
Chronic Diabetic Wounds and Their Treatment with Skin Substitutes

 ,
,
Abstract
1. Introduction
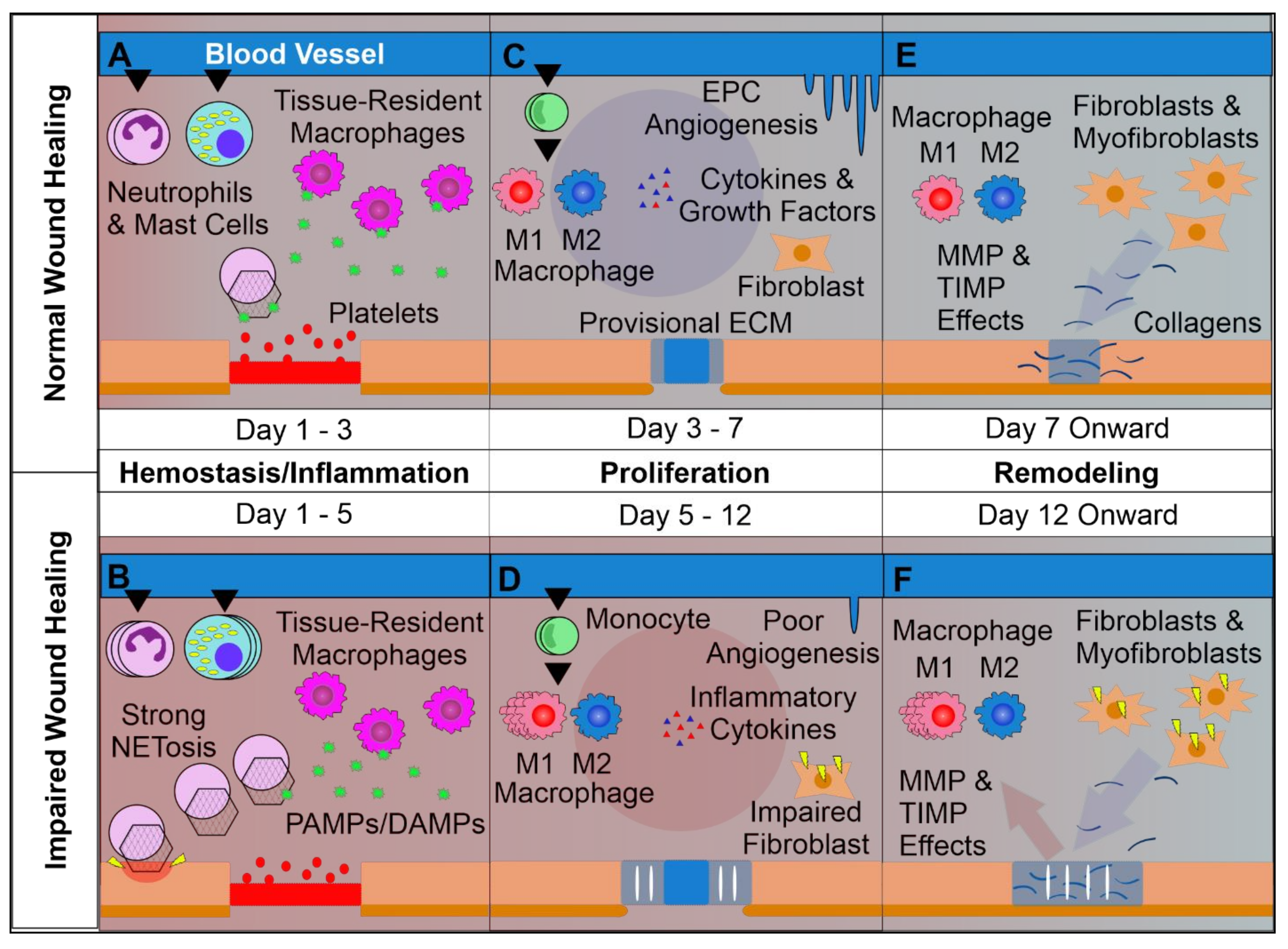
2. Wound Healing Starts with Homeostasis
3. Wound Inflammation Orchestrates Healing and Regeneration
4. Proliferation of Tissue Resident Cells Is Crucial for Wound Closure
5. Wound Remodeling
6. Treatment Strategies for Diabetic Wounds and Ulcers
7. The Use of Skin Substitutes in Diabetic Wound Healing
8. Dermal Scaffolds
9. Acellular Dermal Matrices
10. Cell-Supplemented Dermal Matrices
11. Conclusions
Author Contributions
Funding

Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Cavan, D.; Fernandez, J.D.R.; Huang, Y.; Makaroff, L. IDF releases report of global survey on access to medicines and supplies for people with diabetes. Diabetes Res. Clin. Pract. 2017, 129, 224–225. [Google Scholar]
- Schwarz, P.E.; Gallein, G.; Ebermann, D.; Müller, A.; Lindner, A.; Rothe, U.; Nebel, I.T.; Müller, G. Global Diabetes Survey: An annual report on quality of diabetes care. Diabetes Res. Clin. Pract. 2013, 100, 11–18. [Google Scholar] [CrossRef] [PubMed]
- Reiber, G.E.; McDonell, M.B.; Schleyer, A.M.; Fihn, S.D.; Reda, M.J. A comprehensive system for quality improvement in ambulatory care: Assessing the quality of diabetes care. Patient Educ. Couns. 1995, 26, 337–341. [Google Scholar] [CrossRef]
- Baltzis, D.; Eleftheriadou, I.; Veves, A. Pathogenesis and Treatment of Impaired Wound Healing in Diabetes Mellitus: New Insights. Adv. Ther. 2014, 31, 817–836. [Google Scholar] [CrossRef] [PubMed]
- Periayah, M.H.; Halim, A.S.; Saad, A.Z.M. Mechanism Action of Platelets and Crucial Blood Coagulation Pathways in Hemostasis. Int. J. Hematol. Oncol. Stem Cell Res. 2017, 11, 319–327. [Google Scholar] [PubMed]
- Ridiandries, A.; Tan, J.T.M.; Bursill, C.A. The Role of Chemokines in Wound Healing. Int. J. Mol. Sci. 2018, 19, 3217. [Google Scholar] [CrossRef]
- Gale, A.J. Continuing education course #2: Current understanding of hemostasis. Toxicol. Pathol. 2011, 39, 273–280. [Google Scholar] [PubMed]
- Reinke, J.M.; Sorg, H. Wound repair and regeneration. Eur. Surg. Res. 2012, 49, 35–43. [Google Scholar] [CrossRef]
- Molofsky, A.B.; Savage, A.K.; Locksley, R.M. Interleukin-33 in Tissue Homeostasis, Injury, and Inflammation. Immunity 2015, 42, 1005–1019. [Google Scholar] [CrossRef]
- Landén, N.X.; Li, D.; Ståhle, M. Transition from inflammation to proliferation: A critical step during wound healing. Cell. Mol. Life Sci. 2016, 73, 3861–3885. [Google Scholar] [CrossRef]
- Peiseler, M.; Kubes, P. More friend than foe: The emerging role of neutrophils in tissue repair. J. Clin. Investig. 2019, 129, 2629–2639. [Google Scholar] [CrossRef]
- Wong, S.L.; Demers, M.; Martinod, K.; Gallant, M.; Wang, Y.; Goldfine, A.B.; Kahn, C.R.; Wagner, D.D. Diabetes primes neutrophils to undergo NETosis, which impairs wound healing. Nat. Med. 2015, 21, 815–819. [Google Scholar] [CrossRef]
- Eming, S.A.; Wynn, T.A.; Martin, P. Inflammation and metabolism in tissue repair and regeneration. Science 2017, 356, 1026–1030. [Google Scholar] [CrossRef]
- Kolaczkowska, E.; Kubes, P. Neutrophil recruitment and function in health and inflammation. Nat. Rev. Immunol. 2013, 13, 159–175. [Google Scholar] [CrossRef]
- Yager, D.R.; Nwomeh, B.C. The proteolytic environment of chronic wounds. Wound Repair Regen. 1999, 7, 433–441. [Google Scholar] [CrossRef]
- Trengove, N.J.; Stacey, M.C.; Macauley, S.; Bennett, N.; Gibson, J.; Burslem, F.; Murphy, G.; Schultz, G. Analysis of the acute and chronic wound environments: The role of proteases and their inhibitors. Wound Repair Regen. 1999, 7, 442–452. [Google Scholar] [CrossRef] [PubMed]
- Rømer, J.; Bugge, T.H.; Pyke, C.; Lund, L.R.; Flick, M.J.; Degen, J.L.; Danø, K.; Oslash, R.J. Impaired wound healing in mice with a disrupted plasminogen gene. Nat. Med. 1996, 2, 287–292. [Google Scholar] [CrossRef] [PubMed]
- Etecchio, C.; Micheletti, A.; Cassatella, M.A. Neutrophil-Derived Cytokines: Facts beyond Expression. Front. Immunol. 2014, 5, 508. [Google Scholar] [CrossRef]
- Vatankhah, N.; Jahangiri, Y.; Landry, G.J.; McLafferty, R.B.; Alkayed, N.J.; Moneta, G.L.; Azarbal, A.F. Predictive value of neutrophil-to-lymphocyte ratio in diabetic wound healing. J. Vasc. Surg. 2017, 65, 478–483. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Zhou, X.; Yin, Y.; Mai, Y.; Wang, D.; Zhang, X. Hyperglycemia Induces Neutrophil Extracellular Traps Formation Through an NADPH Oxidase-Dependent Pathway in Diabetic Retinopathy. Front. Immunol. 2019, 9, 3076. [Google Scholar] [CrossRef]
