Abstract
Macrophages are professional antigen presenting cells with intense phagocytic activity, strategically distributed in tissues and cavities. These cells are capable of responding to a wide variety of innate inflammatory stimuli, many of which are signaled by lipid mediators. The distribution of arachidonic acid (AA) among glycerophospholipids and its subsequent release and conversion into eicosanoids in response to inflammatory stimuli such as zymosan, constitutes one of the most studied models. In this work, we used liquid and/or gas chromatography coupled to mass spectrometry to study the changes in the levels of membrane glycerophospholipids of mouse peritoneal macrophages and the implication of group IVA cytosolic phospholipase A2 (cPLA2α) in the process. In the experimental model used, we observed that the acute response of macrophages to zymosan stimulation involves solely the cyclooxygenase-1 (COX-1), which mediates the rapid synthesis of prostaglandins E2 and I2. Using pharmacological inhibition and antisense inhibition approaches, we established that cPLA2α is the enzyme responsible for AA mobilization. Zymosan stimulation strongly induced the hydrolysis of AA-containing choline glycerophospholipids (PC) and a unique phosphatidylinositol (PI) species, while the ethanolamine-containing glycerophospholipids remained constant or slightly increased. Double-labeling experiments with 3H- and 14C-labeled arachidonate unambiguously demonstrated that PC is the major, if not the exclusive source, of AA for prostaglandin E2 production, while both PC and PI appeared to contribute to prostaglandin I2 synthesis. Importantly, in this work we also show that the COX-1-derived prostaglandins produced during the early steps of macrophage activation restrict tumor necrosis factor-α production. Collectively, these findings suggest new approaches and targets to the selective inhibition of lipid mediator production in response to fungal infection.
1. Introduction
When infections caused by viruses, bacteria or fungi occur, macrophages, strategically infiltrated in all tissues, respond by assembling a sequenced and coordinated set of responses to remove the pathogenic agents. Although macrophages are professional antigen presenting cells for T lymphocytes, prior to the immune synapse they also release cytokines and lipid mediators that extensively regulate the progress of inflammation and subsequently tissue remodeling and repair [1,2,3]. Eicosanoids include a class of lipid mediators derived from the metabolism of polyunsaturated fatty acids such as arachidonic acid (AA) by cyclooxygenases (COX), lipoxygenases, cytochrome P450, or non-enzymatic pathways. Among them, prostaglandins (PG) are bioactive signaling molecules derived from COX and the subsequent activity of terminal synthases on AA. Although PG biosynthesis involves several steps catalyzed by different enzymes, an important regulatory step is the hydrolysis of AA from glycerophospholipids by phospholipase A2 enzymes [4,5,6,7].
Zymosan particles are homogenates of Saccharomyces cerevisiae cell walls that have been used extensively as a model to study innate immune responses to fungal infections [8]. Although the recognition of molecular patterns associated with fungal pathogens (through pattern recognition receptors) [8,9], the triggering of phagocytosis [10,11], and the production of macrophage-derived cytokines [12,13] were described in great detail in in vivo or in vitro experimental models, less attention has been given to the study of the distribution of eicosanoid precursor fatty acids such as AA between the different membrane glycerophospholipid species [14,15], and their importance in the regulation of eicosanoid production in the early stages of fungal infection.
In previous work from our laboratory, we analyzed the cellular and molecular events triggered after the stimulation of mouse peritoneal macrophages with zymosan, either native or opsonized, focusing on the phagocytosis process and the associated lipid signaling pathways [15,16,17]. We demonstrated the involvement of several phospholipase A2 forms, each acting on distinct phospholipid pools and releasing different fatty acids [15,18,19]. Also, we showed the importance of coenzyme A-independent transacylation reactions to shape the cellular AA pools and their involvement in some macrophage responses [20,21,22]. In the current work we have utilized advanced mass spectrometry-based lipidomic approaches to characterize the immediate generation of PG and time-dependent changes of AA-containing phospholipid species in zymosan-stimulated mouse peritoneal macrophages. Using pharmacological inhibitors and antisense oligonucleotide approaches, we determined the central role that group IVA cytosolic phospholipase A2 (cPLA2α) plays in generating free AA substrate for PG production. Importantly, our data provide evidence that immediate PG production arises from the hydrolysis of choline-containing glycerophospholipids (PC) and phosphatidylinositol (PI) pools, with no clear involvement of ethanolamine-containing glycerophospholipids (PE). Our work also shows that PG levels negatively modulate the expression on tumor necrosis factor-α (TNFα) levels in activated macrophages, thus suggesting that the cPLA2α/COX-1 pathway regulates the execution of early proinflammatory responses by the macrophages.
2. Materials and Methods
2.1. Reagents
Cell culture medium was from Molecular Probes-Invitrogen (Carlsbad, CA, USA). Organic solvents (Optima® LC/MS grade) were from Fisher Scientific (Madrid, Spain). Lipid standards were from Avanti (Alabaster, AL, USA) or Cayman (Ann Arbor, MI, USA). Silicagel G thin-layer chromatography plates were from Macherey-Nagel (Düren, Germany). [5,6,8,9,11,12,14,15−3H]Arachidonic acid (180 Ci/mmol) and [1−14C]arachidonic acid (50 µCi/mmol) were from PerkinElmer (Boston, MA, USA). Inhibitors were from Cayman. All other reagents were from Sigma-Aldrich (Madrid, Spain).
2.2. Cell Culture and Stimulation Conditions
Resident peritoneal macrophages from Swiss male mice (University of Valladolid Animal House, 10–12 weeks old) were obtained by peritoneal lavage using 5 mL cold phosphate-buffered saline, and cultured in RPMI 1640 medium with 10% heat-inactivated calf serum, 100 U/mL penicillin, and 100 μg/mL streptomycin, as described elsewhere [23]. For antisense inhibition experiments, RAW264.7 macrophage-like cells were used. These cells were grown in Dulbecco’s modified Eagle’s medium supplemented with 10% (v/v) fetal bovine serum, 100 U/mL penicillin, 100 µg/mL streptomycin, and 2 mM L-glutamine at 37 °C in a humidified atmosphere of 5% CO2 at 37 °C, as previously described [24,25]. Zymosan was prepared as described [26]. Only zymosan batches that demonstrated no measurable endogenous phospholipase A2 activity, as measured by in vitro assay under different conditions [27,28,29,30], were used in this study. Cell protein was measured using a commercial kit (BioRad, Hercules, CA, USA).
For labeling of the cells with [3H]AA and [14C]AA, they were first labeled with 0.25 µCi/mL [3H]AA for 20 h and then with 0.1 µCi/mL [14C]AA for 30 min. Labeled AA that had not been incorporated into cellular lipids was removed by washing the cells four times with serum-free medium containing 0.5 mg/mL albumin. After the stimulations, the supernatants were acidified to pH 3.5 with 5 M formic acid and extracted with isopropanol/diethyl ether (1:1.5, v/v), and radiolabeled prostaglandins and free AA were separated by thin-layer chromatography, using ethyl acetate/acetone/acetic acid (90:5:1) as the mobile phase [31]. The cell monolayers were homogenized and the lipids were extracted according to Bligh and Dyer [32]. For separation of phospholipid classes, plates were run twice with chloroform/methanol/28% (w/w) ammonium hydroxide (60:37.5:4, v/v/v) as the mobile phase, using plates impregnated with boric acid [33]. The bands corresponding to the different lipid classes were scraped from the plates and their radioactive content was determined by scintillation counting using a Beckman Coulter LS6500 Liquid Scintillation Counter (Beckman, Fullerton, CA, USA).
