
Spermatogonia Loss Correlates with LAMA 1 Expression in Human Prepubertal Testes Stored for Fertility Preservation

 , , , , , ,
, , , , , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
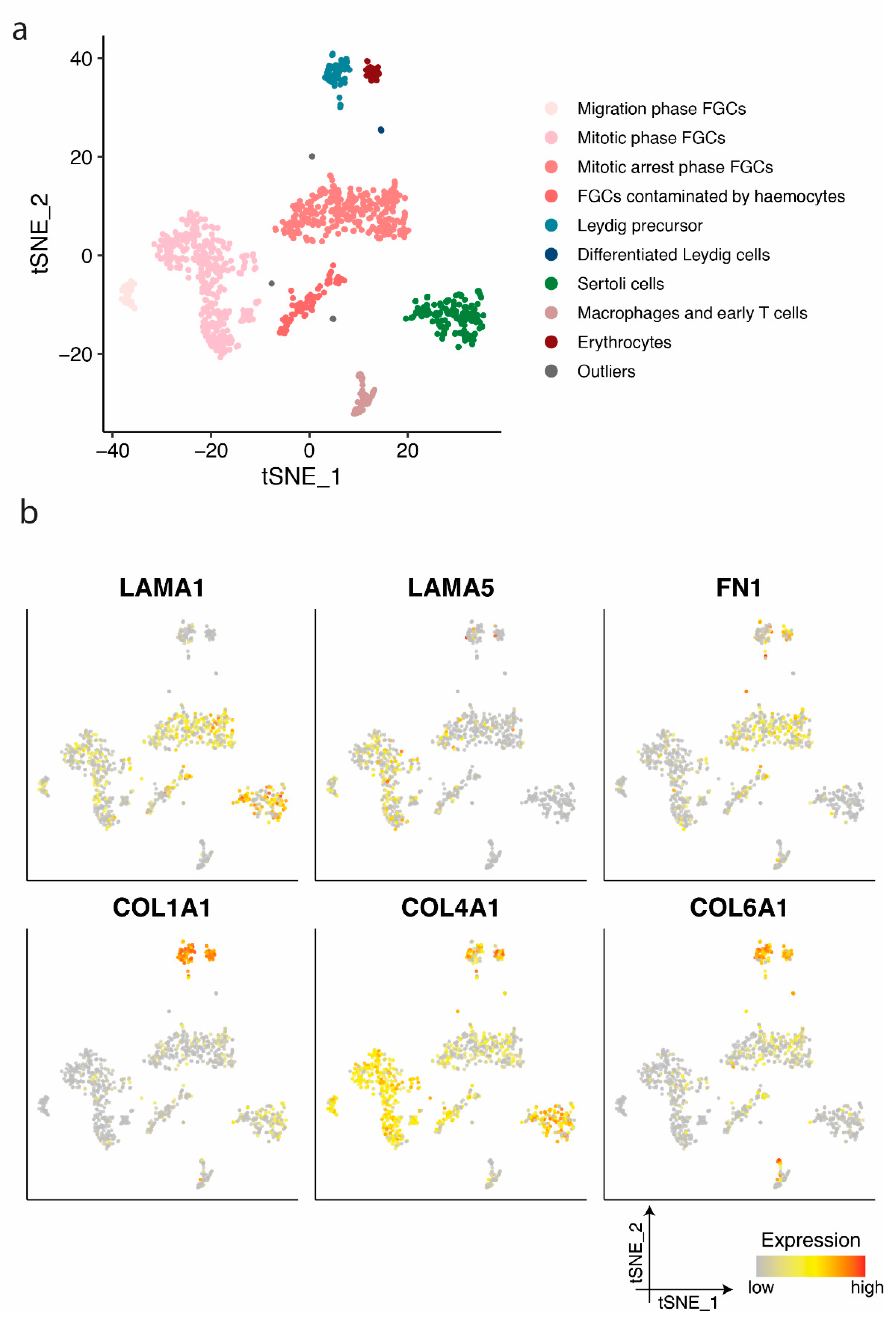
2.1. LAMA 1 and Type IV Collagen are the Main Components of the Pre- and Postnatal Seminiferous BM
2.2. Basement Membrane Disruption upon Testicular Culture Correlates with Germ Cell Loss
3. Discussion
4. Materials and Methods
4.1. Ethical Permission
4.2. Sample Collection and Preparation
4.3. Testicular Explant Tissue Cultures
4.4. Immunofluorescence Staining
4.5. Imaging
4.6. Cell Quantification and Statistical Analysis
4.7. Single-Cell RNA-Sequencing Data Processing
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Giudice, M.G.; de Michele, F.; Poels, J.; Vermeulen, M.; Wyns, C. Update on fertility restoration fromprepubertal spermatogonial stem cells: How far are we fromclinical practice? Stem Cell Res. 2017, 21, 171–177. [Google Scholar] [CrossRef]
- de Michele, F.; Poels, J.; Giudice, M.G.; De Smedt, F.; Ambroise, J.; Vermeulen, M.; Gruson, D.; Wyns, C. In vitro formation of the blood–testis barrier during long-term organotypic culture of human prepubertal tissue: Comparison with a large cohort of pre/peripubertal boys. Mol. Hum. Reprod. 2018, 24, 271–282. [Google Scholar] [CrossRef] [PubMed]
- de Michele, F.; Poels, J.; Weerens, L.; Petit, C.; Evrard, Z.; Ambroise, J.; Gruson, D.; Wyns, C. Preserved seminiferous tubule integrity with spermatogonial survival and induction of Sertoli and Leydig cell maturation after long-term organotypic culture of prepubertal human testicular tissue. Hum. Reprod. 2017, 32, 32–45. [Google Scholar] [CrossRef] [PubMed]
- Medrano, J.V.; Vilanova-Pérez, T.; Fornés-Ferrer, V.; Navarro-Gomezlechon, A.; Martínez-Triguero, M.L.; García, S.; Gómez-Chacón, J.; Povo, I.; Pellicer, A.; Andrés, M.M.; et al. Influence of temperature, serum, and gonadotropin supplementation in short- and long-term organotypic culture of human immature testicular tissue. Fertil. Steril. 2018, 110, 1045–1057.e3. [Google Scholar] [CrossRef]
- Sato, T.; Katagiri, K.; Gohbara, A.; Inoue, K.; Ogonuki, N.; Ogura, A.; Kubota, Y.; Ogawa, T. In vitro production of functional sperm in cultured neonatal mouse testes. Nature 2011, 471, 504. [Google Scholar] [CrossRef]
- Reda, A.; Hou, M.; Winton, T.R.; Chapin, R.E.; Söder, O.; Stukenborg, J.-B. In vitro differentiation of rat spermatogonia into round spermatids in tissue culture. Mol. Hum. Reprod. 2016, 22, 601–612. [Google Scholar] [CrossRef]
- de Rooij, D.G. The nature and dynamics of spermatogonial stem cells. Development 2017, 144, 3022–3030. [Google Scholar] [CrossRef] [PubMed]
- Oatley, J.M.; Brinster, R.L. The germline stem cell niche unit in mammalian testes. Physiol. Rev. 2012, 92, 577–595. [Google Scholar] [CrossRef] [PubMed]