- Brostjan, C.; Oehler, R. The role of neutrophil death in chronic inflammation and cancer. Cell Death Discov. 2020, 6, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Serhan, C.N.; Chiang, N.; Van Dyke, T.E. Resolving inflammation: Dual anti-inflammatory and pro-resolution lipid mediators. Nat. Rev. Immunol. 2008, 8, 349–361. [Google Scholar] [CrossRef] [PubMed]
- Widgerow, A.D. Cellular resolution of inflammation-catabasis. Wound Repair Regen. 2012, 20, 2–7. [Google Scholar] [CrossRef] [PubMed]
- Freire-De-Lima, C.G.; Xiao, Y.Q.; Gardai, S.J.; Bratton, D.L.; Schiemann, W.P.; Henson, P.M. Apoptotic Cells, through Transforming Growth Factor-β, Coordinately Induce Anti-inflammatory and Suppress Pro-inflammatory Eicosanoid and NO Synthesis in Murine Macrophages. J. Biol. Chem. 2006, 281, 38376–38384. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Gu, Z.; Lu, C.; Zhang, T.; Guo, X.; Xue, G.; Zhang, L. Neutrophil Extracellular Traps Are Markers of Wound Healing Impairment in Patients with Diabetic Foot Ulcers Treated in a Multidisciplinary Setting. Adv. Wound Care 2020, 9, 16–27. [Google Scholar] [CrossRef]
- Yan, J.; Tie, G.; Wang, S.; Tutto, A.; Demarco, N.; Khair, L.; Fazzio, T.G.; Messina, L.M. Diabetes impairs wound healing by Dnmt1-dependent dysregulation of hematopoietic stem cells differentiation towards macrophages. Nat. Commun. 2018, 9, 1–13. [Google Scholar] [CrossRef]
- Wellen, K.E.; Hotamisligil, G.S. Inflammation, stress, and diabetes. J. Clin. Investig. 2005, 115, 1111–1119. [Google Scholar] [CrossRef] [PubMed]
- Deshmane, S.L.; Kremlev, S.; Amini, S.; Sawaya, B.E. Monocyte Chemoattractant Protein-1 (MCP-1): An Overview. J. Interf. Cytokine Res. 2009, 29, 313–326. [Google Scholar] [CrossRef] [PubMed]
- Koh, T.J.; DiPietro, L.A. Inflammation and wound healing: The role of the macrophage. Expert Rev. Mol. Med. 2011, 13, e23. [Google Scholar] [CrossRef]
- Tellechea, A.; Leal, E.C.; Kafanas, A.; Auster, M.E.; Kuchibhotla, S.; Ostrovsky, Y.; Tecilazich, F.; Baltzis, D.; Zheng, Y.; Carvalho, E.; et al. Mast Cells Regulate Wound Healing in Diabetes. Diabetes 2016, 65, 2006–2019. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, A.C.D.O.; Andrade, Z.D.A.; Costa, T.F.; Medrado, A.R.A.P. Wound Healing: A Literature Review. An. Bras. Dermatol. 2016, 91, 614–620. [Google Scholar] [CrossRef]
- Pelletier, M.; Maggi, L.; Micheletti, A.; Lazzeri, E.; Tamassia, N.; Constantini, C.; Cosmi, L.; Lunardi, C.; Annunziato, F.; Romagnani, S.; et al. Evidence for a cross-talk between human neutrophils and Th17 cells. Blood 2010, 115, 335–343. [Google Scholar] [CrossRef]
- O’Garra, A.; Vieira, P.L.; Vieira, P.; Goldfeld, A.E. IL-10-producing and naturally occurring CD4+ Tregs: Limiting collateral damage. J. Clin. Investig. 2004, 114, 1372–1378. [Google Scholar] [CrossRef]
- Nosbaum, A.; Prevel, N.; Truong, H.A.; Mehta, P.; Ettinger, M.; Scharschmidt, T.C.; Ali, N.H.; Pauli, M.L.; Abbas, A.K.; Rosenblum, M.D. Cutting Edge: Regulatory T Cells Facilitate Cutaneous Wound Healing. J. Immunol. 2016, 196, 2010–2014. [Google Scholar] [CrossRef]
- Van, Y.H.; Lee, W.H.; Ortiz, S.; Lee, M.H.; Qin, H.J.; Liu, C.P. All-trans retinoic acid inhibits type 1 diabetes by T regulatory (Treg)-dependent suppression of interferon-gamma-producing T-cells without affecting Th17 cells. Diabetes 2009, 58, 146–155. [Google Scholar] [CrossRef]
- Pastar, I.; Stojadinovic, O.; Yin, N.C.; Ramirez, H.; Nusbaum, A.G.; Sawaya, A.; Patel, S.B.; Khalid, L.; Isseroff, R.R.; Tomic-Canic, M. Epithelialization in Wound Healing: A Comprehensive Review. Adv. Wound Care 2014, 3, 445–464. [Google Scholar] [CrossRef]
- Egozi, E.I.; Bs, A.M.F.; Burns, A.L.; Gamelli, R.L.; DiPietro, L.A. Mast cells modulate the inflammatory but not the proliferative response in healing wounds. Wound Repair Regen. 2003, 11, 46–54. [Google Scholar] [CrossRef]
- Tracy, L.E.; Minasian, R.A.; Caterson, E. Extracellular Matrix and Dermal Fibroblast Function in the Healing Wound. Adv. Wound Care 2016, 5, 119–136. [Google Scholar] [CrossRef] [PubMed]
- Zajac, E.; Schweighofer, B.; Kupriyanova, T.A.; Juncker-Jensen, A.; Minder, P.; Quigley, J.P.; Deryugina, E.I. Angiogenic capacity of M1- and M2-polarized macrophages is determined by the levels of TIMP-1 complexed with their secreted proMMP-9. Blood 2013, 122, 4054–4067. [Google Scholar] [CrossRef] [PubMed]
