
Transcriptomic and Functional Evidence Show Similarities between Human Amniotic Epithelial Stem Cells and Keratinocytes

 , ,
, ,  , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Tissue Collection and Cell Culture
2.2. Three Dimensional Skin Equivalent Preparation
2.3. Lentiviral Transduction
2.4. Activation of Notch Signaling Using Jagged-1
2.5. Animal Experiments
2.6. RNA Sequencing
2.7. Bioinformatic Analysis
2.8. Real-Time PCR
2.9. Immunocytochemistry
2.10. Statistical Analysis
3. Results
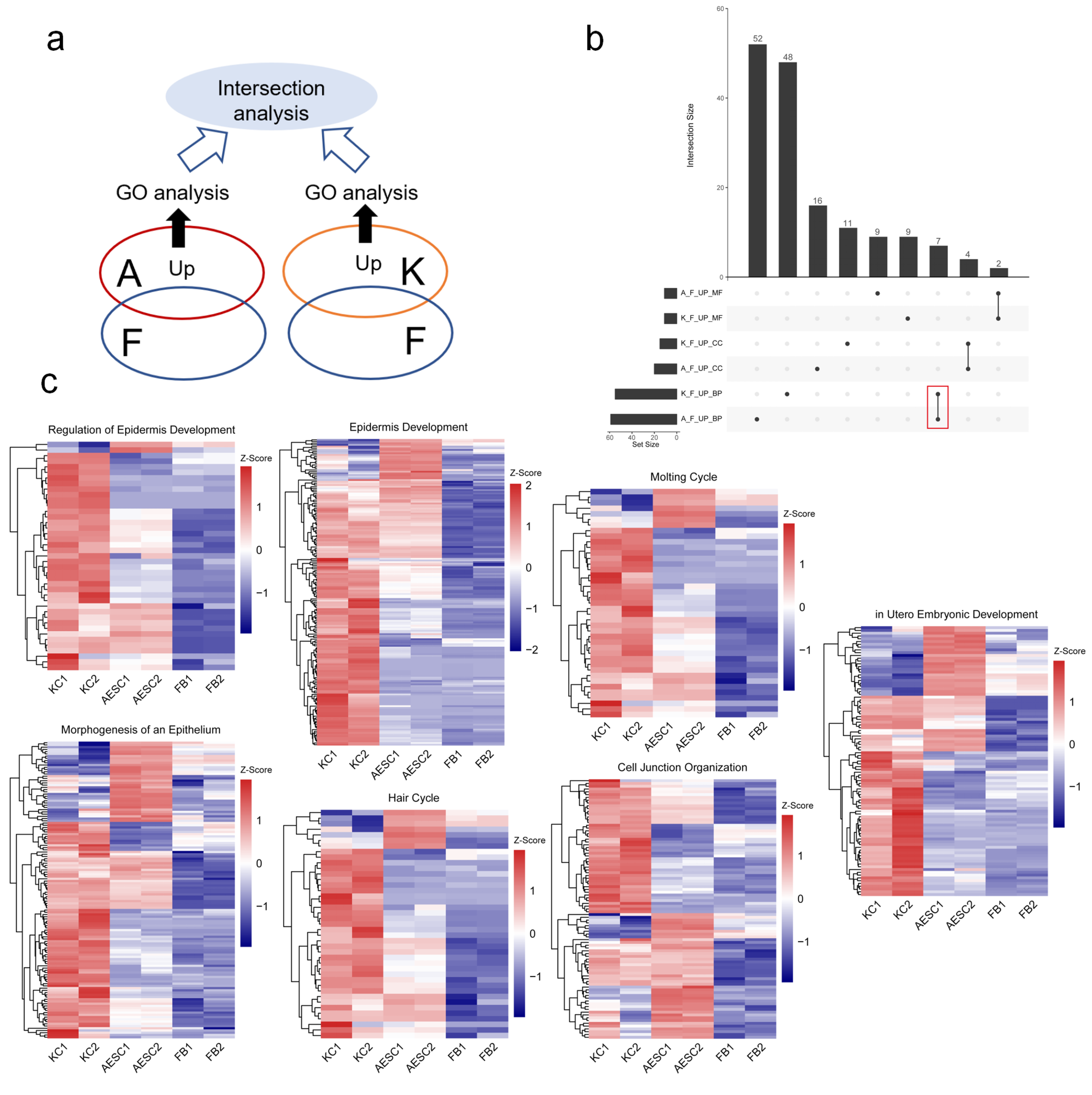
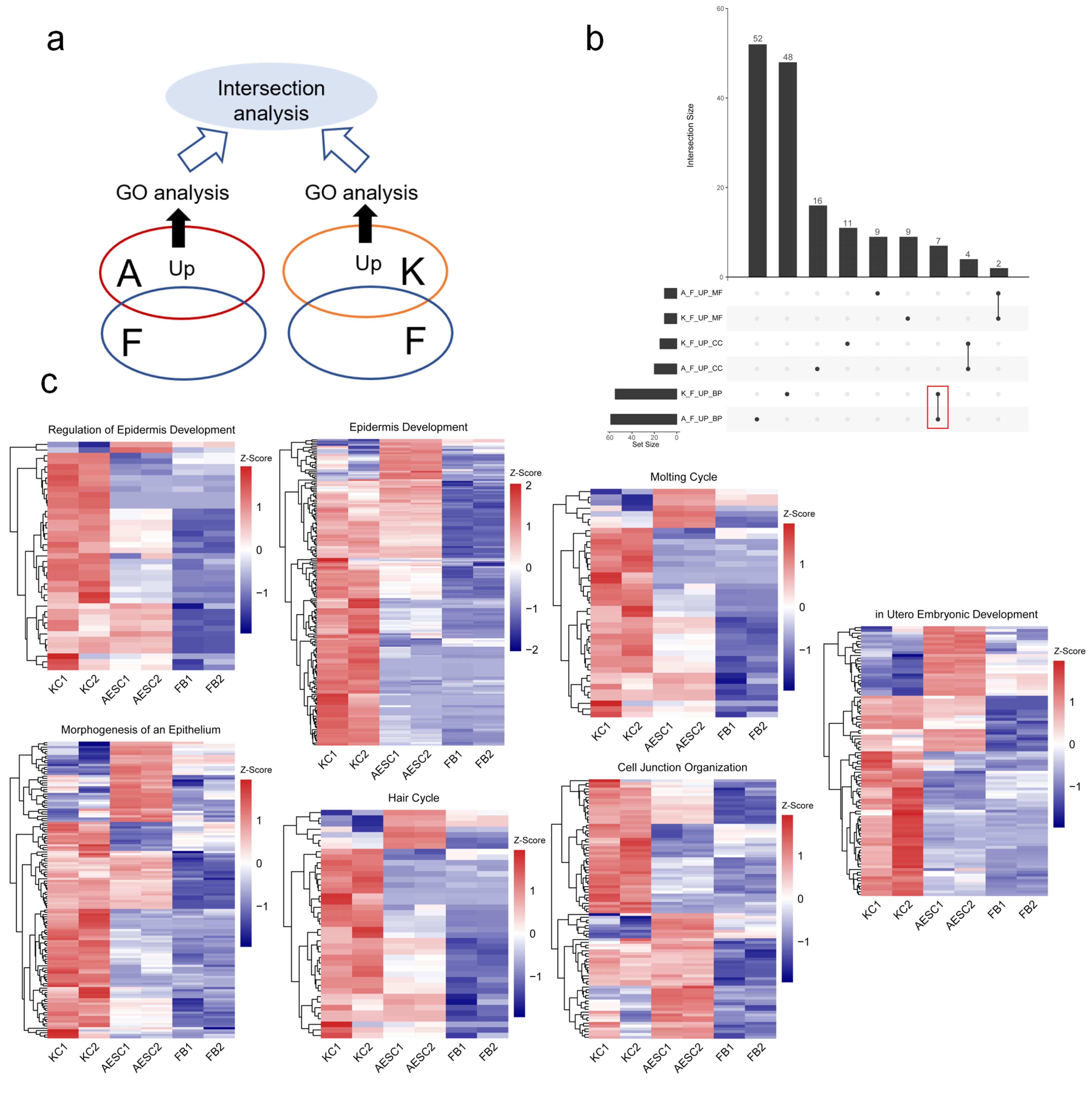
3.1. Similarities between AESCs and KCs in Epidermis-Associated Biological Processes by Transcriptomics
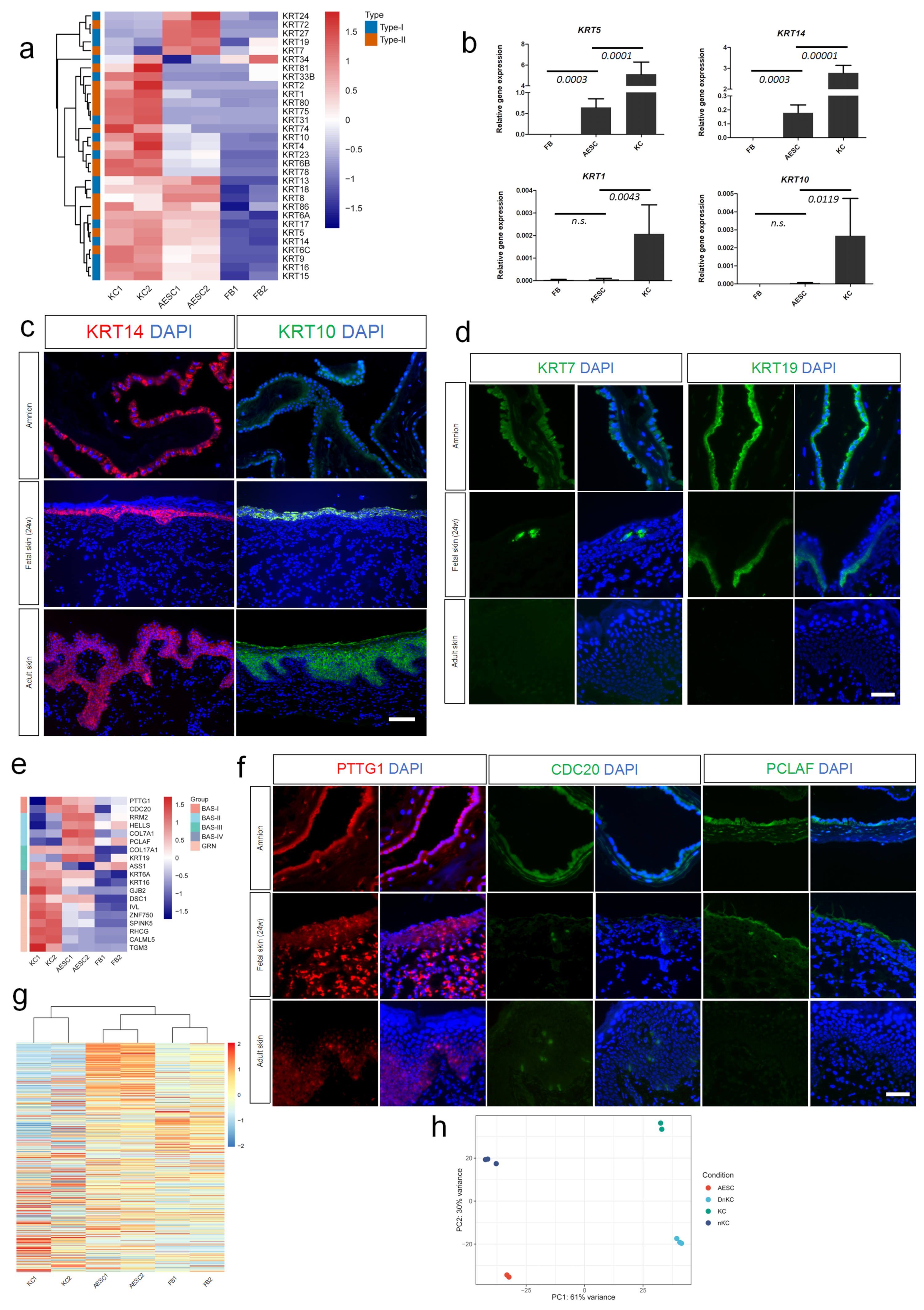
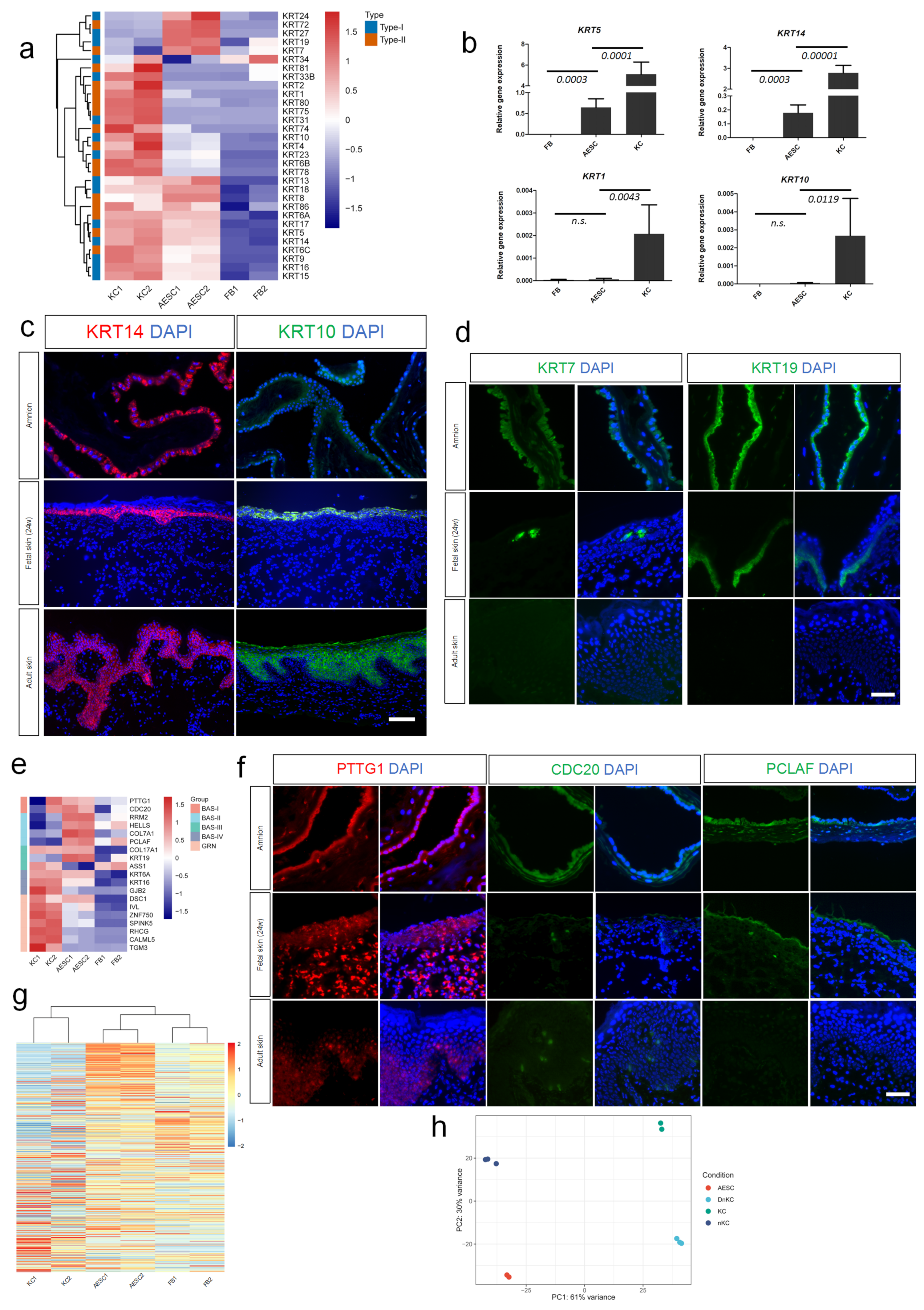
3.2. Similarity of AESCs with Early Stage Immature KCs
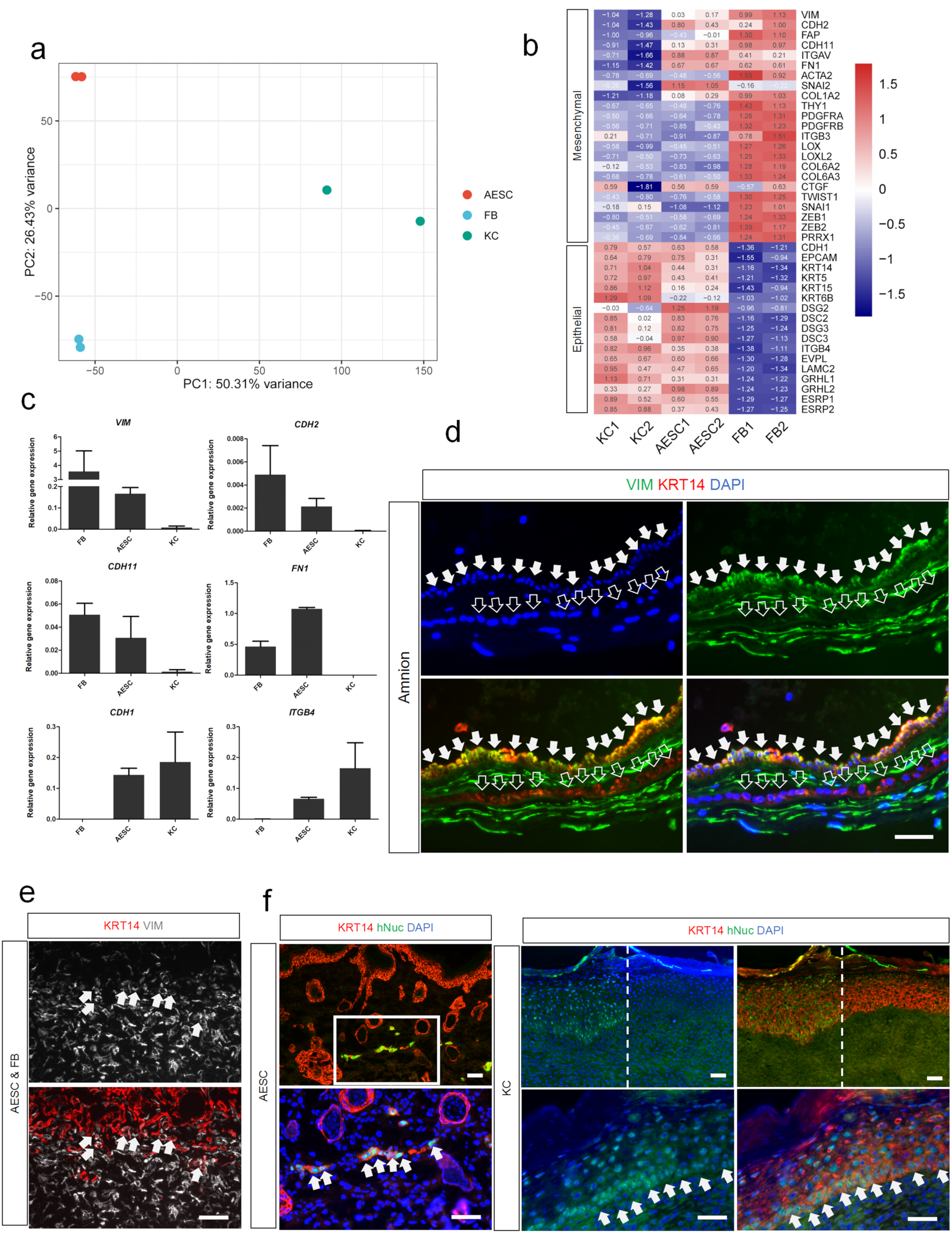
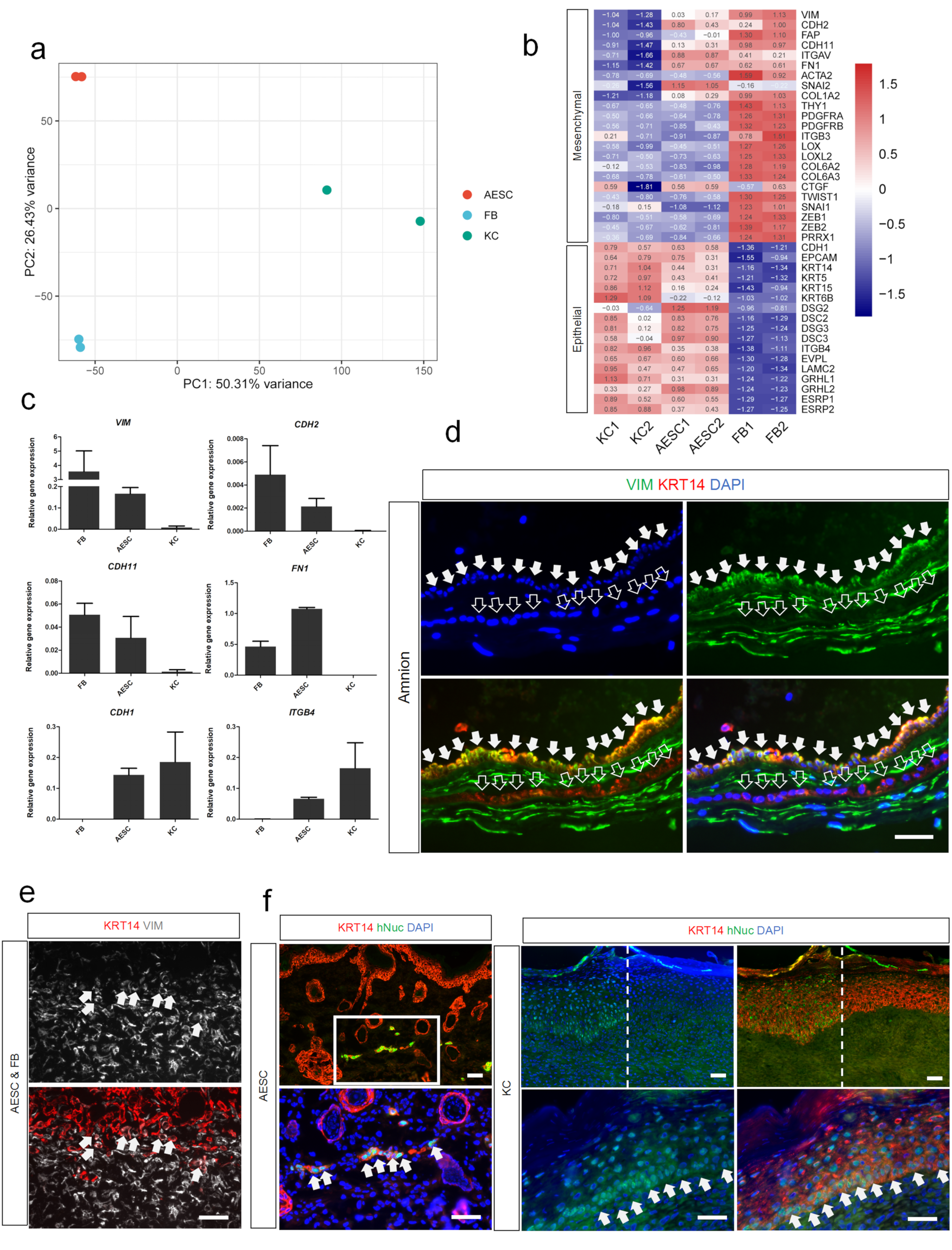
3.3. Differences of AESCs from KCs in Mesenchymal Characteristics
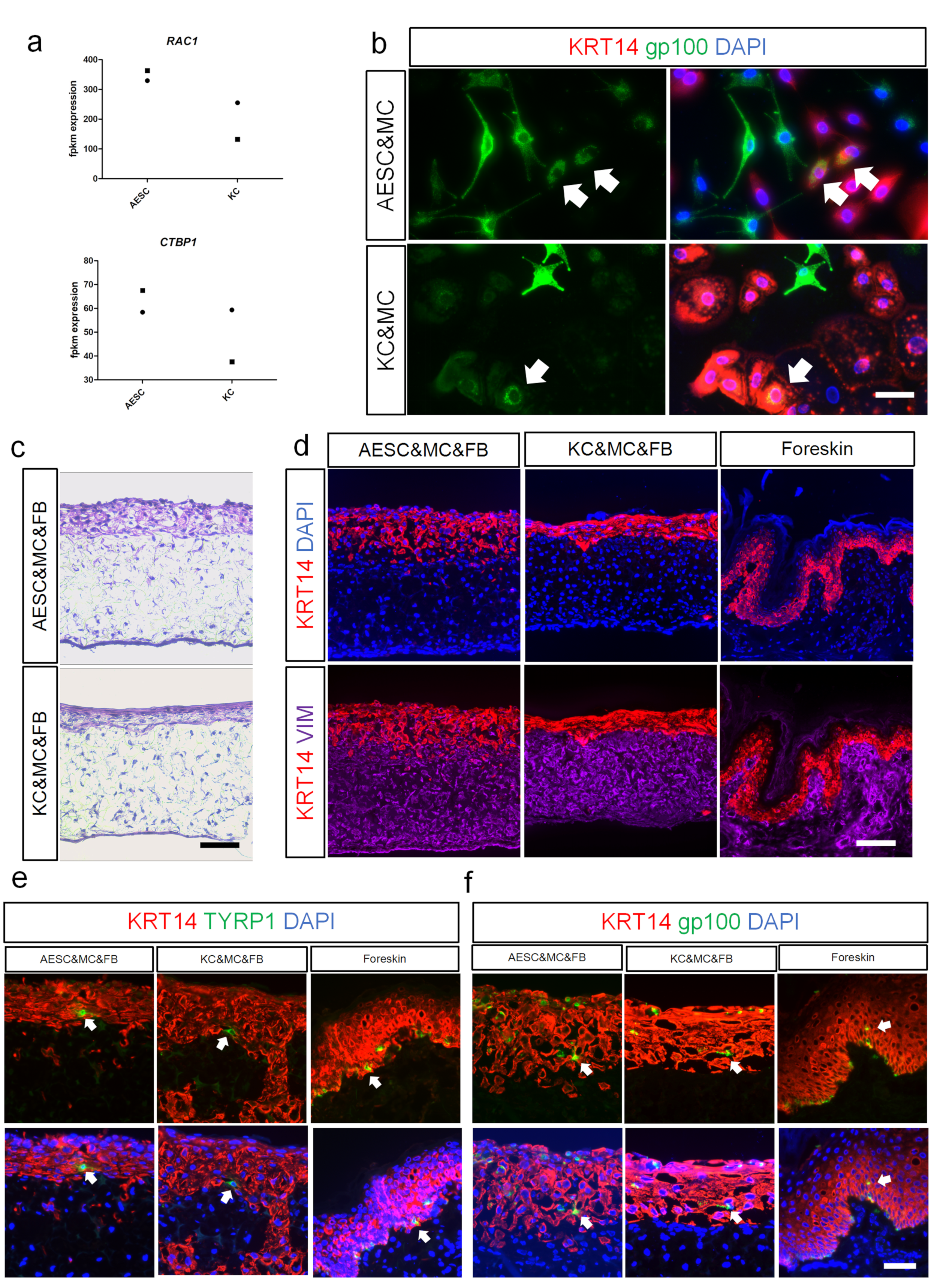
3.4. AESCs Interact with Melanocytes and Phagocytose Melanosomes
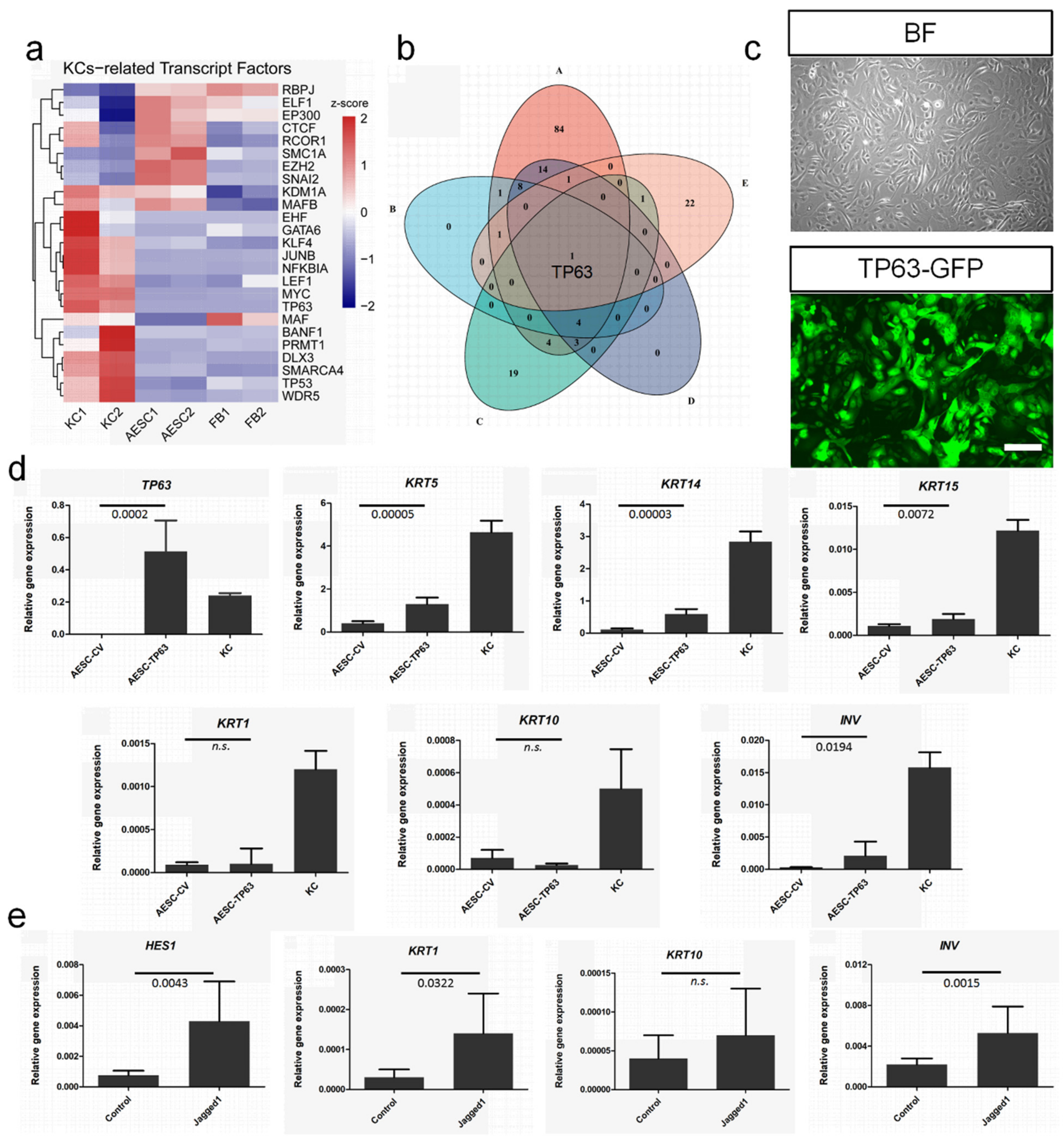
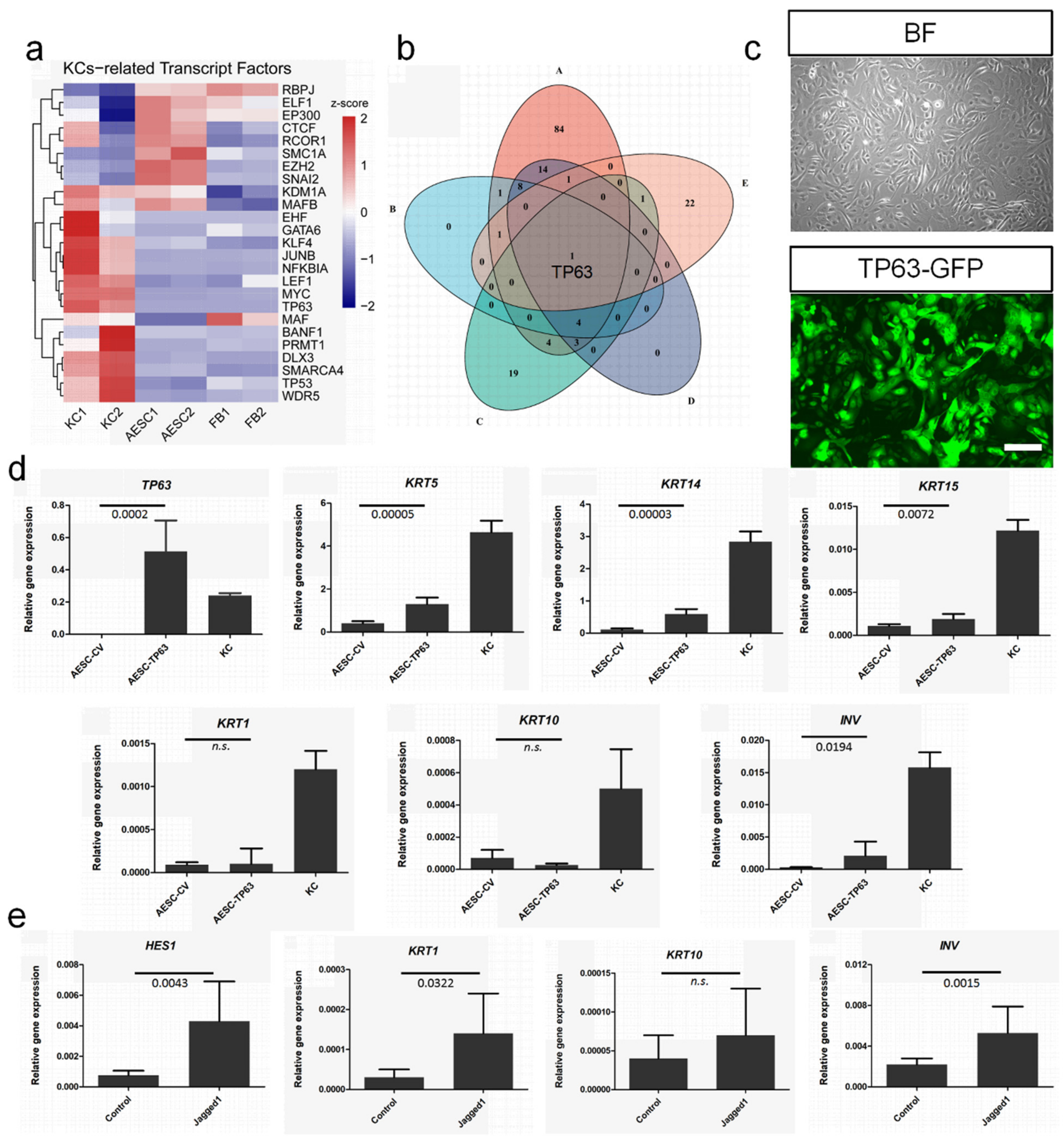
3.5. TP63 Enhancement and NOTCH Activation to Induce Cell Fate Convergence of AESCs to KCs
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hay, R.J.; Johns, N.E.; Williams, H.C.; Bolliger, I.W.; Dellavalle, R.P.; Margolis, D.J.; Marks, R.; Naldi, L.; Weinstock, M.A.; Wulf, S.K.; et al. The global burden of skin disease in 2010: An analysis of the prevalence and impact of skin conditions. J. Investig. Dermatol. 2014, 134, 1527–1534. [Google Scholar] [CrossRef] [Green Version]
- Rahmani Del Bakhshayesh, A.; Annabi, N.; Khalilov, R.; Akbarzadeh, A.; Samiei, M.; Alizadeh, E.; Alizadeh-Ghodsi, M.; Davaran, S.; Montaseri, A. Recent advances on biomedical applications of scaffolds in wound healing and dermal tissue engineering. Artif. Cells Nanomed. Biotechnol. 2018, 46, 691–705. [Google Scholar] [CrossRef] [PubMed]
- Vig, K.; Chaudhari, A.; Tripathi, S.; Dixit, S.; Sahu, R.; Pillai, S.; Dennis, V.A.; Singh, S.R. Advances in skin regeneration using tissue engineering. Int. J. Mol. Sci. 2017, 18, 789. [Google Scholar] [CrossRef] [PubMed]
