
The Transmission of SARS-CoV-2 Infection on the Ocular Surface and Prevention Strategies

 , ,
, ,
Abstract
1. Introduction
2. Ocular Signs of COVID-19 Patients
3. Viral Structure of SARS-CoV-2
4. Method
5. The Ocular Tropism of SARS-CoV-2
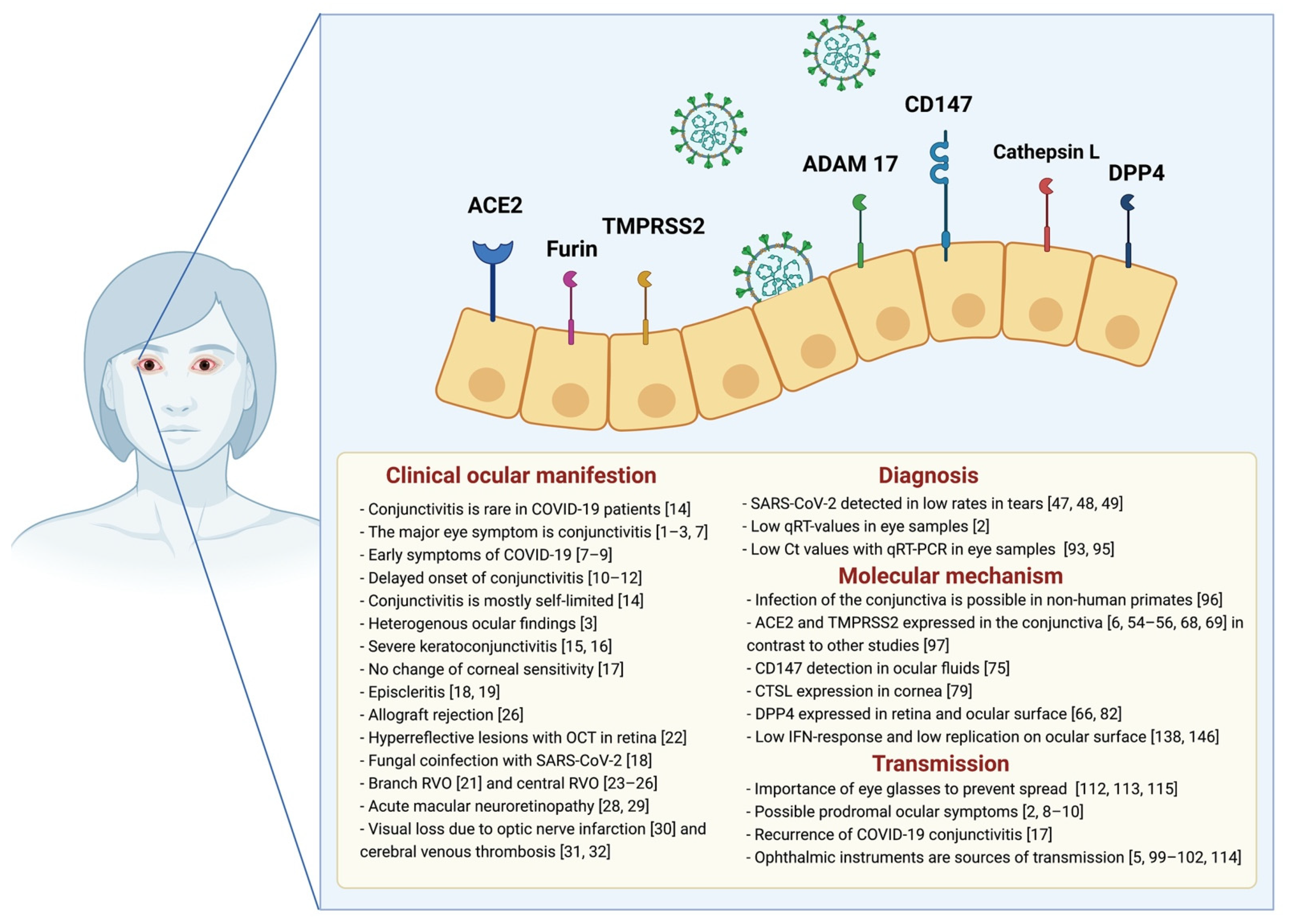
- The ACE2 is confirmed as the primary receptor of SARS-CoV-2 [48] and predominantly present in corneal and conjunctival cells [49,50,51]. Interestingly, retina is a tissue which highly expresses ACE2. These results were proven by analyzing ACE2 expression across different tissues on the mRNA level [52,53]. In addition, the receptor was also determined directly via immunofluorescence staining, both in the cytoplasm as well as on the cell membrane [54]. The main function of ACE2 is the control of the blood pressure via the regulation of blood volume [55]. However, components of the renin-angiotensin system could also be detected in various components of human eyes, yet the function remains unclear [56]. The virus entry is enhanced by overexpression of ACE2, and Monteil et al. recently showed the efficient inhibition of a SARS-CoV-2 infection by using recombinant ACE2. Here recombinant ACE2 blocks SARS-CoV-2 in vitro [57,58,59]. However, studies show that downregulation of ACE2 leads to a proinflammatory process in the lung [59]. It is hypothesized that ADAM17 is relevant for the fusion of SARS-CoV-2 with the host membrane. On the other hand, it is assumed that the shedding of ACE2 leads to a lower ACE2 membrane form and, from this, to a reduced virus entry [60].
- Furin belongs to the family of type I transmembrane serine-proteases. The enzymatic activity is connected to the subtilisin-like catalytic domain [61,62]. The role of furin as an activator of various substrates is well investigated [63]. Collin et al. prove the expression of furin at the conjunctiva and at the corneal and limbal epithelium [64], suggesting a potential entry of SARS-CoV-2 at the ocular surface.
- The transmembrane serine protease 2 (TMPRSS2) is responsible for priming of the S protein and therefore is very important for host entry. Hoffmann et al. published their findings on the blocking of a SARS-CoV-2 infection by using serine protease inhibitors, a method that emphasizes the relevance of this protease [65]. Zhou et al. were able to detect TMPRSS2 on various specimens of the eye [66], and TMPRSS2 is reported as highly expressed in the murine cornea, compared to the conjunctiva [67,68]. Recently published data show that ACE2 and TMPRSS2 is co-expressed at the human conjunctiva [69].
- The disintegrin and metalloproteinase domain 17 (ADAM17) is an important proteinase, expressed by various tissues [57]. It is responsible for the shedding of protein ectodomains and is popular for its shedding of TNFalpha. Additionally, ACE2 is also shed by ADAM17 and upregulated by angiotensin II via type 1 receptors (AT1). However, Palau et al. mentioned the controversial debate about ADAM17 and suggested a protective effect on COVID-19 of ADAM17 inhibition. Altogether, the impact of ADAM17 on SARS-CoV-2 needs to be clarified to understand how ADAM17 regulates the viral entry [70]. In general, ADMA17 is known to be relevant for the morphology of the eye, proven in mouse experiments [71].