- Fadini, G.P.; Miorin, M.; Facco, M.; Bonamico, S.; Baesso, I.; Grego, F.; Menegolo, M.; de Kreutzenberg, S.V.; Tiengo, A.; Agostini, C.; et al. Circulating Endothelial Progenitor Cells Are Reduced in Peripheral Vascular Complications of Type 2 Diabetes Mellitus. J. Am. Coll. Cardiol. 2005, 45, 1449–1457. [Google Scholar] [CrossRef] [PubMed]
- António, N.; Fernandes, R.; Soares, A.; Soares, F.; Lopes, A.; Carvalheiro, T.; Paiva, A.; Pêgo, G.M.; A Providência, L.; Gonçalves, L.; et al. Reduced levels of circulating endothelial progenitor cells in acute myocardial infarction patients with diabetes or pre-diabetes: Accompanying the glycemic continuum. Cardiovasc. Diabetol. 2014, 13, 101. [Google Scholar] [CrossRef]
- Tepper, O.M.; Galiano, R.D.; Capla, J.M.; Kalka, C.; Gagne, P.J.; Jacobowitz, G.R.; Levine, J.P.; Gurtner, G.C. Human Endothelial Progenitor Cells From Type II Diabetics Exhibit Impaired Proliferation, Adhesion, and Incorporation Into Vascular Structures. Circulation 2002, 106, 2781–2786. [Google Scholar] [CrossRef]
- Demidova-Rice, T.N.; Durham, J.T.; Herman, I.M. Wound Healing Angiogenesis: Innovations and Challenges in Acute and Chronic Wound Healing. Adv. Wound Care 2012, 1, 17–22. [Google Scholar] [CrossRef]
- Bonnans, C.; Chou, J.; Werb, Z. Remodelling the extracellular matrix in development and disease. Nat. Rev. Mol. Cell Biol. 2014, 15, 786–801. [Google Scholar] [CrossRef] [PubMed]
- Kendall, R.T.; Feghali-Bostwick, C.A. Fibroblasts in fibrosis: Novel roles and mediators. Front. Pharmacol. 2014, 5, 123. [Google Scholar] [CrossRef] [PubMed]
- He, H.; Xu, J.; Warren, C.M.; Duan, D.; Li, X.; Wu, L.; Iruela-Arispe, M.L. Endothelial cells provide an instructive niche for the differentiation and functional polarization of M2-like macrophages. Blood 2012, 120, 3152–3162. [Google Scholar] [CrossRef] [PubMed]
- Eljaszewicz, A.; Wiese, M.; Helmin-Basa, A.; Jankowski, M.; Gackowska, L.; Kubiszewska, I.; Kaszewski, W.; Michalkiewicz, J.; Zegarski, W. Collaborating with the Enemy: Function of Macrophages in the Development of Neoplastic Disease. Mediat. Inflamm. 2013, 2013, 831387. [Google Scholar] [CrossRef] [PubMed]
- Vitale, I.; Manic, G.; Coussens, L.M.; Kroemer, G.; Galluzzi, L. Macrophages and Metabolism in the Tumor Microenvironment. Cell Metab. 2019, 30, 36–50. [Google Scholar] [CrossRef]
- Fu, L.Q.; Du, W.L.; Cai, M.H.; Yao, J.Y.; Zhao, Y.Y.; Mou, X.Z. The roles of tumor-associated macrophages in tumor angiogenesis and metastasis. Cell. Immunol. 2020, 353, 104119. [Google Scholar] [CrossRef]
- Kim, B.C.; Kim, H.T.; Park, S.H.; Cha, J.C.; Yufit, T.; Kim, S.J.; Falanga, V. Fibroblasts from chronic wounds show altered TGF-beta-signaling and decreased TGF-beta Type II receptor expression. J. Cell Physiol. 2003, 195, 331–336. [Google Scholar] [CrossRef]
- Bannon, P.; Wood, S.; Restivo, T.; Campbell, L.; Hardman, M.J.; Mace, K.A. Diabetes induces stable intrinsic changes to myeloid cells that contribute to chronic inflammation during wound healing in mice. Dis. Model. Mech. 2013, 6, 1434–1447. [Google Scholar] [CrossRef]
- Xue, M.; Jackson, C.J. Extracellular Matrix Reorganization during Wound Healing and Its Impact on Abnormal Scarring. Adv. Wound Care 2015, 4, 119–136. [Google Scholar] [CrossRef] [PubMed]
- Hillege, M.M.G.; Caro, R.A.G.; Offringa, C.; de Witt, G.M.J.; Jaspers, R.T.; Hoogaars, W.M.H. TGF-β Regulates Collagen Type I Expression in Myoblasts and Myotubes via Transient Ctgf and Fgf-2 Expression. Cells 2020, 9, 375. [Google Scholar] [CrossRef] [PubMed]
- Van Doren, S.R. Matrix metalloproteinase interactions with collagen and elastin. Matrix Biol. 2015, 44–46, 224–231. [Google Scholar] [CrossRef]
- Manka, S.W.; Bihan, D.; Farndale, R.W. Structural studies of the MMP-3 interaction with triple-helical collagen introduce new roles for the enzyme in tissue remodelling. Sci. Rep. 2019, 9, 1–14. [Google Scholar] [CrossRef]
- Hesketh, M.; Sahin, K.B.; West, Z.E.; Murray, R.Z. Macrophage Phenotypes Regulate Scar Formation and Chronic Wound Healing. Int. J. Mol. Sci. 2017, 18, 1545. [Google Scholar] [CrossRef]
- Luthringer, M.; Mukherjee, T.; Arguello-Angarita, M.; Granick, M.S.; Alvarez, O.M. Human-derived Acellular Dermal Matrix Grafts for Treatment of Diabetic Foot Ulcers: A Systematic Review and Meta-analysis. Wounds 2020, 32, 57–65. [Google Scholar]
- Yonehiro, L.; Burleson, G.; Sauer, V. Use of a New Acellular Dermal Matrix for Treatment of Nonhealing Wounds in the Lower Extremities of Patients with Diabetes. Wounds 2013, 25, 340–344. [Google Scholar]
