
Altered Ca2+ Homeostasis in Red Blood Cells of Polycythemia Vera Patients Following Disturbed Organelle Sorting during Terminal Erythropoiesis

 , , , ,
, , , ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Human Blood Samples
2.2. Mice Samples
2.3. Flow Cytometry
2.4. Membrane Potential Estimation
2.5. RBC Volume Assays
2.6. Automated Patch-Clamp
2.7. Imaging Flow Cytometry
2.8. Sample Preparation for Tandem Mass Spectrometry (MS/MS) Analysis
2.9. Nanoscale Liquid Chromatography Coupled to Tandem Mass Spectrometry Analysis
2.10. Analyses of MS/MS Data
2.11. SDS-PAGE Immunoblots
2.12. Statistical Analysis
3. Results
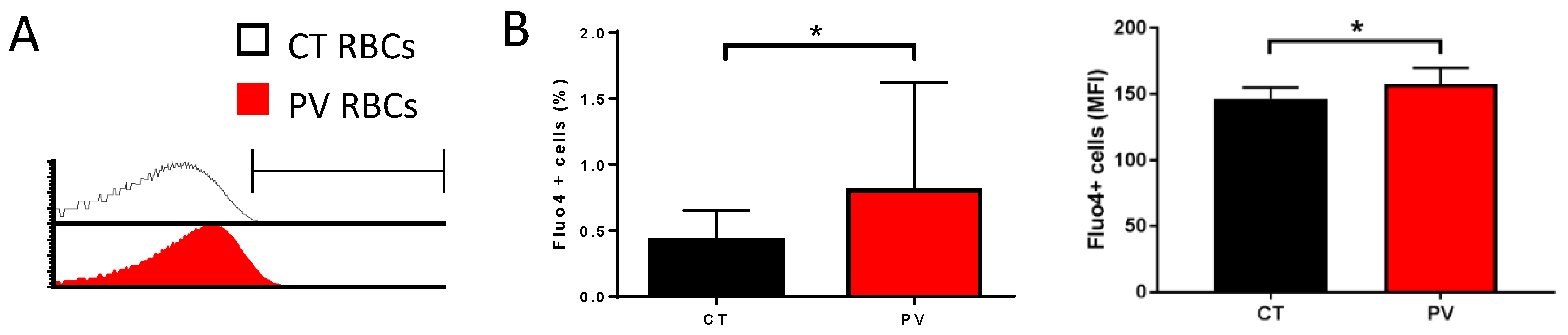
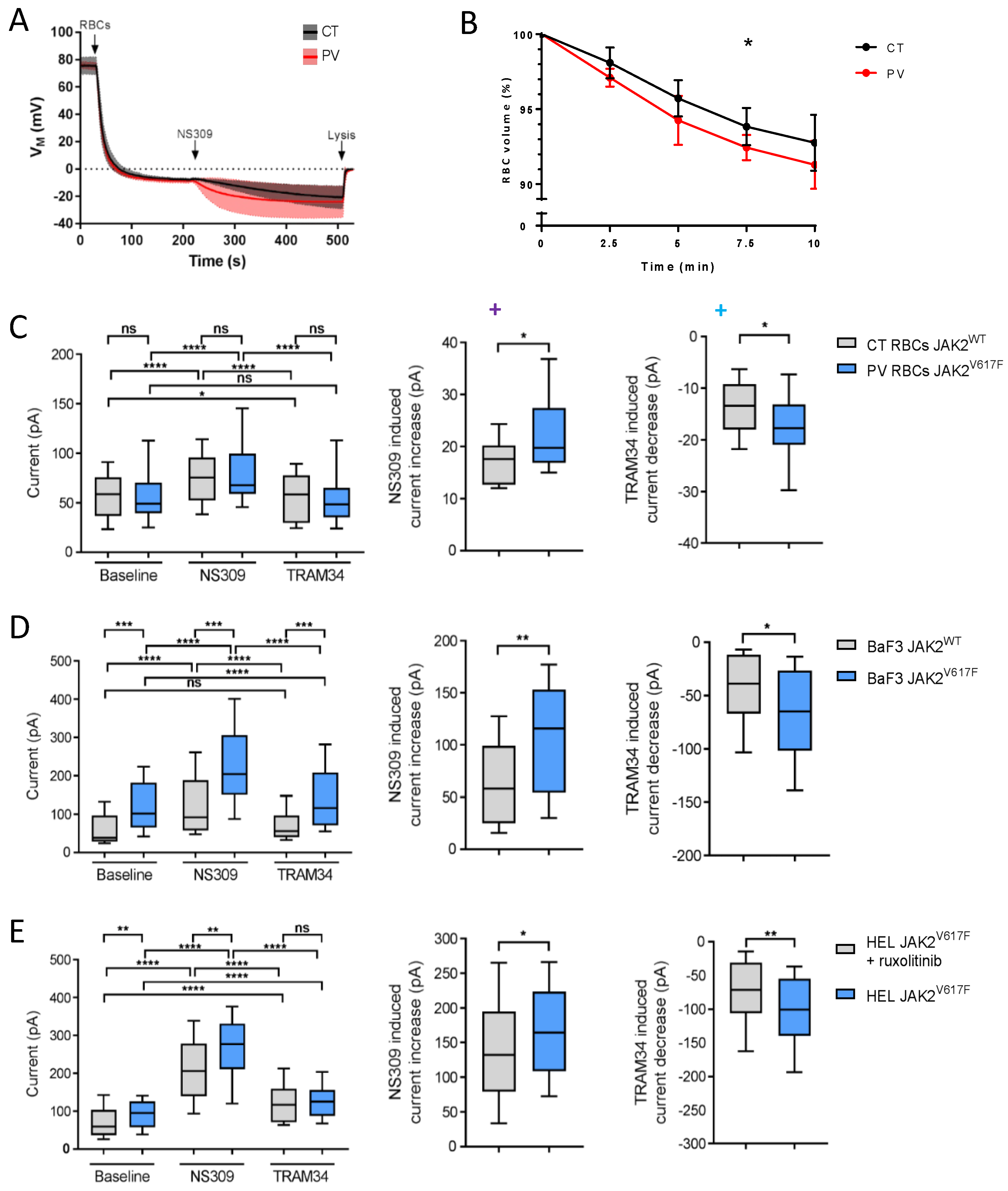
3.1. PV RBCs Show Elevated Free Intracellular Ca2+ Levels and Increased Gárdos Channel Activity
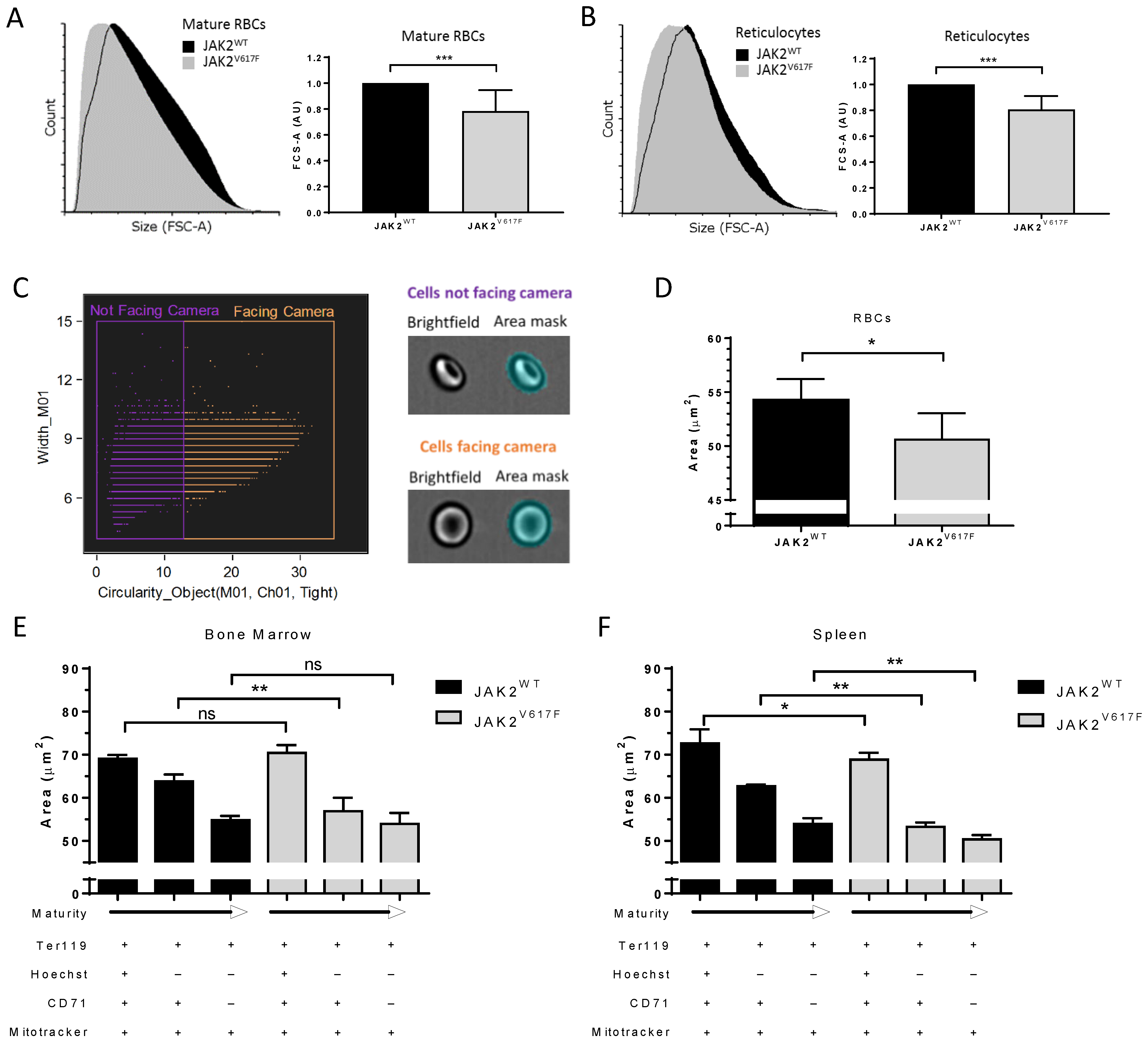
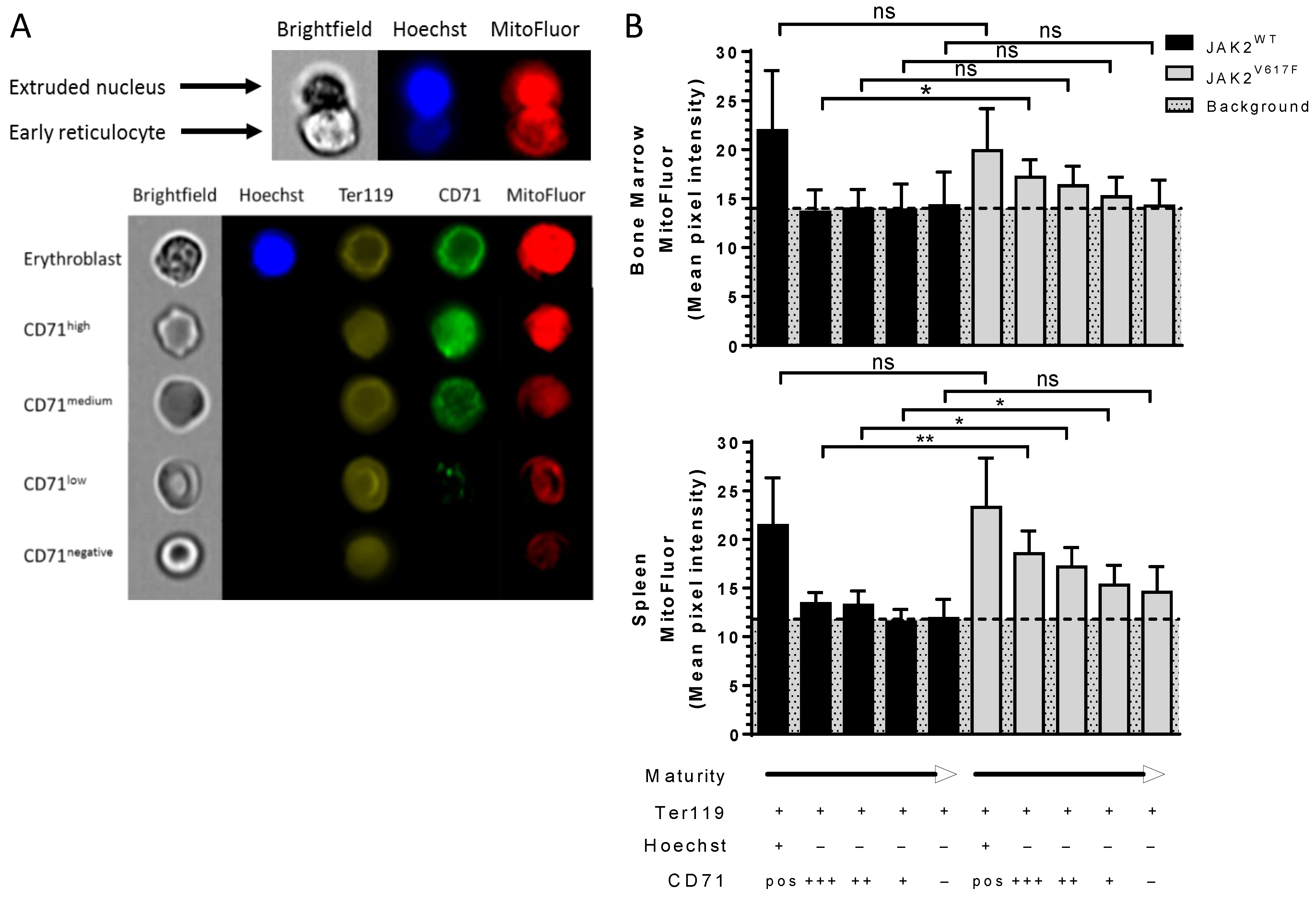
3.2. JAK2V617F Is Associated with Decreased Erythroid Cell Size during Mouse In Vivo Erythropoiesis
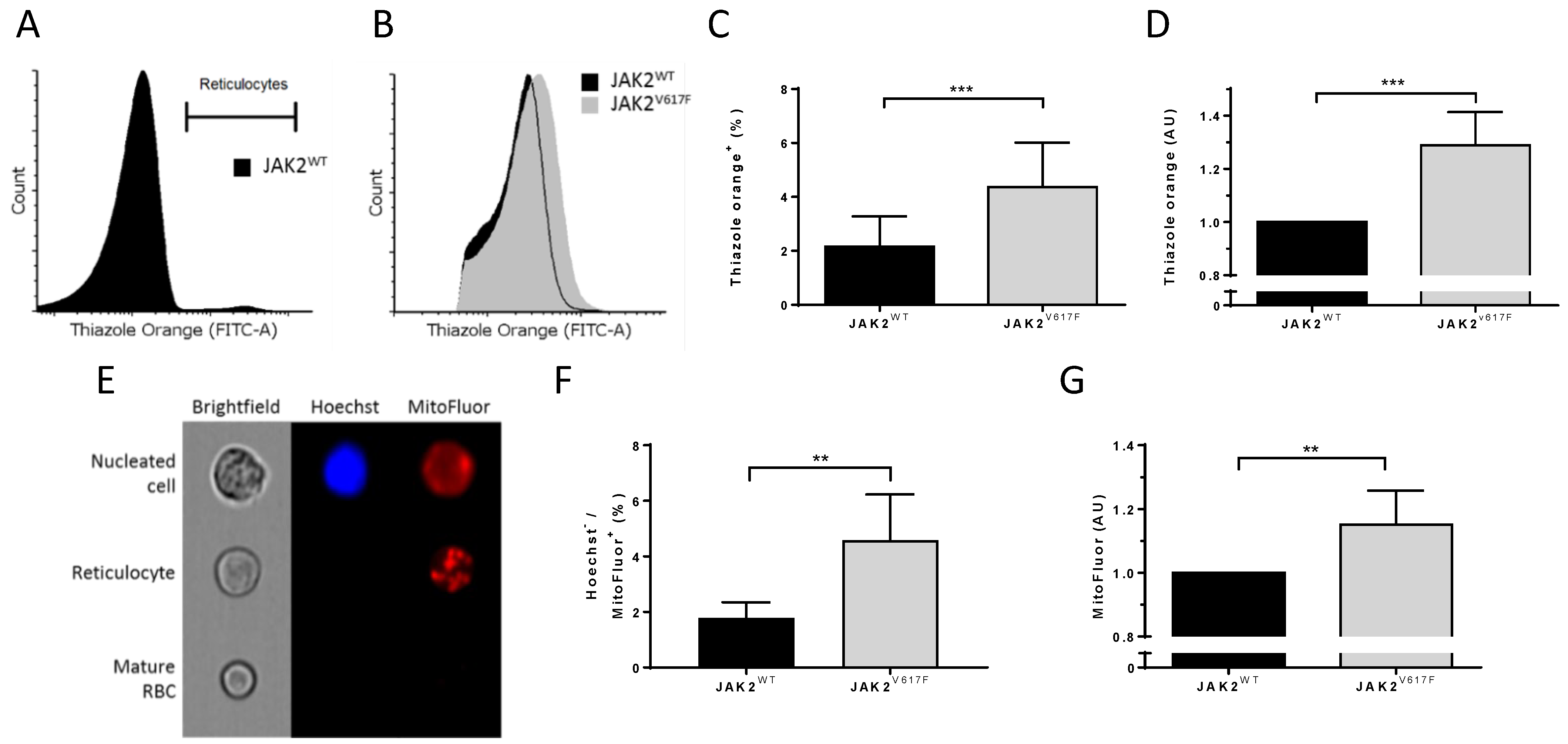
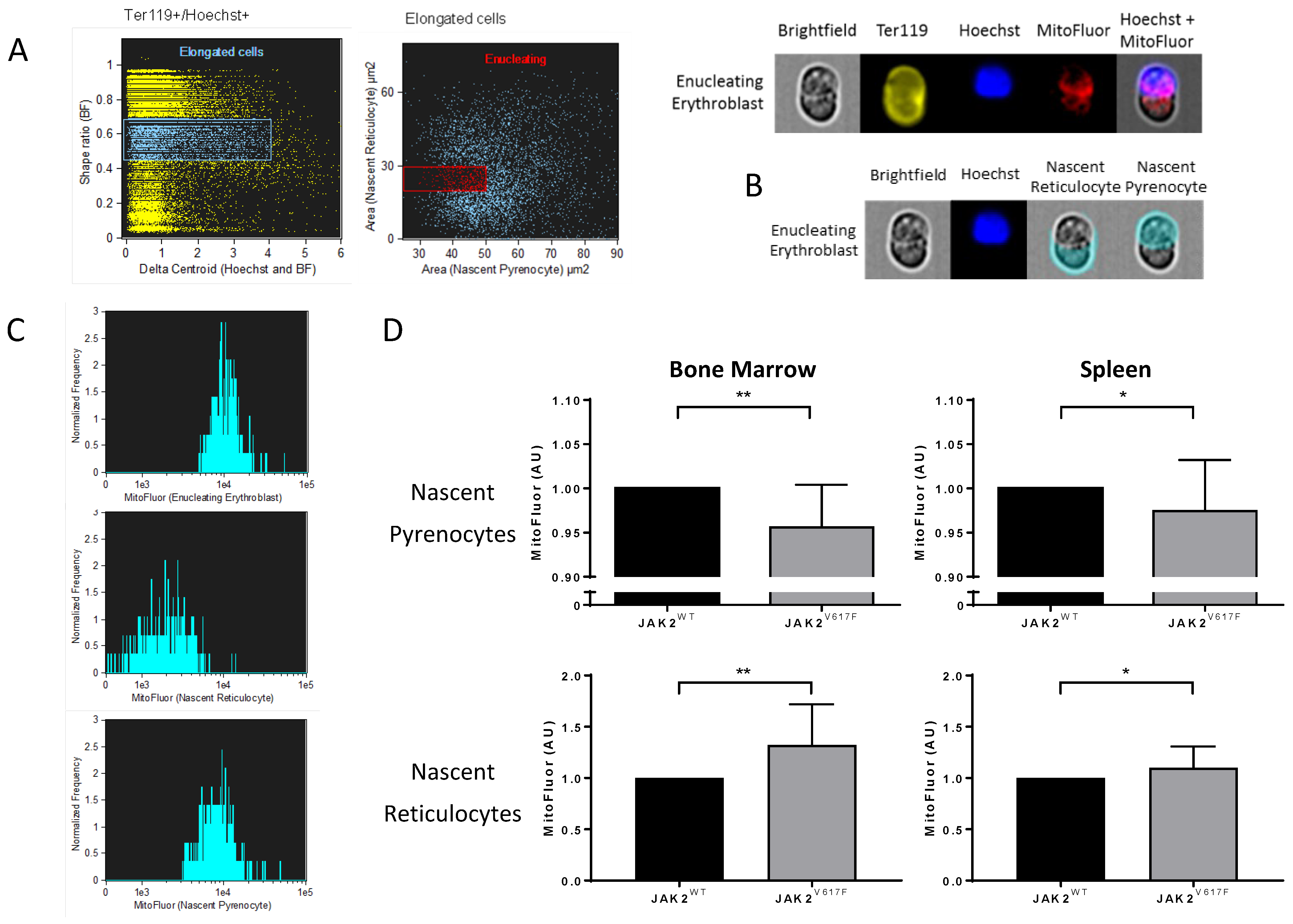
3.3. JAK2V617F Is Associated with Higher Organelle Remnants in Circulating Mouse Reticulocytes
3.4. Altered Organelle Sorting during Enucleation of JAK2V617F Mouse Erythroblasts
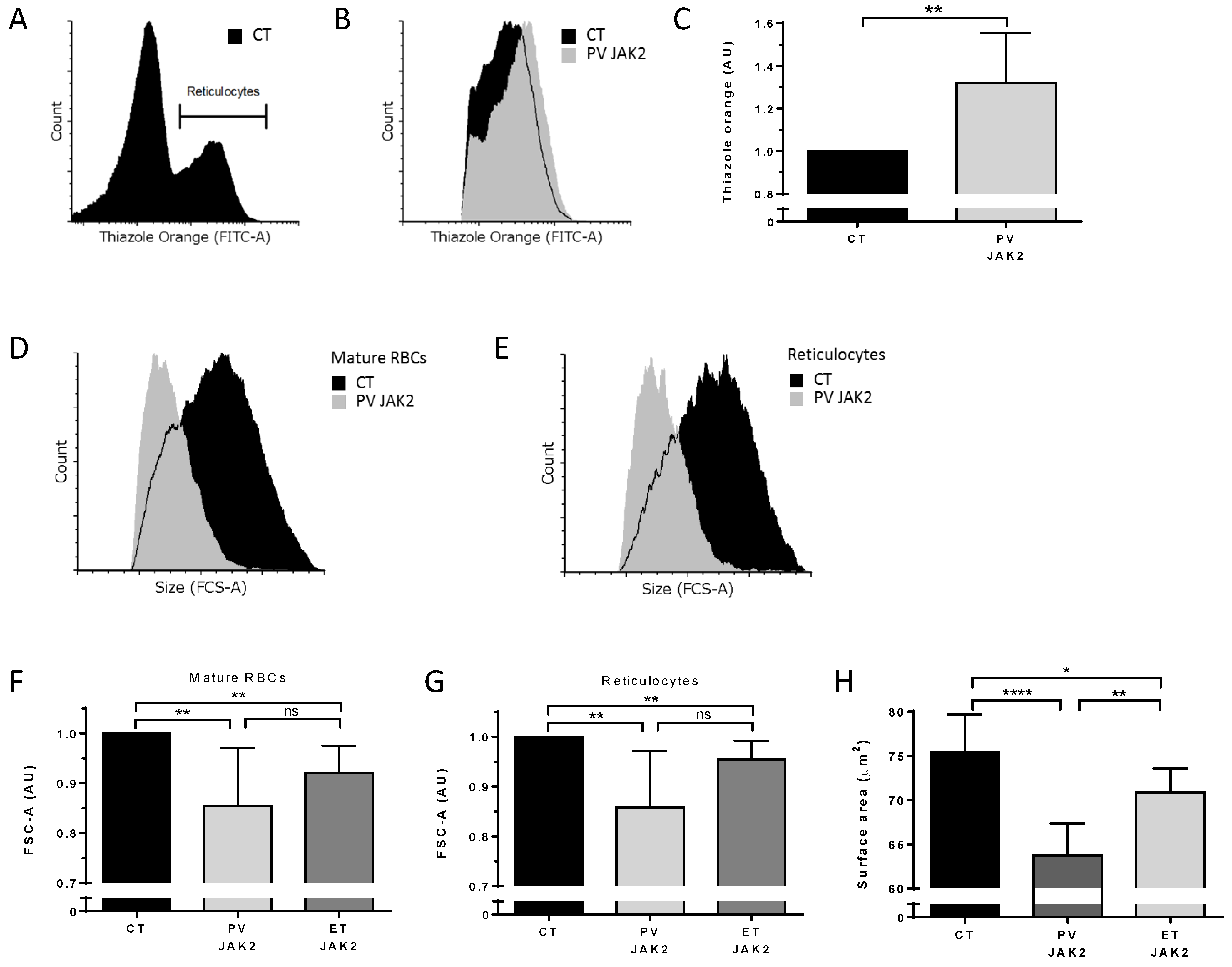
3.5. Higher Ribosomal Content and Smaller Red Cell Size in JAK2V617F PV and ET Patients
3.6. Modified Whole Cell Proteome in Circulating Reticulocytes and Mature RBCs from PV and ET Patients with JAK2V617F
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nangalia, J.; Grinfeld, J.; Green, A.R. Pathogenesis of Myeloproliferative Disorders. Annu. Rev. Pathol. 2016, 11, 101–126. [Google Scholar] [CrossRef] [PubMed]
- Barbui, T.; Finazzi, G.; Falanga, A. Myeloproliferative neoplasms and thrombosis. Blood 2013, 122, 2176–2184. [Google Scholar] [CrossRef] [PubMed]
- Marchioli, R.; Finazzi, G.; Landolfi, R.; Kutti, J.; Gisslinger, H.; Patrono, C.; Marilus, R.; Villegas, A.; Tognoni, G.; Barbui, T. Vascular and neoplastic risk in a large cohort of patients with polycythemia vera. J. Clin. Oncol. 2005, 23, 2224–2232. [Google Scholar] [CrossRef] [PubMed]
- Ji, P. New insights into the mechanisms of mammalian erythroid chromatin condensation and enucleation. Int. Rev. Cell. Mol. Biol. 2015, 316, 159–182. [Google Scholar] [CrossRef]
- Brusson, M.; Cochet, S.; Leduc, M.; Guillonneau, F.; Mayeux, P.; Peyrard, T.; Chomienne, C.; Le Van Kim, C.; Cassinat, B.; Kiladjian, J.-J.; et al. Enhanced calreticulin expression in red cells of polycythemia vera patients harboring the JAK2V617F mutation. Haematologica 2017, 102, e241–e244. [Google Scholar] [CrossRef] [PubMed]
- Bogdanova, A.; Makhro, A.; Wang, J.; Lipp, P.; Kaestner, L. Calcium in red blood cells-a perilous balance. Int. J. Mol. Sci. 2013, 14, 9848–9872. [Google Scholar] [CrossRef] [PubMed]
- Gautier, E.F.; Leduc, M.; Cochet, S.; Bailly, K.; Lacombe, C.; Mohandas, N.; Guillonneau, F.; El Nemer, W.; Mayeux, P. Absolute proteome quantification of highly purified populations of circulating reticulocytes and mature erythrocytes. Blood Adv. 2018, 2, 2646–2657. [Google Scholar] [CrossRef]