- CD147 (HAb18G, basigin, EMMPRIN) is a member of the transmembrane glycoproteins of the immunoglobulin superfamily. Pushkarsky et al. described CD147 as a crucial cofactor for HIV infection, demonstrating a high replication capacity at early stages of infection. Recently published data suggest an interaction of CD147 with SARS-CoV-2 [72]. The authors were able to show that SARS-CoV-2 enters the cell via CD147-mediated endocytosis. This clathrin-independent endocytosis was also described in a mouse model with humanized CD147 mice [72]. Concerning human eyes, the receptor could already be detected in the cornea, the conjunctiva, and in ocular fluids [73,74].
- Cathepsin L (CTSL) is a cysteine protease recently described as involved in the establishment of COVID-19. This lysosomal enzyme is crucial for the initiation of protein degradation, e.g., for the L-lactate dehydrogenase (LDH) [75]. For SARS-CoV-2 a cleavage of the S1 unit has been reported [76]. The human cornea expresses CTSL and is reported to be relevant for corneal angiogenesis [77]. CTLS-inhibitors were mentioned as a potential treatment strategy in COVID-19 patients and could be used as a combination therapy as a useful strategy to block the host cell entry.
- The dipeptidyl peptidase-4 (DPP4) belongs to the family of type I transmembrane proteins, available in the soluble and transmembrane form. DPP4 has a high cleavage potential and therefore various substrates are known to be cleaved. In addition to the Glucagon-like-peptide (GLP-1), additional proinflammatory cytokines and chemokines were inactivated by DPP4 (e.g., RANTES, IP-10, HMGB1) [78]. DPP4 was described as a main receptor for the Middle East respiratory syndrome (MERS) in 2012, and during the early days of the ongoing pandemic it was described as a coreceptor able to bind to the spike protein of SARS-CoV-2 [79]. DPP4 is widely expressed and also described in the retina [80] and the ocular surface [64].
6. Modes of Transmission and Prevention of Viral Infections
6.1. Transmission Routes of SARS-CoV-2
6.2. Prevention of Viral Transmission Triggered by Ocular Infections
7. Therapy for Conjunctivitis Associated with SARS-CoV-2
8. Discussion
9. Conclusions and Future Directions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Guan, W.J.; Ni, Z.Y.; Hu, Y.; Liang, W.H.; Ou, C.Q.; He, J.X.; Liu, L.; Shan, H.; Lei, C.L.; Hui, D.S.C.; et al. Clinical Characteristics of Coronavirus Disease 2019 in China. N. Engl. J. Med. 2020, 382, 1708–1720. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Deng, C.; Chen, X.; Zhang, X.; Chen, B.; Yu, H.; Qin, Y.; Xiao, K.; Zhang, H.; Sun, X. Ocular manifestations and clinical characteristics of 535 cases of COVID-19 in Wuhan, China: A cross-sectional study. Acta Ophthalmol. 2020, 98, e951–e959. [Google Scholar] [CrossRef] [PubMed]
- Wu, P.; Duan, F.; Luo, C.; Liu, Q.; Qu, X.; Liang, L.; Wu, K. Characteristics of Ocular Findings of Patients With Coronavirus Disease 2019 (COVID-19) in Hubei Province, China. JAMA Ophthalmol. 2020. [Google Scholar] [CrossRef]
- Binder, A.M.; Biggs, H.M.; Haynes, A.K.; Chommanard, C.; Lu, X.; Erdman, D.D.; Watson, J.T.; Gerber, S.I. Human Adenovirus Surveillance—United States, 2003–2016. Mmwr. Morb. Mortal. Wkly. Rep. 2017, 66, 1039–1042. [Google Scholar] [CrossRef] [PubMed]
- Napoli, P.E.; Nioi, M.; d’Aloja, E.; Fossarello, M. Safety Recommendations and Medical Liability in Ocular Surgery during the COVID-19 Pandemic: An Unsolved Dilemma. J. Clin. Med. 2020, 9, 1403. [Google Scholar] [CrossRef] [PubMed]
- Napoli, P.E.; Nioi, M.; d’Aloja, E.; Fossarello, M. The Ocular Surface and the Coronavirus Disease 2019: Does a Dual ‘Ocular Route’ Exist? J. Clin. Med. 2020, 9, 1269. [Google Scholar] [CrossRef]
- Zhou, Y.; Duan, C.; Zeng, Y.; Tong, Y.; Nie, Y.; Yang, Y.; Chen, Z.; Chen, C. Ocular Findings and Proportion with Conjunctival SARS-COV-2 in COVID-19 Patients. Ophthalmology 2020. [Google Scholar] [CrossRef]
- Inomata, T.; Kitazawa, K.; Kuno, T.; Sung, J.; Nakamura, M.; Iwagami, M.; Takagi, H.; Midorikawa-Inomata, A.; Zhu, J.; Fujimoto, K.; et al. Clinical and Prodromal Ocular Symptoms in Coronavirus Disease: A Systematic Review and Meta-Analysis. Invest. Ophthalmol. Vis. Sci. 2020, 61, 29. [Google Scholar] [CrossRef]
- Hong, N.; Yu, W.; Xia, J.; Shen, Y.; Yap, M.; Han, W. Evaluation of ocular symptoms and tropism of SARS-CoV-2 in patients confirmed with COVID-19. Acta Ophthalmol. 2020. [Google Scholar] [CrossRef]
- Sindhuja, K.; Lomi, N.; Asif, M.I.; Tandon, R. Clinical profile and prevalence of conjunctivitis in mild COVID-19 patients in a tertiary care COVID-19 hospital: A retrospective cross-sectional study. Indian J. Ophthalmol. 2020, 68, 1546–1550. [Google Scholar] [CrossRef]
- Nayak, B.; Poddar, C.; Panigrahi, M.K.; Tripathy, S.; Mishra, B. Late manifestation of follicular conjunctivitis in ventilated patient following COVID-19 positive severe pneumonia. Indian J. Ophthalmol. 2020, 68, 1675–1677. [Google Scholar] [CrossRef] [PubMed]
- Navel, V.; Chiambaretta, F.; Dutheil, F. Haemorrhagic conjunctivitis with pseudomembranous related to SARS-CoV-2. Am. J. Ophthalmol. Case Rep. 2020, 19, 100735. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Liu, M.; Zhang, Z.; Qiao, K.; Huang, T.; Chen, M.; Xin, N.; Huang, Z.; Liu, L.; Zhang, G.; et al. Ocular manifestations of a hospitalised patient with confirmed 2019 novel coronavirus disease. Br. J. Ophthalmol. 2020. [Google Scholar] [CrossRef] [PubMed]
