
Co-Expression of Nogo-A in Dopaminergic Neurons of the Human Substantia Nigra Pars Compacta Is Reduced in Parkinson’s Disease

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
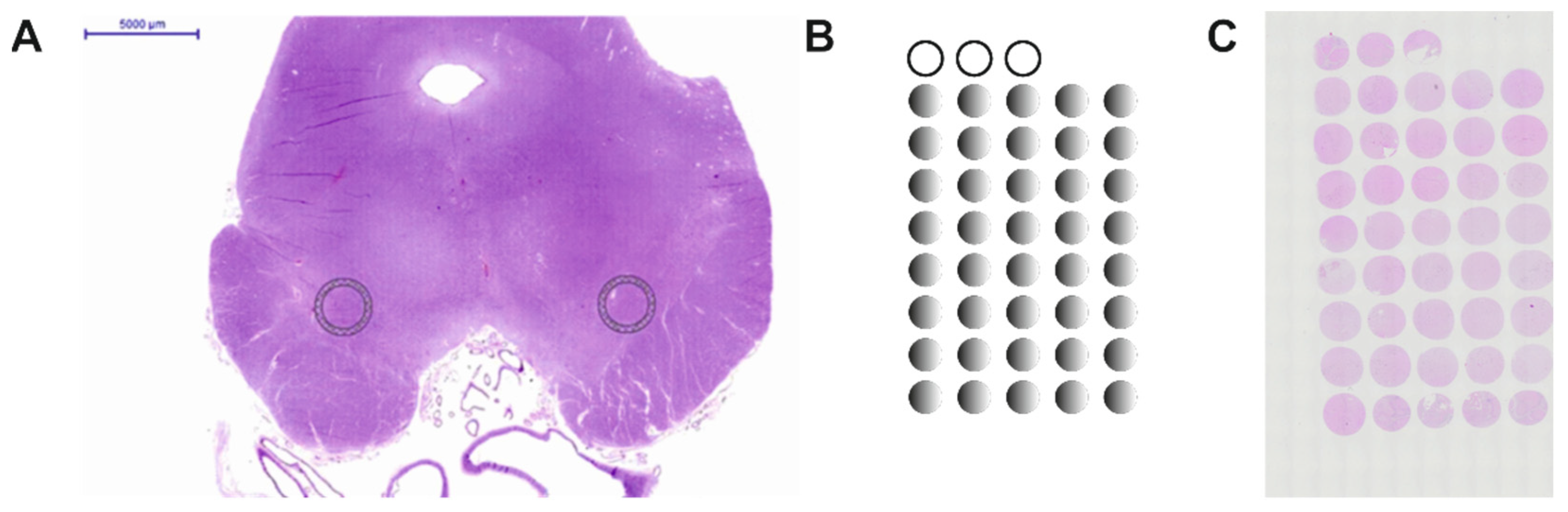
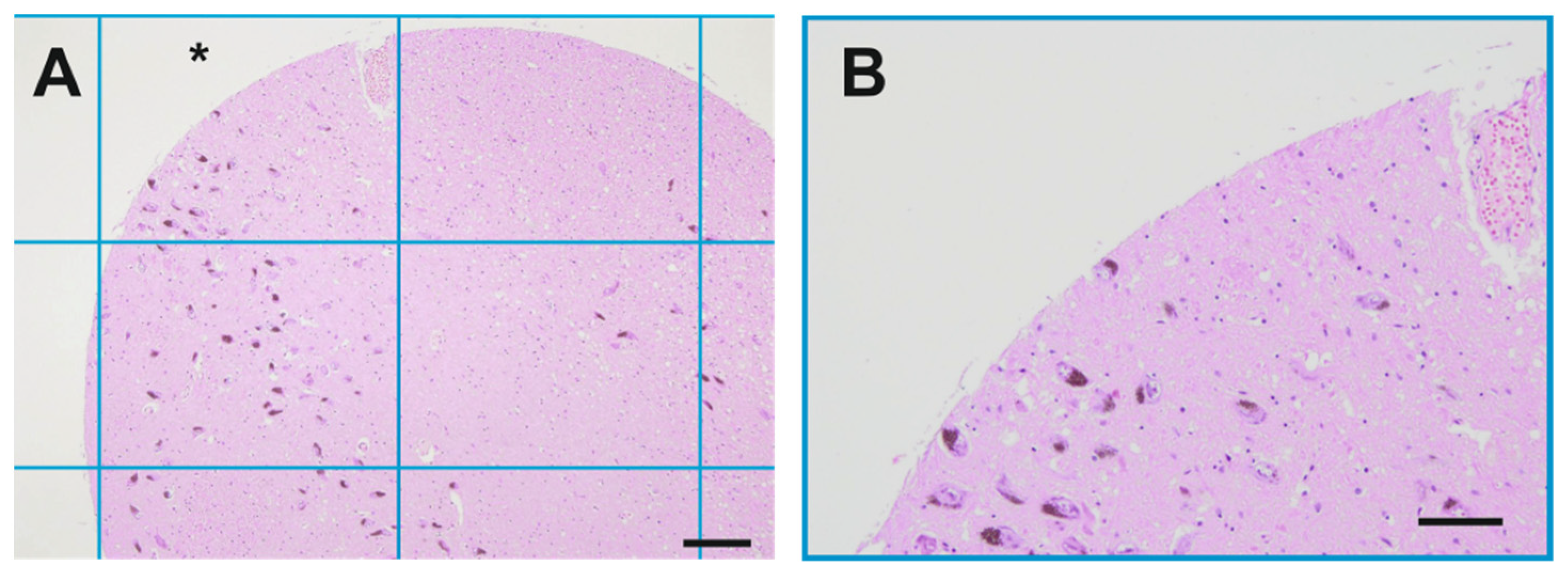
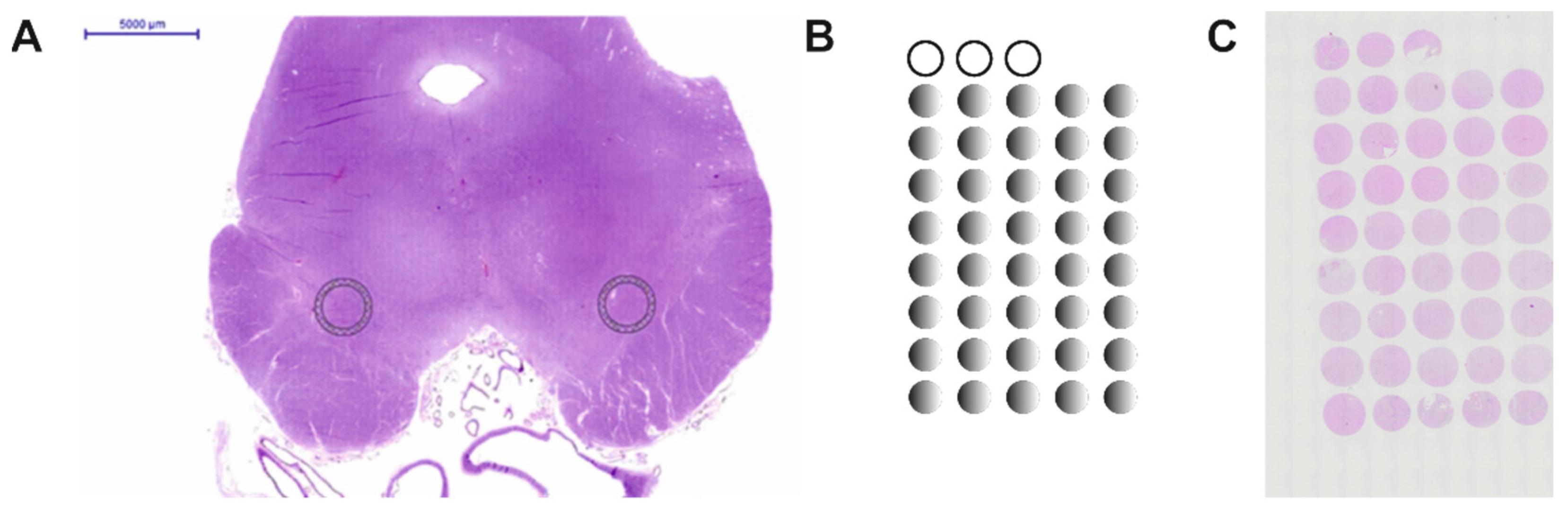
2.1. Construction of the Human Substantia Nigra Pars Compacta TMA
2.2. Immunohistochemistry
2.3. Cell Count Analysis
2.4. Statistical Analysis
3. Results