- Furuya, K.; Zheng, Y.W.; Sako, D.; Iwasaki, K.; Zheng, D.X.; Ge, J.Y.; Liu, L.P.; Furuta, T.; Akimoto, K.; Yagi, H.; et al. Enhanced hepatic differentiation in the subpopulation of human amniotic stem cells under 3D multicellular microenvironment. World J. Stem Cells 2019, 11, 705–721. [Google Scholar] [CrossRef]
- Miki, T.; Lehmann, T.; Cai, H.; Stolz, D.B.; Strom, S.C. Stem cell characteristics of amniotic epithelial cells. Stem Cells 2005, 23, 1549–1559. [Google Scholar] [CrossRef] [Green Version]
- Miki, T.; Marongiu, F.; Dorko, K.; Ellis, E.C.; Strom, S.C. Isolation of amniotic epithelial stem cells. Curr. Protoc. Stem Cell Biol. 2010, 12, 1E. 3.1–1E. 3.10. [Google Scholar] [CrossRef]
- Ilancheran, S.; Michalska, A.; Peh, G.; Wallace, E.M.; Pera, M.; Manuelpillai, U. Stem cells derived from human fetal membranes display multilineage differentiation potential. Biol. Reprod. 2007, 77, 577–588. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hori, J.; Wang, M.; Kamiya, K.; Takahashi, H.; Sakuragawa, N. Immunological characteristics of amniotic epithelium. Cornea 2006, 25, S53–S58. [Google Scholar] [CrossRef]
- Insausti, C.L.; Blanquer, M.; García-Hernandez, A.M.; Castellanos, G.; Moraleda, J.M. Amniotic membrane-derived stem cells: Immunomodulatory properties and potential clinical application. Stem Cells Cloning 2014, 7, 53–63. [Google Scholar] [CrossRef] [Green Version]
- Niknejad, H.; Peirovi, H.; Jorjani, M.; Ahmadiani, A.; Ghanavi, J.; Seifalian, A.M. Properties of the amniotic membrane for potential use in tissue engineering. Eur. Cells Mater. 2008, 15, 88–99. [Google Scholar] [CrossRef]
- Zhang, Q.; Lai, D. Application of human amniotic epithelial cells in regenerative medicine: A systematic review. Stem Cell Res. Ther. 2020, 11, 439. [Google Scholar] [CrossRef]
- Regauer, S.; Franke, W.W.; Virtanen, I. Intermediate filament cytoskeleton of amnion epithelium and cultured amnion epithelial cells: Expression of epidermal cytokeratins in cells of a simple epithelium. J. Cell Biol. 1985, 100, 997–1009. [Google Scholar] [CrossRef] [Green Version]
- Jiang, L.W.; Chen, H.; Lu, H. Using human epithelial amnion cells in human de-epidermized dermis for skin regeneration. J. Dermatol. Sci. 2016, 81, 26–34. [Google Scholar] [CrossRef] [PubMed]