- Hasan, S.; Lacout, C.; Marty, C.; Cuingnet, M.; Solary, E.; Vainchenker, W.; Villeval, J.-L. JAK2V617F expression in mice amplifies early hematopoietic cells and gives them a competitive advantage that is hampered by IFNα. Blood 2013, 122, 1464–1477. [Google Scholar] [CrossRef]
- Croker, B.A.; Metcalf, D.; Robb, L.; Wei, W.; Mifsud, S.; DiRago, L.; Cluse, L.A.; Sutherland, K.D.; Hartley, L.; Williams, E.; et al. SOCS3 is a critical physiological negative regulator of G-CSF signaling and emergency granulopoiesis. Immunity 2004, 20, 153–165. [Google Scholar] [CrossRef]
- Almarza, E.; Segovia, J.C.; Guenechea, G.; Gomez, S.G.; Ramirez, A.; Bueren, J.A. Regulatory elements of the vav gene drive transgene expression in hematopoietic stem cells from adult mice. Exp. Hematol. 2004, 32, 360–364. [Google Scholar] [CrossRef] [PubMed]
- Georgiades, P.; Ogilvy, S.; Duval, H.; Licence, D.R.; Charnock-Jones, D.S.; Smith, S.K.; Print, C.G. VavCre transgenic mice: A tool for mutagenesis in hematopoietic and endothelial lineages. Genesis 2002, 34, 251–256. [Google Scholar] [CrossRef]
- Macey, R.I.; Adorante, J.S.; Orme, F.W. Erythrocyte membrane potentials determined by hydrogen ion distribution. Biochim. Biophys. Acta 1978, 512, 284–295. [Google Scholar] [CrossRef]
- Fermo, E.; Monedero-Alonso, D.; Petkova-Kirova, P.; Makhro, A.; Peres, L.; Bouyer, G.; Marcello, A.P.; Longo, F.; Graziadei, G.; Barcellini, W.; et al. Gardos channelopathy: Functional analysis of a novel KCNN4 variant. Blood Adv. 2020, 4, 6336–6341. [Google Scholar] [CrossRef]
- Rotordam, M.G.; Fermo, E.; Becker, N.; Barcellini, W.; Bruggemann, A.; Fertig, N.; Egee, S.; Rapedius, M.; Bianchi, P.; Kaestner, L. A novel gain-of-function mutation of Piezo1 is functionally affirmed in red blood cells by high-throughput patch clamp. Haematologica 2019, 104, e179–e183. [Google Scholar] [CrossRef]
- Monedero Alonso, D.; Peres, L.; Hatem, A.; Bouyer, G.; Egee, S. The Chloride Conductance Inhibitor NS3623 Enhances the Activity of a Non-selective Cation Channel in Hyperpolarizing Conditions. Front. Physiol. 2021, 12, 743094. [Google Scholar] [CrossRef] [PubMed]
- Kulak, N.A.; Pichler, G.; Paron, I.; Nagaraj, N.; Mann, M. Minimal, encapsulated proteomic-sample processing applied to copy-number estimation in eukaryotic cells. Nat. Methods 2014, 11, 319–324. [Google Scholar] [CrossRef]
- Cox, J.; Matic, I.; Hilger, M.; Nagaraj, N.; Selbach, M.; Olsen, J.V.; Mann, M. A practical guide to the MaxQuant computational platform for SILAC-based quantitative proteomics. Nat Protoc. 2009, 4, 698–705. [Google Scholar] [CrossRef] [PubMed]
- Tyanova, S.; Temu, T.; Sinitcyn, P.; Carlson, A.; Hein, M.Y.; Geiger, T.; Mann, M.; Cox, J. The Perseus computational platform for comprehensive analysis of (prote)omics data. Nat. Methods 2016, 13, 731–740. [Google Scholar] [CrossRef] [PubMed]
- Bessman, D. Erythropoiesis during recovery from iron deficiency: Normocytes and macrocytes. Blood 1977, 50, 987–993. [Google Scholar] [CrossRef] [PubMed]
- Spangler, R.; Sytkowski, A.J. c-myc is an erythropoietin early response gene in normal erythroid cells: Evidence for a protein kinase C-mediated signal. Blood 1992, 79, 52–57. [Google Scholar] [CrossRef]
- Spotts, G.D.; Hann, S.R. Enhanced translation and increased turnover of c-myc proteins occur during differentiation of murine erythroleukemia cells. Mol. Cell. Biol. 1990, 10, 3952–3964. [Google Scholar] [CrossRef] [PubMed]