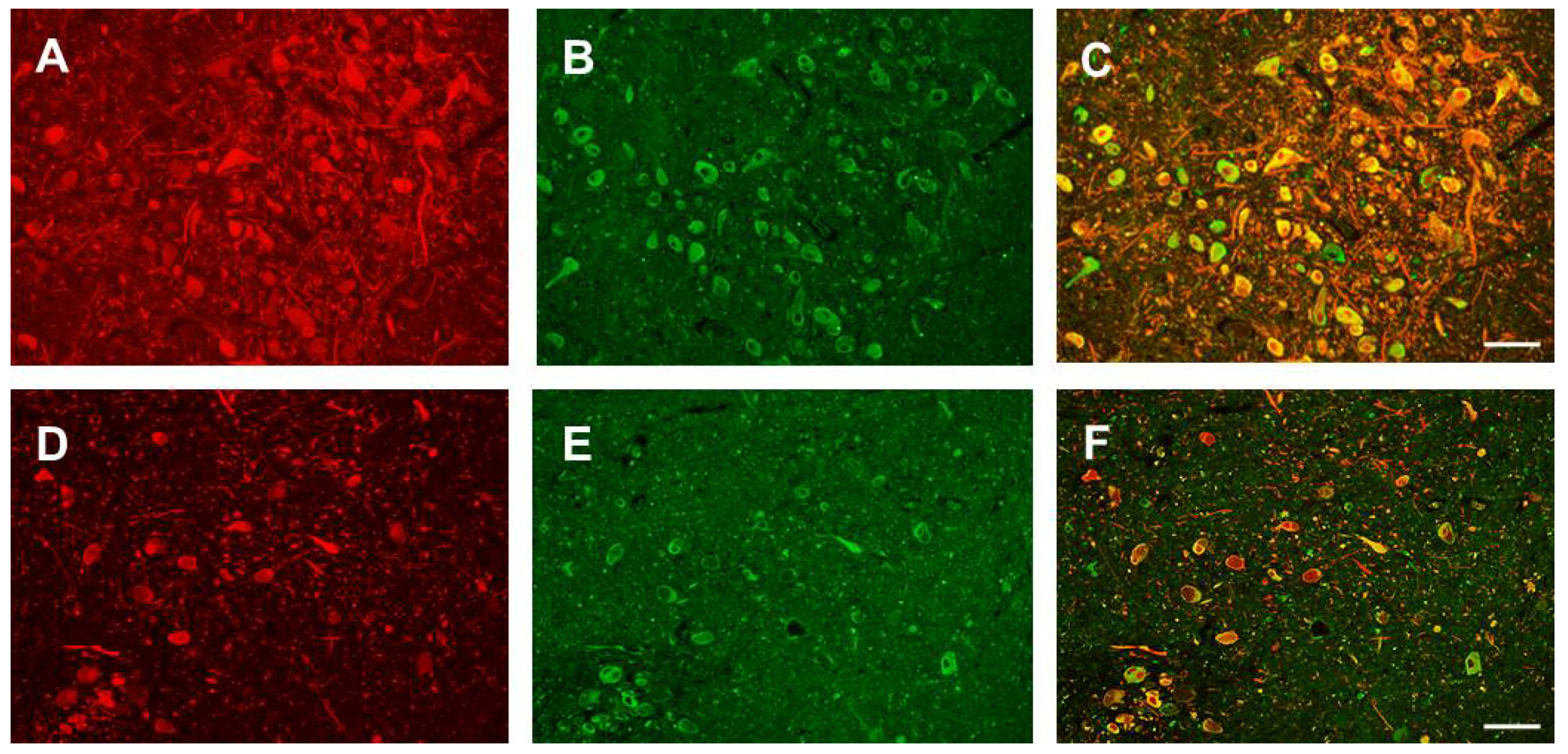
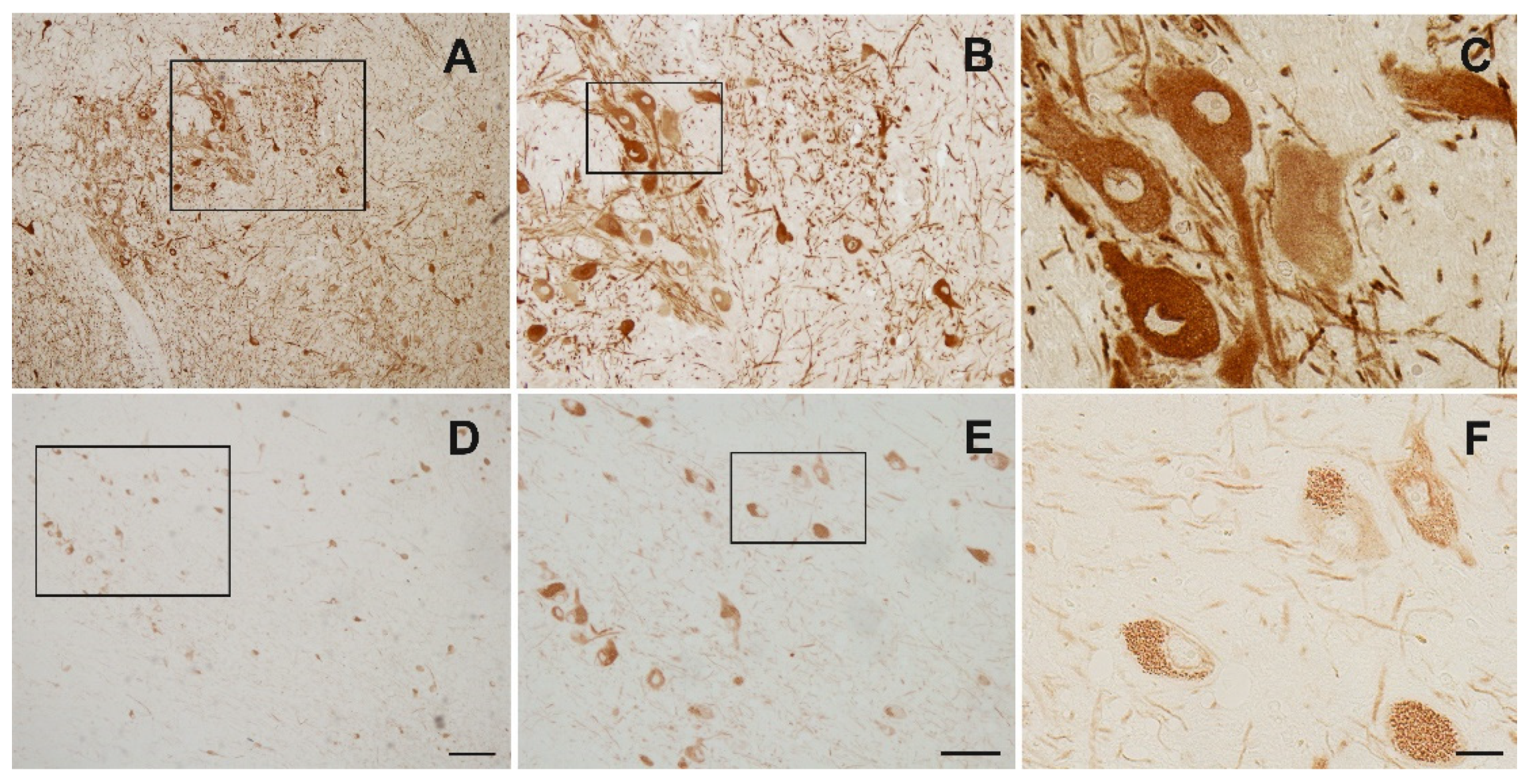
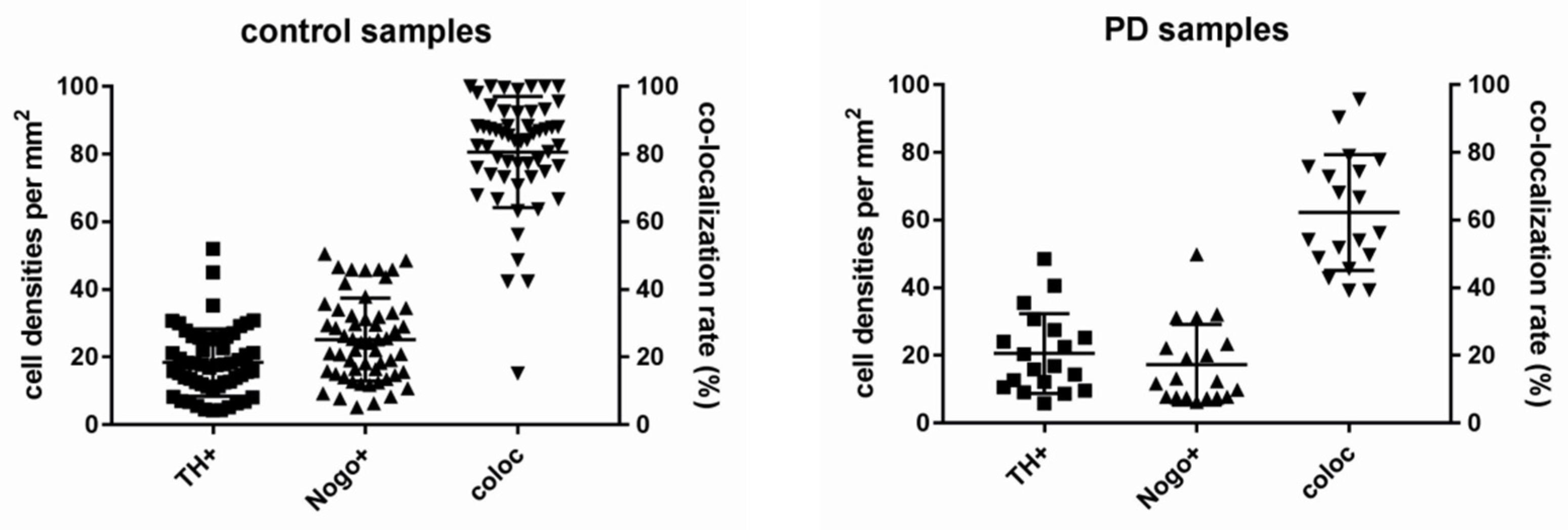
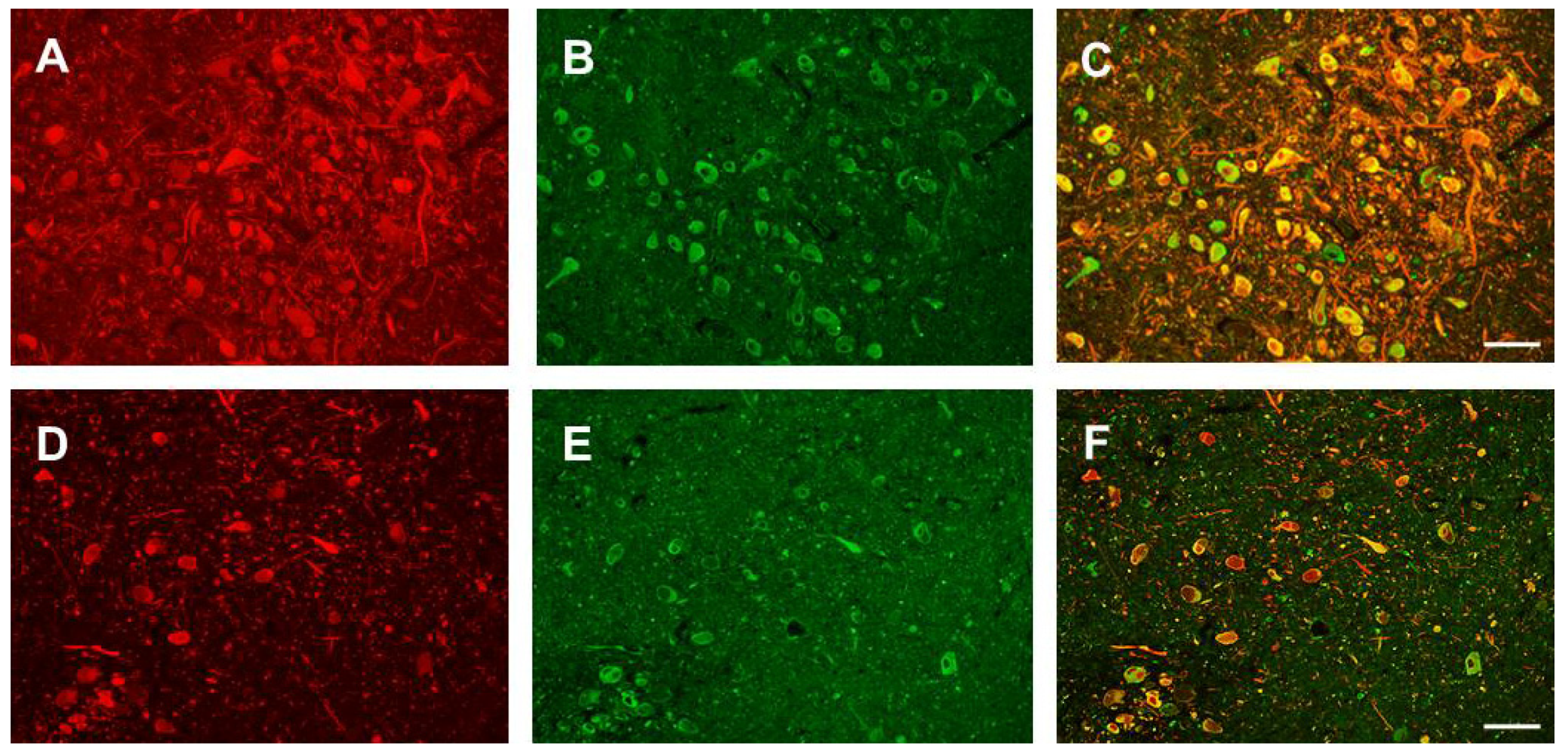
3.1. Nogo-A Is Expressed in DAneurons Human SNc
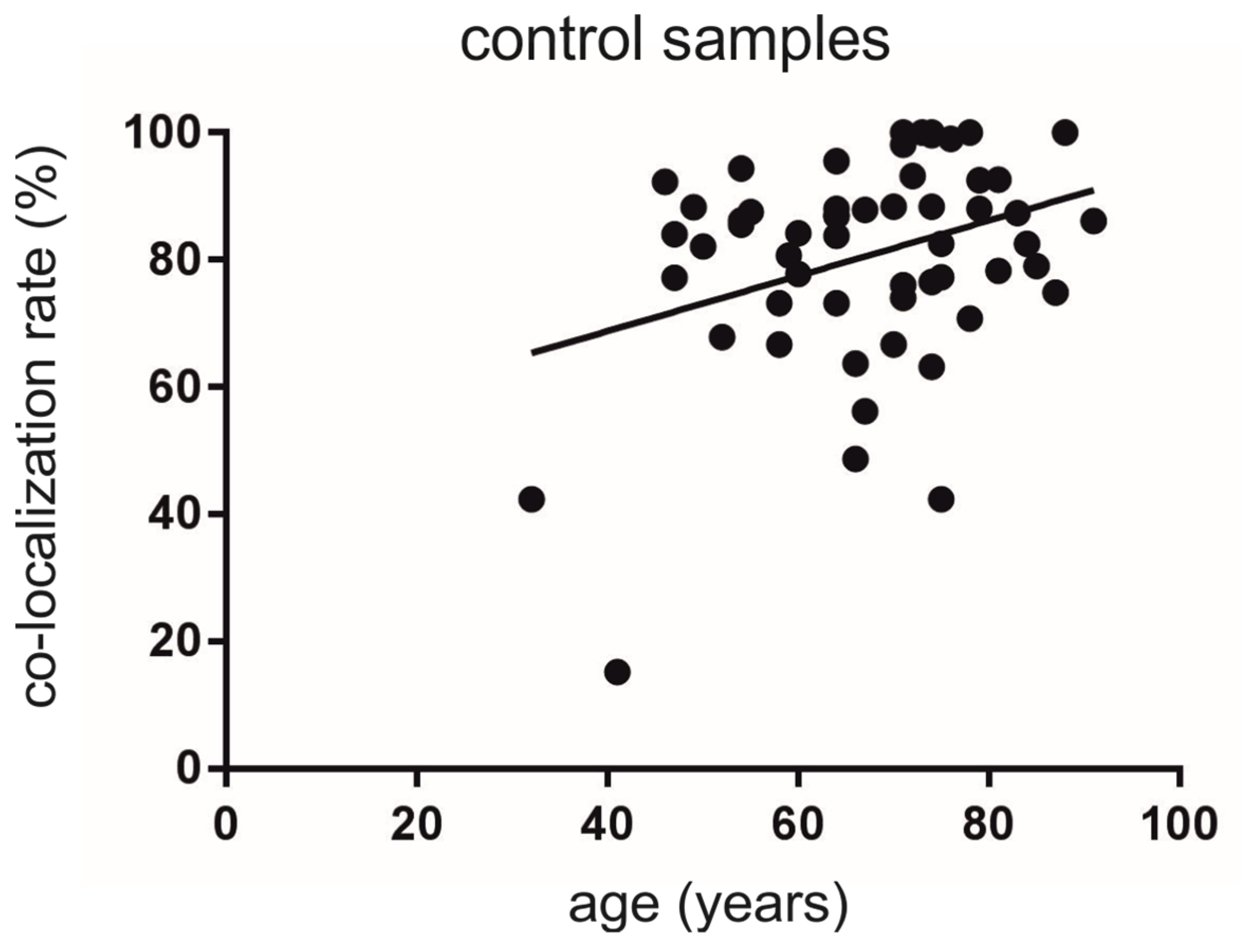
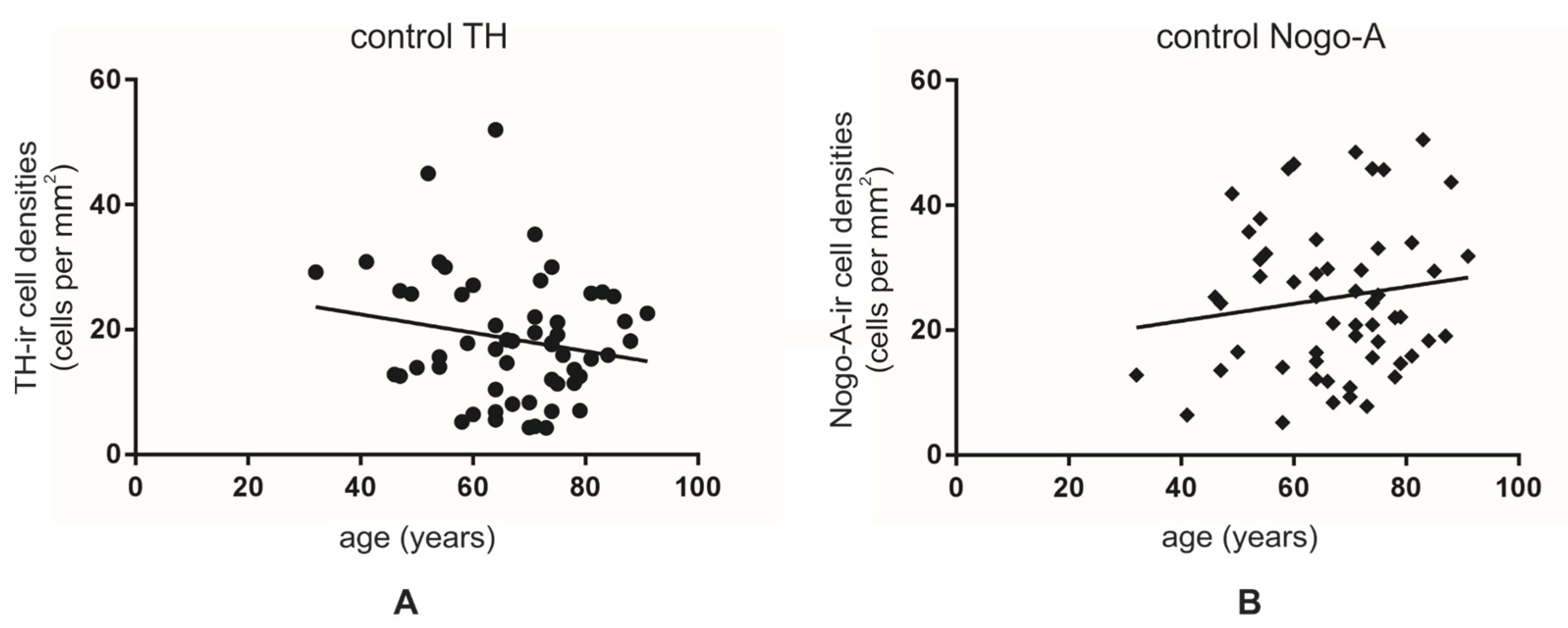
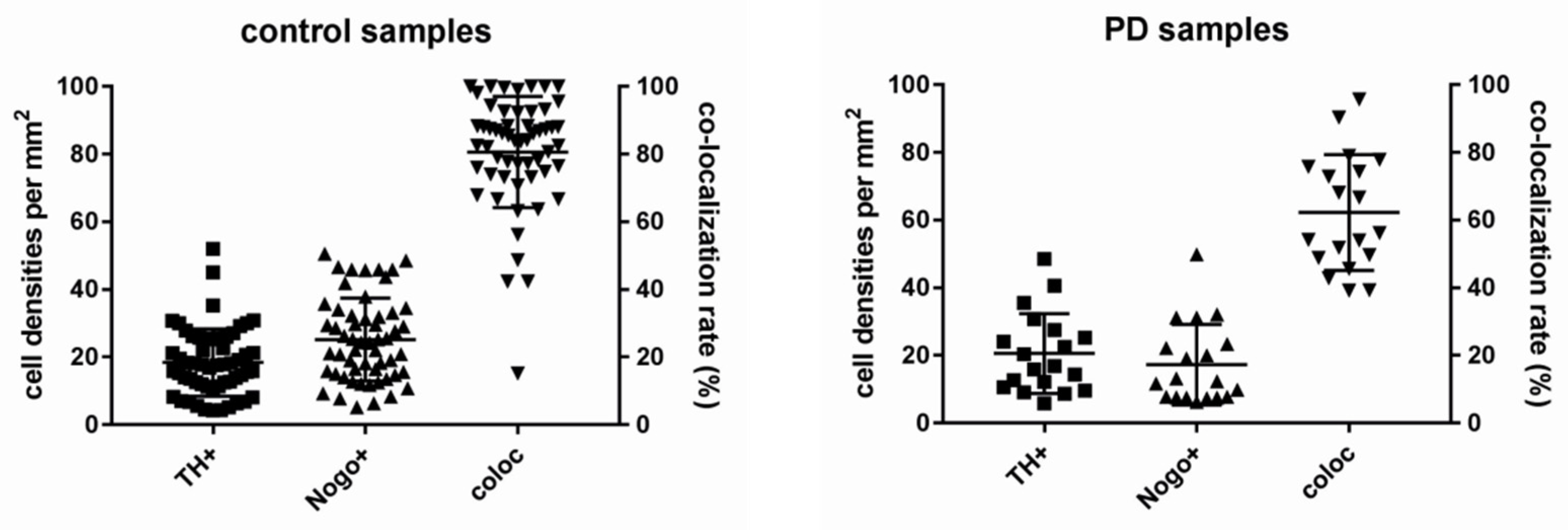
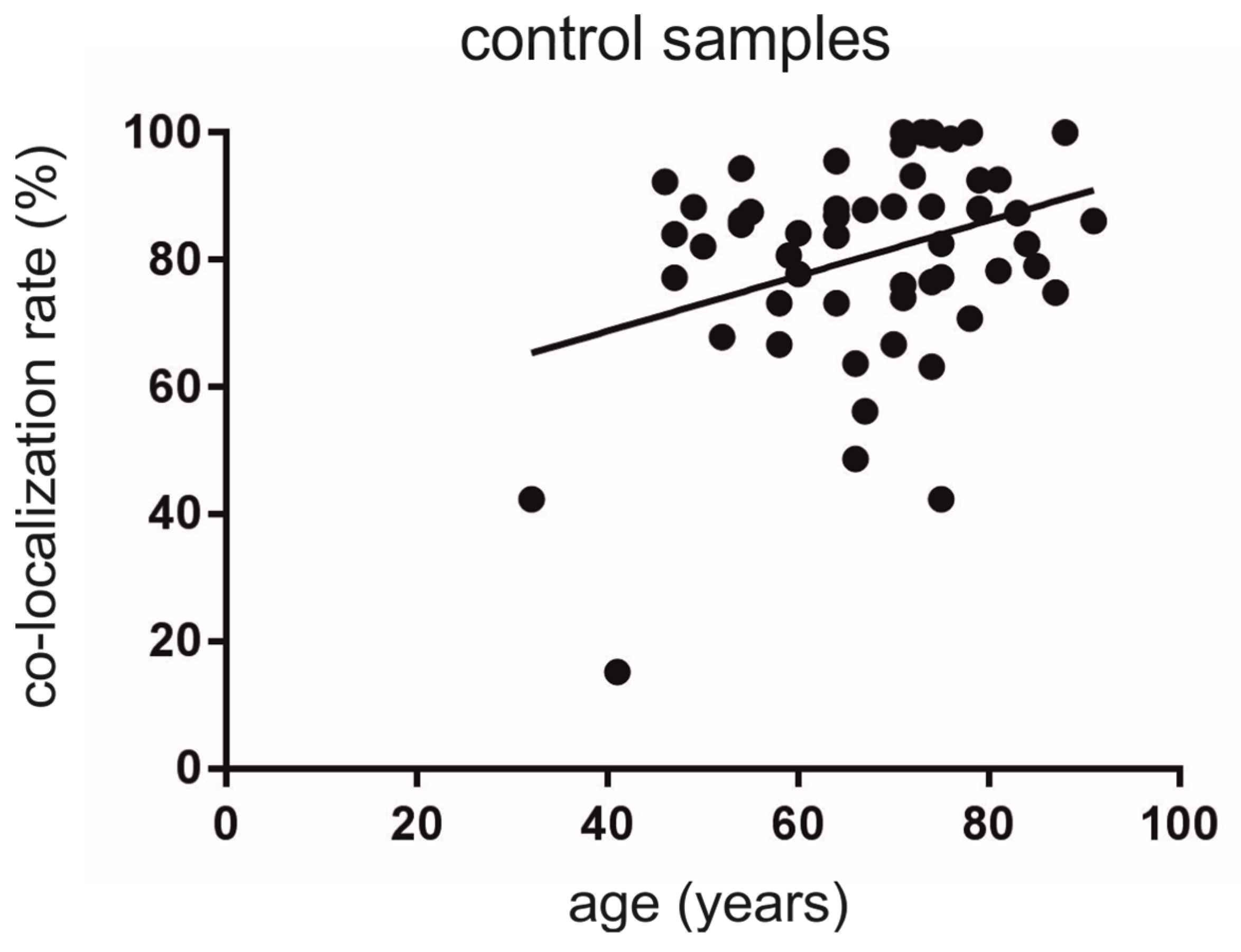
3.2. Co-Localization Rates Increase with Age in the Non-Diseased Brains
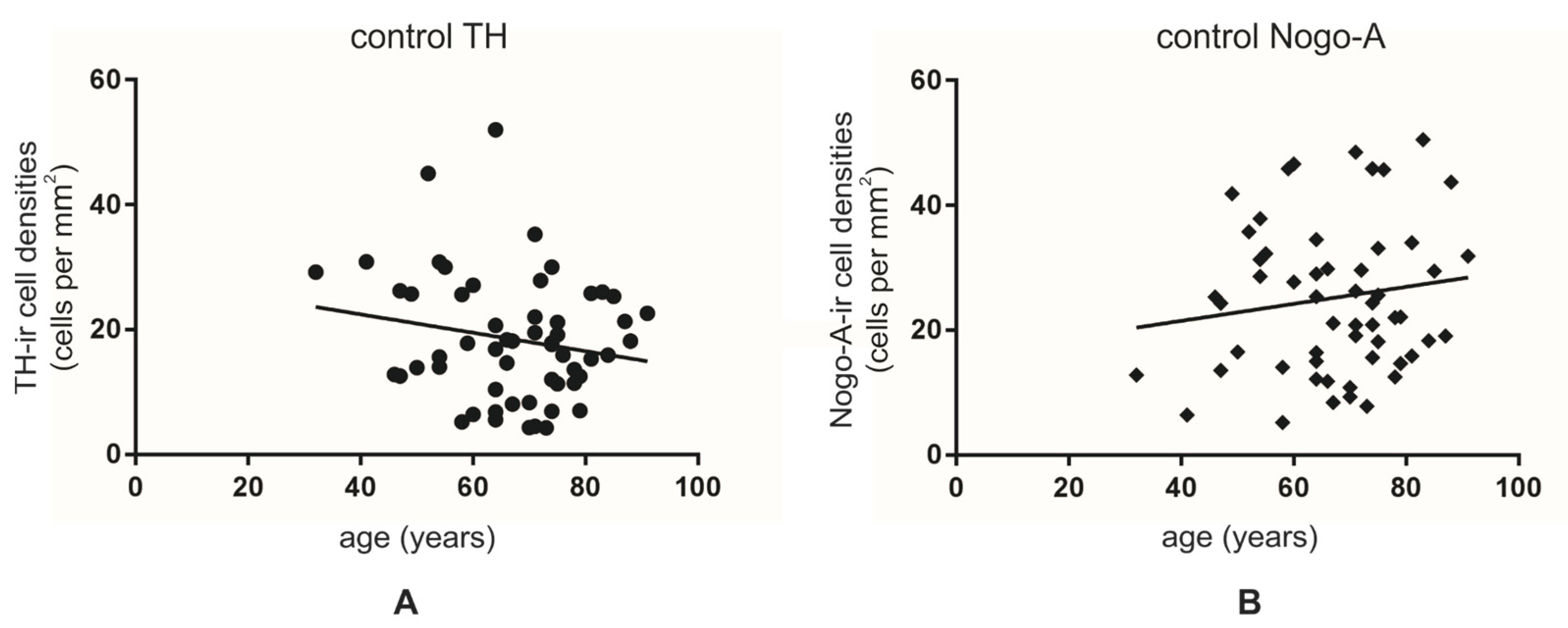
