
CXCL12-Abundant Reticular (CAR) Cells Direct Megakaryocyte Protrusions across the Bone Marrow Sinusoid Wall

 , and
, and 
Abstract
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Mouse Lines
2.2. Femur Sectioning and Staining
2.3. Confocal Laser Scanning Microscopy
2.4. Electron Microscopy
2.5. Quantification of MK Protrusions
3. Results
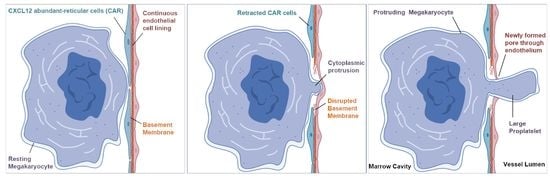
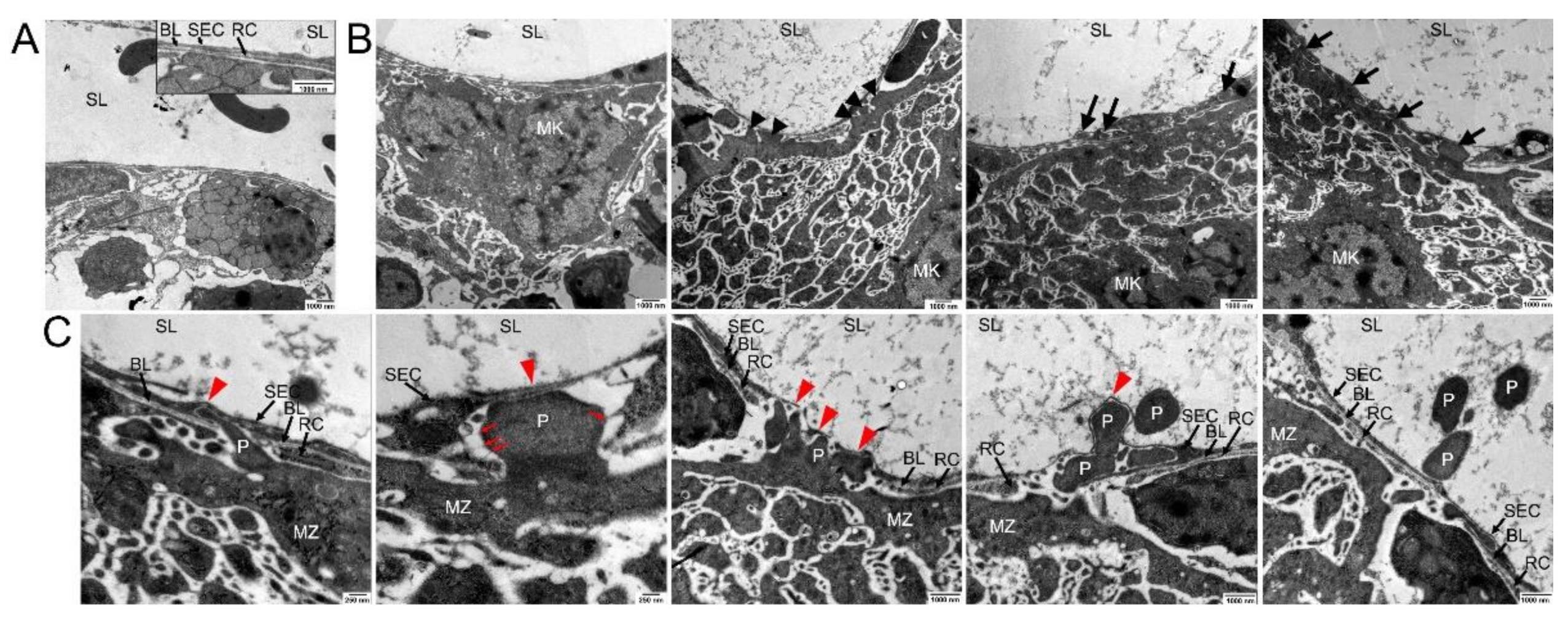
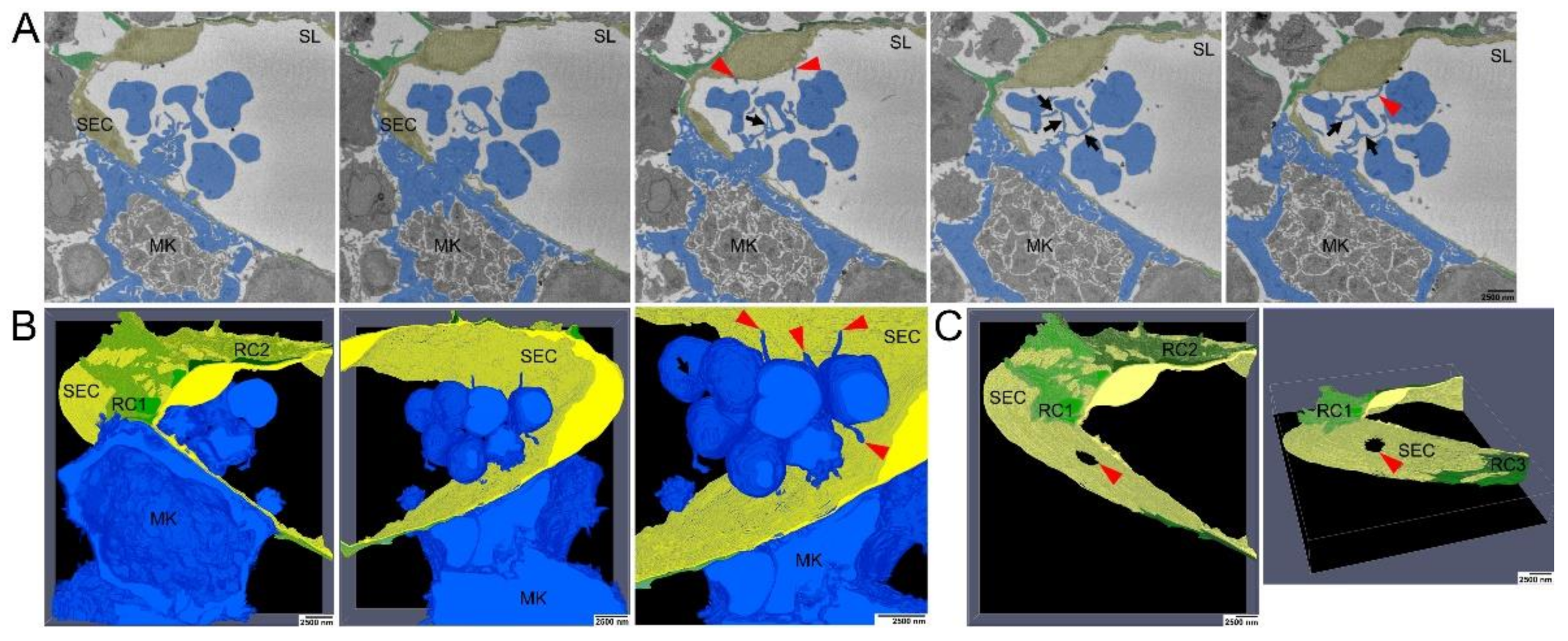
3.1. The Megakaryocyte Demarcation Membrane System during Transendothelial Passage
3.2. Ultrastructure of Megakaryocyte Protrusions during the Transendothelial Passage
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Italiano, J.E., Jr.; Lecine, P.; Shivdasani, R.A.; Hartwig, J.H. Blood platelets are assembled principally at the ends of proplatelet processes produced by differentiated megakaryocytes. J. Cell Biol. 1999, 147, 1299–1312. [Google Scholar] [CrossRef]
- De Bruyn, P.P.H.; Michelson, S.; Thomas, T.B. Migration of Blood Cells of Bone Marrow through Sinusoidal Wall. J. Morphol. 1971, 133, 417. [Google Scholar] [CrossRef] [PubMed]
- Slayton, W.B.; Li, X.M.; Butler, J.; Guthrie, S.M.; Jorgensen, M.L.; Wingard, J.R.; Scott, E.W. The role of the donor in the repair of the marrow vascular niche following hematopoietic stem cell transplant. Stem Cells 2007, 25, 2945–2955. [Google Scholar] [CrossRef]
- Junt, T.; Schulze, H.; Chen, Z.; Massberg, S.; Goerge, T.; Krueger, A.; Wagner, D.D.; Graf, T.; Italiano, J.E., Jr.; Shivdasani, R.A.; et al. Dynamic visualization of thrombopoiesis within bone marrow. Science 2007, 317, 1767–1770. [Google Scholar] [CrossRef]
- Nishimura, S.; Nagasaki, M.; Kunishima, S.; Sawaguchi, A.; Sakata, A.; Sakaguchi, H.; Ohmori, T.; Manabe, I.; Italiano, J.E.; Ryu, T.; et al. IL-1 alpha induces thrombopoiesis through megakaryocyte rupture in response to acute platelet needs. J. Cell Biol. 2015, 209, 453–466. [Google Scholar] [CrossRef] [PubMed]
- Potts, K.S.; Farley, A.; Dawson, C.A.; Rimes, J.; Biben, C.; de Graaf, C.; Potts, M.A.; Stonehouse, O.J.; Carmagnac, A.; Gangatirkar, P.; et al. Membrane budding is a major mechanism of in vivo platelet biogenesis. J. Exp. Med. 2020, 217, e20191206. [Google Scholar] [CrossRef] [PubMed]