- Jayapal, S.R.; Lee, K.L.; Ji, P.; Kaldis, P.; Lim, B.; Lodish, H.F. Down-regulation of Myc is essential for terminal erythroid maturation. J. Biol. Chem. 2010, 285, 40252–40265. [Google Scholar] [CrossRef]
- Xie, S.; Lin, H.; Sun, T.; Arlinghaus, R.B. Jak2 is involved in c-Myc induction by Bcr-Abl. Oncogene 2002, 21, 7137–7146. [Google Scholar] [CrossRef][Green Version]
- Thom, C.S.; Traxler, E.A.; Khandros, E.; Nickas, J.M.; Zhou, O.Y.; Lazarus, J.E.; Silva, A.P.; Prabhu, D.; Yao, Y.; Aribeana, C.; et al. Trim58 degrades Dynein and regulates terminal erythropoiesis. Dev. Cell. 2014, 30, 688–700. [Google Scholar] [CrossRef]
- Kobayashi, I.; Ubukawa, K.; Sugawara, K.; Asanuma, K.; Guo, Y.M.; Yamashita, J.; Takahashi, N.; Sawada, K.; Nunomura, W. Erythroblast enucleation is a dynein-dependent process. Exp. Hematol. 2016, 44, 247–256.e212. [Google Scholar] [CrossRef] [PubMed]
- Socoro-Yuste, N.; Dagher, M.C.; Gonzalez De Peredo, A.; Mondet, J.; Zaccaria, A.; Roux Dalvai, F.; Plo, I.; Cahn, J.Y.; Mossuz, P. Ph(-) myeloproliferative neoplasm red blood cells display deregulation of IQGAP1-Rho GTPase signaling depending on CALR/JAK2 status. Biochim. Biophys. Acta 2016, 1863, 2758–2765. [Google Scholar] [CrossRef]
- Ji, P.; Jayapal, S.R.; Lodish, H.F. Enucleation of cultured mouse fetal erythroblasts requires Rac GTPases and mDia2. Nat. Cell. Biol. 2008, 10, 314–321. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, S.; De Zan, T.; Ishizaki, T.; Yasuda, S.; Kamijo, H.; Yamada, D.; Aoki, T.; Kiyonari, H.; Kaneko, H.; Shimizu, R.; et al. Loss of a Rho-regulated actin nucleator, mDia2, impairs cytokinesis during mouse fetal erythropoiesis. Cell Rep. 2013, 5, 926–932. [Google Scholar] [CrossRef]
- Socoro-Yuste, N.; Cokic, V.P.; Mondet, J.; Plo, I.; Mossuz, P. Quantitative Proteome Heterogeneity in Myeloproliferative Neoplasm Subtypes and Association with JAK2 Mutation Status. Mol. Cancer Res. 2017, 15, 852–861. [Google Scholar] [CrossRef]
- Ubukawa, K.; Guo, Y.M.; Takahashi, M.; Hirokawa, M.; Michishita, Y.; Nara, M.; Tagawa, H.; Takahashi, N.; Komatsuda, A.; Nunomura, W.; et al. Enucleation of human erythroblasts involves non-muscle myosin IIB. Blood 2012, 119, 1036–1044. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Ramirez, T.; Ji, P.; Jayapal, S.R.; Lodish, H.F.; Murata-Hori, M. Mammalian erythroblast enucleation requires PI3K-dependent cell polarization. J. Cell Sci. 2012, 125, 340–349. [Google Scholar] [CrossRef] [PubMed]
- Holaska, J.M.; Black, B.E.; Love, D.C.; Hanover, J.A.; Leszyk, J.; Paschal, B.M. Calreticulin Is a receptor for nuclear export. J. Cell Biol. 2001, 152, 127–140. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Prak, L.; Rayon-Estrada, V.; Thiru, P.; Flygare, J.; Lim, B.; Lodish, H.F. ZFP36L2 is required for self-renewal of early burst-forming unit erythroid progenitors. Nature 2013, 499, 92–96. [Google Scholar] [CrossRef]
- Falchi, M.; Varricchio, L.; Martelli, F.; Marra, M.; Picconi, O.; Tafuri, A.; Girelli, G.; Uversky, V.N.; Migliaccio, A.R. The Calreticulin control of human stress erythropoiesis is impaired by JAK2V617F in polycythemia vera. Exp. Hematol. 2017, 50, 53–76. [Google Scholar] [CrossRef]