- Siu, M.K.; Cheng, C.Y. Extracellular matrix: Recent advances on its role in junction dynamics in the seminiferous epithelium during spermatogenesis. Biol. Reprod. 2004, 71, 375–391. [Google Scholar] [CrossRef]
- Flenkenthaler, F.; Windschuttl, S.; Frohlich, T.; Schwarzer, J.U.; Mayerhofer, A.; Arnold, G.J. Secretome analysis of testicular peritubular cells: A window into the human testicular microenvironment and the spermatogonial stem cell niche in man. J. Proteome Res. 2014, 13, 1259–1269. [Google Scholar] [CrossRef]
- de Rooij, D.G. The spermatogonial stem cell niche. Microsc. Res. Tech. 2009, 72, 580–585. [Google Scholar] [CrossRef] [PubMed]
- Baert, Y.; Stukenborg, J.-B.; Landreh, M.; De Kock, J.; Jörnvall, H.; Söder, O.; Goossens, E. Derivation and characterization of a cytocompatible scaffold from human testis. Hum. Reprod. 2015, 30, 256–267. [Google Scholar] [CrossRef] [PubMed]
- Miner, J.H.; Cunningham, J.; Sanes, J.R. Roles for Laminin in Embryogenesis: Exencephaly, Syndactyly, and Placentopathy in Mice Lacking the Laminin α 5 Chain. J. Cell Biol. 1998, 143, 1713–1723. [Google Scholar] [CrossRef] [PubMed]
- Miner, J.H.; Yurchenco, P.D. Laminin functions in tissue morphogenesis. Annu. Rev. Cell Dev. Biol. 2004, 20, 255–284. [Google Scholar] [CrossRef] [PubMed]
- Ekblom, P.; Lonai, P.; Talts, J.F. Expression and biological role of laminin-1. Matrix Biol. 2003, 22, 35–47. [Google Scholar] [CrossRef]
- Klaffky, E.; Williams, R.; Yao, C.C.; Ziober, B.; Kramer, R.; Sutherland, A. Trophoblast-specific expression and function of the integrin alpha 7 subunit in the peri-implantation mouse embryo. Dev. Biol. 2001, 239, 161–175. [Google Scholar] [CrossRef]
- Agelopoulou, R.; Magre, S. Expression of fibronectin and laminin in fetal male gonads in vivo and in vitro with and without testicular morphogenesis. Cell Differ. 1987, 21, 31–36. [Google Scholar] [CrossRef]
- Häger, M.; Gawlik, K.; Nyström, A.; Sasaki, T.; Durbeej, M. Laminin α1 Chain Corrects Male Infertility Caused by Absence of Laminin α2 Chain. Am. J. Pathol. 2005, 167, 823–833. [Google Scholar] [CrossRef]
- Gao, Y.; Chen, H.; Lui, W.Y.; Lee, W.M.; Cheng, C.Y. Basement Membrane Laminin alpha2 Regulation of BTB Dynamics via Its Effects on F-Actin and Microtubule Cytoskeletons Is Mediated Through mTORC1 Signaling. Endocrinology 2017, 158, 963–978. [Google Scholar] [CrossRef][Green Version]
- Gao, Y.; Mruk, D.; Chen, H.; Lui, W.Y.; Lee, W.M.; Cheng, C.Y. Regulation of the blood-testis barrier by a local axis in the testis: Role of laminin alpha2 in the basement membrane. FASEB J. 2017, 31, 584–597. [Google Scholar] [CrossRef]
- Pöllanen, P.P.; Kallajoki, M.; Risteliz, L.; Risteli, J.; Suominen, J.J.O. Laminin and type 1V collagen in the human testis. Int. J. Androl. 1985, 8, 337–347. [Google Scholar] [CrossRef] [PubMed]