- Brown, E.; Carlin, L.M.; Nerlov, C.; Lo Celso, C.; Poole, A.W. Multiple membrane extrusion sites drive megakaryocyte migration into bone marrow blood vessels. Life Sci. Alliance 2018, 1, e201800061. [Google Scholar] [CrossRef] [PubMed]
- Eckly, A.; Scandola, C.; Oprescu, A.; Michel, D.; Rinckel, J.Y.; Proamer, F.; Hoffmann, D.; Receveur, N.; Leon, C.; Bear, J.E.; et al. Megakaryocytes use in vivo podosome-like structures working collectively to penetrate the endothelial barrier of bone marrow sinusoids. J. Thromb. Haemost. 2020. [Google Scholar] [CrossRef] [PubMed]
- Sugiyama, T.; Kohara, H.; Noda, M.; Nagasawa, T. Maintenance of the hematopoietic stem cell pool by CXCL12-CXCR4 chemokine signaling in bone marrow stromal cell niches. Immunity 2006, 25, 977–988. [Google Scholar] [CrossRef] [PubMed]
- Gomariz, A.; Helbling, P.M.; Isringhausen, S.; Suessbier, U.; Becker, A.; Boss, A.; Nagasawa, T.; Paul, G.; Goksel, O.; Szekely, G.; et al. Quantitative spatial analysis of haematopoiesis-regulating stromal cells in the bone marrow microenvironment by 3D microscopy. Nat. Commun. 2018, 9, 2532. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Varas, F.; Stadtfeld, M.; Heck, S.; Faust, N.; Graf, T. CD41-YFP mice allow in vivo labeling of megakaryocytic cells and reveal a subset of platelets hyperreactive to thrombin stimulation. Exp. Hematol. 2007, 35, 490–499. [Google Scholar] [CrossRef]
- Ding, L.; Morrison, S.J. Haematopoietic stem cells and early lymphoid progenitors occupy distinct bone marrow niches. Nature 2013, 495, 231–235. [Google Scholar] [CrossRef]
- Semeniak, D.; Kulawig, R.; Stegner, D.; Meyer, I.; Schwiebert, S.; Bösing, H.; Eckes, B.; Nieswandt, B.; Schulze, H. Proplatelet formation is selectively inhibited by collagen type I through Syk-independent GPVI signaling. J. Cell Sci. 2016, 129, 3473–3484. [Google Scholar] [CrossRef] [PubMed]
- Sorokin, L.M.; Pausch, F.; Durbeej, M.; Ekblom, P. Differential expression of five laminin alpha (1-5) chains in developing and adult mouse kidney. Dev. Dyn. 1997, 210, 446–462. [Google Scholar] [CrossRef]