- Liu, Y.; Bao, H.; Mei, Y.; Han, X.; Ji, P. Formin Links Erythroid Cytoskeleton to Organelle Clearance through Escrt-III Complex during Reticulocyte Maturation. Blood 2019, 134, 151. [Google Scholar] [CrossRef]
- Goerttler, P.S.; Kreutz, C.; Donauer, J.; Faller, D.; Maiwald, T.; Marz, E.; Rumberger, B.; Sparna, T.; Schmitt-Graff, A.; Wilpert, J.; et al. Gene expression profiling in polycythaemia vera: Overexpression of transcription factor NF-E2. Br. J. Haematol. 2005, 129, 138–150. [Google Scholar] [CrossRef] [PubMed]
- Gothwal, M.; Wehrle, J.; Aumann, K.; Zimmermann, V.; Grunder, A.; Pahl, H.L. A novel role for nuclear factor-erythroid 2 in erythroid maturation by modulation of mitochondrial autophagy. Haematologica 2016, 101, 1054–1064. [Google Scholar] [CrossRef]
- Moura, P.L.; Hawley, B.R.; Mankelow, T.J.; Griffiths, R.E.; Dobbe, J.G.G.; Streekstra, G.J.; Anstee, D.J.; Satchwell, T.J.; Toye, A.M. Non-muscle myosin II drives vesicle loss during human reticulocyte maturation. Haematologica 2018, 103, 1997–2007. [Google Scholar] [CrossRef] [PubMed]
- Lithanatudom, P.; Wannatung, T.; Leecharoenkiat, A.; Svasti, S.; Fucharoen, S.; Smith, D.R. Enhanced activation of autophagy in beta-thalassemia/Hb E erythroblasts during erythropoiesis. Ann. Hematol. 2011, 90, 747–758. [Google Scholar] [CrossRef] [PubMed]
- Khandros, E.; Thom, C.S.; D’Souza, J.; Weiss, M.J. Integrated protein quality-control pathways regulate free alpha-globin in murine beta-thalassemia. Blood 2012, 119, 5265–5275. [Google Scholar] [CrossRef] [PubMed]
- Butthep, P.; Wisedpanichkij, R.; Jindadamrongwech, S.; Fucharoen, S. Elevated erythropoietin and cytokines levels are related to impaired reticulocyte maturation in thalassemic patients. Blood Cells Mol. Dis. 2015, 54, 170–176. [Google Scholar] [CrossRef] [PubMed]
- De Franceschi, L. Pathophisiology of sickle cell disease and new drugs for the treatment. Mediterr. J. Hematol. Infect. Dis. 2009, 1, e2009024. [Google Scholar] [CrossRef] [PubMed]
- Carayon, K.; Chaoui, K.; Ronzier, E.; Lazar, I.; Bertrand-Michel, J.; Roques, V.; Balor, S.; Terce, F.; Lopez, A.; Salome, L.; et al. Proteolipidic composition of exosomes changes during reticulocyte maturation. J. Biol. Chem. 2011, 286, 34426–34439. [Google Scholar] [CrossRef] [PubMed]
- Franco, S.S.; De Falco, L.; Ghaffari, S.; Brugnara, C.; Sinclair, D.A.; Matte, A.; Iolascon, A.; Mohandas, N.; Bertoldi, M.; An, X.; et al. Resveratrol accelerates erythroid maturation by activation of FoxO3 and ameliorates anemia in beta-thalassemic mice. Haematologica 2014, 99, 267–275. [Google Scholar] [CrossRef]
- Jagadeeswaran, R.; Vazquez, B.A.; Thiruppathi, M.; Ganesh, B.B.; Ibanez, V.; Cui, S.; Engel, J.D.; Diamond, A.M.; Molokie, R.E.; DeSimone, J.; et al. Pharmacological inhibition of LSD1 and mTOR reduces mitochondrial retention and associated ROS levels in the red blood cells of sickle cell disease. Exp. Hematol. 2017, 50, 46–52. [Google Scholar] [CrossRef]
- Andrews, D.A.; Yang, L.; Low, P.S. Phorbol ester stimulates a protein kinase C-mediated agatoxin-TK-sensitive calcium permeability pathway in human red blood cells. Blood 2002, 100, 3392–3399. [Google Scholar] [CrossRef] [PubMed]