2.3. Liquid Chromatography/Mass Spectrometry (LC/MS) Analyses of Prostaglandins
Analysis of prostaglandins by LC/MS was carried out exactly as described elsewhere [15,33], using an Agilent 1260 Infinity high-performance liquid chromatograph equipped with an Agilent G1311C quaternary pump and an Agilent G1329B Autosampler, coupled to an API2000 triple quadrupole mass spectrometer (Applied Biosystems, Carlsbad, CA, USA). Quantification was carried out by integrating the chromatographic peaks of each species and comparing with a calibration curve made with analytical standards.
2.4. Liquid Chromatography/Mass Spectrometry (LC/MS) Analyses of Phospholipids
This was carried out exactly as described elsewhere [15,22,34,35], using a high-performance liquid chromatograph equipped with a binary pump Hitachi LaChrom Elite L-2130 and a Hitachi Autosampler L-2200 (Merck, Madrid, Spain), coupled on-line to a Bruker Esquire 6000 ion-trap mass spectrometer (Bruker Daltonics, Bremen, Germany). Phospholipid molecular species were identified by multiple reaction monitoring experiments on chromatographic effluent by comparison with previously published data [15,22,34,35].
2.5. Gas chromatography/Mass Spectrometry (GC/MS) Analyses
Total lipids from approximately 107 cells were extracted according to Bligh and Dyer [32], and internal standards were added. Phospholipids were separated from neutral lipids by thin-layer chromatography, using n-hexane/diethyl ether/acetic acid (70:30:1, v/v/v) as the mobile phase [36]. The phospholipid bands were scraped from the plate, and fatty acid methyl esters were obtained from the various lipid fractions by transmethylation with 0.5 M KOH in methanol for 60 min at 37 °C [37,38,39,40]. Analysis was carried out using an Agilent 7890A gas chromatograph coupled to an Agilent 5975C mass-selective detector operated in electron impact mode (EI, 70 eV), equipped with an Agilent 7693 autosampler and an Agilent DB23 column (60 m length × 0.25 mm internal diameter × 0.15 µm film thickness) (Agilent Technologies, Santa Clara, CA, USA). Data analysis was carried out with the Agilent G1701EA MSD Productivity Chemstation software, revision E.02.00 [37,38,39,40].
2.6. iPLA2β Antisense Inhibition Studies
The iPLA2β antisense oligonucleotide used in this study has been described in previous studies from our laboratory [41,42,43,44]. The oligonucleotides used were as follows: antisense, 5′-CTC CTT CAC CCG GAA TGG GT; sense, 5′-ACC CAT TCC GGG TGA AGG AG. Both sense and antisense oligonucleotides contained phosphorothioate linkages to limit degradation. The oligonucleotides were mixed with Lipofectamine RNAiMAX (Thermo Fisher Scientific, Walthman, MA, USA) following the manufacturer’s instructions. Oligonucleotide treatment and culture conditions were not toxic for the cells as assessed by trypan blue dye exclusion and by quantitating cellular protein.
2.7. Quantitative PCR
Total RNA was extracted from the cells with TRIzol reagent (Invitrogen, Carlsbad, CA, USA) according to the manufacturer’s instructions, and 2 µg RNA was reverse transcribed using random primers and oligo d(T) and Moloney murine leukemia virus reverse transcriptase (Ambion, Austin, TX, USA). Quantitative PCR was carried out with an ABI 7500 machine (Applied Biosystems, Carlsbad, CA, USA) using Brilliant III Ultra-Fast SYBR Green qPCR Master Mix (Agilent Technologies, Santa Clara, CA, USA). Cycling conditions were as follows: 1 cycle at 95 °C for 3 min and 40 cycles at 95 °C for 12 s, 60 °C for 15 s, and 72 °C for 28 s [45]. The replicates were averaged, and fold induction was determined in ΔΔCt-based fold-change calculations, with cyclophilin A as a control [46]. Primer sequences are available upon request.
2.8. Data Analysis
The results are shown as means ± standard error of the mean and were analyzed for statistical significance by t-test (two groups) or by ANOVA (more than two groups), followed Tukey’s post hoc test, using GraphPad Prism software. A value of p < 0.05 was considered statistically significant.
3. Results
3.1. Immediate Synthesis of Prostaglandins by Macrophages
Figure 1A shows that acute stimulation of murine resident peritoneal macrophages with yeast-derived zymosan results in the immediate production of PGE2 (Figure 1A) and PGI2 (recovered as 6-keto-PGF1α; Figure 1B). These two species accounted for more than >90% of total AA metabolites produced at these short times.
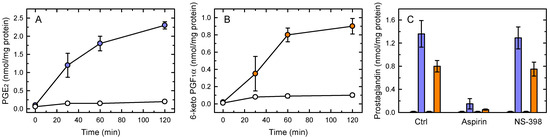
Figure 1.
PG production by zymosan-stimulated macrophages. The cells were left untreated (open symbols) or treated with 1 mg/mL zymosan (colored symbols) for the indicated times, and the levels of PGE2 (A) and 6-keto-PGF1α (B) in the supernatants were measured by LC/MS. (C) Effect of COX inhibitors on PG production. Aspirin (20 µM), NS-398 (5 µM) or neither (Ctrl) were present at the time the cells were stimulated with zymosan. After 1 h, PGE2 (purple bars) and 6-keto-PGF1α (orange bars) were measured by LC/MS. The small bars to the left of the colored bars represent the basal PG production in the absence of stimulus. The results, given as nmol PG per mg of total cell protein, are shown as means ± standard error (n = 6).
Previous studies have demonstrated that COX-1 is constitutively expressed in macrophages, whereas the COX-2 isoform is induced only after at least 2–3 h of cell stimulation [47,48]. In agreement with these observations, PG production was completely blocked by the general inhibitor aspirin, whereas the selective COX-2 inhibitor NS-398 had no discernible effect (Figure 1C). Thus these results demonstrate that immediate PG production in activated macrophages is due to COX-1.
3.2. Analysis of the AA Mobilization Response
To analyze the availability of AA substrate for PG synthesis, we measured AA mobilization by the zymosan-stimulated macrophages (Figure 2A). GC/MS analyses of AA content in the macrophages after 1 h of zymosan stimulation indicated that the cells lost 20–25% of their initial AA content, equaling to 10–15 nmol/mg protein, an amount well above what was necessary to sustain the COX-1-dependent PG production shown in Figure 1A. In turn, the data indicated that a substantial part of the released AA remains as unmetabolized free fatty acid. Figure 2A also shows that the zymosan-stimulated response was almost completely prevented by the presence in the incubation medium of the cPLA2α inhibitors pyrrophenone [49] and methyl arachidonyl fluorophosphonate (MFP) [50], highlighting the role of cPLA2α as the major mediator of the response. MFP is also a potent inhibitor of iPLA2β [51]; however the selective inhibitors bromoenol lactone (BEL) [52] and the fluoroketone FKGK18 [53] had no discernible effect, ruling out a significant role for iPLA2β in the response (Figure 2A). This conclusion was further supported by experiments utilizing cells deficient in iPLA2β by the use of an antisense oligonucleotide which we and others have previously used with success to reduce the expression of this enzyme in a variety of cells [54,55,56]. Since we were unable to find reliable antibodies against murine iPLA2β, the efficiency of antisense inhibition was analyzed by determining mRNA levels by qPCR. We detected an mRNA decrease of 60–70% (Figure 2B). iPLA2β-deficient cells, however, did not show any significant reduction of their capacity to release AA to the extracellular medium in response to zymosan (Figure 2C), providing additional evidence that the enzyme does not play a significant role in the response.
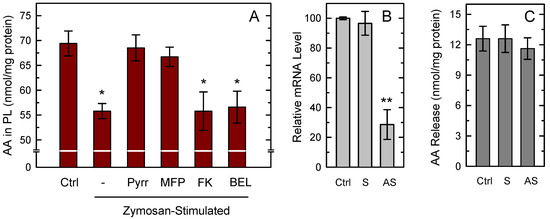
Figure 2.