- Virtanen, I.; Lohi, J.; Tani, T.; Korhonen, M.; Burgeson, R.E.; Lehto, V.P.; Leivo, I. Distinct Changes in the Laminin Composition of Basement Membranes in Human Seminiferous Tubules during Development and Degeneration. Am. J. Pathol. 1997, 150, 1421–1431. [Google Scholar] [PubMed]
- Ooba, T.; Ishikawa, T.; Yamaguchi, K.; Kondo, Y.; Sakamoto, Y.; Fujisawa, M. Expression and distribution of laminin chains in the testis for patients with azoospermia. J. Androl. 2008, 29, 147–152. [Google Scholar] [CrossRef] [PubMed]
- Dobashi, M.; Fujisawa, M.; Naito, I.; Yamazaki, T.; Okada, H.; Kamidono, S. Distribution of type IV collagen subtypes in human testes and their association with spermatogenesis. Fertil. Steril. 2003, 80, 755–760. [Google Scholar] [CrossRef]
- Santamaria, L.; Martinez-Onsurbe, P.; Paniagua, R.; Nistal, M. Laminin, type IV collagen, and fibronectin in normal and cryptorchid human testes. An immunohistochemical study. Int. J. Androl. 1990, 13, 135–146. [Google Scholar] [CrossRef]
- Virtanen, I.; Gullberg, D.; Rissanen, J.; Kivilaakso, E.; Kiviluoto, T.; Laitinen, L.A.; Lehto, V.P.; Ekblom, P. Laminin alpha1-chain shows a restricted distribution in epithelial basement membranes of fetal and adult human tissues. Exp. Cell Res. 2000, 257, 298–309. [Google Scholar] [CrossRef]
- Domogatskaya, A.; Rodin, S.; Tryggvason, K. Functional diversity of laminins. Annu. Rev. Cell Dev. Biol. 2012, 28, 523–553. [Google Scholar] [CrossRef]
- Yamada, M.; Sekiguchi, K. Molecular Basis of Laminin-Integrin Interactions. Curr. Top. Membr. 2015, 76, 197–229. [Google Scholar]
- Li, L.; Dong, J.; Yan, L.; Yong, J.; Liu, X.; Hu, Y.; Fan, X.; Wu, X.; Guo, H.; Wang, X.; et al. Single-Cell RNA-Seq Analysis Maps Development of Human Germline Cells and Gonadal Niche Interactions. Cell Stem Cell 2017, 20, 858–873.e4. [Google Scholar] [CrossRef]
- Guo, J.; Nie, X.; Giebler, M.; Mlcochova, H.; Wang, Y.; Grow, E.J.; Kim, R.; Tharmalingam, M.; Matilionyte, G.; Lindskog, C.; et al. The Dynamic Transcriptional Cell Atlas of Testis Development during Human Puberty. Cell Stem Cell 2020, 26, 262–276.e4. [Google Scholar] [CrossRef]
- Guo, F.; Yan, L.; Guo, H.; Li, L.; Hu, B.; Zhao, Y.; Yong, J.; Hu, Y.; Wang, X.; Wei, Y.; et al. The Transcriptome and DNA Methylome Landscapes of Human Primordial Germ Cells. Cell 2015, 161, 1437–1452. [Google Scholar] [CrossRef] [PubMed]
- Stukenborg, J.-B.; Alves-Lopes, J.P.; Kurek, M.; Albalushi, H.; Reda, A.; Keros, V.; Töhönen, V.; Bjarnason, R.; Romerius, P.; Sundin, M.; et al. Spermatogonial quantity in human prepubertal testicular tissue collected for fertility preservation prior to potentially sterilizing therapy. Hum. Reprod. 2018, 33, 1677–1683. [Google Scholar] [CrossRef]
- Sohni, A.; Tan, K.; Song, H.W.; Burow, D.; de Rooij, D.G.; Laurent, L.; Hsieh, T.C.; Rabah, R.; Hammoud, S.S.; Vicini, E.; et al. The Neonatal and Adult Human Testis Defined at the Single-Cell Level. Cell Rep. 2019, 26, 1501–1517.e4. [Google Scholar] [CrossRef] [PubMed]