- Belser, J.A.; Rota, P.A.; Tumpey, T.M. Ocular tropism of respiratory viruses. Microbiol. Mol. Biol. Rev. 2013, 77, 144–156. [Google Scholar] [CrossRef]
- Guemes-Villahoz, N.; Burgos-Blasco, B.; Garcia-Feijoo, J.; Saenz-Frances, F.; Arriola-Villalobos, P.; Martinez-de-la-Casa, J.M.; Benitez-Del-Castillo, J.M.; Herrera de la Muela, M. Conjunctivitis in COVID-19 patients: Frequency and clinical presentation. Graefe’s Arch. Clin. Exp. Ophthalmol. 2020, 258, 2501–2507. [Google Scholar] [CrossRef]
- Cheema, M.; Aghazadeh, H.; Nazarali, S.; Ting, A.; Hodges, J.; McFarlane, A.; Kanji, J.N.; Zelyas, N.; Damji, K.F.; Solarte, C. Keratoconjunctivitis as the initial medical presentation of the novel coronavirus disease 2019 (COVID-19). Can. J. Ophthalmol. J. Can. D’ophtalmologie 2020. [Google Scholar] [CrossRef]
- Guo, D.; Xia, J.; Wang, Y.; Zhang, X.; Shen, Y.; Tong, J.P. Relapsing viral keratoconjunctivitis in COVID-19: A case report. Virol. J. 2020, 17, 97. [Google Scholar] [CrossRef]
- Pirraglia, M.P.; Ceccarelli, G.; Cerini, A.; Visioli, G.; d’Ettorre, G.; Mastroianni, C.M.; Pugliese, F.; Lambiase, A.; Gharbiya, M. Retinal involvement and ocular findings in COVID-19 pneumonia patients. Sci. Rep. 2020, 10, 17419. [Google Scholar] [CrossRef]
- Otaif, W.; Al Somali, A.I.; Al Habash, A. Episcleritis as a possible presenting sign of the novel coronavirus disease: A case report. Am. J. Ophthalmol. Case Rep. 2020, 20, 100917. [Google Scholar] [CrossRef]
- Mendez Mangana, C.; Barraquer Kargacin, A.; Barraquer, R.I. Episcleritis as an ocular manifestation in a patient with COVID-19. Acta Ophthalmol. 2020, 98, e1056–e1057. [Google Scholar] [CrossRef]
- Sheth, J.U.; Narayanan, R.; Goyal, J.; Goyal, V. Retinal vein occlusion in COVID-19: A novel entity. Indian J. Ophthalmol. 2020, 68, 2291–2293. [Google Scholar] [CrossRef] [PubMed]
- Marinho, P.M.; Marcos, A.A.A.; Romano, A.C.; Nascimento, H.; Belfort, R., Jr. Retinal findings in patients with COVID-19. Lancet 2020, 395, 1610. [Google Scholar] [CrossRef]
- Invernizzi, A.; Pellegrini, M.; Messenio, D.; Cereda, M.; Olivieri, P.; Brambilla, A.M.; Staurenghi, G. Impending Central Retinal Vein Occlusion in a Patient with Coronavirus Disease 2019 (COVID-19). Ocul. Immunol. Inflamm. 2020, 28, 1290–1292. [Google Scholar] [CrossRef] [PubMed]
- Walinjkar, J.A.; Makhija, S.C.; Sharma, H.R.; Morekar, S.R.; Natarajan, S. Central retinal vein occlusion with COVID-19 infection as the presumptive etiology. Indian J. Ophthalmol. 2020, 68, 2572–2574. [Google Scholar] [CrossRef] [PubMed]
- Gaba, W.H.; Ahmed, D.; Al Nuaimi, R.K.; Dhanhani, A.A.; Eatamadi, H. Bilateral Central Retinal Vein Occlusion in a 40-Year-Old Man with Severe Coronavirus Disease 2019 (COVID-19) Pneumonia. Am. J. Case Rep. 2020, 21, e927691. [Google Scholar] [CrossRef]
- Acharya, S.; Diamond, M.; Anwar, S.; Glaser, A.; Tyagi, P. Unique case of central retinal artery occlusion secondary to COVID-19 disease. IDCases 2020, 21, e00867. [Google Scholar] [CrossRef]
- Jin, S.X.; Juthani, V. Acute Corneal Endothelial Graft Rejection with Coinciding COVID-19 Infection. Cornea 2020. [Google Scholar] [CrossRef]
- Gascon, P.; Briantais, A.; Bertrand, E.; Ramtohul, P.; Comet, A.; Beylerian, M.; Sauvan, L.; Swiader, L.; Durand, J.M.; Denis, D. Covid-19-Associated Retinopathy: A Case Report. Ocul. Immunol. Inflamm. 2020, 28, 1293–1297. [Google Scholar] [CrossRef]
- Virgo, J.; Mohamed, M. Paracentral acute middle maculopathy and acute macular neuroretinopathy following SARS-CoV-2 infection. Eye 2020, 34, 2352–2353. [Google Scholar] [CrossRef]
- Tavakoli, M.; Sotoudeh, H.; Rezaei, A.; Saadatpour, Z.; Vaphides, M.S.; Kline, L.B. Optic nerve infarction in a patient with coronavirus disease 2019. J. Neuro-Ophthalmol. Off. J. N. Am. Neuro-Ophthalmol. Soc. 2021. [Google Scholar] [CrossRef]
- Cyr, D.G.; Vicidomini, C.M.; Siu, N.Y.; Elmann, S.E. Severe Bilateral Vision Loss in 2 Patients With Coronavirus Disease 2019. J. Neuro-Ophthalmol. Off. J. N. Am. Neuro-Ophthalmol. Soc. 2020, 40, 403–405. [Google Scholar] [CrossRef] [PubMed]
- Cavalcanti, D.D.; Raz, E.; Shapiro, M.; Dehkharghani, S.; Yaghi, S.; Lillemoe, K.; Nossek, E.; Torres, J.; Jain, R.; Riina, H.A.; et al. Cerebral Venous Thrombosis Associated with COVID-19. Ajnr. Am. J. Neuroradiol. 2020, 41, 1370–1376. [Google Scholar] [CrossRef] [PubMed]
- Louten, J. Virus Structure and Classification. Essent. Hum. Virol. 2016, 19–29. [Google Scholar] [CrossRef]
- Prasad, B.V.; Schmid, M.F. Principles of virus structural organization. Adv. Exp. Med. Biol. 2012, 726, 17–47. [Google Scholar] [CrossRef] [PubMed]
- Geller, C.; Varbanov, M.; Duval, R.E. Human coronaviruses: Insights into environmental resistance and its influence on the development of new antiseptic strategies. Viruses 2012, 4, 3044–3068. [Google Scholar] [CrossRef]
- Hu, B.; Guo, H.; Zhou, P.; Shi, Z.-L. Characteristics of SARS-CoV-2 and COVID-19. Nat. Rev. Microbiol. 2020. [Google Scholar] [CrossRef]
- Payne, S. Family Coronaviridae. Viruses 2017, 149–158. [Google Scholar] [CrossRef]
- Chen, J.; Wang, R.; Wang, M.; Wei, G.W. Mutations Strengthened SARS-CoV-2 Infectivity. J. Mol. Biol. 2020, 432, 5212–5226. [Google Scholar] [CrossRef]