- Schulze, H.; Korpal, M.; Hurov, J.; Kim, S.W.; Zhang, J.; Cantley, L.C.; Graf, T.; Shivdasani, R.A. Characterization of the megakaryocyte demarcation membrane system and its role in thrombopoiesis. Blood 2006, 107, 3868–3875. [Google Scholar] [CrossRef]
- Kokkaliaris, K.D.; Kunz, L.; Cabezas-Wallscheid, N.; Christodoulou, C.; Renders, S.; Camargo, F.; Trumpp, A.; Scadden, D.T.; Schroeder, T. Adult blood stem cell localization reflects the abundance of reported bone marrow niche cell types and their combinations. Blood J. Am. Soc. Hematol. 2020, 136, 2296–2307. [Google Scholar] [CrossRef] [PubMed]
- Omatsu, Y.; Sugiyama, T.; Kohara, H.; Kondoh, G.; Fujii, N.; Kohno, K.; Nagasawa, T. The essential functions of adipo-osteogenic progenitors as the hematopoietic stem and progenitor cell niche. Immunity 2010, 33, 387–399. [Google Scholar] [CrossRef]
- Eltoukhy, H.S.; Sinha, G.; Moore, C.; Guiro, K.; Rameshwar, P. CXCL12-abundant reticular cells (CAR) cells: A review of the literature with relevance to cancer stem cell survival. J. Cancer 2016, 4, e1004. [Google Scholar] [CrossRef]
- Avecilla, S.T.; Hattori, K.; Heissig, B.; Tejada, R.; Liao, F.; Shido, K.; Jin, D.K.; Dias, S.; Zhang, F.; Hartman, T.E. Chemokine-mediated interaction of hematopoietic progenitors with the bone marrow vascular niche is required for thrombopoiesis. Nat. Med. 2004, 10, 64–71. [Google Scholar] [CrossRef]
- Burger, J.A.; Peled, A. CXCR4 antagonists: Targeting the microenvironment in leukemia and other cancers. Leukemia 2009, 23, 43–52. [Google Scholar] [CrossRef]
- Micallef, I.N.; Jacobsen, E.D.; Shaughnessy, P.; Gandhi, P.J.; Calandra, G.; van Rhee, F.; Uberti, J. G-CSF plus plerixafor (Mozobil) to mobilize hematopoietic stem cells in patients with thrombocytopenia or leukopenia prior to auto-SCT. Bone Marrow Transplant. 2013, 48, 303–304. [Google Scholar] [CrossRef] [PubMed]
- Gustine, J.N.; Xu, L.; Yang, G.; Liu, X.; Kofides, A.; Tsakmaklis, N.; Munshi, M.; Demos, M.; Guerrera, M.L.; Meid, K.; et al. Bone marrow involvement and subclonal diversity impairs detection of mutated CXCR4 by diagnostic next-generation sequencing in Waldenström macroglobulinaemia. Br. J. Haematol. 2021. [Google Scholar] [CrossRef]