- Foller, M.; Kasinathan, R.S.; Koka, S.; Lang, C.; Shumilina, E.; Birnbaumer, L.; Lang, F.; Huber, S.M. TRPC6 contributes to the Ca(2+) leak of human erythrocytes. Cell. Physiol. Biochem. 2008, 21, 183–192. [Google Scholar] [CrossRef] [PubMed]
- Zarychanski, R.; Schulz, V.P.; Houston, B.L.; Maksimova, Y.; Houston, D.S.; Smith, B.; Rinehart, J.; Gallagher, P.G. Mutations in the mechanotransduction protein PIEZO1 are associated with hereditary xerocytosis. Blood 2012, 120, 1908–1915. [Google Scholar] [CrossRef] [PubMed]
- Makhro, A.; Hanggi, P.; Goede, J.S.; Wang, J.; Bruggemann, A.; Gassmann, M.; Schmugge, M.; Kaestner, L.; Speer, O.; Bogdanova, A. N-methyl-D-aspartate receptors in human erythroid precursor cells and in circulating red blood cells contribute to the intracellular calcium regulation. Am. J. Physiol. Cell Physiol. 2013, 305, C1123–C1138. [Google Scholar] [CrossRef] [PubMed]
- Belkacemi, A.; Trost, C.F.; Tinschert, R.; Flormann, D.; Malihpour, M.; Wagner, C.; Meyer, M.R.; Beck, A.; Flockerzi, V. The TRPV2 channel mediates Ca2+ influx and the Delta9-THC-dependent decrease in osmotic fragility in red blood cells. Haematologica 2021, 106, 2246–2250. [Google Scholar] [CrossRef]
- Lew, V.L.; Tsien, R.Y.; Miner, C.; Bookchin, R.M. Physiological [Ca2+]i level and pump-leak turnover in intact red cells measured using an incorporated Ca chelator. Nature 1982, 298, 478–481. [Google Scholar] [CrossRef] [PubMed]
- Pasini, E.M.; Kirkegaard, M.; Mortensen, P.; Lutz, H.U.; Thomas, A.W.; Mann, M. In-depth analysis of the membrane and cytosolic proteome of red blood cells. Blood 2006, 108, 791–801. [Google Scholar] [CrossRef]
- Zambo, B.; Varady, G.; Padanyi, R.; Szabo, E.; Nemeth, A.; Lango, T.; Enyedi, A.; Sarkadi, B. Decreased calcium pump expression in human erythrocytes is connected to a minor haplotype in the ATP2B4 gene. Cell Calcium 2017, 65, 73–79. [Google Scholar] [CrossRef] [PubMed]
- Silver, R.T.; Krichevsky, S. Distinguishing essential thrombocythemia JAK2V617F from polycythemia vera: Limitations of erythrocyte values. Haematologica 2019, 104, 2200. [Google Scholar] [CrossRef] [PubMed]
- Lew, V.L.; Tiffert, T. On the Mechanism of Human Red Blood Cell Longevity: Roles of Calcium, the Sodium Pump, PIEZO1, and Gardos Channels. Front. Physiol. 2017, 8, 977. [Google Scholar] [CrossRef]
- Kaestner, L.; Bernhardt, I. Ion channels in the human red blood cell membrane: Their further investigation and physiological relevance. Bioelectrochemistry 2002, 55, 71–74. [Google Scholar] [CrossRef]
- Steffen, P.; Jung, A.; Nguyen, D.B.; Muller, T.; Bernhardt, I.; Kaestner, L.; Wagner, C. Stimulation of human red blood cells leads to Ca2+-mediated intercellular adhesion. Cell Calcium 2011, 50, 54–61. [Google Scholar] [CrossRef]
- Kaestner, L.; Steffen, P.; Nguyen, D.B.; Wang, J.; Wagner-Britz, L.; Jung, A.; Wagner, C.; Bernhardt, I. Lysophosphatidic acid induced red blood cell aggregation in vitro. Bioelectrochemistry 2012, 87, 89–95. [Google Scholar] [CrossRef]
- Mohandas, N.; Evans, E. Sickle erythrocyte adherence to vascular endothelium. Morphologic correlates and the requirement for divalent cations and collagen-binding plasma proteins. J. Clin. Investig. 1985, 76, 1605–1612. [Google Scholar] [CrossRef]