- Wang, R.; Chen, J.; Gao, K.; Hozumi, Y.; Yin, C.; Wei, G.W. Analysis of SARS-CoV-2 mutations in the United States suggests presence of four substrains and novel variants. Commun. Biol. 2021, 4, 228. [Google Scholar] [CrossRef]
- Korber, B.; Fischer, W.M.; Gnanakaran, S.; Yoon, H.; Theiler, J.; Abfalterer, W.; Hengartner, N.; Giorgi, E.E.; Bhattacharya, T.; Foley, B.; et al. Tracking Changes in SARS-CoV-2 Spike: Evidence that D614G Increases Infectivity of the COVID-19 Virus. Cell 2020, 182, 812–827 e819. [Google Scholar] [CrossRef]
- Mueller, N.F.; Wagner, C.; Frazar, C.D.; Roychoudhury, P.; Lee, J.; Moncla, L.H.; Pelle, B.; Richardson, M.; Ryke, E.; Xie, H.; et al. Viral genomes reveal patterns of the SARS-CoV-2 outbreak in Washington State. medRxiv 2020. [Google Scholar] [CrossRef]
- Kirchdoerfer, R.N.; Cottrell, C.A.; Wang, N.; Pallesen, J.; Yassine, H.M.; Turner, H.L.; Corbett, K.S.; Graham, B.S.; McLellan, J.S.; Ward, A.B. Pre-fusion structure of a human coronavirus spike protein. Nature 2016, 531, 118–121. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Li, W.; Farzan, M.; Harrison, S.C. Structure of SARS coronavirus spike receptor-binding domain complexed with receptor. Science 2005, 309, 1864–1868. [Google Scholar] [CrossRef]
- Yamauchi, Y.; Helenius, A. Virus entry at a glance. J. Cell Sci. 2013, 126, 1289. [Google Scholar] [CrossRef] [PubMed]
- Xia, J.; Tong, J.; Liu, M.; Shen, Y.; Guo, D. Evaluation of coronavirus in tears and conjunctival secretions of patients with SARS-CoV-2 infection. J. Med. Virol. 2020, 92, 589–594. [Google Scholar] [CrossRef] [PubMed]
- Karimi, S.; Arabi, A.; Shahraki, T.; Safi, S. Detection of severe acute respiratory syndrome Coronavirus-2 in the tears of patients with Coronavirus disease 2019. Eye 2020, 34, 1220–1223. [Google Scholar] [CrossRef] [PubMed]
- Arora, R.; Goel, R.; Kumar, S.; Chhabra, M.; Saxena, S.; Manchanda, V.; Pumma, P. Evaluation of SARS-CoV-2 in Tears of Patients with Moderate to Severe COVID-19. Ophthalmology 2020. [Google Scholar] [CrossRef]
- Zamorano Cuervo, N.; Grandvaux, N. ACE2: Evidence of role as entry receptor for SARS-CoV-2 and implications in comorbidities. eLife 2020, 9. [Google Scholar] [CrossRef]
- Reinhold, A.; Tzankov, A.; Matter, M.; Mihic-Probst, D.; Scholl, H.P.N.; Meyer, P. Ocular pathology and occasionally detectable intraocular SARS-CoV-2 RNA in five fatal COVID-19 cases. Ophthalmic Res. 2021. [Google Scholar] [CrossRef]
- Shen, J.; Wu, J.; Yang, Y.; Wang, P.; Luo, T.; Guo, Y.; Zhao, J.; Dai, W.; Han, Y.; Zhu, P.; et al. The paradoxical problem with COVID-19 ocular infection: Moderate clinical manifestation and potential infection risk. Comput. Struct. Biotechnol. J. 2021, 19, 1063–1071. [Google Scholar] [CrossRef]
- Sahu, D.K.; Pradhan, D.; Naik, P.K.; Kar, B.; Ghosh, G.; Rath, G. Smart polymeric eye gear: A possible preventive measure against ocular transmission of COVID-19. Med. Hypotheses. 2020, 144, 110288. [Google Scholar] [CrossRef] [PubMed]
- Sungnak, W.; Huang, N.; Becavin, C.; Berg, M.; Queen, R.; Litvinukova, M.; Talavera-Lopez, C.; Maatz, H.; Reichart, D.; Sampaziotis, F.; et al. SARS-CoV-2 entry factors are highly expressed in nasal epithelial cells together with innate immune genes. Nat. Med. 2020. [Google Scholar] [CrossRef] [PubMed]
- Senanayake, P.; Drazba, J.; Shadrach, K.; Milsted, A.; Rungger-Brandle, E.; Nishiyama, K.; Miura, S.; Karnik, S.; Sears, J.E.; Hollyfield, J.G. Angiotensin II and its receptor subtypes in the human retina. Investig. Ophthalmol. Vis. Sci. 2007, 48, 3301–3311. [Google Scholar] [CrossRef]
- Grajewski, R.S.; Rokohl, A.C.; Becker, M.; Dewald, F.; Lehmann, C.; Fatkenheuer, G.; Cursiefen, C.; Klein, F.; Heindl, L.M. A missing link between SARS-CoV-2 and the eye?: ACE2 expression on the ocular surface. J. Med. Virol. 2021, 93, 78–79. [Google Scholar] [CrossRef] [PubMed]
- Oudit, G.Y.; Crackower, M.A.; Backx, P.H.; Penninger, J.M. The role of ACE2 in cardiovascular physiology. Trends Cardiovasc Med. 2003, 13, 93–101. [Google Scholar] [CrossRef]
- Holappa, M.; Vapaatalo, H.; Vaajanen, A. Many Faces of Renin-angiotensin System—Focus on Eye. Open Ophthalmol. J. 2017, 11, 122–142. [Google Scholar] [CrossRef]
- Zipeto, D.; Palmeira, J.D.F.; Arganaraz, G.A.; Arganaraz, E.R. ACE2/ADAM17/TMPRSS2 Interplay May Be the Main Risk Factor for COVID-19. Front. Immunol. 2020, 11, 576745. [Google Scholar] [CrossRef] [PubMed]
- Monteil, V.; Kwon, H.; Prado, P.; Hagelkruys, A.; Wimmer, R.A.; Stahl, M.; Leopoldi, A.; Garreta, E.; Hurtado Del Pozo, C.; Prosper, F.; et al. Inhibition of SARS-CoV-2 Infections in Engineered Human Tissues Using Clinical-Grade Soluble Human ACE2. Cell 2020, 181, 905–913.e907. [Google Scholar] [CrossRef]
- Imai, Y.; Kuba, K.; Rao, S.; Huan, Y.; Guo, F.; Guan, B.; Yang, P.; Sarao, R.; Wada, T.; Leong-Poi, H.; et al. Angiotensin-converting enzyme 2 protects from severe acute lung failure. Nature 2005, 436, 112–116. [Google Scholar] [CrossRef]
- Palau, V.; Riera, M.; Soler, M.J. ADAM17 inhibition may exert a protective effect on COVID-19. Nephrol. Dial. Transpl. 2020, 35, 1071–1072. [Google Scholar] [CrossRef]
- Brooke, G.N.; Prischi, F. Structural and functional modelling of SARS-CoV-2 entry in animal models. Sci. Rep. 2020, 10, 15917. [Google Scholar] [CrossRef] [PubMed]
