fish Hepatocytes. Cells, 11(12), 1870. https://doi.org/10.3390/cells11121870