- Noh, J.Y.; Lim, K.M.; Bae, O.N.; Chung, S.M.; Lee, S.W.; Joo, K.M.; Lee, S.D.; Chung, J.H. Procoagulant and prothrombotic activation of human erythrocytes by phosphatidic acid. Am. J. Physiol. Heart Circ. Physiol. 2010, 299, H347–H355. [Google Scholar] [CrossRef] [PubMed]
- Borst, O.; Abed, M.; Alesutan, I.; Towhid, S.T.; Qadri, S.M.; Foller, M.; Gawaz, M.; Lang, F. Dynamic adhesion of eryptotic erythrocytes to endothelial cells via CXCL16/SR-PSOX. Am. J. Physiol. Cell Physiol. 2012, 302, C644–C651. [Google Scholar] [CrossRef] [PubMed]
- Smeets, M.W.; Bierings, R.; Meems, H.; Mul, F.P.; Geerts, D.; Vlaar, A.P.; Voorberg, J.; Hordijk, P.L. Platelet-independent adhesion of calcium-loaded erythrocytes to von Willebrand factor. PLoS ONE 2017, 12, e0173077. [Google Scholar] [CrossRef] [PubMed]
- Perez-Riverol, Y.; Csordas, A.; Bai, J.; Bernal-Llinares, M.; Hewapathirana, S.; Kundu, D.J.; Inuganti, A.; Griss, J.; Mayer, G.; Eisenacher, M.; et al. The PRIDE database and related tools and resources in 2019: Improving support for quantification data. Nucl. Acids Res. 2019, 47, D442–D450. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cells | Comparison | More Abundant in X Group | More Abundant in Y Group | On Group X Off Group Y | On Group Y Off Group X |
|---|---|---|---|---|---|
| X vs. Y | |||||
| Red Blood Cells | PV JAK2 vs. CT | 11 | 10 | 14 | 23 |
| ET JAK2 vs. CT | 12 | 18 | 11 | 24 | |
| PV JAK2 vs. ET JAK2 | 13 | 12 | 14 | 5 | |
| Reticulocytes | PV JAK2 vs. CT | 39 | 11 | 83 | 5 |
| ET JAK2 vs. CT | 55 | 14 | 215 | 1 | |
| PV JAK2 vs. ET JAK2 | 33 | 49 | 6 | 117 |
| Red Blood Cells PV JAK2 vs. CT | Gene Name | Accession Number | p-Value | Fold Change | Subcellular Location | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Protein | Cytoplasm | Plasma Membrane | Mitochondria | Nucleus | Cytoskeleton | Endoplasmic Reticulum | Endosome | ||||
| Myosin-10 | MYH10 | P35580 | 0.0273 | 8.0 | C | Pm | N | S | |||
| SEC14-like protein 2 | SEC14L2 | O76054 | 0.0457 | 1.8 | C | N | |||||
| Charged multivesicular body protein 5 | CHMP5 | Q9NZZ3 | 0.0081 | 1.5 | C | N | E | ||||
| 14-3-3 protein gamma | YWHAG | P61981 | 0.0079 | 1.4 | C | Pm | M | ||||
| Gamma-glutamylcyclotransferase | GGCT | O75223 | 0.0170 | 1.4 | C | ||||||
| Probable tRNA(His) guanylyltransferase | THG1L | Q9NWX6 | 0.0098 | 1.4 | C | M | |||||
| Alcohol dehydrogenase [NADP(+)] | AKR1A1 | P14550 | 0.0360 | 1.3 | C | Pm | |||||
| 14-3-3 protein beta/alpha | YWHAB | P31946-2 | 0.0299 | 1.3 | C | Pm | M | N | |||
| Retinal dehydrogenase 1 | ALDH1A1 | P00352 | 0.0235 | 1.3 | C | ||||||
| NHL repeat-containing protein 2 | NHLRC2 | Q8NBF2 | 0.0382 | 1.2 | C | ||||||
| Transforming protein RhoA | RHOA | P61586 | 0.0275 | 1.2 | C | Pm | S | Er | E | ||
| Mannose-1-phosphate guanyltransferase alpha | GMPPA | Q96IJ6 | 0.0227 | −1.2 | C | ||||||
| Gamma-enolase | ENO2 | P09104 | 0.0494 | −1.3 | C | Pm | |||||
| Adenosine kinase | ADK | P55263 | 0.0230 | −1.3 | C | Pm | N | ||||
| Thioredoxin | TXN | P10599 | 0.0391 | −1.3 | C | N | |||||
| Eukaryotic translation initiation factor 2 subunit 2 | EIF2S2 | P20042 | 0.0258 | −1.5 | C | ||||||
| Prostaglandin E synthase 3 | PTGES3 | Q15185-4 | 0.0240 | −1.5 | C | N | |||||