- Fatimah, S.S.; Chua, K.; Tan, G.C.; Azmi, T.I.; Tan, A.E.; Rahman, H.A. Organotypic culture of human amnion cells in air-liquid interface as a potential substitute for skin regeneration. Cytotherapy 2013, 15, 1030–1041. [Google Scholar] [CrossRef]
- Yu, S.C.; Xu, Y.Y.; Li, Y.; Xu, B.; Sun, Q.; Li, F.; Zhang, X.G. Construction of tissue engineered skin with human amniotic mesenchymal stem cells and human amniotic epithelial cells. Eur. Rev. Med. Pharmacol. Sci. 2015, 19, 4627–4635. [Google Scholar]
- Li, H.; Chu, Y.; Zhang, Z.; Zhang, G.; Jiang, L.; Wu, H.; Liu, S.; Yu, C.; Jin, Y. Construction of bilayered tissue-engineered skin with human amniotic mesenchymal cells and human amniotic epithelial cells. Artif. Organs 2012, 36, 911–919. [Google Scholar] [CrossRef] [PubMed]
- Wolf, H.J.; Schmidt, W.; Drenckhahn, D. Immunocytochemical analysis of the cytoskeleton of the human amniotic epithelium. Cell Tissue Res. 1991, 266, 385–389. [Google Scholar] [CrossRef]
- Liu, L.P.; Li, Y.M.; Guo, N.N.; Li, S.; Ma, X.; Zhang, Y.X.; Gao, Y.; Huang, J.L.; Zheng, D.X.; Wang, L.Y.; et al. Therapeutic potential of patient iPSC-derived iMelanocytes in autologous transplantation. Cell Rep. 2019, 27, 455–466.e5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moll, R.; Divo, M.; Langbein, L. The human keratins: Biology and pathology. Histochem. Cell Biol. 2008, 129, 705–733. [Google Scholar] [CrossRef] [Green Version]
- Byrne, C.; Tainsky, M.; Fuchs, E. Programming gene expression in developing epidermis. Development 1994, 120, 2369–2383. [Google Scholar] [CrossRef] [PubMed]
- Tan, K.K.; Salgado, G.; Connolly, J.E.; Chan, J.K.; Lane, E.B. Characterization of fetal keratinocytes, showing enhanced stem cell-like properties: A potential source of cells for skin reconstruction. Stem Cell Rep. 2014, 3, 324–338. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Drummond, M.L.; Guerrero-Juarez, C.F.; Tarapore, E.; MacLean, A.L.; Stabell, A.R.; Wu, S.C.; Gutierrez, G.; That, B.T.; Benavente, C.A.; et al. Single cell transcriptomics of human epidermis identifies basal stem cell transition states. Nat. Commun. 2020, 11, 4239. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Lloyd, S.M.; Kweon, J.; Gamalong, G.M.; Bao, X. Epidermal progenitors suppress GRHL3-mediated differentiation through intronic polyadenylation promoted by CPSF-HNRNPA3 collaboration. Nat. Commun. 2021, 12, 448. [Google Scholar] [CrossRef]