3.3. Lower Numbers of TH-ir Neurons in Normal Aging
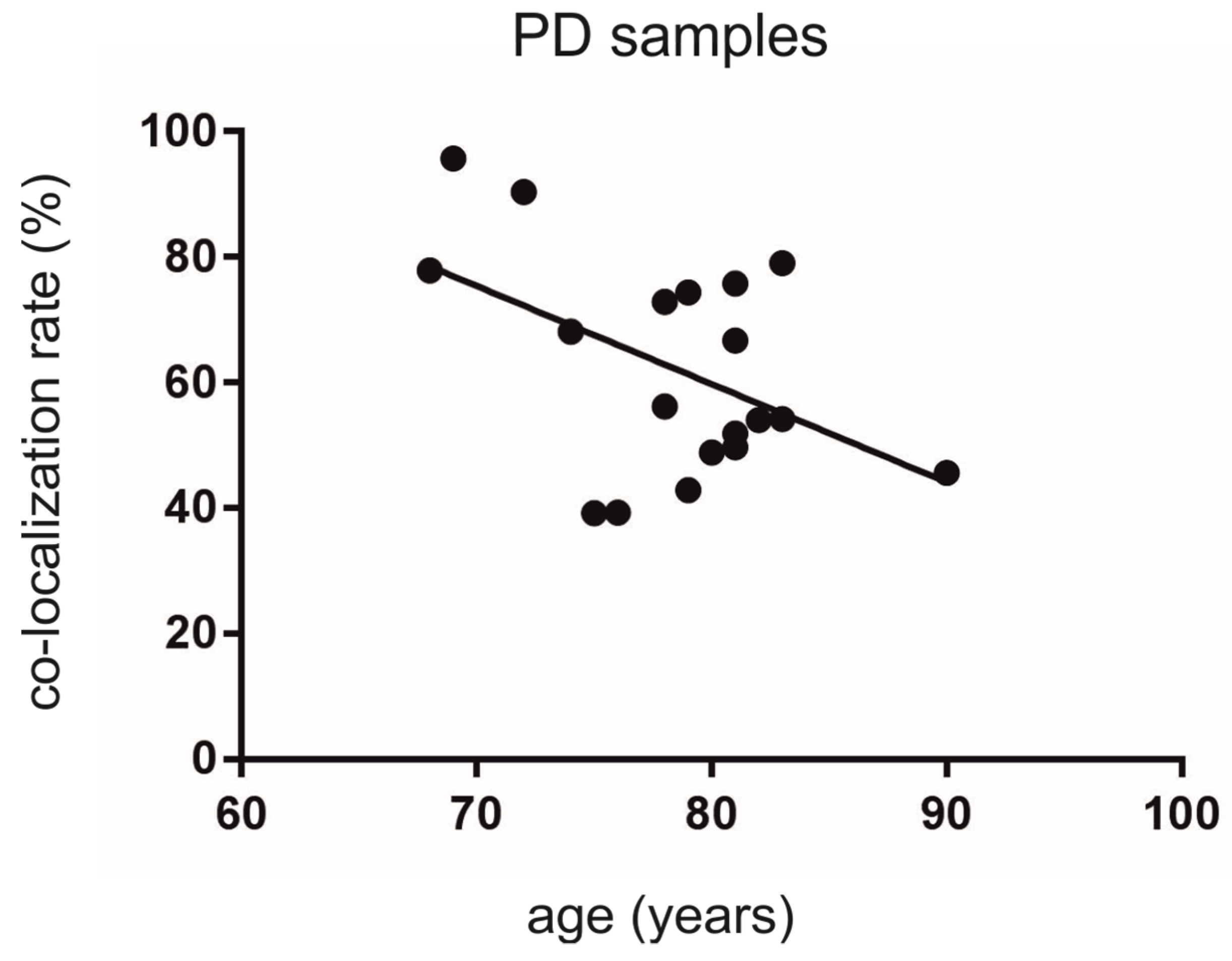
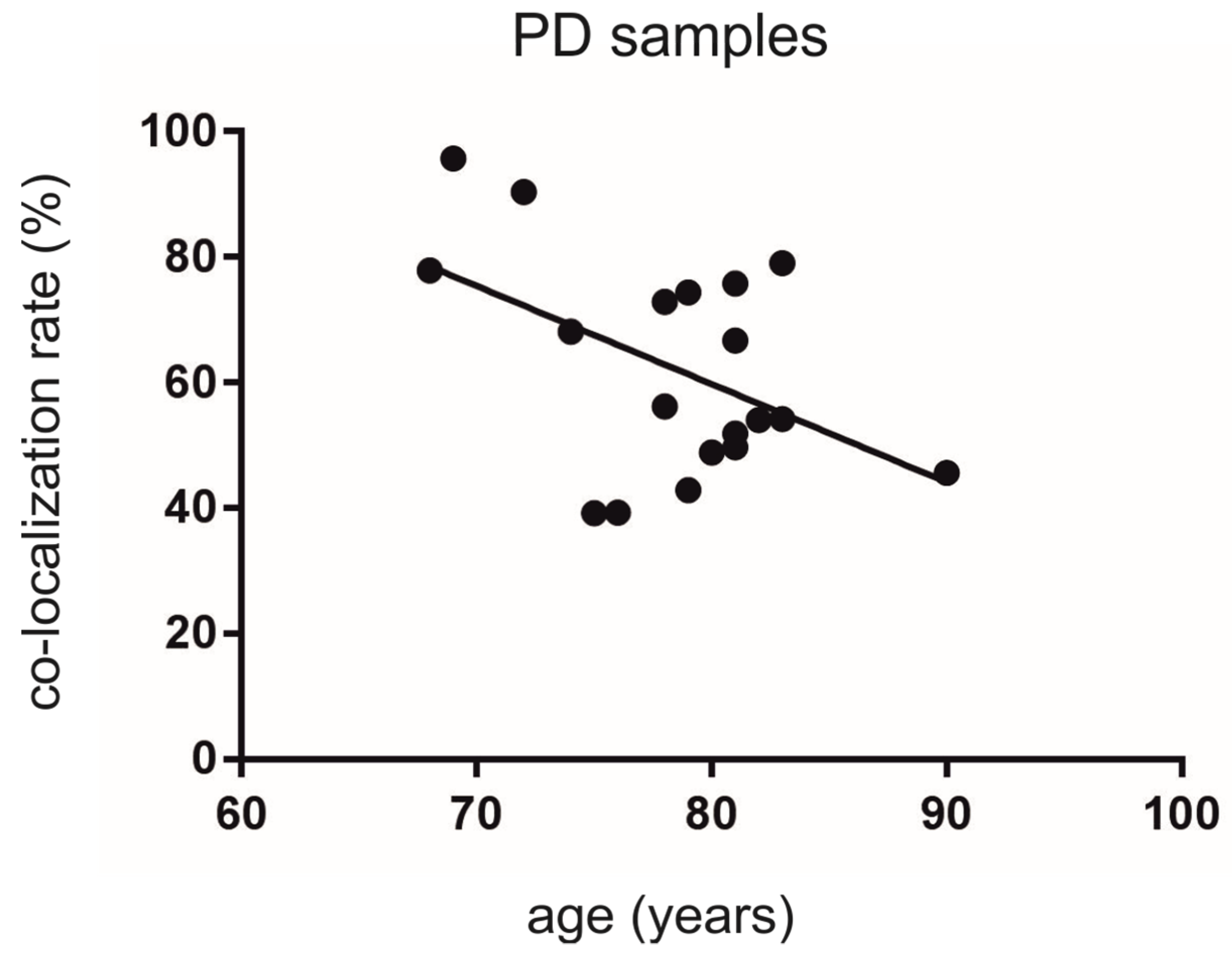
3.4. Co-Localization Rates in PD Decrease Depending on Age
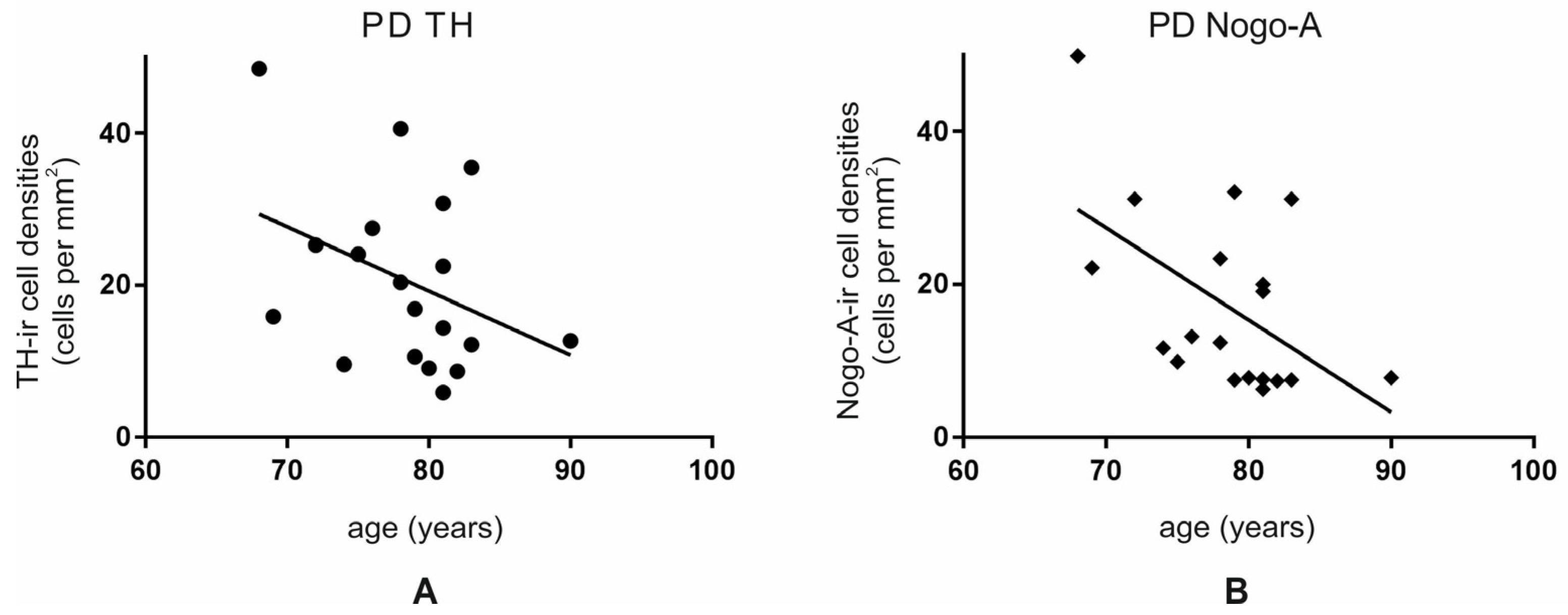
3.5. Age Is Associated with Lower Numbers of TH-ir and Nogo-A-ir Neurons in PD
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
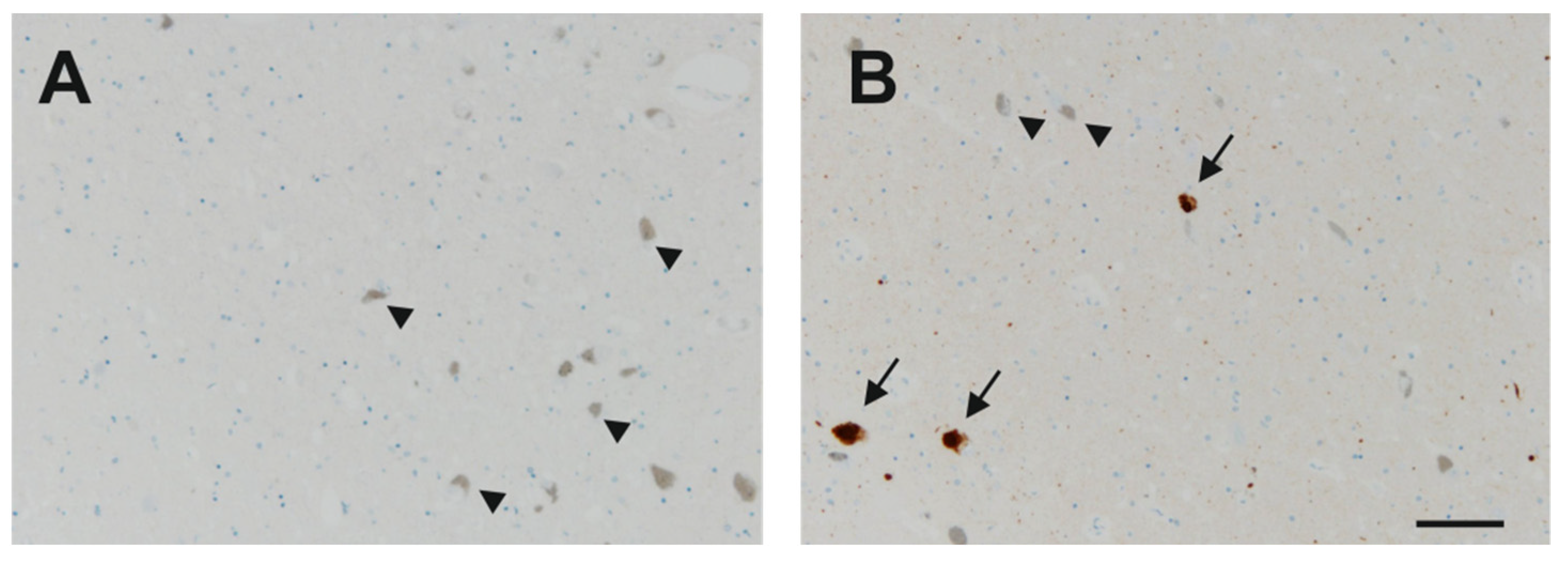
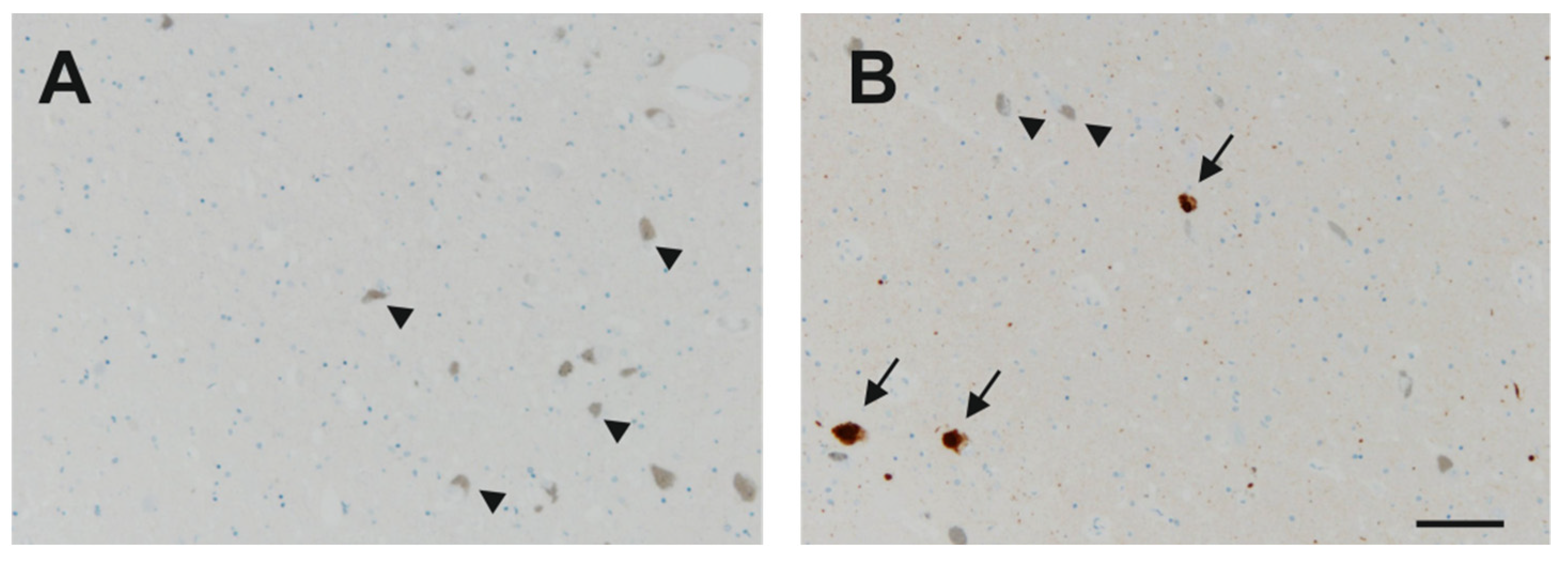