- Wainwright, D.J.; Bury, S.B. Acellular dermal matrix in the management of the burn patient. Aesthet. Surg. J. 2011, 31, 13S–23S. [Google Scholar] [CrossRef]
- Yde, S.S.; Brunbjerg, M.E.; Damsgaard, T.E. Acellular dermal matrices in breast reconstructions: A literature review. J. Plast. Surg. Hand Surg. 2016, 50, 187–196. [Google Scholar]
- Guo, X.; Mu, D.; Gao, F. Efficacy and safety of acellular dermal matrix in diabetic foot ulcer treatment: A systematic review and meta-analysis. Int. J. Surg. 2017, 40, 1–7. [Google Scholar] [CrossRef]
- Hayes, E.M.; Tsaousi, A.; Di Gregoli, K.; Jenkinson, S.R.; Bond, A.R.; Johnson, J.L.; Bevan, L.; Thomas, A.C.; Newby, A.C. Classical and Alternative Activation and Metalloproteinase Expression Occurs in Foam Cell Macrophages in Male and Female ApoE Null Mice in the Absence of T and B Lymphocytes. Front. Immunol. 2014, 5. [Google Scholar] [CrossRef]
- Maione, A.G.; Smith, A.; Kashpur, O.; Yanez, V.; Knight, E.; Mooney, D.J.; Veves, A.; Tomic-Canic, M.; Garlick, J.A. Altered ECM deposition by diabetic foot ulcer-derived fibroblasts implicates fibronectin in chronic wound repair. Wound Repair Regen. 2016, 24, 630–643. [Google Scholar] [CrossRef]
- Hamed, S.; Ullmann, Y.; Egozi, D.; Daod, E.; Hellou, E.; Ashkar, M.; Gilhar, A.; Teot, L. Fibronectin Potentiates Topical Erythropoietin-Induced Wound Repair in Diabetic Mice. J. Investig. Dermatol. 2011, 131, 1365–1374. [Google Scholar] [CrossRef]
- Lewis, J.; Lipp, A. Pressure-relieving interventions for treating diabetic foot ulcers. Cochrane Database Syst. Rev. 2013, 2013. [Google Scholar] [CrossRef] [PubMed]
- Reyzelman, A.; Crews, R.T.; Moore, J.C.; Moore, L.; Mukker, J.S.; Offutt, S.; Tallis, A.; Turner, W.B.; Vayser, D.; Winters, C.; et al. Clinical effectiveness of an acellular dermal regenerative tissue matrix compared to standard wound management in healing diabetic foot ulcers: A prospective, randomised, multicentre study. Int. Wound J. 2009, 6, 196–208. [Google Scholar] [CrossRef] [PubMed]
- Perez-Favila, A.; Martinez-Fierro, M.L.; Rodriguez-Lazalde, J.G.; A Cid-Baez, M.; Zamudio-Osuna, M.D.J.; Martinez-Blanco, M.D.R.; Mollinedo-Montaño, E.F.; Rodriguez-Sanchez, I.P.; Castañeda-Miranda, R.; Garza-Veloz, I. Current Therapeutic Strategies in Diabetic Foot Ulcers. Medicina 2019, 55, 714. [Google Scholar] [CrossRef] [PubMed]
- Cazzell, S.; Moyer, P.M.; Samsell, B.; Dorsch, K.; McLean, J.; Moore, M.A. A Prospective, Multicenter, Single-Arm Clinical Trial for Treatment of Complex Diabetic Foot Ulcers with Deep Exposure Using Acellular Dermal Matrix. Adv. Ski. Wound Care 2019, 32, 409–415. [Google Scholar] [CrossRef] [PubMed]
- Walters, J.; Cazzell, S.; Pham, H.; Vayser, D.; Reyzelman, A. Healing Rates in a Multicenter Assessment of a Sterile, Room Temperature, Acellular Dermal Matrix Versus Conventional Care Wound Management and an Active Comparator in the Treatment of Full-Thickness Diabetic Foot Ulcers. Eplasty 2016, 16, e10. [Google Scholar]
- Cazzell, S.; Vayser, D.; Pham, H.; Walters, J.; Reyzelman, A.; Samsell, B.; Dorsch, K.; Moore, M. A randomized clinical trial of a human acellular dermal matrix demonstrated superior healing rates for chronic diabetic foot ulcers over conventional care and an active acellular dermal matrix comparator. Wound Repair Regen. 2017, 25, 483–497. [Google Scholar] [CrossRef] [PubMed]
- Martinson, M. A comparative analysis of skin substitutes used in the management of diabetic foot ulcers. J. Wound Care 2016, 25, S8–S17. [Google Scholar] [CrossRef]
- Snyder, R.J.; Shimozaki, K.; Tallis, A.; Kerzner, M.; Reyzelman, A.; Lintzeris, D.; Bell, D.; Rutan, R.L.; Rosenblum, B. A Prospective, Randomized, Multicenter, Controlled Evaluation of the Use of Dehy-drated Amniotic Membrane Allograft Compared to Standard of Care for the Closure of Chronic Diabetic Foot Ulcer. Wounds 2016, 28, 70–77. [Google Scholar]
- Marston, W.A.; Hanft, J.; Norwood, P.; Pollak, R. The Efficacy and Safety of Dermagraft in Improving the Healing of Chronic Diabetic Foot Ulcers: Results of a prospective randomized trial. Diabetes Care 2003, 26, 1701–1705. [Google Scholar] [CrossRef]
- Steed, D.L.; Attinger, C.; Colaizzi, T.; Rn, M.C.; Franz, M.; Harkless, L.; Bs, A.J.; Moosa, H.; Robson, M.; Serena, T.; et al. Guidelines for the treatment of diabetic ulcers. Wound Repair Regen. 2006, 14, 680–692. [Google Scholar] [CrossRef]