- Pastushenko, I.; Mauri, F.; Song, Y.; de Cock, F.; Meeusen, B.; Swedlund, B.; Impens, F.; Van Haver, D.; Opitz, M.; Thery, M.; et al. Fat1 deletion promotes hybrid EMT state, tumour stemness and metastasis. Nature 2021, 589, 448–455. [Google Scholar] [CrossRef] [PubMed]
- Moreiras, H.; Neto, M.; Bento-Lopes, L.; Escrevente, C.; Ramalho, J.; Seabra, M.; Barral, D. Melanocore uptake by keratinocytes occurs through phagocytosis and involves Protease-activated receptor-2 activation. bioRxiv 2021. [Google Scholar] [CrossRef]
- Singh, S.K.; Nizard, C.; Kurfurst, R.; Bonte, F.; Schnebert, S.; Tobin, D.J. The silver locus product (Silv/gp100/Pmel17) as a new tool for the analysis of melanosome transfer in human melanocyte-keratinocyte co-culture. Exp. Dermatol. 2008, 17, 418–426. [Google Scholar] [CrossRef]
- Ma, H.J.; Zhao, G.; Zi, S.X.; Li, D.G.; Liu, W.; Yang, Q.Q. Efficacy of quantifying melanosome transfer with flow cytometry in a human melanocyte-HaCaT keratinocyte co-culture system in vitro. Exp. Dermatol. 2009, 19, e282–e285. [Google Scholar] [CrossRef] [PubMed]
- Boehnke, K.; Mirancea, N.; Pavesio, A.; Fusenig, N.E.; Boukamp, P.; Stark, H.J. Effects of fibroblasts and microenvironment on epidermal regeneration and tissue function in long-term skin equivalents. Eur. J. Cell Biol. 2007, 86, 731–746. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Mistry, D.S.; Sen, G.L. Highly rapid and efficient conversion of human fibroblasts to keratinocyte-like cells. J. Investig. Dermatol. 2014, 134, 335–344. [Google Scholar] [CrossRef] [Green Version]
- Yang, A.; Schweitzer, R.; Sun, D.; Kaghad, M.; Walker, N.; Bronson, R.T.; Tabin, C.; Sharpe, A.; Caput, D.; Crum, C.; et al. p63 is essential for regenerative proliferation in limb, craniofacial and epithelial development. Nature 1999, 398, 714–718. [Google Scholar] [CrossRef]