- Nieschlag, E.; Behre, H. Andrology Male Reproductive Health and Dysfunction, 3rd ed.; Springer: Berlin/Heidelberg, Germany, 2010; pp. 1–629. [Google Scholar]
- Kerr, C.L.; Hill, C.M.; Blumenthal, P.D.; Gearhart, J.D. Expression of pluripotent stem cell markers in the human fetal ovary. Hum. Reprod. 2008, 23, 589–599. [Google Scholar] [CrossRef][Green Version]
- Anderson, R.A.; Fulton, N.; Cowan, G.; Coutts, S.; Saunders, P.T.K. Conserved and divergent patterns of expression of DAZL, VASA and OCT4 in the germ cells of the human fetal ovary and testis. BMC Dev. Biol. 2007, 7, 136. [Google Scholar] [CrossRef] [PubMed]
- Murdock, M.H.; David, S.; Swinehart, I.T.; Reing, J.E.; Tran, K.; Gassei, K.; Orwig, K.E.; Badylak, S.F. Human Testis Extracellular Matrix Enhances Human Spermatogonial Stem Cell Survival In Vitro. Tissue Eng. Part A 2019, 25, 663–676. [Google Scholar] [CrossRef] [PubMed]
- Marjorie, A. Colour Atlas of Life Before Birth: Normal Fetal Development; Wolfe Medical Publications: London, UK, 1999; p. 1216. [Google Scholar]
- Butler, A.; Hoffman, P.; Smibert, P.; Papalexi, E.; Satija, R. Integrating single-cell transcriptomic data across different conditions, technologies, and species. Nat. Biotechnol. 2018, 36, 411. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kurek, M.; Åkesson, E.; Yoshihara, M.; Oliver, E.; Cui, Y.; Becker, M.; Alves-Lopes, J.P.; Bjarnason, R.; Romerius, P.; Sundin, M.; et al. Spermatogonia Loss Correlates with LAMA 1 Expression in Human Prepubertal Testes Stored for Fertility Preservation. Cells 2021, 10, 241. https://doi.org/10.3390/cells10020241
Kurek M, Åkesson E, Yoshihara M, Oliver E, Cui Y, Becker M, Alves-Lopes JP, Bjarnason R, Romerius P, Sundin M, et al. Spermatogonia Loss Correlates with LAMA 1 Expression in Human Prepubertal Testes Stored for Fertility Preservation. Cells. 2021; 10(2):241. https://doi.org/10.3390/cells10020241
Chicago/Turabian StyleKurek, Magdalena, Elisabet Åkesson, Masahito Yoshihara, Elizabeth Oliver, Yanhua Cui, Martin Becker, João Pedro Alves-Lopes, Ragnar Bjarnason, Patrik Romerius, Mikael Sundin, and et al. 2021. "Spermatogonia Loss Correlates with LAMA 1 Expression in Human Prepubertal Testes Stored for Fertility Preservation" Cells 10, no. 2: 241. https://doi.org/10.3390/cells10020241
APA StyleKurek, M., Åkesson, E., Yoshihara, M., Oliver, E., Cui, Y., Becker, M., Alves-Lopes, J. P., Bjarnason, R., Romerius, P., Sundin, M., Norén Nyström, U., Langenskiöld, C., Vogt, H., Henningsohn, L., Petersen, C., Söder, O., Guo, J., Mitchell, R. T., Jahnukainen, K., & Stukenborg, J.-B. (2021). Spermatogonia Loss Correlates with LAMA 1 Expression in Human Prepubertal Testes Stored for Fertility Preservation. Cells, 10(2), 241. https://doi.org/10.3390/cells10020241