- Lu, D.; Jiang, Y.; Deng, W.; Zhang, Y.; Liang, Z.; Wu, Q.; Jiang, X.; Zhang, L.; Gao, F.; Cao, Y.; et al. Long-Term Outcomes of BMMSC Compared with BMMNC for Treatment of Critical Limb Is-chemia and Foot Ulcer in Patients with Diabetes. Cell Transplant. 2019, 28, 645–652. [Google Scholar] [CrossRef]
- Yang, H.Y.; Fierro, F.; So, M.; Yoon, D.J.; Nguyen, A.V.; Gallegos, A.; Bagood, M.D.; Rojo-Castro, T.; Alex, A.; Stewart, H.; et al. Combination product of dermal matrix, human mesenchymal stem cells, and timolol promotes diabetic wound healing in mice. STEM CELLS Transl. Med. 2020, 9, 1353–1364. [Google Scholar] [CrossRef]
- Hart, C.E.; Loewen-Rodriguez, A.; Lessem, J. Dermagraft: Use in the Treatment of Chronic Wounds. Adv. Wound Care 2012, 1, 138–141. [Google Scholar] [CrossRef] [PubMed]
- Boháč, M.; Danišovič, Ľ.; Koller, J.; Dragúňová, J.; Varga, I. What happens to an acellular dermal matrix after implantation in the human body? A histological and electron microscopic study. Eur. J. Histochem. 2018, 62, 2873. [Google Scholar] [CrossRef] [PubMed]
- Larsen, L.; Tchanque-Fossuo, C.N.; Gorouhi, F.; Boudreault, D.; Nguyen, C.; Fuentes, J.J.; Crawford, R.W.; Dahle, S.E.; Whetzel, T.; Isseroff, R.R. Combination therapy of autologous adipose mesenchymal stem cell-enriched, high-density lipoaspirate and topical timolol for healing chronic wounds. J. Tissue Eng. Regen. Med. 2018, 12, 186–190. [Google Scholar] [CrossRef]
- Moon, K.C.; Suh, H.S.; Kim, K.B.; Han, S.K.; Young, K.W.; Lee, J.W. Potential of Allogeneic Adipose-Derived Stem Cell–Hydrogel Complex for Treating Diabetic Foot Ulcers. Diabetes 2019, 68, 837–846. [Google Scholar] [CrossRef] [PubMed]
- Choudhury, S.; Das, A. Advances in generation of three-dimensional skin equivalents: Pre-clinical studies to clinical therapies. Cytotherapy 2021, 23, 1–9. [Google Scholar] [CrossRef]
- Álvaro-Afonso, F.J.; García-Álvarez, Y.; Lázaro-Martínez, J.L.; Kakagia, D.; Papanas, N. Advances in Dermoepidermal Skin Substitutes for Diabetic Foot Ulcers. Curr. Vasc. Pharmacol. 2020, 18, 182–192. [Google Scholar] [CrossRef]
- Greer, N.; Foman, N.; Dorrian, J.; Fitzgerald, P.; Macdonald, R.; Rutks, I.; Wilt, T. Advanced Wound Care Therapies for Non-Healing Diabetic, Venous, and Arterial Ulcers: A Systematic Review. Ann. Inter. Med. 2013, 159, 532–542. [Google Scholar] [CrossRef]
- Witherel, C.E.; Graney, P.L.; Freytes, D.O.; Weingarten, M.S.; Spiller, K.L. Response of human macrophages to wound matrices in vitro. Wound Repair Regen. 2016, 24, 514–524. [Google Scholar] [CrossRef] [PubMed]
- Sun, W.Q.; Leung, P. Calorimetric study of extracellular tissue matrix degradation and instability after gamma irradiation. Acta Biomater. 2008, 4, 817–826. [Google Scholar] [CrossRef] [PubMed]
- Gouk, S.S.; Lim, T.M.; Teoh, S.H.; Sun, W.Q. Alterations of human acellular tissue matrix by gamma irradiation: Histology, biomechanical property, stability, in vitro cell repopulation, and remodeling. J. Biomed. Mater. Res. 2007, 84, 205–217. [Google Scholar] [CrossRef]
- Postlethwaite, E.A.; Seyer, J.M.; Kang, A.H. Chemotactic attraction of human fibroblasts to type I, II, and III collagens and collagen-derived peptides. Proc. Natl. Acad. Sci. USA 1978, 75, 871–875. [Google Scholar] [CrossRef] [PubMed]
- Galt, S.W.; Lindemann, S.; Medd, D.; Allen, L.L.; Kraiss, L.W.; Harris, E.S.; Prescott, S.M.; McIntyre, T.M.; Weyrich, A.S.; Zimmerman, G.A. Differential Regulation of Matrix Metalloproteinase-9 by Monocytes Adherent to Collagen and Platelets. Circ. Res. 2001, 89, 509–516. [Google Scholar] [CrossRef] [PubMed]
- Sondag, C.M.; Combs, C.K. Adhesion of monocytes to type I collagen stimulates an APP-dependent proinflammatory signaling response and release of Aβ1-40. J. Neuroinflamm. 2010, 7, 22. [Google Scholar] [CrossRef] [PubMed]
- Khalaji, S.; Zondler, L.; Kleinjan, F.; Nolte, U.; Mulaw, M.A.; Danzer, K.M.; Weishaupt, J.H.; Gottschalk, K.-E. Age Increases Monocyte Adhesion on Collagen. Sci. Rep. 2017, 7, srep46532. [Google Scholar] [CrossRef] [PubMed]
- Schultz, H.S.; Guo, L.; Keller, P.; Fleetwood, A.J.; Sun, M.; Guo, W.; Ma, C.; Hamilton, J.A.; Bjørkdahl, O.; Berchtold, M.W.; et al. OSCAR-collagen signaling in monocytes plays a proinflammatory role and may contribute to the pathogenesis of rheumatoid arthritis. Eur. J. Immunol. 2016, 46, 952–963. [Google Scholar] [CrossRef]
- Bao, P.; Kodra, A.; Tomic-Canic, M.; Golinko, M.S.; Ehrlich, H.P.; Brem, H. The Role of Vascular Endothelial Growth Factor in Wound Healing. J. Surg. Res. 2009, 153, 347–358. [Google Scholar] [CrossRef]