AA mobilization from macrophages. (A) The cells were either unstimulated (Ctrl) or stimulated with 1 mg/mL zymosan for 1 h in the absence (-) or presence of the following inhibitors: 1 µM pyrrophenone (Pyrr), 10 µM methyl arachidonyl fluorophosphonate (MFP), 10 µM FKGK18 (FK), or 10 µM bromoenol lactone (BEL). Afterward, total content of AA in phospholipids was measured by GC/MS. Results are shown as means ± standard error of the mean (n = 6). * p < 0.05, significance of stimulated cells versus control cells. (B) The cells were treated for 36 h with sense (S) or antisense (AS) oligonucleotide, or vehicle (Ctrl), and iPLA2β mRNA levels were determined by quantitative PCR. (C) After the treatments, the cells were stimulated with zymosan, and AA release was determined by GC/MS. The fatty acid release was calculated by subtracting the amount of phospholipid-bound AA in unstimulated cells from that in stimulated cells. Results, given as nmol AA per mg of total cell protein, are shown as mean values ± standard error of the mean. (n = 4). ** p < 0.01, significantly different from control cells.
Having established the role of cPLA2α as the mediator of zymosan-stimulated release, in the next series of experiments we set out to determine whether phospholipid pools existed that selectively accounted for PG production. In the context of this work, the term phospholipid pool refers to each of the AA-containing phospholipid classes present in the cells. In the first place, we determined by LC/MS the time-course of changes in AA-containing phospholipid molecular species upon stimulation with zymosan (Figure 3). All AA-containing PC species experienced marked decreases with time. A single PI species, namely PI(18:0/20:4) also showed strong time-dependent decreases. In contrast, PE species showed little to no decreases and, as a matter of fact, some of the species slightly increased with time. It has to be noted in this regard that, during cell activation, rapid transfer of AA moieties from PC to PE occurs via CoA-independent transacylation reactions [57,58], which obscures the actual contribution of PE species to overall AA mobilization.
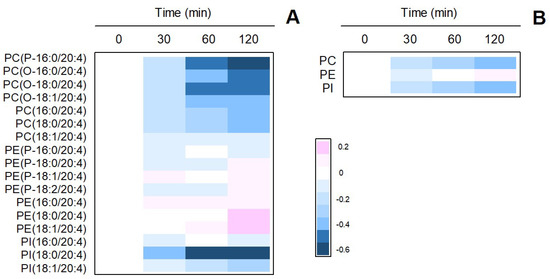
Figure 3.
Time-dependent changes of AA-containing phospholipid species in zymosan-stimulated macrophages. (A) The cells were stimulated with 1 mg/ml zymosan for the times indicated. The figure shows a heat map of all species identified by LC/MS. (B) Sum of AA-containing species within phospholipid classes. The phospholipids are designated according to Fahy et al. [59]. Fatty chains within the different phospholipid species are designated by their number of carbons followed by a colon, and the number of double bonds. A designation of O- before the first fatty chain indicates that the sn-1 position is ether linked, whereas a P- designation indicates a plasmalogen form (sn-1 vinyl ether linkage). Phospholipids containing two ester bonds have no designation. The experiment shown is representative of three independent determinations with duplicate incubations.
Collectively, the data of Figure 3 point at PC and PI as likely precursors of the AA being used for PG production during zymosan activation of the macrophages. To directly examine this possibility, we took advantage of the fact that the cellular AA phospholipids can be differentiated by double-labeling them with [3H]AA and [14C]AA at different times. The cells were first labeled with [3H]AA for 20 h, a time frame long enough to allow for the radiolabeled fatty acid to equilibrate among phospholipids and thus resemble the endogenous distribution of AA [60]. Under these conditions, the order of incorporation of radiolabel in phospholipid classes was PE > PC ≫ PI. After the 20h incubation period, the cells were pulse-labeled with [14C]AA for 30 min. At these short labeling times, the distribution of radiolabeled AA phospholipids dramatically differs from that seen at long incubation times, in that most of the radiolabel is incorporated into PC. PI and PE incorporate much lesser amounts [60]. Subsequent to the double-labeling, the cells were treated with 1 mg/mL zymosan for 1 h, and the 3H/14C ratios were determined in the phospholipid classes as well as in the prostaglandins and free AA liberated to the incubation medium (Figure 4). The 3H/14C ratio for PGE2 was very close to that of PC, strongly suggesting that PC was indeed the principal originator of the AA being converted to this PG. As for 6-keto-PGF1α, its 3H/14C ratio was intermediate between those of PC and PI, consistent with a contribution of both phospholipid classes. Note in contrast the high 3H/14C value for free AA, which was intermediate between that of PE and those of PC and PI, suggesting that free AA had been derived from all phospholipid classes (Figure 4).
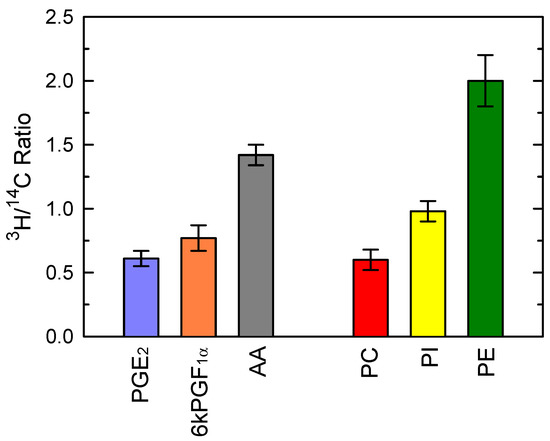
Figure 4.
Phospholipid sources of prostaglandins produced by zymosan-stimulated macrophages. The cells, labeled with both [3H]AA and [14C]AA were treated with 1 mg/mL zymosan for 1 h. Afterward, the 3H/14C ratio of extracellularly liberated PGE2, 6-keto-PGF1α and AA was calculated. The 3H/14C ratio for AA-containing PC, PI and PE is shown for comparison.
3.3. COX-1-Mediated Prostaglandin Production Regulates TNFα Production
Recent evidence suggests the existence of a regulatory interplay between the production of certain cytokines or chemokines and the synthesis/action of eicosanoids or their receptors. This is exemplified by studies showing that TNFα and PGE2 act synergistically to induce IL-8 expression [61], and that IL-10, a cytokine with proven anti-inflammatory functions, contributes to PGE2 signaling through the upregulation of the EP4 receptor [62]. Therefore it seemed reasonable to investigate whether a connection existed in our system between early PG production and cytokine expression.
Figure 5A shows that zymosan stimulation of the macrophages induced the expression of Tnf, the gene coding for the cytokine TNFα. Tnf induction occurred very rapidly, reaching a maximum at 2 h of incubation and decreasing thereafter. Given that the peak of Tnf induction occurred within the same time frame as the COX-1-dependent PG production by the macrophages, we sought to analyze the possibility of whether the two events are related. We conducted measurements in the presence of 20 µM aspirin which, as shown in Figure 1, completely ablates early PG production by the macrophages. Figure 5B shows that the presence of aspirin increased Tnf gene induction by 2-fold. Importantly, if the incubations received exogenous PGE2 (0.5–1 µM) to overcome the inhibition of COX-1 by aspirin, the Tnf induction levels dose-dependently decreased to reach the levels found in the absence of aspirin (Figure 5B). These data suggest that PGE2 produced by COX-1 during the early steps of macrophage activation restricts TNFα production in macrophages.
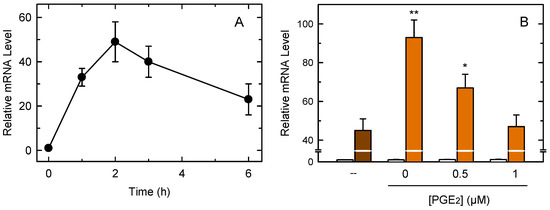
Figure 5.