- Blanpain, C.; Lowry, W.E.; Pasolli, H.A.; Fuchs, E. Canonical notch signaling functions as a commitment switch in the epidermal lineage. Genes Dev. 2006, 20, 3022–3035. [Google Scholar] [CrossRef] [Green Version]
- Cordle, J.; Johnson, S.; Tay, J.Z.; Roversi, P.; Wilkin, M.B.; de Madrid, B.H.; Shimizu, H.; Jensen, S.; Whiteman, P.; Jin, B.; et al. A conserved face of the Jagged/Serrate DSL domain is involved in Notch trans-activation and cis-inhibition. Nat. Struct. Mol. Biol. 2008, 15, 849–857. [Google Scholar] [CrossRef] [Green Version]
- Artavanis-Tsakonas, S.; Rand, M.D.; Lake, R.J. Notch signaling: Cell fate control and signal integration in development. Science 1999, 284, 770–776. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nyfeler, Y.; Kirch, R.D.; Mantei, N.; Leone, D.P.; Radtke, F.; Suter, U.; Taylor, V. Jagged1 signals in the postnatal subventricular zone are required for neural stem cell self-renewal. EMBO J. 2005, 24, 3504–3515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roobrouck, V.D.; Ulloa-Montoya, F.; Verfaillie, C.M. Self-renewal and differentiation capacity of young and aged stem cells. Exp. Cell Res. 2008, 314, 1937–1944. [Google Scholar] [CrossRef]
- Gaunt, G.; Ramin, K. Immunological tolerance of the human fetus. Am. J. Perinatol. 2001, 18, 299–312. [Google Scholar] [CrossRef] [PubMed]
- Jackson, B.W.; Grund, C.; Schmid, E.; Bürki, K.; Franke, W.W.; Illmensee, K. Formation of cytoskeletal elements during mouse embryogenesis. Intermediate filaments of the cytokeratin type and desmosomes in preimplantation embryos. Differentiation 1980, 17, 161–179. [Google Scholar] [CrossRef]
- Fuchs, E.; Green, H. Changes in keratin gene expression during terminal differentiation of the keratinocyte. Cell 1980, 19, 1033–1042. [Google Scholar] [CrossRef]
- Moll, R.; Dhouailly, D.; Sun, T.T. Expression of keratin 5 as a distinctive feature of epithelial and biphasic mesotheliomas. An immunohistochemical study using monoclonal antibody AE14. Virchows Arch. B Cell Pathol. Incl. Mol. Pathol. 1989, 58, 129–145. [Google Scholar] [CrossRef]
- Bickenbach, J.R.; Greer, J.M.; Bundman, D.S.; Rothnagel, J.A.; Roop, D.K. Loricrin expression is coordinated with other epidermal proteins and the appearance of lipid lamellar granules in development. J. Investig. Dermatol. 1995, 104, 405–410. [Google Scholar] [CrossRef] [Green Version]