- Adu-Agyeiwaah, Y.; Grant, M.B.; Obukhov, A.G. The Potential Role of Osteopontin and Furin in Worsening Disease Outcomes in COVID-19 Patients with Pre-Existing Diabetes. Cells 2020, 9, 2528. [Google Scholar] [CrossRef] [PubMed]
- Thomas, G. Furin at the cutting edge: From protein traffic to embryogenesis and disease. Nat. Rev. Mol. Cell Biol. 2002, 3, 753–766. [Google Scholar] [CrossRef] [PubMed]
- Collin, J.; Queen, R.; Zerti, D.; Dorgau, B.; Georgiou, M.; Djidrovski, I.; Hussain, R.; Coxhead, J.M.; Joseph, A.; Rooney, P.; et al. Co-expression of SARS-CoV-2 entry genes in the superficial adult human conjunctival, limbal and corneal epithelium suggests an additional route of entry via the ocular surface. Ocul. Surf. 2021, 19, 190–200. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, M.; Kleine-Weber, H.; Schroeder, S.; Kruger, N.; Herrler, T.; Erichsen, S.; Schiergens, T.S.; Herrler, G.; Wu, N.H.; Nitsche, A.; et al. SARS-CoV-2 Cell Entry Depends on ACE2 and TMPRSS2 and Is Blocked by a Clinically Proven Protease Inhibitor. Cell 2020, 181, 271–280.e278. [Google Scholar] [CrossRef]
- Zhou, L.; Xu, Z.; Castiglione, G.M.; Soiberman, U.S.; Eberhart, C.G.; Duh, E.J. ACE2 and TMPRSS2 are expressed on the human ocular surface, suggesting susceptibility to SARS-CoV-2 infection. Ocul. Surf. 2020, 18, 537–544. [Google Scholar] [CrossRef] [PubMed]
- Ma, D.; Chen, C.B.; Jhanji, V.; Xu, C.; Yuan, X.L.; Liang, J.J.; Huang, Y.; Cen, L.P.; Ng, T.K. Expression of SARS-CoV-2 receptor ACE2 and TMPRSS2 in human primary conjunctival and pterygium cell lines and in mouse cornea. Eye 2020, 34, 1212–1219. [Google Scholar] [CrossRef]
- Zimmerman, K.; Kearns, F.; Tzekov, R. Natural protection of ocular surface from viral infections—A hypothesis. Med. Hypotheses. 2020, 143, 110082. [Google Scholar] [CrossRef]
- Ma, R.; Gan, L.; Jiang, S.; Ding, P.; Chen, D.; Wu, J.; Qian, J. High expression of SARS-CoV-2 entry factors in human conjunctival goblet cells. Exp. Eye Res. 2021, 205, 108501. [Google Scholar] [CrossRef]
- Haga, S.; Yamamoto, N.; Nakai-Murakami, C.; Osawa, Y.; Tokunaga, K.; Sata, T.; Yamamoto, N.; Sasazuki, T.; Ishizaka, Y. Modulation of TNF-alpha-converting enzyme by the spike protein of SARS-CoV and ACE2 induces TNF-alpha production and facilitates viral entry. Proc. Natl. Acad. Sci. USA 2008, 105, 7809–7814. [Google Scholar] [CrossRef]
- Sel, S.; Kalinski, T.; Enssen, I.; Kaiser, M.; Nass, N.; Trau, S.; Wollensak, G.; Brauer, L.; Jager, K.; Paulsen, F. Expression analysis of ADAM17 during mouse eye development. Ann. Anat. 2012, 194, 334–338. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Chen, W.; Zhang, Z.; Deng, Y.; Lian, J.Q.; Du, P.; Wei, D.; Zhang, Y.; Sun, X.X.; Gong, L.; et al. CD147-spike protein is a novel route for SARS-CoV-2 infection to host cells. Signal Transduct. Target. Ther. 2020, 5, 283. [Google Scholar] [CrossRef] [PubMed]
- Maatta, M.; Tervahartiala, T.; Kaarniranta, K.; Tang, Y.; Yan, L.; Tuukkanen, J.; Sorsa, T. Immunolocalization of EMMPRIN (CD147) in the human eye and detection of soluble form of EMMPRIN in ocular fluids. Curr. Eye Res. 2006, 31, 917–924. [Google Scholar] [CrossRef]
- Li, Y.P.; Ma, Y.; Wang, N.; Jin, Z.B. Eyes on coronavirus. Stem Cell Res. 2021, 51, 102200. [Google Scholar] [CrossRef] [PubMed]
- Ohshita, T.; Hiroi, Y. Cathepsin L plays an important role in the lysosomal degradation of L-lactate dehydrogenase. Biosci. Biotechnol. Biochem. 2006, 70, 2254–2261. [Google Scholar] [CrossRef][Green Version]
- Liu, T.; Luo, S.; Libby, P.; Shi, G.P. Cathepsin L-selective inhibitors: A potentially promising treatment for COVID-19 patients. Pharmacol. Ther. 2020, 213, 107587. [Google Scholar] [CrossRef] [PubMed]
- Coppini, L.P.; Visniauskas, B.; Costa, E.F.; Filho, M.N.; Rodrigues, E.B.; Chagas, J.R.; Farah, M.E.; Barros, N.M.; Carmona, A.K. Corneal angiogenesis modulation by cysteine cathepsins: In vitro and in vivo studies. Exp. Eye Res. 2015, 134, 39–46. [Google Scholar] [CrossRef]
- Mulvihill, E.E.; Drucker, D.J. Pharmacology, physiology, and mechanisms of action of dipeptidyl peptidase-4 inhibitors. Endocr. Rev. 2014, 35, 992–1019. [Google Scholar] [CrossRef]
- Li, Y.; Zhang, Z.; Yang, L.; Lian, X.; Xie, Y.; Li, S.; Xin, S.; Cao, P.; Lu, J. The MERS-CoV Receptor DPP4 as a Candidate Binding Target of the SARS-CoV-2 Spike. Iscience 2020, 23, 101160. [Google Scholar] [CrossRef]
- Li, H.; Zhang, J.; Lin, L.; Xu, L. Vascular protection of DPP-4 inhibitors in retinal endothelial cells in in vitro culture. Int. Immunopharmacol. 2019, 66, 162–168. [Google Scholar] [CrossRef]
- Kutter, J.S.; Spronken, M.I.; Fraaij, P.L.; Fouchier, R.A.; Herfst, S. Transmission routes of respiratory viruses among humans. Curr. Opin. Virol. 2018, 28, 142–151. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Lu, X.; Jiang, B.; Du, Y.; Yang, Y.; Qian, H.; Liu, B.; Lin, C.; Jia, L.; Chen, L.; et al. Adenovirus-associated acute conjunctivitis in Beijing, China, 2011-2013. BMC Infect. Dis. 2018, 18, 135. [Google Scholar] [CrossRef] [PubMed]
- Sow, A.S.; Kane, H.; Ka, A.M.; Hanne, F.T.; Ndiaye, J.M.M.; Diagne, J.P.; Nguer, M.; Sow, S.; Saheli, Y.; Sy, E.H.M.; et al. Senegalese experience with acute viral conjunctivitis. J. Fr. D’ophtalmologie 2017, 40, 297–302. [Google Scholar] [CrossRef] [PubMed]