Analysis of Tnf expression in macrophages. (A) The cells were treated with 1 mg/mL zymosan for different periods of time. Afterward, Tnf expression was studied by qPCR. (B) The cells were stimulated with zymosan in the absence (brown bar) or presence (orange bars) of 20 µM aspirin plus the indicated concentrations of PGE2. Open bars denote incubations in the absence of zymosan. After the 1-h incubations, Tnf expression was studied by qPCR. The results, are shown as means ± standard error (n = 6). ** p < 0.01 or * p < 0.05, significance of incubations in the presence of aspirin versus incubations in the absence of aspirin.
4. Discussion
Macrophages are known to respond to a wide variety of stimuli by mobilizing free AA and efficiently converting it to a number of oxygenated products with key roles in inflammation [4,5]. In this work we describe the early generation of PG by mouse peritoneal macrophages, a process involving COX-1, and how induction of the Tnf gene is influenced by this event. The immediate PG production by activated peritoneal macrophages is shown to involve cPLA2α activation to provide free AA, which is known to constitute a limiting factor for eicosanoid production [63,64,65].
For our studies, we have utilized yeast-derived zymosan to activate the cells. Zymosan has been widely used for many years to investigate the phospholipase A2-dependent pathways for lipid mediator production in murine peritoneal macrophages [66,67,68,69,70]. These cells have been found to contain high amounts of AA in membrane phospholipids; 20–25% of total fatty acid content is AA [71,72]. Importantly, macrophages also exhibit a characteristic distribution of AA among phospholipids, with PE, not PC, constituting the richest AA containing class, and PI containing much lesser amounts than PE or PC [18,21]. Among molecular species, the ethanolamine plasmalogens are markedly enriched with AA [56,73]. Further, AA does not distribute uniformly among membranes. Rather, specific phospholipid pools appear to exist that may accumulate AA at specific membrane locations within the cell [57,74]. There is now abundant evidence that the multiple phospholipase A2s present in cells mobilize AA for eicosanoid biosynthesis with different spatial and temporal characteristics [75]. Thus, not all cellular AA pools may be accessible to the same phospholipases. Hence AA compartmentalization may also constitute another limiting factor for eicosanoid biosynthesis.
Given this asymmetric distribution of AA in cells, in this work we considered the intriguing possibility that, depending on the phospholipid source of free AA, certain eicosanoids could be produced in preference over others. We explored this point by taking advantage of the fact that the incorporation and distribution of AA between cellular AA pools greatly depends upon the time of incubation of the cells with the fatty acid. At short incubation times (up to 2 h), the phospholipid classes that incorporate most of the exogenous fatty acid are PC and PI, whereas PE is labeled more slowly [60,76]. However, at long incubation times (>6 h), PE becomes the major AA-containing class due to the continuing transfer of AA moieties from PC to PE via CoA-independent transacylation reactions [60,76]. Thus, by labeling the cells with [14C]AA at short incubation times and [3H]AA at long incubation times, we could establish a different 3H/14C ratio for each phospholipid class [50,77].
By comparing the 3H/14C ratios of PGE2 and 6-keto-PGF1α with those of the various phospholipid classes, it was possible to establish precursor/product relationships. Although the interpretation of these data may be complicated by the phenomenon of mixing AA pools as well as the molecular heterogeneity of each phospholipid class [50], some definite conclusions can be drawn. The finding that the 3H/14C ratio for PGE2 is almost identical to that of PC strongly suggests that this PG arises mostly, if not exclusively, from the AA liberated from PC. As for 6-keto-PGF1α, the stable product of PGI2, its 3H/14C ratio is intermediate between those of PC and PI, suggesting that both of these pools participate in the process, albeit a contribution of PE, if minor, cannot be ruled out.
In mouse resident peritoneal macrophages, PE constitutes the major AA-containing class (45–50% of total cellular AA), followed by PC (35–40%), and PI (5–15%) [18,21]. Note that the 3H/14C ratio for extracellular free AA is considerably higher than that of the prostaglandins, thus suggesting a substantial contribution of PE, in addition to PC and PI, to AA release. Because cPLA2α is responsible for AA mobilization under these conditions, the participation of all kinds of phospholipids to overall AA release is fully consistent with the view that this enzyme does not distinguish among phospholipid head groups [78].
We stress that our double-labeling approach does not allow to quantify the relative contribution of each phospholipid class to overall AA release, which could be highly dependent on the absolute amounts of AA in each phospholipid class and their cellular compartmentalization. Collectively however, the data highlight the central role that PC plays in PG production, thus providing strong support to the concept that specific phospholipid pools are linked to the formation of specific eicosanoids. These conclusions are consistent with previous work in neutrophils [79] and macrophages [15] suggesting that PC molecular species are major donors of the AA used for the formation of lipoxygenase products, and also with comprehensive lipidomic studies in macrophages [14] suggesting as well the importance of PC as a major AA source for eicosanoid biosynthesis. Importantly, our work also highlights another striking function for AA-linked PC in activated cells, i.e., to donate AA moieties to replenish the fatty acid that is lost from PE species via direct transacylation. While the physiological and/or pathophysiological consequences of such transfer are yet to be fully established, it seems likely that the reaction may constitute an important point of control of the whole eicosanoid response, because diverting AA from PC to PE prevents the fatty acid from being used by cPLA2α to feed COX-1 for immediate PG production. The relative importance of these two AA fluxes arising from PC, i.e., cPLA2α-mediated release of AA versus direct channeling to PE, may be determined by the nature of stimulus and activation conditions.
The direct transacylation of AA moieties from PC to PE explains well our finding that, although PE contributes to early AA mobilization, the overall amount of AA in this phospholipid class remains relatively constant. The finding that the AA released from PE appears not to contribute significantly to early PG production by the activated macrophages was unanticipated. However, this result would be fully consistent with the view that maintaining high quantities of AA within PE phospholipid species may not be necessarily related to regulatory aspects of AA homeostasis and eicosanoid metabolism but to biophysical effects and interactions of AA-containing PE molecules with other membrane components to sustain different biological responses [80]. We have recently shown that the plasmalogen subclass of AA-containing PE participates in the execution of certain macrophage response such as bacterial lipopolysaccharide priming [20] or phagocytosis [17], but not others such as eicosanoid synthesis (this study). As discussed elsewhere [81,82], the relative content of AA-containing PE, especially the plasmalogen subclass, which is frequently found in lipid rafts [83,84,85], may affect key cellular properties such as fluidity, tendency to fusion, packing, thickness, and density, thereby influencing membrane transport and transmembrane signaling. Moreover, recent research has placed certain AA-containing PE phospholipid species at the center of the cellular machinery leading to ferroptotic cell death [86,87]. The relevance of some of these phospholipid species to neurodegenerative and neurodevelopmental disorders has also been emphasized [88]. It is conceivable that the AA deacylation/reacylation processes that PE molecular species are engaged in during cellular activation may be related to remodeling processes that are needed to place particular AA-containing PE species in the appropriate cellular compartments. This would lead to interaction with specific cellular components that lead to the execution of specific responses.