| Sorting nexin-6 | SNX6 | Q9UNH7 | 0.0076 | −1.5 | C | N | E | ||||
| DnaJ homolog subfamily B member 2 | DNAJB2 | P25686 | 0.0145 | −1.6 | C | N | Er | ||||
| Semaphorin-7A | SEMA7A | O75326 | 0.0047 | −1.6 | Pm | ||||||
| Guanine nucleotide-binding protein subunit alpha-13 | GNA13 | Q14344 | 0.0486 | −1.7 | C | Pm | N | ||||
 | |||||||||||
| Reticulocytes PV JAK2 vs. CT | Gene Name | Accession Number | p-Value | Fold Change | Subcellular Location | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Protein | Cytoplasm | Plasma Membrane | Mitochondria | Nucleus | Cytoskeleton | Endoplasmic Reticulum | Ribosomes | Golgi Apparatus | Endosome | ||||
| Transferrin receptor protein 1 | TFRC | P02786 | 0.0439 | 6.4 | Pm | E | |||||||
| ATP synthase subunit alpha, mitochondrial | ATP5A1 | P25705 | 0.0012 | 4.3 | M | ||||||||
| 60 kDa heat shock protein, mitochondrial | HSPD1 | P10809 | 0.0212 | 3.9 | M | ||||||||
| Methionine–tRNA ligase, cytoplasmic | MARS | P56192 | 0.0100 | 3.8 | C | ||||||||
| Prohibitin | PHB | P35232 | 0.0253 | 3.3 | C | Pm | M | N | |||||
| Myosin-9 | MYH9 | P35579 | 0.0221 | 3.1 | C | N | S | ||||||
| Stress-70 protein, mitochondrial | HSPA9 | P38646 | 0.0151 | 2.7 | M | ||||||||
| Isocitrate dehydrogenase [NADP], mitochondrial | IDH2 | P48735 | 0.0493 | 2.4 | M | ||||||||
| Protein-L-isoaspartate(D-aspartate) O-methyltransferase | PCMT1 | P22061 | 0.0466 | 2.4 | C | ||||||||
| Vacuolar protein sorting-associated protein 35 | VPS35 | Q96QK1 | 0.0356 | 2.3 | C | E | |||||||
| 60S ribosomal protein L22 | RPL22 | P35268 | 0.0056 | 2.0 | C | N | R | ||||||
| Synembryn-A | RIC8A | Q9NPQ8-4 | 0.0292 | 2.0 | C | Pm | |||||||
| Voltage-dependent anion-selective channel protein 3 | VDAC3 | Q9Y277 | 0.0127 | 2.0 | M | N | |||||||
| RNA 3-terminal phosphate cyclase | RTCA | O00442 | 0.0355 | 2.0 | N | ||||||||
| Protein disulfide-isomerase | P4HB | P07237 | 0.0232 | 1.9 | C | Pm | S | Er | G | ||||
| 40S ribosomal protein S7 | RPS7 | P62081 | 0.0080 | 1.8 | C | N | S | R | |||||
| 60S ribosomal protein L23a | RPL23A | P62750 | 0.0106 | 1.8 | C | N | R | ||||||
| Glucose 1,6-bisphosphate synthase | PGM2L1 | Q6PCE3 | 0.0358 | 1.8 | C | ||||||||
| Trifunctional enzyme subunit beta, mitochondrial | HADHB | P55084-2 | 0.0004 | 1.8 | M | ||||||||
| Glutamine synthetase | GLUL | P15104 | 0.0127 | 1.8 | C | Pm | M | N | Er | ||||
| 40S ribosomal protein S18 | RPS18 | P62269 | 0.0494 | 1.8 | C | N | R | ||||||
| 60S ribosomal protein L11 | RPL11 | P62913 | 0.0410 | 1.7 | C | N | R | ||||||
| Schlafen family member 14 | SLFN14 | P0C7P3-2 | 0.0323 | 1.7 | C | N | |||||||
| 60S ribosomal protein L8 | RPL8 | P62917 | 0.0117 | 1.7 | C | R | |||||||
| 60S ribosomal protein L13a | RPL13A | P40429 | 0.0423 | 1.6 | C | N | R | ||||||
| 60S ribosomal protein L7a | RPL7A | P62424 | 0.0085 | 1.6 | C | N | R | ||||||
| Tyrosine-protein phosphatase non-receptor type 11 | PTPN11 | Q06124 | 0.0421 | 1.6 | C | Pm | M | N | |||||
| Trifunctional purine biosynthetic protein adenosine-3 | GART | P22102 | 0.0009 | 1.6 | C | ||||||||
| Coatomer subunit delta | ARCN1 | P48444 | 0.0006 | 1.5 | C | Er | G | ||||||
| 60S ribosomal protein L12 | RPL12 | P30050 | 0.0366 | 1.5 | C | N | R | ||||||