- Zhou, P.; Yang, X.L.; Wang, X.G.; Hu, B.; Zhang, L.; Zhang, W.; Si, H.R.; Zhu, Y.; Li, B.; Huang, C.L.; et al. A pneumonia outbreak associated with a new coronavirus of probable bat origin. Nature 2020, 579, 270–273. [Google Scholar] [CrossRef]
- Alonso, W.J.; Nascimento, F.C.; Shapiro, J.; Schuck-Paim, C. Facing ubiquitous viruses: When hand washing is not enough. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 2013, 56, 617. [Google Scholar] [CrossRef]
- Seah, I.Y.J.; Anderson, D.E.; Kang, A.E.Z.; Wang, L.; Rao, P.; Young, B.E.; Lye, D.C.; Agrawal, R. Assessing Viral Shedding and Infectivity of Tears in Coronavirus Disease 2019 (COVID-19) Patients. Ophthalmology 2020, 127, 977–979. [Google Scholar] [CrossRef]
- Meduri, A.; Oliverio, G.W.; Mancuso, G.; Giuffrida, A.; Guarneri, C.; Venanzi Rullo, E.; Nunnari, G.; Aragona, P. Ocular surface manifestation of COVID-19 and tear film analysis. Sci. Rep. 2020, 10, 20178. [Google Scholar] [CrossRef]
- Bayyoud, T.; Iftner, A.; Iftner, T.; Bartz-Schmidt, K.U.; Rohrbach, J.M.; Ueffing, M.; Schindler, M.; Thaler, S. Absence of Severe Acute Respiratory Syndrome-Coronavirus-2 RNA in Human Corneal Tissues. Cornea 2021, 40, 342–347. [Google Scholar] [CrossRef]
- Napoli, P.E.; Mangoni, L.; Gentile, P.; Braghiroli, M.; Fossarello, M. A Panel of Broad-Spectrum Antivirals in Topical Ophthalmic Medications from the Drug Repurposing Approach during and after the Coronavirus Disease 2019 Era. J. Clin. Med. 2020, 9, 2441. [Google Scholar] [CrossRef]
- Sawant, O.B.; Singh, S.; Wright, R.E., 3rd; Jones, K.M.; Titus, M.S.; Dennis, E.; Hicks, E.; Majmudar, P.A.; Kumar, A.; Mian, S.I. Prevalence of SARS-CoV-2 in human post-mortem ocular tissues. Ocul. Surf. 2021, 19, 322–329. [Google Scholar] [CrossRef]
- Casagrande, M.; Fitzek, A.; Spitzer, M.S.; Puschel, K.; Glatzel, M.; Krasemann, S.; Norz, D.; Lutgehetmann, M.; Pfefferle, S.; Schultheiss, M. Presence of SARS-CoV-2 RNA in the Cornea of Viremic Patients With COVID-19. JAMA Ophthalmol. 2021. [Google Scholar] [CrossRef]
- Layden, J.E.; Ghinai, I.; Pray, I.; Kimball, A.; Layer, M.; Tenforde, M.W.; Navon, L.; Hoots, B.; Salvatore, P.P.; Elderbrook, M.; et al. Pulmonary Illness Related to E-Cigarette Use in Illinois and Wisconsin—Final Report. N. Engl. J. Med. 2020, 382, 903–916. [Google Scholar] [CrossRef] [PubMed]
- Deng, W.; Bao, L.; Gao, H.; Xiang, Z.; Qu, Y.; Song, Z.; Gong, S.; Liu, J.; Liu, J.; Yu, P.; et al. Ocular conjunctival inoculation of SARS-CoV-2 can cause mild COVID-19 in rhesus macaques. Nat. Commun. 2020, 11, 4400. [Google Scholar] [CrossRef] [PubMed]
- Lange, C.; Wolf, J.; Auw-Haedrich, C.; Schlecht, A.; Boneva, S.; Lapp, T.; Horres, R.; Agostini, H.; Martin, G.; Reinhard, T.; et al. Expression of the COVID-19 receptor ACE2 in the human conjunctiva. J. Med. Virol. 2020. [Google Scholar] [CrossRef] [PubMed]
- Brooks, J.T.; Beezhold, D.H.; Noti, J.D.; Coyle, J.P.; Derk, R.C.; Blachere, F.M.; Lindsley, W.G. Maximizing Fit for Cloth and Medical Procedure Masks to Improve Performance and Reduce SARS-CoV-2 Transmission and Exposure, 2021. Mmwr. Morb. Mortal. Wkly. Rep. 2021, 70, 254–257. [Google Scholar] [CrossRef]
- Coroneo, M.T. The eye as the discrete but defensible portal of coronavirus infection. Ocul. Surf. 2021, 19, 176–182. [Google Scholar] [CrossRef]
- Felfeli, T.; Mandelcorn, E.D. Assessment of Simulated Respiratory Droplet Spread During an Ophthalmologic Slitlamp Examination. JAMA Ophthalmol. 2020, 138, 1099–1101. [Google Scholar] [CrossRef]
- Shah, S.H.; Garg, A.K.; Patel, S.; Yim, W.; Jokerst, J.V.; Chao, D.L. Assessment of Respiratory Droplet Transmission During the Ophthalmic Slit-Lamp Exam: A Particle Tracking Analysis. Am. J. Ophthalmol. 2020, 222, 76–81. [Google Scholar] [CrossRef]
- Mannan, R.; Pruthi, A.; Sud, R.; Khanduja, S. Slit lamp examination during COVID-19: Where should the protective barrier be? Indian J. Ophthalmol. 2021, 69, 376–383. [Google Scholar] [CrossRef]
- Kampf, G. Efficacy of ethanol against viruses in hand disinfection. J. Hosp. Infect. 2018, 98, 331–338. [Google Scholar] [CrossRef]
- Thuret, G.; Courrier, E.; Poinard, S.; Gain, P.; Baud’Huin, M.; Martinache, I.; Cursiefen, C.; Maier, P.; Hjortdal, J.; Sanchez Ibanez, J.; et al. One threat, different answers: The impact of COVID-19 pandemic on cornea donation and donor selection across Europe. Br. J. Ophthalmol. 2020. [Google Scholar] [CrossRef] [PubMed]
- Roy, A.; Chaurasia, S.; Fernandes, M.; Das, S. Impact of nationwide COVID-19 lockdown on keratoplasty and eye banking in India: A survey of cornea surgeons and eye banks. Indian J. Ophthalmol. 2021, 69, 706–708. [Google Scholar] [CrossRef] [PubMed]
- Nair, A.G.; Narayanan, N.; Ali, M.J. A Survey on the Impact of COVID-19 on Lacrimal Surgery: The Asia-Pacific Perspective. Clin. Ophthalmol. 2020, 14, 3789–3799. [Google Scholar] [CrossRef] [PubMed]
- Elfalah, M.; AlRyalat, S.A.; Toro, M.D.; Rejdak, R.; Zweifel, S.; Nazzal, R.; Abu-Ameerh, M.; Ababneh, O.; Gharaibeh, A.; Sharif, Z.; et al. Delayed Intravitreal Anti-VEGF Therapy for Patients During the COVID-19 Lockdown: An Ethical Endeavor. Clin. Ophthalmol. 2021, 15, 661–669. [Google Scholar] [CrossRef] [PubMed]
- Borrelli, E.; Grosso, D.; Vella, G.; Sacconi, R.; Battista, M.; Querques, L.; Zucchiatti, I.; Prascina, F.; Bandello, F.; Querques, G. Short-term outcomes of patients with neovascular exudative AMD: The effect of COVID-19 pandemic. Graefe’s Arch. Clin. Exp. Ophthalmol. 2020, 258, 2621–2628. [Google Scholar] [CrossRef]