- Scheibner, K.; Schirge, S.; Burtscher, I.; Büttner, M.; Sterr, M.; Yang, D.; Böttcher, A.; Ansarullah; Irmler, M.; Beckers, J.; et al. Epithelial cell plasticity drives endoderm formation during gastrulation. Nat. Cell Biol. 2021, 23, 692–703. [Google Scholar] [CrossRef] [PubMed]
- Nieto, M.A.; Huang, R.Y.; Jackson, R.A.; Thiery, J.P. EMT: 2016. Cell 2016, 166, 21–45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abell, A.N.; Jordan, N.V.; Huang, W.; Prat, A.; Midland, A.A.; Johnson, N.L.; Granger, D.A.; Mieczkowski, P.A.; Perou, C.M.; Gomez, S.M.; et al. MAP3K4/CBP-regulated H2B acetylation controls epithelial-mesenchymal transition in trophoblast stem cells. Cell Stem Cell 2011, 8, 525–537. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, M.; Bardia, A.; Wittner, B.S.; Stott, S.L.; Smas, M.E.; Ting, D.T.; Isakoff, S.J.; Ciciliano, J.C.; Wells, M.N.; Shah, A.M.; et al. Circulating breast tumor cells exhibit dynamic changes in epithelial and mesenchymal composition. Science 2013, 339, 580–584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, R.Y.; Wong, M.K.; Tan, T.Z.; Kuay, K.T.; Ng, A.H.; Chung, V.Y.; Chu, Y.S.; Matsumura, N.; Lai, H.C.; Lee, Y.F.; et al. An EMT spectrum defines an anoikis-resistant and spheroidogenic intermediate mesenchymal state that is sensitive to e-cadherin restoration by a src-kinase inhibitor, saracatinib (AZD0530). Cell Death Dis. 2013, 4, e915. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, W.; Kang, Y. Epithelial-mesenchymal plasticity in cancer progression and metastasis. Dev. Cell 2019, 49, 361–374. [Google Scholar] [CrossRef]
- Li, B.; Zheng, Y.W.; Sano, Y.; Taniguchi, H. Evidence for mesenchymal-epithelial transition associated with mouse hepatic stem cell differentiation. PLoS ONE 2011, 6, e17092. [Google Scholar] [CrossRef] [Green Version]
- Richardson, L.S.; Taylor, R.N.; Menon, R. Reversible EMT and MET mediate amnion remodeling during pregnancy and labor. Sci. Signal. 2020, 13, eaay1486. [Google Scholar] [CrossRef]