| Argininosuccinate lyase | ASL | P04424 | 0.0373 | 1.5 | C | ||||||||
| 40S ribosomal protein S2 | RPS2 | P15880 | 0.0059 | 1.5 | C | N | R | ||||||
| Ras GTPase-activating protein-binding protein 1 | G3BP1 | Q13283 | 0.0354 | 1.5 | C | N | R | ||||||
| Aspartate–tRNA ligase, cytoplasmic | DARS | P14868 | 0.0360 | 1.4 | C | ||||||||
| Heterogeneous nuclear ribonucleoprotein Q | SYNCRIP | O60506-3 | 0.0391 | 1.4 | C | N | Er | R | |||||
| 14-3-3 protein beta/alpha | YWHAB | P31946-2 | 0.0410 | 1.3 | C | Pm | M | N | |||||
| Eukaryotic translation initiation factor 3 subunit J | EIF3J | O75822 | 0.0356 | 1.3 | C | ||||||||
| Cysteine protease ATG4A | ATG4A | Q8WYN0 | 0.0101 | 1.3 | C | ||||||||
| Double-stranded RNA-binding protein Staufen homolog 1 | STAU1 | O95793-2 | 0.0248 | 1.2 | C | Pm | S | Er | |||||
| Spermine synthase | SMS | P52788 | 0.0244 | −1.5 | C | ||||||||
| Dipeptidyl peptidase 9 | DPP9 | Q86TI2-4 | 0.0112 | −1.5 | C | N | S | ||||||
| Alpha-actinin-4 | ACTN4 | O43707 | 0.0044 | −1.5 | C | N | S | R | |||||
| Ribosyldihydronicotinamide dehydrogenase [quinone] | NQO2 | P16083 | 0.0390 | −1.6 | C | N | |||||||
| Anamorsin | CIAPIN1 | Q6FI81-3 | 0.0218 | −1.7 | C | M | N | ||||||
| Xaa-Pro dipeptidase | PEPD | P12955 | 0.0258 | −1.7 | |||||||||
| Probable E3 ubiquitin-protein ligase HECTD4 | HECTD4 | Q9Y4D8-5 | 0.0242 | −1.7 | Pm | ||||||||
| Importin-4 | IPO4 | Q8TEX9 | 0.0176 | −1.9 | C | N | |||||||
| Glutathione S-transferase Mu 3 | GSTM3 | P21266 | 0.0182 | −1.9 | C | N | |||||||
| Rab GDP dissociation inhibitor alpha | GDI1 | P31150 | 0.0250 | −2.0 | C | G | |||||||
| Eukaryotic initiation factor 4A-III | EIF4A3 | P38919 | 0.0295 | −2.1 | C | N | |||||||
 | |||||||||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Buks, R.; Dagher, T.; Rotordam, M.G.; Monedero Alonso, D.; Cochet, S.; Gautier, E.-F.; Chafey, P.; Cassinat, B.; Kiladjian, J.-J.; Becker, N.; et al. Altered Ca2+ Homeostasis in Red Blood Cells of Polycythemia Vera Patients Following Disturbed Organelle Sorting during Terminal Erythropoiesis. Cells 2022, 11, 49. https://doi.org/10.3390/cells11010049
Buks R, Dagher T, Rotordam MG, Monedero Alonso D, Cochet S, Gautier E-F, Chafey P, Cassinat B, Kiladjian J-J, Becker N, et al. Altered Ca2+ Homeostasis in Red Blood Cells of Polycythemia Vera Patients Following Disturbed Organelle Sorting during Terminal Erythropoiesis. Cells. 2022; 11(1):49. https://doi.org/10.3390/cells11010049
Chicago/Turabian StyleBuks, Ralfs, Tracy Dagher, Maria Giustina Rotordam, David Monedero Alonso, Sylvie Cochet, Emilie-Fleur Gautier, Philippe Chafey, Bruno Cassinat, Jean-Jacques Kiladjian, Nadine Becker, and et al. 2022. "Altered Ca2+ Homeostasis in Red Blood Cells of Polycythemia Vera Patients Following Disturbed Organelle Sorting during Terminal Erythropoiesis" Cells 11, no. 1: 49. https://doi.org/10.3390/cells11010049
APA StyleBuks, R., Dagher, T., Rotordam, M. G., Monedero Alonso, D., Cochet, S., Gautier, E.-F., Chafey, P., Cassinat, B., Kiladjian, J.-J., Becker, N., Plo, I., Egée, S., & El Nemer, W. (2022). Altered Ca2+ Homeostasis in Red Blood Cells of Polycythemia Vera Patients Following Disturbed Organelle Sorting during Terminal Erythropoiesis. Cells, 11(1), 49. https://doi.org/10.3390/cells11010049