- Aggarwal, S.; Jain, P.; Jain, A. COVID-19 and cataract surgery backlog in Medicare beneficiaries. J. Cataract Refract. Surg. 2020, 46, 1530–1533. [Google Scholar] [CrossRef]
- McGhee, C.N.J.; Dean, S.; Freundlich, S.E.N.; Gokul, A.; Ziaei, M.; Patel, D.V.; Niederer, R.L.; Danesh-Meyer, H.V. Microdroplet and spatter contamination during phacoemulsification cataract surgery in the era of COVID-19. Clin. Exp. Ophthalmol. 2020, 48, 1168–1174. [Google Scholar] [CrossRef]
- Wong, J.K.W.; Kwok, J.; Chan, J.C.H.; Shih, K.C.; Qin, R.; Lau, D.; Lai, J.S.M. Aerosolization and Fluid Spillage During Phacoemulsification in Human Subjects. Clin. Ophthalmol. 2021, 15, 307–313. [Google Scholar] [CrossRef]
- Lu, C.-w.; Liu, X.-f.; Jia, Z.-f. 2019-nCoV transmission through the ocular surface must not be ignored. Lancet 2020, 395. [Google Scholar] [CrossRef]
- Zeng, W.; Wang, X.; Li, J.; Yang, Y.; Qiu, X.; Song, P.; Xu, J.; Wei, Y. Association of Daily Wear of Eyeglasses With Susceptibility to Coronavirus Disease 2019 Infection. JAMA Ophthalmol. 2020. [Google Scholar] [CrossRef]
- Aytogan, H.; Ayintap, E.; Ozkalay Yilmaz, N. Detection of Coronavirus Disease 2019 Viral Material on Environmental Surfaces of an Ophthalmology Examination Room. JAMA Ophthalmol. 2020, 138, 990–993. [Google Scholar] [CrossRef] [PubMed]
- Alai, N. Enhancing best practices in ophthalmology for prevention of nosocomial epidemic keratoconjunctivitis infections. Curr. Med. Res. Opin. 2016, 32, 1757–1758. [Google Scholar] [CrossRef] [PubMed]
- Dai, L.; Gao, G.F. Viral targets for vaccines against COVID-19. Nat. Rev. Immunol. 2021, 21, 73–82. [Google Scholar] [CrossRef] [PubMed]
- Pushpakom, S.; Iorio, F.; Eyers, P.A.; Escott, K.J.; Hopper, S.; Wells, A.; Doig, A.; Guilliams, T.; Latimer, J.; McNamee, C.; et al. Drug repurposing: Progress, challenges and recommendations. Nat. Rev. Drug Discov. 2019, 18, 41–58. [Google Scholar] [CrossRef]
- Khan, A.; Benthin, C.; Zeno, B.; Albertson, T.E.; Boyd, J.; Christie, J.D.; Hall, R.; Poirier, G.; Ronco, J.J.; Tidswell, M.; et al. A pilot clinical trial of recombinant human angiotensin-converting enzyme 2 in acute respiratory distress syndrome. Crit. Care 2017, 21, 234. [Google Scholar] [CrossRef]
- Latil, M.; Camelo, S.; Veillet, S.; Lafont, R.; Dilda, P.J. Developing new drugs that activate the protective arm of the renin-angiotensin system as a potential treatment for respiratory failure in COVID-19 patients. Drug Discov. Today 2021. [Google Scholar] [CrossRef] [PubMed]
- Lambert, D.W.; Clarke, N.E.; Hooper, N.M.; Turner, A.J. Calmodulin interacts with angiotensin-converting enzyme-2 (ACE2) and inhibits shedding of its ectodomain. FEBS Lett. 2008, 582, 385–390. [Google Scholar] [CrossRef] [PubMed]
- Cohen, J.B.; Hanff, T.C.; Corrales-Medina, V.; William, P.; Renna, N.; Rosado-Santander, N.R.; Rodriguez-Mori, J.E.; Spaak, J.; Andrade-Villanueva, J.; Chang, T.I.; et al. Randomized elimination and prolongation of ACE inhibitors and ARBs in coronavirus 2019 (REPLACE COVID) Trial Protocol. J. Clin. Hypertens. 2020, 22, 1780–1788. [Google Scholar] [CrossRef]
- Ahmad, S.I. 5-Fluorouracil in combination with deoxyribonucleosides and deoxyribose as possible therapeutic options for the Coronavirus, COVID-19 infection. Med. Hypotheses. 2020, 142, 109754. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, M.; Schroeder, S.; Kleine-Weber, H.; Muller, M.A.; Drosten, C.; Pohlmann, S. Nafamostat Mesylate Blocks Activation of SARS-CoV-2: New Treatment Option for COVID-19. Antimicrob. Agents Chemother. 2020, 64. [Google Scholar] [CrossRef] [PubMed]
- Smieszek, S.P.; Przychodzen, B.P.; Polymeropoulos, M.H. Amantadine disrupts lysosomal gene expression: A hypothesis for COVID19 treatment. Int. J. Antimicrob. Agents 2020, 55, 106004. [Google Scholar] [CrossRef]
- Barnett, B.P.; Wahlin, K.; Krawczyk, M.; Spencer, D.; Welsbie, D.; Afshari, N.; Chao, D. Potential of Ocular Transmission of SARS-CoV-2: A Review. Vision 2020, 4, 40. [Google Scholar] [CrossRef] [PubMed]
- Zhou, N.; Pan, T.; Zhang, J.; Li, Q.; Zhang, X.; Bai, C.; Huang, F.; Peng, T.; Zhang, J.; Liu, C.; et al. Glycopeptide Antibiotics Potently Inhibit Cathepsin L in the Late Endosome/Lysosome and Block the Entry of Ebola Virus, Middle East Respiratory Syndrome Coronavirus (MERS-CoV), and Severe Acute Respiratory Syndrome Coronavirus (SARS-CoV). J. Biol. Chem. 2016, 291, 9218–9232. [Google Scholar] [CrossRef] [PubMed]
- Padmanabhan, P.; Desikan, R.; Dixit, N.M. Targeting TMPRSS2 and Cathepsin B/L together may be synergistic against SARS-CoV-2 infection. PLoS Comput. Biol. 2020, 16, e1008461. [Google Scholar] [CrossRef] [PubMed]
- Ulrich, H.; Pillat, M.M. CD147 as a Target for COVID-19 Treatment: Suggested Effects of Azithromycin and Stem Cell Engagement. Stem. Cell Rev. Rep. 2020, 16, 434–440. [Google Scholar] [CrossRef] [PubMed]
- Shilts, J.; Crozier, T.W.M.; Greenwood, E.J.D.; Lehner, P.J.; Wright, G.J. No evidence for basigin/CD147 as a direct SARS-CoV-2 spike binding receptor. Sci. Rep. 2021, 11, 413. [Google Scholar] [CrossRef]