The other striking feature of the present work is the finding that early PG production modulates the extent of TNFα induction. Macrophages possess the ability to generate significant PG amounts immediately after exposure to stimuli, as shown in the present study. This effect has been attributed to the relatively high level of COX-1 expression of macrophages compared to many other cell types [47]. Our results agree with this view because pharmacological inhibition of COX-1, but not COX-2, almost completely abolishes PG production. Importantly, Tnf induction is markedly elevated when COX-1 is inhibited, and the effect can be reversed by addition of exogenous PGE2 to the incubation media. These results highlight a direct relationship between the two responses. While exploring the specific target(s) within the molecular machinery of Tnf induction through which prostaglandins exert their effect falls outside of the scope of the present work, we speculate that, in analogy with other studies [89,90], prostaglandins, acting through their specific receptors, may increase the intracellular cAMP concentration, which in turn leads to reduced gene expression. Exploring this issue will be the focus of future work from our laboratory. The finding that the early induction of cytokines such as TNFα is negatively regulated by prostaglandins may have a significant impact on the overall inflammatory response of the macrophages to immune innate stimuli, and provides a pharmacological target to manipulate the extent of such response. In turn, since the PG response depends on the supply of free AA by cPLA2α activation, our results also highlight the key role that this enzyme plays in the early macrophage responses to microbial infection that help modulate the expression of genes involved in the inflammatory response.
5. Conclusions
The possibility that distinct AA-containing glycerophospholipid pools, including the plasmalogens, are used by inflammatory stimuli to elicit a specific eicosanoid response, constitutes an intriguing current line of research. In this paper we contribute to strengthening this concept by showing that PC is a key source of AA used for the immediate synthesis of PGE2, and likely also of PGI2. Thus the compartmentalized distribution of AA among membrane phospholipid classes may constitute an effective means to regulate both the levels and nature of eicosanoids produced under different stimulatory conditions. Furthermore, our data also reinforce the important role that cPLA2α and COX-1 play in the innate inflammatory response by modulating the early expression of genes such as Tnf, thus contributing to limit inflammation. Altogether, these results provide novel information to increase our understanding of the cellular pathways that regulate AA bioavailability, and its subsequent conversion to eicosanoids. In addition, the data help to improve our understanding of the host defense to fungal aggression, which may facilitate the development of therapies to treat these challenging and difficult to treat disorders.
Author Contributions
Conceptualization, J.B.; Data curation, A.M.A., J.P.R. and C.G.; Formal analysis, A.M.A., J.P.R., C.G. and J.M.R.; Funding acquisition, M.A.B. and J.B.; Investigation, A.M.A., J.P.R., C.G. and J.M.R.; Methodology, A.M.A., J.P.R., C.G., J.M.R. and J.B.; Project administration, M.A.B. and J.B.; Resources, M.A.B. and J.B.; Supervision, M.A.B. and J.B.; Validation, A.M.A., J.P.R., C.G., M.A.B. and J.B.; Writing–original draft, J.P.R.and J.B.; Writing–review & editing, A.M.A., J.P.R., C.G., M.A.B. and J.B. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Spanish Ministry of Science and Innovation, grant number PID2019-105989RB-I00. CIBERDEM is an initiative of Instituto de Salud Carlos III.
Institutional Review Board Statement
All procedures involving animals were carried out under the supervision of the Institutional Committee of Animal Care and Usage of the University of Valladolid (approval number 7406000; date: 04/19/2016; renewed: 10/08/2019), and are in accordance with the guidelines established by the Spanish Ministry of Agriculture, Food, and Environment and the European Union.
Data Availability Statement
Data are contained within the article or supplementary material.
Acknowledgments
We thank Montse Duque for excellent technical assistance.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
Abbreviations
| AA | arachidonic acid |
| COX | cyclooxygenase |
| GC/MS | gas chromatography coupled to mass spectrometry |
| LC/MS | liquid chromatography coupled to mass spectrometry |
| cPLA2α | group IVA cytosolic phospholipase A2 |
| iPLA2β | group VIA calcium-independent phospholipase A2 |
| PC | choline-containing glycerophospholipids |
| PE | ethanolamine-containing glycerophospholipids |
| PI | phosphatidylinositol |
| PG | prostaglandin |
| TNFα | tumor necrosis factor-α |
References
- Murray, P.J.; Wynn, T.A. Protective and pathogenic functions of macrophage subsets. Nat. Rev. Immunol. 2011, 11, 723–737. [Google Scholar] [CrossRef]
- Ginhoux, F.; Jung, S. Monocytes and macrophages: Developmental pathways and tissue homeostasis. Nat. Rev. Immunol. 2014, 14, 392–404. [Google Scholar] [CrossRef]
- Stumpo, R.; Kauer, M.; Martin, S.; Kolb, H. Alternative activation of macrophages by IL-10. Pathobiology 1999, 67, 245–248. [Google Scholar] [CrossRef] [PubMed]
- Dennis, E.A.; Norris, P.C. Eicosanoid storm in infection and inflammation. Nat. Rev. Immunol. 2015, 15, 511–523. [Google Scholar] [CrossRef] [PubMed]
- Astudillo, A.M.; Balgoma, D.; Balboa, M.A.; Balsinde, J. Dynamics of arachidonic acid mobilization by inflammatory cells. Biochim. Biophys. Acta 2012, 1821, 249–256. [Google Scholar] [CrossRef] [PubMed]
- Guijas, C.; Rodríguez, J.P.; Rubio, J.M.; Balboa, M.A.; Balsinde, J. Phospholipase A2 regulation of lipid droplet formation. Biochim. Biophys. Acta 2014, 1841, 1661–1671. [Google Scholar] [CrossRef] [PubMed]
- Buczynski, M.W.; Dumlao, D.S.; Dennis, E.A. An integrated omics analysis of eicosanoid biology. J. Lipid Res. 2009, 50, 1015–1038. [Google Scholar] [CrossRef]
- Underhill, D.M. Macrophage recognition of zymosan particles. J. Endotoxin Res. 2003, 9, 176–180. [Google Scholar] [CrossRef]
- Taylor, P.R.; Brown, G.D.; Herre, J.; Williams, D.L.; Willment, J.A.; Gordon, S. The Role of SIGNR1 and the β-glucan receptor (dectin-1) in the nonopsonic recognition of yeast by specific macrophages. J. Immunol. 2004, 172, 1157–1162. [Google Scholar] [CrossRef] [PubMed]