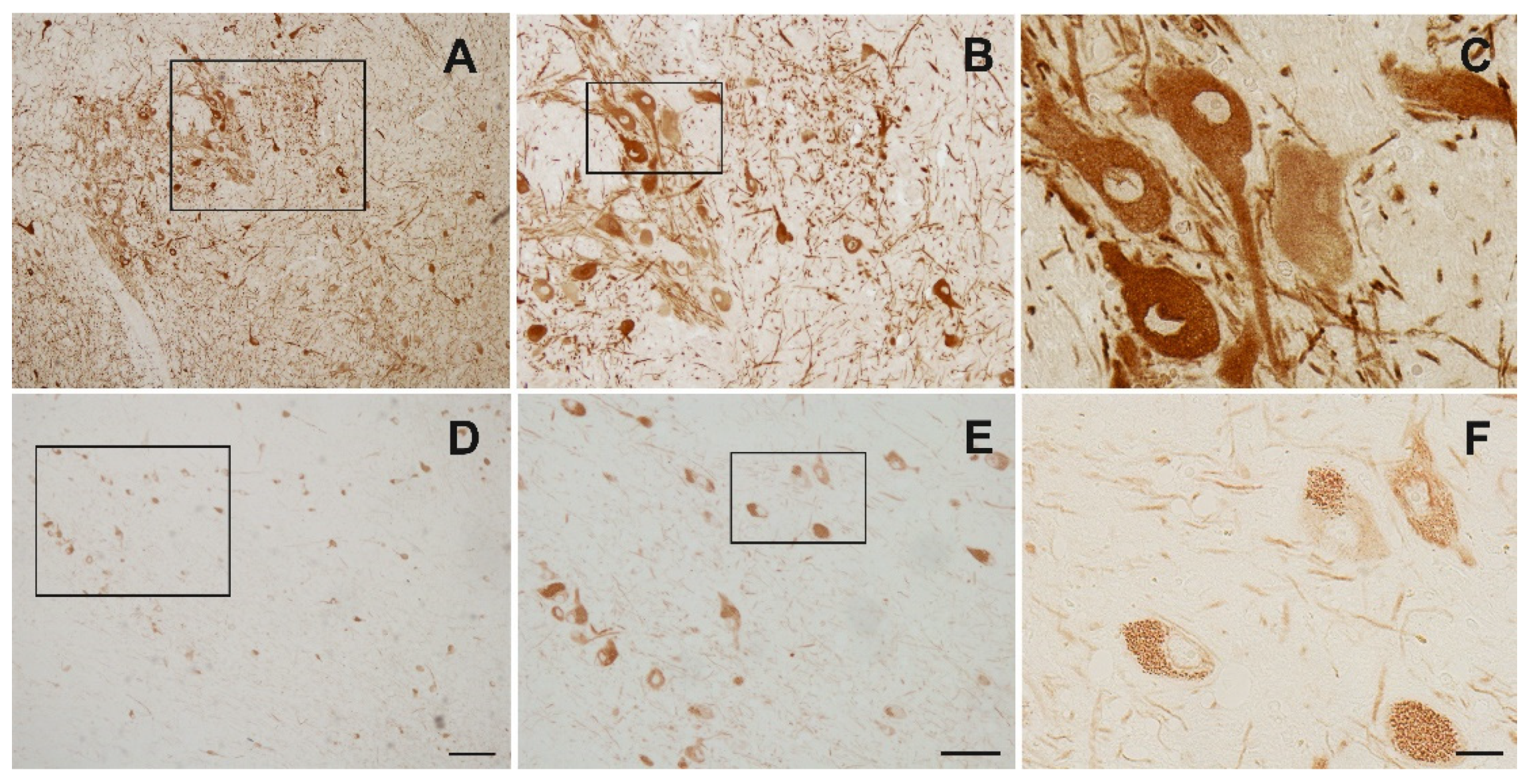
Appendix A. Immunohistochemistry for α-Synuclein

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
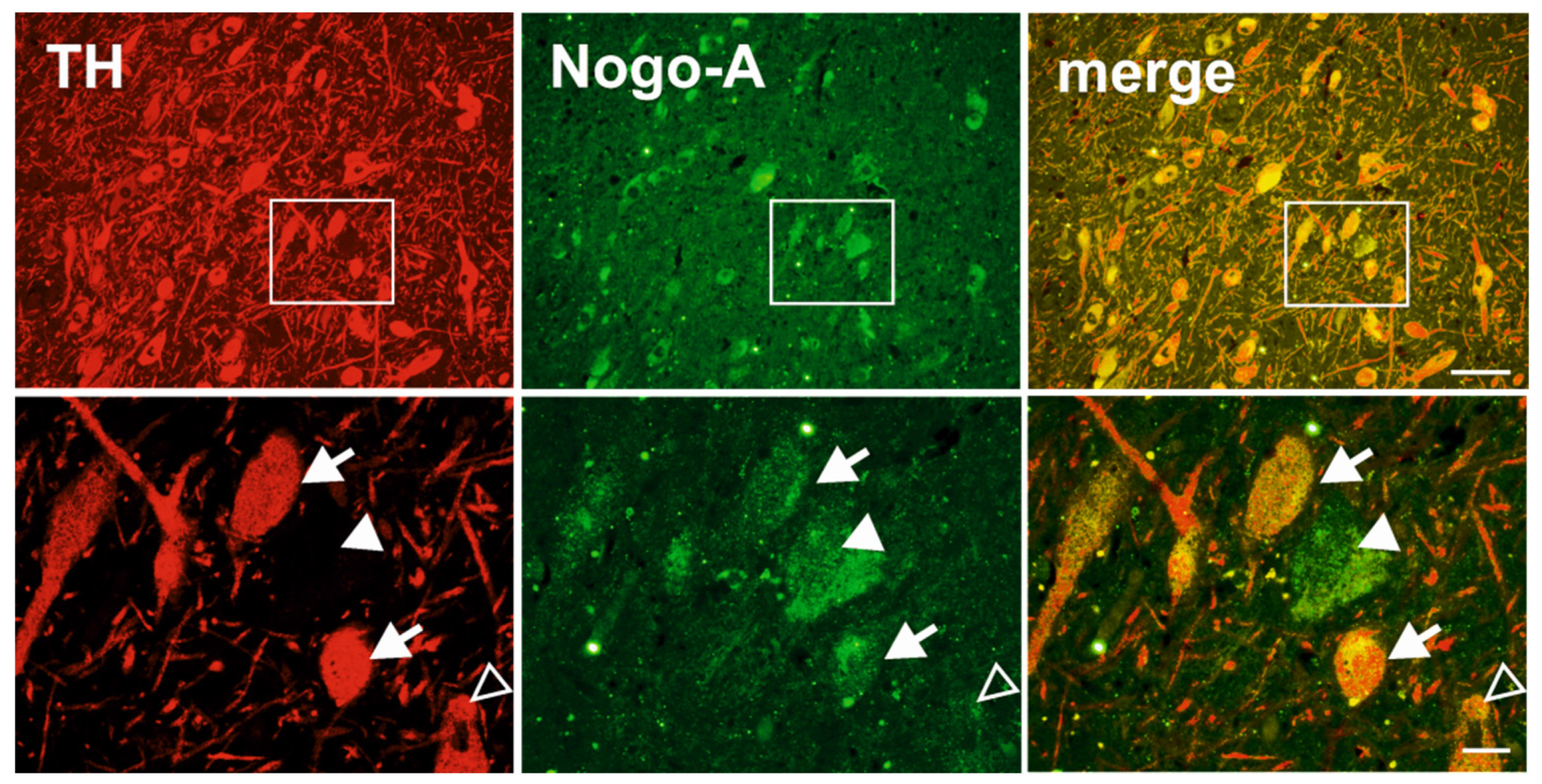{kind=link}
{kind=link}
{kind=link}
| Sample Number | Age |
|---|---|
| 1 | 32 |
| 2 | 46 |
| 3 | 50 |
| 4 | 52 |
| 5 | 55 |
| 6 | 58 |
| 7 | 59 |
| 8 | 64 |
| 9 | 64 |
| 10 | 64 |
| 11 | 67 |
| 12 | 70 |
| 13 | 73 |
| 14 | 74 |
| 15 | 74 |
| 16 | 74 |
| 17 | 75 |
| 18 | 79 |
| 19 | 79 |