- Hernandez, P.A.; Gorlin, R.J.; Lukens, J.N.; Taniuchi, S.; Bohinjec, J.; Francois, F.; Klotman, M.E.; Diaz, G.A. Mutations in the chemokine receptor gene CXCR4 are associated with WHIM syndrome, a combined immunodeficiency disease. Nat. Genet. 2003, 34, 70–74. [Google Scholar] [CrossRef]
- Campbell, F.R. Ultrastructural Studies of Transmural Migration of Blood-Cells in Bone-Marrow of Rats, Mice and Guinea-Pigs. Am. J. Anat. 1972, 135, 521–535. [Google Scholar] [CrossRef]
- Malara, A.; Currao, M.; Gruppi, C.; Celesti, G.; Viarengo, G.; Buracchi, C.; Laghi, L.; Kaplan, D.L.; Balduini, A. Megakaryocytes contribute to the bone marrow-matrix environment by expressing fibronectin, type IV collagen, and laminin. Stem Cells 2014, 32, 926–937. [Google Scholar] [CrossRef] [PubMed]
- Semeniak, D.; Faber, K.; Oftering, P.; Manukjan, G.; Schulze, H. Impact of Itga2-Gp6-double collagen receptor deficient mice for bone marrow megakaryocytes and platelets. PLoS ONE 2019, 14, e0216839. [Google Scholar] [CrossRef] [PubMed]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wagner, N.; Mott, K.; Upcin, B.; Stegner, D.; Schulze, H.; Ergün, S. CXCL12-Abundant Reticular (CAR) Cells Direct Megakaryocyte Protrusions across the Bone Marrow Sinusoid Wall. Cells 2021, 10, 722. https://doi.org/10.3390/cells10040722
Wagner N, Mott K, Upcin B, Stegner D, Schulze H, Ergün S. CXCL12-Abundant Reticular (CAR) Cells Direct Megakaryocyte Protrusions across the Bone Marrow Sinusoid Wall. Cells. 2021; 10(4):722. https://doi.org/10.3390/cells10040722
Chicago/Turabian StyleWagner, Nicole, Kristina Mott, Berin Upcin, David Stegner, Harald Schulze, and Süleyman Ergün. 2021. "CXCL12-Abundant Reticular (CAR) Cells Direct Megakaryocyte Protrusions across the Bone Marrow Sinusoid Wall" Cells 10, no. 4: 722. https://doi.org/10.3390/cells10040722
APA StyleWagner, N., Mott, K., Upcin, B., Stegner, D., Schulze, H., & Ergün, S. (2021). CXCL12-Abundant Reticular (CAR) Cells Direct Megakaryocyte Protrusions across the Bone Marrow Sinusoid Wall. Cells, 10(4), 722. https://doi.org/10.3390/cells10040722