- Barrientos, S.; Stojadinovic, O.; Golinko, M.S.; Brem, H.; Tomic-Canic, M. Growth factors and cytokines in wound healing. Wound Repair Regen. 2008, 16, 585–601. [Google Scholar] [CrossRef]
- Karin, M.; Clevers, H. Reparative inflammation takes charge of tissue regeneration. Nat. Cell Biol. 2016, 529, 307–315. [Google Scholar] [CrossRef]
- Larsen, A.M.H.; Kuczek, D.E.; Kalvisa, A.; Siersbæk, M.S.; Thorseth, M.-L.; Johansen, A.Z.; Carretta, M.; Grøntved, L.; Vang, O.; Madsen, D.H. Collagen Density Modulates the Immunosuppressive Functions of Macrophages. J. Immunol. 2020, 205, 1461–1472. [Google Scholar] [CrossRef]
- Gauglitz, G.G.; Korting, H.C.; Pavicic, T.; Ruzicka, T.; Jeschke, M.G. Hypertrophic Scarring and Keloids: Pathomechanisms and Current and Emerging Treatment Strategies. Mol. Med. 2010, 17, 113–125. [Google Scholar] [CrossRef]
- Paige, J.T.; Kremer, M.; Landry, J.; Hatfield, A.S.; Wathieu, D.; Brug, A.; Lightell, D.J.; Spiller, K.L.; Woods, T.C. Modulation of inflammation in wounds of diabetic patients treated with porcine urinary bladder matrix. Regen. Med. 2019, 14, 269–277. [Google Scholar] [CrossRef] [PubMed]
- Newton, D.; Khan, F.; Belch, J.; Mitchell, M.; Leese, G. Blood flow changes in diabetic foot ulcers treated with dermal replacement therapy. J. Foot Ankle Surg. 2002, 41, 233–237. [Google Scholar] [CrossRef]
- Pliszczynski, J.; Nita, M.; Kowalewski, C.; Wozniak, K.; Eljaszewicz, A.; Moniuszko, M.; Kaminski, A.; Sladowski, D.L.; Zimek, Z.; Majewski, S.; et al. Transplantation of a New Biological Product in Rare Diseases, Such as Epidermolysis Bullosa: Response and Clinical Outcome. Transplant. Proc. 2020, 52, 2239–2243. [Google Scholar] [CrossRef] [PubMed]
- Macadam, S.A.; Lennox, P.A. Acellular dermal matrices: Use in reconstructive and aesthetic breast surgery. Can. J. Plast. Surg. 2012, 20, 75–89. [Google Scholar] [CrossRef] [PubMed]
- El Masry, M.S.; Chaffee, S.; Das Ghatak, P.; Mathew-Steiner, S.S.; Das, A.; Higuita-Castro, N.; Roy, S.; Anani, R.A.; Sen, C.K. Stabilized collagen matrix dressing improves wound macrophage function and epithelialization. FASEB J. 2019, 33, 2144–2155. [Google Scholar] [CrossRef] [PubMed]
- Bergijk, E.C.; Munaut, C.; Baelde, J.J.; Prins, F.; Foidart, J.M.; Hoedemaeker, P.J.; Bruijn, J.A. A histologic study of the extracellular matrix during the development of glomerulosclerosis in murine chronic graft-versus-host disease. Am. J. Pathol. 1992, 140, 1147–1156. [Google Scholar]
- Dixit, S.; Baganizi, D.R.; Sahu, R.; Dosunmu, E.; Chaudhari, A.; Vig, K.; Pillai, S.R.; Singh, S.R.; Dennis, V.A. Immunological challenges associated with artificial skin grafts: Available solutions and stem cells in future design of synthetic skin. J. Biol. Eng. 2017, 11, 49. [Google Scholar] [CrossRef]
- Nahabedian, M.Y. Prosthetic Breast Reconstruction and Red Breast Syndrome: Demystification and a Review of the Literature. Plast. Reconstr. Surg. Glob. Open 2019, 7, e2108. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.C.; Brown, A.M.; Kulber, D.A.; Moliver, C.L.; Kuehnert, M.J. The Role of Endotoxin in Sterile Inflammation After Implanted Acellular Dermal Matrix: Red Breast Syndrome Explained? Aesthet. Surg. J. 2020, 40, 392–399. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; He, W.; Luo, G.; Wu, J. Fundamental immunology of skin transplantation and key strategies for tolerance induction. Arch. Immunol. Ther. Exp. 2013, 61, 397–405. [Google Scholar] [CrossRef] [PubMed]
- Sharma, V.; Patel, N.; Kohli, N.; Ravindran, N.; Hook, L.; Mason, C.; García-Gareta, E. Viscoelastic, physical, and bio-degradable properties of dermal scaffolds and related cell behaviour. Biomed. Mater. 2016, 11, 055001. [Google Scholar] [CrossRef]
- Nita, M.; Pliszczynski, J.; Kowalewski, C.; Wozniak, K.; Eljaszewicz, A.; Moniuszko, M.; Kaminski, A.; Sladowski, D.L.; Zimek, Z.; Majewski, S.; et al. New Treatment of Wound Healing With Allogenic Acellular Human Skin Graft: Preclinical Assessment and In Vitro Study. Transplant. Proc. 2020, 52, 2204–2207. [Google Scholar] [CrossRef]
- Greco, K.V.; Francis, L.; Somasundaram, M.; Greco, G.; English, N.R.; Roether, A.J.; Boccaccini, A.R.; Sibbons, P.; Ansari, T. Characterisation of porcine dermis scaffolds decellularised using a novel non-enzymatic method for biomedical applications. J. Biomater. Appl. 2015, 30, 239–253. [Google Scholar] [CrossRef]