- Goodridge, H.S.; Underhill, D.M.; Touret, N. Mechanisms of Fc receptor and dectin-1 activation for phagocytosis. Traffic 2012, 13, 1062–1071. [Google Scholar] [CrossRef] [PubMed]
- Goodridge, H.S.; Reyes, C.N.; Becker, C.A.; Katsumoto, T.R.; Ma, J.; Wolf, A.J.; Bose, N.; Chan, A.S.H.; Magee, A.S.; Danielson, M.E.; et al. Activation of the innate immune receptor dectin-1 upon formation of a “phagocytic synapse”. Nature 2011, 472, 471–475. [Google Scholar] [CrossRef]
- Walachowski, S.; Tabouret, G.; Foucras, G. Triggering dectin-1-pathway alone is not sufficient to induce cytokine production by murine macrophages. PLoS One 2016, 11, 1–26. [Google Scholar] [CrossRef]
- Takahara, K.; Tokieda, S.; Nagaoka, K.; Inaba, K. Efficient capture of Candida albicans and zymosan by SIGNR1 augments TLR2-dependent TNF-α production. Int. Immunol. 2012, 24, 89–96. [Google Scholar] [CrossRef]
- Rouzer, C.A.; Ivanova, P.T.; Byrne, M.O.; Milne, S.B.; Brown, H.A.; Marnett, L.J. Lipid profiling reveals glycerophospholipid remodeling in zymosan-stimulated macrophages. Biochemistry 2007, 46, 6026–6042. [Google Scholar] [CrossRef] [PubMed]
- Gil-de-Gómez, L.; Astudillo, A.M.; Guijas, C.; Magrioti, V.; Kokotos, G.; Balboa, M.A.; Balsinde, J. Cytosolic group IVA and calcium-independent group VIA phospholipase A2s act on distinct phospholipid pools in zymosan-stimulated mouse peritoneal macrophages. J. Immunol. 2014, 192, 752–762. [Google Scholar] [CrossRef] [PubMed]
- Gil-de-Gómez, L.; Astudillo, A.M.; Meana, C.; Rubio, J.M.; Guijas, C.; Balboa, M.A.; Balsinde, J. A phosphatidylinositol species acutely generated by activated macrophages regulates innate immune responses. J. Immunol. 2013, 190, 5169–5177. [Google Scholar] [CrossRef] [PubMed]
- Rubio, J.M.; Astudillo, A.M.; Casas, J.; Balboa, M.A.; Balsinde, J. Regulation of phagocytosis in macrophages by membrane ethanolamine plasmalogens. Front. Immunol. 2018, 9, 1723. [Google Scholar] [CrossRef] [PubMed]
- Monge, P.; Garrido, A.; Rubio, J.M.; Magrioti, V.; Kokotos, G.; Balboa, M.A.; Balsinde, J. The contribution of cytosolic group IVA and calcium-independent group VIA phospholipase A2s to adrenic acid mobilization in murine macrophages. Biomolecules 2020, 10, 542. [Google Scholar] [CrossRef] [PubMed]
- Astudillo, A.M.; Meana, C.; Bermúdez, M.A.; Pérez-Encabo, A.; Balboa, M.A.; Balsinde, J. Release of anti-inflammatory palmitoleic acid and its positional isomers by mouse peritoneal macrophages. Biomedicines 2020, 8, 480. [Google Scholar] [CrossRef] [PubMed]
- Gil-de-Gómez, L.; Astudillo, A.M.; Lebrero, P.; Balboa, M.A.; Balsinde, J. Essential role for ethanolamine plasmalogen hydrolysis in bacterial lipopolysaccharide priming of macrophages for enhanced arachidonic acid release. Front. Immunol. 2017, 8, 1251. [Google Scholar] [CrossRef] [PubMed]
- Gil-de-Gómez, L.; Monge, P.; Rodríguez, J.P.; Astudillo, A.M.; Balboa, M.A.; Balsinde, J. Phospholipid arachidonic acid remodeling during phagocytosis in mouse peritoneal macrophages. Biomedicines 2020, 8, 274. [Google Scholar] [CrossRef]
- Astudillo, A.M.; Pérez-Chacón, G.; Meana, C.; Balgoma, D.; Pol, A.; del Pozo, M.A.; Balboa, M.A.; Balsinde, J. Altered arachidonate distribution in macrophages from caveolin-1 null mice leading to reduced eicosanoid synthesis. J. Biol. Chem. 2011, 286, 35299–35307. [Google Scholar] [CrossRef] [PubMed]
- Balsinde, J.; Fernández, B.; Diez, E. Regulation of arachidonic acid release in mouse peritoneal macrophages. The role of extracellular calcium and protein kinase C. J. Immunol. 1990, 144, 4298–4304. [Google Scholar] [PubMed]
- Ruipérez, V.; Astudillo, M.A.; Balboa, M.A.; Balsinde, J. Coordinate regulation of TLR-mediated arachidonic acid mobilization in macrophages by group IVA and group V phospholipase A2s. J. Immunol. 2009, 182, 3877–3883. [Google Scholar] [CrossRef] [PubMed]
- Pindado, J.; Balsinde, J.; Balboa, M.A. TLR3-dependent induction of nitric oxide synthase in RAW 264.7 macrophage-like cells via a cytosolic phospholipase A2/cyclooxygenase-2 pathway. J. Immunol. 2007, 179, 4821–4828. [Google Scholar] [CrossRef]
- Balsinde, J.; Balboa, M.A.; Dennis, E.A. Identification of a third pathway for arachidonic acid mobilization and prostaglandin production in activated P388D1 macrophage-like cells. J. Biol. Chem. 2000, 275, 22544–22549. [Google Scholar] [CrossRef]
- Balboa, M.A.; Pérez, R.; Balsinde, J. Amplification mechanisms of inflammation: Paracrine stimulation of arachidonic acid mobilization by secreted phospholipase A2 is regulated by cytosolic phospholipase A2-derived hydroperoxyeicosatetraenoic acid. J. Immunol. 2003, 171, 989–994. [Google Scholar] [CrossRef]
- Balsinde, J.; Balboa, M.A.; Insel, P.A.; Dennis, E.A. Differential regulation of phospholipase D and phospholipase A2 by protein kinase C in P388D1 macrophages. Biochem. J. 1997, 321, 805–809. [Google Scholar] [CrossRef]
- Balboa, M.A.; Balsinde, J.; Dennis, E.A. Involvement of phosphatidate phosphohydrolase in arachidonic acid mobilization in human amnionic WISH cells. J. Biol. Chem. 1998, 273, 7684–7690. [Google Scholar] [CrossRef] [PubMed]
- Balboa, M.A.; Balsinde, J.; Dillon, D.A.; Carman, G.M.; Dennis, E.A. Proinflammatory macrophage-activating properties of the novel phospholipid diacylglycerol pyrophosphate. J. Biol. Chem. 1999, 274, 522–526. [Google Scholar] [CrossRef] [PubMed]
- Smith, B.J.; Ross, R.M.; Ayers, C.R.; Wills, M.R.; Savory, J. Rapid separation of prostaglandins by linear high performance thin layer chromatography. J. Liquid. Chromatogr. 1983, 6, 1265–1272. [Google Scholar] [CrossRef]
- Bligh, E.G.; Dyer, W.J. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef]
- Fine, J.B.; Sprecher, H. Unidimensional thin-layer chromatography of phospholipids on boric acid-impregnated plates. J. Lipid Res. 1982, 23, 660–663. [Google Scholar] [CrossRef]
- Rubio, J.M.; Rodríguez, J.P.; Gil-de-Gómez, L.; Guijas, C.; Balboa, M.A.; Balsinde, J. Group V secreted phospholipase A2 is up-regulated by interleukin-4 in human macrophages and mediates phagocytosis via hydrolysis of ethanolamine phospholipids. J. Immunol. 2015, 194, 3327–3339. [Google Scholar] [CrossRef]
- Balgoma, D.; Astudillo, A.M.; Pérez-Chacón, G.; Montero, O.; Balboa, M.A.; Balsinde, J. Markers of monocyte activation revealed by lipidomic profiling of arachidonic acid-containing phospholipids. J. Immunol. 2010, 184, 3857–3865. [Google Scholar] [CrossRef]
- Diez, E.; Balsinde, J.; Aracil, M.; Schüller, A. Ethanol induces release of arachidonic acid but not synthesis of eicosanoids in mouse peritoneal macrophages. Biochim. Biophys. Acta 1987, 921, 82–89. [Google Scholar] [CrossRef]