| 20 | 83 |
| 21 | 84 |
| 22 | 85 |
| 23 | 91 |
| Sample Number | Age |
|---|---|
| 1 | 41 |
| 2 | 47 |
| 3 | 47 |
| 4 | 49 |
| 5 | 54 |
| 6 | 54 |
| 7 | 54 |
| 8 | 58 |
| 9 | 60 |
| 10 | 60 |
| 11 | 64 |
| 12 | 64 |
| 13 | 64 |
| 14 | 66 |
| 15 | 66 |
| 16 | 67 |
| 17 | 70 |
| 18 | 71 |
| 19 | 71 |
| 20 | 71 |
| 21 | 71 |
| 22 | 72 |
| 23 | 74 |
| 24 | 74 |
| 25 | 75 |
| 26 | 75 |
| 27 | 76 |
| 28 | 78 |
| 29 | 78 |
| 30 | 81 |
| 31 | 81 |
| 32 | 87 |
| 33 | 88 |
| Sample Number | Gender | Age |
|---|---|---|
| 1 | Female | 69 |
| 2 | Female | 75 |
| 3 | Female | 76 |
| 4 | Female | 80 |
| 5 | Female | 81 |
| 6 | Female | 83 |
| 7 | Male | 68 |
| 8 | Male | 72 |
| 9 | Male | 74 |
| 10 | Male | 78 |
| 11 | Male | 78 |
| 12 | Male | 79 |
| 13 | Male | 79 |
| 14 | Male | 81 |
| 15 | Male | 81 |
| 16 | Male | 81 |
| 17 | Male | 82 |
| 18 | Male | 83 |
| 19 | Male | 90 |
Appendix B. Immunohistochemistry for Establishment of Best Staining Method