- Abbasi, S.; Sinha, S.; Labit, E.; Rosin, N.L.; Yoon, G.; Rahmani, W.; Jaffer, A.; Sharma, N.; Hagner, A.; Shah, P.; et al. Distinct regulatory programs control the latent regenerative potential of dermal fibroblasts during wound healing. Cell Stem Cell 2020, 27, 396–412.e6. [Google Scholar] [CrossRef]
- Jiang, D.; Christ, S.; Correa-Gallegos, D.; Ramesh, P.; Kalgudde Gopal, S.; Wannemacher, J.; Mayr, C.H.; Lupperger, V.; Yu, Q.; Ye, H.; et al. Injury triggers fascia fibroblast collective cell migration to drive scar formation through N-cadherin. Nat. Commun. 2020, 11, 5653. [Google Scholar] [CrossRef]
- Sinha, M.; Sen, C.K.; Singh, K.; Das, A.; Ghatak, S.; Rhea, B.; Blackstone, B.; Powell, H.M.; Khanna, S.; Roy, S. Direct conversion of injury-site myeloid cells to fibroblast-like cells of granulation tissue. Nat. Commun. 2018, 9, 936. [Google Scholar] [CrossRef] [Green Version]
- Iacovides, D.; Rizki, G.; Lapathitis, G.; Strati, K. Direct conversion of mouse embryonic fibroblasts into functional keratinocytes through transient expression of pluripotency-related genes. Stem Cell Res. Ther. 2016, 7, 98. [Google Scholar] [CrossRef] [Green Version]
- Xie, F.; Tang, X.; Zhang, Q.; Deng, C. Reprogramming human adipose tissue stem cells using epidermal keratinocyte extracts. Mol. Med. Rep. 2015, 11, 182–188. [Google Scholar] [CrossRef] [Green Version]
- Kurita, M.; Araoka, T.; Hishida, T.; O’Keefe, D.D.; Takahashi, Y.; Sakamoto, A.; Sakurai, M.; Suzuki, K.; Wu, J.; Yamamoto, M.; et al. In vivo reprogramming of wound-resident cells generates skin epithelial tissue. Nature 2018, 561, 243–247. [Google Scholar] [CrossRef] [PubMed]
- Koster, M.I. p63 in skin development and ectodermal dysplasias. J. Investig. Dermatol. 2010, 130, 2352–2358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, B.C.; Lefort, K.; Mandinova, A.; Antonini, D.; Devgan, V.; Della Gatta, G.; Koster, M.I.; Zhang, Z.; Wang, J.; Tommasi di Vignano, A.; et al. Cross-regulation between Notch and p63 in keratinocyte commitment to differentiation. Genes Dev. 2006, 20, 1028–1042. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rangarajan, A.; Talora, C.; Okuyama, R.; Nicolas, M.; Mammucari, C.; Oh, H.; Aster, J.C.; Krishna, S.; Metzger, D.; Chambon, P.; et al. Notch signaling is a direct determinant of keratinocyte growth arrest and entry into differentiation. EMBO J. 2001, 20, 3427–3436. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Category | Description | AESCs-Up Gene Ratio | KCs-Up Gene Ratio |