- Astudillo, A.M.; Pérez-Chacón, G.; Balgoma, D.; Gil-de-Gómez, L.; Ruipérez, V.; Guijas, C.; Balboa, M.A.; Balsinde, J. Influence of cellular arachidonic acid levels on phospholipid remodeling and CoA-independent transacylase activity in human monocytes and U937 cells. Biochim. Biophys. Acta 2011, 1811, 97–103. [Google Scholar] [CrossRef] [PubMed]
- Valdearcos, M.; Esquinas, E.; Meana, C.; Gil-de-Gómez, L.; Guijas, C.; Balsinde, J.; Balboa, M.A. Subcellular localization and role of lipin-1 in human macrophages. J. Immunol. 2011, 186, 6004–6013. [Google Scholar] [CrossRef] [PubMed]
- Guijas, C.; Pérez-Chacón, G.; Astudillo, A.M.; Rubio, J.M.; Gil-de-Gómez, L.; Balboa, M.A.; Balsinde, J. Simultaneous activation of p38 and JNK by arachidonic acid stimulates the cytosolic phospholipase A2-dependent synthesis of lipid droplets in human monocytes. J. Lipid Res. 2012, 53, 2343–2354. [Google Scholar] [CrossRef] [PubMed]
- Guijas, C.; Meana, C.; Astudillo, A.M.; Balboa, M.A.; Balsinde, J. Foamy monocytes are enriched in cis-7-hexadecenoic fatty acid (16:1n-9), a possible biomarker for early detection of cardiovascular disease. Cell Chem. Biol. 2016, 23, 689–699. [Google Scholar] [CrossRef]
- Balboa, M.A.; Balsinde, J. Involvement of calcium-independent phospholipase A2 in hydrogen peroxide-induced accumulation of free fatty acids in human U937 cells. J. Biol. Chem. 2002, 277, 40384–40389. [Google Scholar] [CrossRef]
- Balboa, M.A.; Sáez, Y.; Balsinde, J. Calcium-independent phospholipase A2 is required for lysozyme secretion in U937 promonocytes. J. Immunol. 2003, 170, 5276–5280. [Google Scholar] [CrossRef] [PubMed]
- Pérez, R.; Melero, R.; Balboa, M.A.; Balsinde, J. Role of group VIA calcium-independent phospholipase A2 in arachidonic acid release, phospholipid fatty acid incorporation, and apoptosis in U937 cells responding to hydrogen peroxide. J. Biol. Chem. 2004, 279, 40385–40391. [Google Scholar] [CrossRef]
- Pérez, R.; Balboa, M.A.; Balsinde, J. Involvement of group VIA calcium-independent phospholipase A2 in macrophage engulfment of hydrogen peroxide-treated U937 cells. J. Immunol. 2006, 176, 2555–2561. [Google Scholar] [CrossRef]
- Valdearcos, M.; Esquinas, E.; Meana, C.; Peña, L.; Gil-de-Gómez, L.; Balsinde, J.; Balboa, M.A. Lipin-2 reduces proinflammatory signaling induced by saturated fatty acids in macrophages. J. Biol. Chem. 2012, 287, 10894–10904. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔ Ct method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Rouzer, C.A.; Kingsley, P.J.; Wang, H.; Zhang, H.; Morrow, J.D.; Dey, S.K.; Marnett, L.J. Cyclooxygenase-1-dependent prostaglandin synthesis modulates tumor necrosis factor-α secretion in lipopolysaccharide-challenged murine resident peritoneal macrophages. J. Biol. Chem. 2004, 279, 34256–34268. [Google Scholar] [CrossRef]
- Suram, S.; Silveira, L.J.; Mahaffey, S.; Brown, G.D.; Bonventre, J.V.; Williams, D.L.; Gow, N.A.R.; Bratton, D.L.; Murphy, R.C.; Leslie, C.C. Cytosolic phospholipase A2α and eicosanoids regulate expression of genes in macrophages involved in host defense and inflammation. PLoS ONE 2013, 8, e69002. [Google Scholar] [CrossRef] [PubMed]
- Ono, T.; Yamada, K.; Chikazawa, Y.; Ueno, M.; Nakamoto, S.; Okuno, T.; Seno, K. Characterization of a novel inhibitor of cytosolic phospholipase A2α, pyrrophenone. Biochem. J. 2002, 363, 727–735. [Google Scholar] [CrossRef] [PubMed]
- Balsinde, J.; Dennis, E.A. Distinct roles in signal transduction for each of the phospholipase A2 enzymes present in P388D1 macrophages. J. Biol. Chem. 1996, 271, 6758–6765. [Google Scholar] [CrossRef]
- Shirai, Y.; Balsinde, J.; Dennis, E.A. Localization and functional interrelationships among cytosolic group IV, secreted group V, and Ca2+-independent group VI phospholipase A2s in P388D1 macrophages using GFP/RFP constructs. Biochim. Biophys. Acta 2005, 1735, 119–129. [Google Scholar] [CrossRef]
- Balsinde, J.; Balboa, M.A. Cellular regulation and proposed biological functions of group VIA calcium-independent phospholipase A2 in activated cells. Cell. Signal. 2005, 17, 1052–1062. [Google Scholar] [CrossRef]
- Bone, R.N.; Gai, Y.; Magrioti, V.; Kokotou, M.G.; Ali, T.; Lei, X.; Tse, H.M.; Kokotos, G.; Ramanadham, S. Inhibition of Ca2+-independent phospholipase A2β (iPLA2β) ameliorates islet infiltration and incidence of diabetes in NOD mice. Diabetes 2015, 64, 541–554. [Google Scholar] [CrossRef] [PubMed]
- Balsinde, J.; Balboa, M.A.; Dennis, E.A. Antisense inhibition of group VI Ca2+-independent phospholipase A2 blocks phospholipid fatty acid remodeling in murine P388D1 macrophages. J. Biol. Chem. 1997, 272, 29317–29321. [Google Scholar] [CrossRef] [PubMed]
- Carnevale, K.A.; Cathcart, M.K. Calcium-independent phospholipase A2 is required for human monocyte chemotaxis to monocyte chemoattractant protein 1. J. Immunol. 2001, 167, 3414–3421. [Google Scholar] [CrossRef] [PubMed]
- Smani, T.; Zakharov, S.I.; Leno, E.; Csutora, P.; Trepakova, E.S.; Bolotina, V.M. Ca2+-independent phospholipase A2 is a novel determinant of store-operated Ca2+ entry. J. Biol. Chem. 2003, 278, 11909–11915. [Google Scholar] [CrossRef] [PubMed]
- Chilton, F.H.; Fonteh, A.N.; Surette, M.E.; Triggiani, M.; Winkler, J.D. Control of arachidonate levels within inflammatory cells. Biochim. Biophys. Acta 1996, 1299, 1–15. [Google Scholar] [CrossRef]
- Yamashita, A.; Hayashi, Y.; Nemoto-Sasaki, Y.; Ito, M.; Oka, S.; Tanikawa, T.; Waku, K.; Sugiura, T. Acyltransferases and transacylases that determine the fatty acid composition of glycerolipids and the metabolism of bioactive lipid mediators in mammalian cells and model organisms. Prog. Lipid Res. 2014, 53, 18–81. [Google Scholar] [CrossRef]
- Fahy, E.; Subramaniam, S.; Brown, H.A.; Glass, C.K.; Merrill, A.H., Jr.; Murphy, R.C.; Raetz, C.R.; Russell, D.W.; Seyama, Y.; Shaw, W.; et al. A comprehensive classification system for lipids. J. Lipid Res. 2005, 46, 839–861. [Google Scholar] [CrossRef]
- Balsinde, J.; Fernández, B.; Solís-Herruzo, J.A. Increased incorporation of arachidonic acid into phospholipids in zymosan-stimulated mouse peritoneal macrophages. Eur. J. Biochem. 1994, 221, 1013–1018. [Google Scholar] [CrossRef]
- Neuschäfer-Rube, F.; Pathe-Neuschäfer-Rube, A.; Hippenstiel, S.; Püschel, G.P. PGE2 enhanced TNFα-mediated IL-8 induction in monocytic cell lines and PBMC. Cytokine 2019, 113, 105–116. [Google Scholar] [CrossRef]
- Samiea, A.; Yoon, J.S.J.; Cheung, S.T.; Chamberlain, T.C.; Mui, A.L.F. Interleukin-10 contributes to PGE2 signalling through upregulation of EP4 via SHIP1 and STAT3. PLoS ONE 2020, 15, 1–18. [Google Scholar] [CrossRef]