- Nie, C.; Yang, D.; Xu, J.; Si, Z.; Jin, X.; Zhang, J. Locally Administered Adipose-Derived Stem Cells Accelerate Wound Healing through Differentiation and Vasculogenesis. Cell Transplant. 2011, 20, 205–216. [Google Scholar] [CrossRef]
- Nie, C.; Zhang, G.; Yang, D.; Liu, T.; Liu, D.; Xu, J.; Zhang, J. Targeted delivery of adipose-derived stem cells via acellular dermal matrix enhances wound repair in diabetic rats. J. Tissue Eng. Regen. Med. 2012, 9, 224–235. [Google Scholar] [CrossRef] [PubMed]
- Stasch, T.; Hoehne, J.; Huynh, T.; De Baerdemaeker, R.; Grandel, S.; Herold, C. Débridement and Autologous Lipotransfer for Chronic Ulceration of the Diabetic Foot and Lower Limb Improves Wound Healing. Plast. Reconstr. Surg. 2015, 136, 1357–1366. [Google Scholar] [CrossRef] [PubMed]
- Maacha, S.; Sidahmed, H.; Jacob, S.; Gentilcore, G.; Calzone, R.; Grivel, J.-C.; Cugno, C. Paracrine Mechanisms of Mesenchymal Stromal Cells in Angiogenesis. Stem Cells Int. 2020, 2020, 4356359. [Google Scholar] [CrossRef]
- Ankrum, J.A.; Ong, J.F.; Karp, J.M. Mesenchymal stem cells: Immune evasive, not immune privileged. Nat. Biotechnol. 2014, 32, 252–260. [Google Scholar] [CrossRef] [PubMed]
- Gentile, P.; Sterodimas, A.; Pizzicannella, J.; Dionisi, L.; De Fazio, D.; Calabrese, C.; Garcovich, S. Systematic Review: Allogenic Use of Stromal Vascular Fraction (SVF) and Decellularized Extracellular Matrices (ECM) as Advanced Therapy Medicinal Products (ATMP) in Tissue Regeneration. Int. J. Mol. Sci. 2020, 21, 4982. [Google Scholar] [CrossRef]


{kind=link}
| Name of Clinical TRIAL | Last Update | Clinical Trial ID | Status | Conclusions | Publications (PMID) | |
|---|---|---|---|---|---|---|
| Acellular Dermal Matrices | Effect of Meso Wound Matrix in the Treatment of DFUs | 22 October 2020 | NCT04182451 | Active, not recruiting | No results available | |
| Comparative Effectiveness of Two Acellular Matrices (Dermacell vs. Integra) for Management of Deep Diabetic Foot Ulcers | 2 September 2020 | NCT03476876 | Recruiting | No results available | ||
| DermACELL AWM® in Chronic Wagner Grade 3/4 Diabetic Foot Ulcers | 6 September 2019 | NCT03044132 | Completed | DermACELL healed complex DFUs with exposed bone. Results suggest wound closure if given time. | [68] 31361269 | |
| DermACELL in Subjects With Chronic Wounds of the Lower Extremities | 14 March 2018 | NCT01970163 | Completed | DermACELL increases in healing rates in DFUs compared with conventional care options | [69] 26933467 | |
| DermACELL-treated subjects had higher wound closure than those treated with ADM Graftjacket. | [70] 28544150 | |||||
| OASIS Wound Matrix (Oasis) Mechanism of Action | 9 June 2011 | NCT00570141 | Completed | 7 of 13 wounds closed fully after 12 weeks. | ||
| Acellular Porcine Dermal Matrix Wound Dressing in the Management of Diabetic Foot Ulcers | 7 June 2011 | NCT01353495 | Completed | Submitted; Pending | ||
| Skin Substitutes | AMNIOEXCEL® Plus vs. A Marketed Comparator vs. SOC in the Management of Diabetic Foot Ulcers | 20 July 2020 | NCT03547635 | Completed | 13,000 treatments: All matrices roughly equivalent in closure over 12 weeks. SIS & UBM healed more quickly and cost less. | [71] 27681811 |
| Half of patients treated achieved wound closure vs. none with SOC | [72] 26978860 | |||||
| Multi Center Site, Controlled Trial Comparing a Bioengineered Skin Substitute to a Human Skin Allograft | 26 June 2019 | NCT01676272 | Completed | No results available | ||
| Clinical Outcomes After Treatment With RestrataTM Wound Matrix in Diabetic Foot Ulcers (DFU) | 13 August 2018 | NCT03312595 | Completed | No results available | ||
| Pivotal Trial of Dermagraft(R) to Treat Diabetic Foot Ulcers | 21 May 2018 | NCT01181453 | Completed | Benefit for chronic DFUs >6 weeks duration | [73] 12766097 | |
| Dermagraft(R) for the Treatment of Patients With Diabetic Foot Ulcers | 21 May 2018 | NCT01181440 | Completed | Dermagraft-treated patients have better healing than SOC. | [73] 12766097 | |
| Growth Factors | BB-101 (Recombinant Human for Treatment of Diabetic Lower Leg and Foot Ulcers | 14 September 2020 | NCT03888053 | Recruiting | No results available | |
| A Randomized Trial on Platelet Rich Plasma Versus Saline Dressing of Diabetic Foot Ulcers | 16 September 2019 | NCT04090008 | Completed | No results available | ||
| Efficacy and Safety of Heberprot-P® in Patients With Advanced Diabetic Foot Ulcer in Dasman Diabetes Institute. | 4 August 2017 | NCT03239457 | Completed | No results available | ||