|---|---|---|---|
| BP | Epidermis development | 100/1644 | 160/2562 |
| Cell junction organization | 62/1644 | 69/2562 | |
| Morphogenesis of epithelium | 73/1644 | 96/2562 | |
| Regulation of epidermis development | 20/1644 | 38/2562 | |
| Molting cycle | 23/1644 | 33/2562 | |
| Hair cycle | 23/1644 | 33/2562 | |
| In utero embryonic development | 46/1644 | 72/2562 | |
| CC | Cell–cell junction | 93/1732 | 110/2666 |
| Cornified envelope | 25/1732 | 42/2666 | |
| Cell–cell adherens junction | 30/1732 | 32/2666 | |
| Cell cortex | 52/1732 | 65/2666 | |
| MF | Cadherin binding | 75/1659 | 112/2588 |
| Rho GTPase binding | 38/1659 | 46/2588 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, L.-P.; Zheng, D.-X.; Xu, Z.-F.; Zhou, H.-C.; Wang, Y.-C.; Zhou, H.; Ge, J.-Y.; Sako, D.; Li, M.; Akimoto, K.; et al. Transcriptomic and Functional Evidence Show Similarities between Human Amniotic Epithelial Stem Cells and Keratinocytes. Cells 2022, 11, 70. https://doi.org/10.3390/cells11010070
Liu L-P, Zheng D-X, Xu Z-F, Zhou H-C, Wang Y-C, Zhou H, Ge J-Y, Sako D, Li M, Akimoto K, et al. Transcriptomic and Functional Evidence Show Similarities between Human Amniotic Epithelial Stem Cells and Keratinocytes. Cells. 2022; 11(1):70. https://doi.org/10.3390/cells11010070
Chicago/Turabian StyleLiu, Li-Ping, Dong-Xu Zheng, Zheng-Fang Xu, Hu-Cheng Zhou, Yun-Cong Wang, Hang Zhou, Jian-Yun Ge, Daisuke Sako, Mi Li, Kazunori Akimoto, and et al. 2022. "Transcriptomic and Functional Evidence Show Similarities between Human Amniotic Epithelial Stem Cells and Keratinocytes" Cells 11, no. 1: 70. https://doi.org/10.3390/cells11010070
APA StyleLiu, L.-P., Zheng, D.-X., Xu, Z.-F., Zhou, H.-C., Wang, Y.-C., Zhou, H., Ge, J.-Y., Sako, D., Li, M., Akimoto, K., Li, Y.-M., & Zheng, Y.-W. (2022). Transcriptomic and Functional Evidence Show Similarities between Human Amniotic Epithelial Stem Cells and Keratinocytes. Cells, 11(1), 70. https://doi.org/10.3390/cells11010070