- Solerte, S.B.; D’Addio, F.; Trevisan, R.; Lovati, E.; Rossi, A.; Pastore, I.; Dell’Acqua, M.; Ippolito, E.; Scaranna, C.; Bellante, R.; et al. Sitagliptin Treatment at the Time of Hospitalization Was Associated With Reduced Mortality in Patients With Type 2 Diabetes and COVID-19: A Multicenter, Case-Control, Retrospective, Observational Study. Diabetes Care 2020, 43, 2999–3006. [Google Scholar] [CrossRef]
- Greber, U.F.; Willetts, M.; Webster, P.; Helenius, A. Stepwise dismantling of adenovirus 2 during entry into cells. Cell 1993, 75, 477–486. [Google Scholar] [CrossRef]
- Georgi, F.; Greber, U.F. The Adenovirus Death Protein—A small membrane protein controls cell lysis and disease. FEBS Lett. 2020, 594, 1861–1878. [Google Scholar] [CrossRef]
- Sette, A.; Crotty, S. Adaptive immunity to SARS-CoV-2 and COVID-19. Cell 2021, 184, 861–880. [Google Scholar] [CrossRef]
- Matsuura, K.; Terasaka, Y.; Miyazaki, D.; Shimizu, Y.; Inoue, Y. Comparative study of topical regimen for adenoviral kearoconjunctivitis by 0.1% fluorometholone with and without polyvinyl alcohol iodine. Jpn. J. Ophthalmol. 2021, 65, 107–114. [Google Scholar] [CrossRef] [PubMed]
- Oran, D.P.; Topol, E.J. Prevalence of Asymptomatic SARS-CoV-2 Infection: A Narrative Review. Ann. Int. Med. 2020, 173, 362–367. [Google Scholar] [CrossRef] [PubMed]
- Blanco-Melo, D.; Nilsson-Payant, B.E.; Liu, W.C.; Uhl, S.; Hoagland, D.; Moller, R.; Jordan, T.X.; Oishi, K.; Panis, M.; Sachs, D.; et al. Imbalanced Host Response to SARS-CoV-2 Drives Development of COVID-19. Cell 2020, 181, 1036–1045 e1039. [Google Scholar] [CrossRef]
- Bastard, P.; Rosen, L.B.; Zhang, Q.; Michailidis, E.; Hoffmann, H.H.; Zhang, Y.; Dorgham, K.; Philippot, Q.; Rosain, J.; Beziat, V.; et al. Autoantibodies against type I IFNs in patients with life-threatening COVID-19. Science 2020, 370. [Google Scholar] [CrossRef]
- Zhang, Q.; Bastard, P.; Liu, Z.; Le Pen, J.; Moncada-Velez, M.; Chen, J.; Ogishi, M.; Sabli, I.K.D.; Hodeib, S.; Korol, C.; et al. Inborn errors of type I IFN immunity in patients with life-threatening COVID-19. Science 2020, 370. [Google Scholar] [CrossRef] [PubMed]
- Chigbu, D.I.; Labib, B.A. Pathogenesis and management of adenoviral keratoconjunctivitis. Infect. Drug Resist. 2018, 11, 981–993. [Google Scholar] [CrossRef]
- Miner, J.J.; Platt, D.J.; Ghaznavi, C.M.; Chandra, P.; Santeford, A.; Menos, A.M.; Dong, Z.; Wang, E.R.; Qian, W.; Karozichian, E.S.; et al. HSV-1 and Zika Virus but Not SARS-CoV-2 Replicate in the Human Cornea and Are Restricted by Corneal Type III Interferon. Cell Rep. 2020, 33, 108339. [Google Scholar] [CrossRef]
- Hoffmann, H.H.; Schneider, W.M.; Rice, C.M. Interferons and viruses: An evolutionary arms race of molecular interactions. Trends Immunol. 2015, 36, 124–138. [Google Scholar] [CrossRef]
- Al-Khatib, K.; Williams, B.R.; Silverman, R.H.; Halford, W.; Carr, D.J. Dichotomy between survival and lytic gene expression in RNase L- and PKR-deficient mice transduced with an adenoviral vector expressing murine IFN-beta following ocular HSV-1 infection. Exp. Eye Res. 2005, 80, 167–173. [Google Scholar] [CrossRef]
- Austin, B.A.; Halford, W.P.; Williams, B.R.; Carr, D.J. Oligoadenylate synthetase/protein kinase R pathways and alphabeta TCR+ T cells are required for adenovirus vector: IFN-gamma inhibition of herpes simplex virus-1 in cornea. J. Immunol. 2007, 178, 5166–5172. [Google Scholar] [CrossRef]
- Napoli, P.E.; Nioi, M.; Mangoni, L.; Gentile, P.; Braghiroli, M.; d’Aloja, E.; Fossarello, M. Fourier-Domain OCT Imaging of the Ocular Surface and Tear Film Dynamics: A Review of the State of the Art and an Integrative Model of the Tear Behavior During the Inter-Blink Period and Visual Fixation. J. Clin. Med. 2020, 9, 668. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Name of Potential Drugs | Targets | Function |
|---|---|---|
| APN01 | ACE2 | Soluble ACE2 [115] [NCT04335136] |
| Angiotensin receptor blocker | ACE2 | ACE2 modulator [116] |
| Melatonin | ACE2 | Calmodulin inhibitor [117] |
| Toremifene | ACE2 | Nonsteroidal antiestrogen [NCT04531748] |
| 5-fluorouracil | ADAM-17 | ADAM-17 enhancer [119] |
| Camostat mesylate | TMPRSS | TMPRSS inhibitor [65] |
| Nafamostat mesylate | TMPRSS | TMPRSS inhibitor [120] |
| Amantadine | CTSL | CTSL inhibitor [121] |
| Teicoplanin | CTSL | CTSL inhibitor [122,123] |
| Meplazumab | CD147 | Anti-CD147 [NCT04275245] |
| Azithromycin | CD147 | Indirect CD147 inhibition [125] |
| Sitagliptin | DPP4 | DDP4 inhibitor [127] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kitazawa, K.; Deinhardt-Emmer, S.; Inomata, T.; Deshpande, S.; Sotozono, C. The Transmission of SARS-CoV-2 Infection on the Ocular Surface and Prevention Strategies. Cells 2021, 10, 796. https://doi.org/10.3390/cells10040796
Kitazawa K, Deinhardt-Emmer S, Inomata T, Deshpande S, Sotozono C. The Transmission of SARS-CoV-2 Infection on the Ocular Surface and Prevention Strategies. Cells. 2021; 10(4):796. https://doi.org/10.3390/cells10040796
Chicago/Turabian StyleKitazawa, Koji, Stefanie Deinhardt-Emmer, Takenori Inomata, Sharvari Deshpande, and Chie Sotozono. 2021. "The Transmission of SARS-CoV-2 Infection on the Ocular Surface and Prevention Strategies" Cells 10, no. 4: 796. https://doi.org/10.3390/cells10040796
APA StyleKitazawa, K., Deinhardt-Emmer, S., Inomata, T., Deshpande, S., & Sotozono, C. (2021). The Transmission of SARS-CoV-2 Infection on the Ocular Surface and Prevention Strategies. Cells, 10(4), 796. https://doi.org/10.3390/cells10040796