- Leslie, C.C. Cytosolic phospholipase A₂: Physiological function and role in disease. J. Lipid Res. 2015, 56, 1386–1402. [Google Scholar] [CrossRef] [PubMed]
- Astudillo, A.M.; Balboa, M.A.; Balsinde, J. Selectivity of phospholipid hydrolysis by phospholipase A2 enzymes in activated cells leading to polyunsaturated fatty acid mobilization. Biochim. Biophys. Acta 2019, 1864, 772–783. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Chacón, G.; Astudillo, A.M.; Balgoma, D.; Balboa, M.A.; Balsinde, J. 2009. Control of free arachidonic acid levels by phospholipases A2 and lysophospholipid acyltransferases. Biochim. Biophys. Acta 2009, 1791, 1103–1113. [Google Scholar] [CrossRef]
- Humes, J.L.; Bonney, R.J.; Pelus, L.; Dahlgren, M.E.; Sadowski, S.J.; Kuehl, F.A.; Davies, P. Macrophages synthesise and release prostaglandins in response to inflammatory stimuli. Nature 1977, 269, 149–151. [Google Scholar] [CrossRef]
- Scott, W.A.; Zrike, J.M.; Hamill, A.L.; Kempe, J.; Cohn, Z.A. Regulation of arachidonic acid metabolites in macrophages. J. Exp. Med. 1980, 152, 324–335. [Google Scholar] [CrossRef]
- Emilsson, A.; Sundler, R. Evidence for a catalytic role of phospholipase A in phorbol diester- and zymosan-induced mobilization of arachidonic acid in mouse peritoneal macrophages. Biochim. Biophys. Acta 1986, 876, 533–542. [Google Scholar] [CrossRef]
- Balsinde, J.; Fernández, B.; Solís-Herruzo, J.A.; Diez, E. Pathways for arachidonic acid mobilization in zymosan-stimulated mouse peritoneal macrophages. Biochim. Biophys. Acta 1992, 1136, 75–82. [Google Scholar] [CrossRef]
- Qiu, Z.H.; Leslie, C.C. Protein kinase C-dependent and -independent pathways of mitogen-activated protein kinase activation in macrophages by stimuli that activate phospholipase A2. J. Biol Chem. 1994, 269, 19480–19487. [Google Scholar] [CrossRef]
- Rouzer, C.A.; Ivanova, P.T.; Byrne, M.O.; Milne, S.B.; Marnett, L.J.; Brown, H.A. Lipid profiling reveals arachidonate deficiency in RAW264.7 cells: Structural and functional implications. Biochemistry 2006, 45, 14795–14808. [Google Scholar] [CrossRef] [PubMed]
- Astudillo, A.M.; Meana, C.; Guijas, C.; Pereira, L.; Lebrero, R.; Balboa, M.A.; Balsinde, J. Occurrence and biological activity of palmitoleic acid isomers in phagocytic cells. J. Lipid Res. 2018, 59, 237–249. [Google Scholar] [CrossRef]
- Sugiura, T.; Nakajima, M.; Sekiguchi, N.; Nakagawa, Y.; Waku, K. Different fatty chain compositions of alkenylacyl, alkylacyl and diacyl phospholipids in rabbit alveolar macrophages: High amounts of arachidonic acid in ether phospholipids. Lipids 1983, 18, 125–129. [Google Scholar] [CrossRef]
- MacDonald, J.I.S.; Sprecher, H. Phospholipid fatty acid remodeling in mammalian cells. Biochim. Biophys. Acta 1991, 1084, 105–121. [Google Scholar] [CrossRef]
- Mouchlis, V.D.; Dennis, E.A. Phospholipase A2 catalysis and lipid mediator lipidomics. Biochim. Biophys. Acta 2019, 1864, 766–771. [Google Scholar] [CrossRef] [PubMed]
- Fonteh, A.N.; Chilton, F.H. Rapid remodeling of arachidonate from phosphatidylcholine to phosphatidylethanolamine pools during mast cell activation. J. Immunol. 1992, 148, 1784–1791. [Google Scholar]
- Fonteh, A.N.; Chilton, F.H. Mobilization of different arachidonate pools and their roles in the generation of leukotrienes and free arachidonic acid during immunologic activation of mast cells. J. Immunol. 1993, 150, 563–570. [Google Scholar] [PubMed]
- Dennis, E.A.; Cao, J.; Hsu, Y.H.; Magrioti, V.; Kokotos, G. Phospholipase A2 enzymes: Physical structure, biological function, disease implication, chemical inhibition, and therapeutic intervention. Chem. Rev. 2011, 111, 6130–6185. [Google Scholar] [CrossRef]
- Chilton, F.H. Potential phospholipid source(s) of arachidonate used for the synthesis of leukotrienes by the human neutrophil. Biochem. J. 1989, 258, 327–333. [Google Scholar] [CrossRef] [PubMed]
- Brown, H.A.; Murphy, R.C. Working towards an exegesis for lipids in biology. Nat. Chem. Biol. 2009, 5, 602–606. [Google Scholar] [CrossRef]
- Lebrero, P.; Astudillo, A.M.; Rubio, J.M.; Fernández-Caballero, J.; Kokotos, G.; Balboa, M.A.; Balsinde, J. Cellular plasmalogen content does not influence arachidonic acid levels or distribution in macrophages: A role for cytosolic phospholipase A2γ in phospholipid remodeling. Cells 2019, 8, 799. [Google Scholar] [CrossRef]
- Guijas, C.; Bermúdez, M.A.; Meana, C.; Astudillo, A.M.; Pereira, L.; Fernández-Caballero, L.; Balboa, M.A.; Balsinde, J. Neutral lipids are not a source of arachidonic acid for lipid mediator signaling in human foamy monocytes. Cells 2019, 8, 941. [Google Scholar] [CrossRef]
- Dean, J.M.; Lodhi, I.J. Structural and functional roles of ether lipids. Protein Cell 2018, 9, 196–206. [Google Scholar] [CrossRef]
- Koivuniemi, A. The biophysical properties of plasmalogens originating from their unique molecular architecture. FEBS Lett. 2017, 591, 2700–2713. [Google Scholar] [CrossRef]
- Honsho, M.; Fujiki, Y. Plasmalogen homeostasis. Regulation of plasmalogen biosynthesis and its physiological consequence in mammals. FEBS Lett. 2017, 591, 2720–2729. [Google Scholar] [CrossRef]
- Zou, Y.; Henry, W.S.; Ricq, E.L.; Graham, E.T.; Phadnis, V.V.; Maretich, P.; Paradkar, S.; Boehnke, N.; Deik, A.A.; Reinhardt, F.; et al. Plasticity of ether lipids promotes ferroptosis susceptibility and evasion. Nature 2020, 585, 603–608. [Google Scholar] [CrossRef] [PubMed]
- Kapralov, A.A.; Yang, Q.; Dar, H.H.; Tyurina, Y.Y.; Anthonymuthu, T.S.; Kim, R.; Croix, C.M.S.; Mikulska-Ruminska, K.; Liu, B.; Shrivastava, I.H.; et al. Redox lipid reprogramming commands susceptibility of macrophages and microglia to ferroptotic death. Nat. Chem. Biol. 2020, 16, 278–290. [Google Scholar] [CrossRef] [PubMed]
- Dorninger, F.; Forss-Petter, S.; Wimmer, I.; Berger, J. Plasmalogens, platelet-activating factor and beyond. Ether lipids in signaling and neurodegeneration. Neurobiol. Dis. 2020, 145, 105061. [Google Scholar] [CrossRef] [PubMed]
- Yamane, H.; Sugimoto, Y.; Tanaka, S.; Ichikawa, A. Prostaglandin E2 receptors EP2 and EP4 differentially modulate TNF-α and IL-6 production induced by lipopolysaccharide in mouse peritoneal neutrophils. Biochem. Biophys. Res. Commun. 2000, 278, 224–228. [Google Scholar] [CrossRef]
- Treffkorn, L.; Scheibe, R.; Maruyama, T.; Dieter, P. PGE2 exerts its effect on the LPS-induced release of TNF-α, ET-1, IL-lα, IL-6 and IL-10 via the EP2 and EP4 receptor in rat liver macrophages. Prostaglandins Other Lipid Mediat. 2004, 74, 113–123. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).