| A Phase 3 Clinical Trial to Assess the Effectiveness of BioChaperone PDGF-BB In the Treatment of Chronic Diabetic Foot Ulcer | 29 June 2017 | NCT02236793 | Completed | No results available | ||
| A Study Evaluating Topical Recombinant Human Vascular Endothelial Growth Factor (Telbermin) for Induction of Healing of Chronic, Diabetic Foot Ulcers | 11 May 2017 | NCT00069446 | Completed | No results available | ||
| Comparative Study of 3 Dose Regimens of BioChaperone to Becaplermin Gel for the Treatment of Diabetic Foot Ulcer | 15 December 2014 | NCT01098357 | Completed | No results available | ||
| Efficacy and Safety of rhEGF in Diabetic Foot Ulcer Patients With Uncontrolled Diabetic Mellitus | 4 August 2014 | NCT01629199 | Completed | No results available | ||
| Evaluation of the Safety Follow-up of Becaplermin or Placebo Gel Following Treatment of Chronic, Full Thickness Diabetic Ulcers | 8 June 2011 | NCT00740922 | Completed | No results available | ||
| Gene Therapy to Improve Wound Healing in Patients With Diabetes | 20 November 2007 | NCT00065663 | Completed | No results available | ||
| Misc. | Utilization of Platelet Gel for Treatment of Diabetic Foot Ulcers | 4 December 2015 | NCT02134132 | Completed | No results available | |
| Evaluation of the Effect of Vivostat Platelet Rich Fibrin (PRF) in the Treatment of Diabetic Foot Ulcers | 12 October 2011 | NCT00770939 | Completed | No results available | ||
| MMPs | Matrix Metalloproteinase-1/Tissue Inhibitor of Metalloproteinase-1 (MMP-1/TIMP-1) Ratio and Diabetic Foot Ulcers (DIAB-MMP2) | 18 December 2013 | NCT00935051 | Completed | No results available | |
| Mixed | Phase 2b Study of GAM501 in the Treatment of Diabetic Ulcers of the Lower Extremities (MATRIX) | 10 February 2010 | NCT00493051 | Completed | [74] 17199833 | |
| Stem Cells | Phase 1, Open-Label Safety Study of Umbilical Cord Lining Mesenchymal Stem Cells (Corlicyte®) to Heal Chronic Diabetic Foot Ulcers | 6 August 2020 | NCT04104451 | Recruiting | No results available | |
| Clinical Study of Adipose-derived Stem Cells in the Treatment of Diabetic Foot | 16 April 2019 | NCT03916211 | Not yet recruiting | No results available | ||
| Comparison of Autologous MSCs and Mononuclear Cells on Diabetic Critical Limb Ischemia and Foot Ulcer | 1 December 2010 | NCT00955669 | Completed | BMMSCs led to increased blood flow and wound healing compared to BMMNCs | [75] 30917698 | |
| Endothelial Progenitor Cells | Cryopreserved Human Umbilical Cord (TTAX01) for Late Stage, Complex Non-healing Diabetic Foot Ulcers (AMBULATE DFU II) | 24 November 2020 | NCT04450693 | Recruiting | No results available | |
| Cryopreserved Human Umbilical Cord (TTAX01) for Late Stage, Complex Non-healing Diabetic Foot Ulcers (AMBULATE DFU) | 5 November 2020 | NCT04176120 | Recruiting | No results available | ||
| Antibodies | The Safety, Tolerability, Pharmacokinetics, and Pharmacodynamics of UTTR1147A in Participants With Neuropathic Non-Healing Diabetic Foot Ulcers | 21 November 2018 | NCT02833389 | Completed | No results available |
| Composition | Material | Additives |
|---|---|---|
| Dermal | Autogenic | None—Fully Acellular |
| Epidermal | Allogenic | Acellular with Remnants |
| Full Skin | Xenogenic | Cellular (MSCs) |
| Synthetic | Molecular (MMPs/TIMPs/Growth Factors/Cytokines) | |
| Mixed |
| Property | Elaboration |
|---|---|
| Non-immunogenic |
|
| Bio-compatible |
|
| Regenerative |
|
| Protective |
|
| Non-pathogenic |
|
| Durable |
|
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Holl, J.; Kowalewski, C.; Zimek, Z.; Fiedor, P.; Kaminski, A.; Oldak, T.; Moniuszko, M.; Eljaszewicz, A. Chronic Diabetic Wounds and Their Treatment with Skin Substitutes. Cells 2021, 10, 655. https://doi.org/10.3390/cells10030655
Holl J, Kowalewski C, Zimek Z, Fiedor P, Kaminski A, Oldak T, Moniuszko M, Eljaszewicz A. Chronic Diabetic Wounds and Their Treatment with Skin Substitutes. Cells. 2021; 10(3):655. https://doi.org/10.3390/cells10030655
Chicago/Turabian StyleHoll, Jordan, Cezary Kowalewski, Zbigniew Zimek, Piotr Fiedor, Artur Kaminski, Tomasz Oldak, Marcin Moniuszko, and Andrzej Eljaszewicz. 2021. "Chronic Diabetic Wounds and Their Treatment with Skin Substitutes" Cells 10, no. 3: 655. https://doi.org/10.3390/cells10030655
APA StyleHoll, J., Kowalewski, C., Zimek, Z., Fiedor, P., Kaminski, A., Oldak, T., Moniuszko, M., & Eljaszewicz, A. (2021). Chronic Diabetic Wounds and Their Treatment with Skin Substitutes. Cells, 10(3), 655. https://doi.org/10.3390/cells10030655