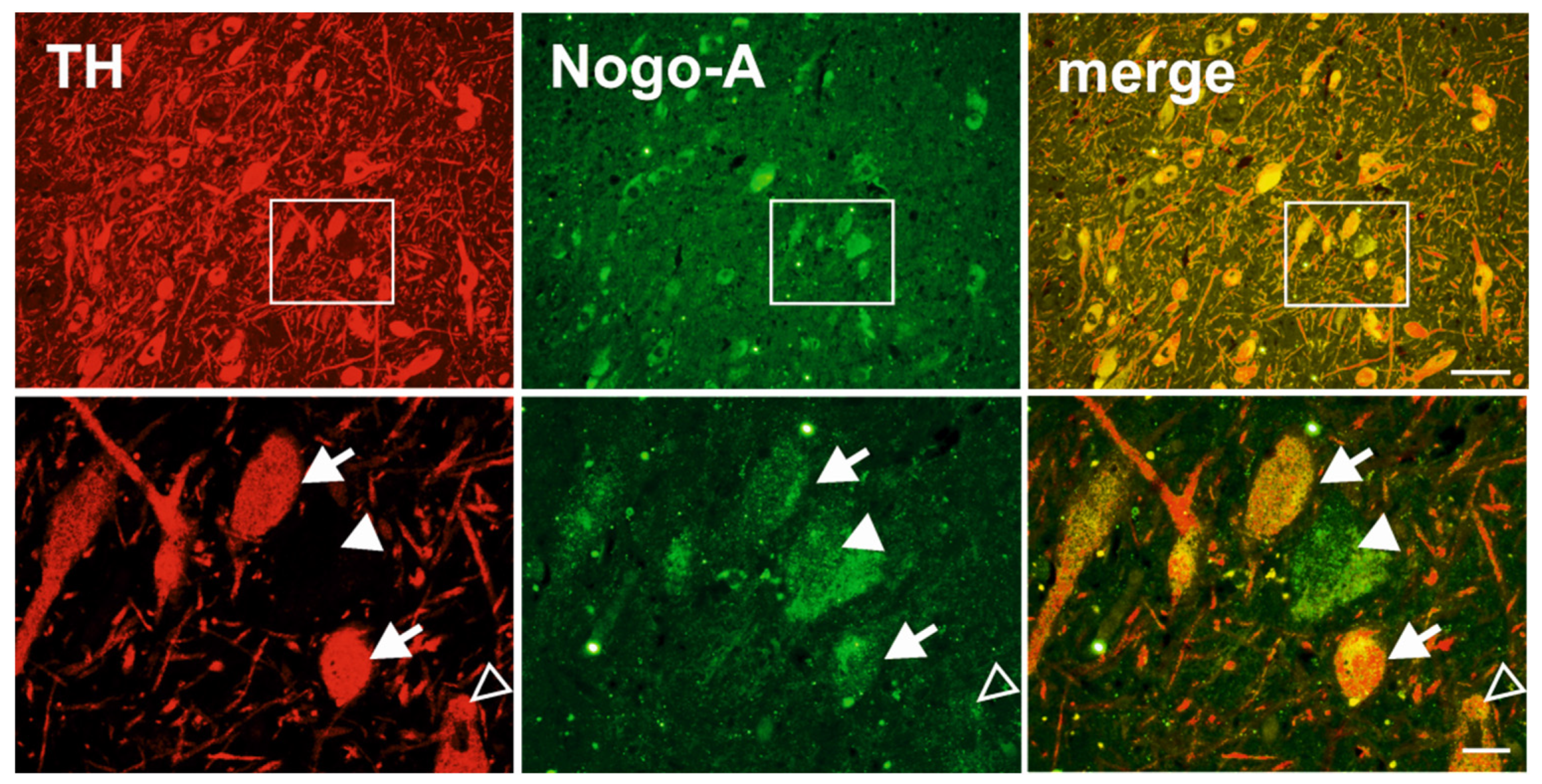
Appendix C. A Subpopulation of Neurons in the SNc Express Nogo-A Only
Appendix D. Cell Count Approach
References
- GBD 2016 Parkinson's Disease Collaborators. Global, regional, and national burden of Parkinson’s disease, 1990–2016: A systematic analysis for the Global Burden of Disease Study 2016. Lancet Neurol. 2018, 17, 939–953. [Google Scholar] [CrossRef] [Green Version]
- Bogetofte, H.; Alamyar, A.; Blaabjerg, M.; Meyer, M. Levodopa Therapy for Parkinson’s Disease: History, Current Status and Perspectives. CNS Neurol. Disord. Drug Targets 2020, 19, 572–583. [Google Scholar] [CrossRef]
- Lees, A.J.; Hardy, J.; Revesz, T. Parkinson’s disease. Lancet 2009, 373, 2055–2066. [Google Scholar] [CrossRef]
- Surmeier, D.J.; Obeso, J.A.; Halliday, G.M. Selective neuronal vulnerability in Parkinson disease. Nat. Rev. Neurosci. 2017, 18, 101–113. [Google Scholar] [CrossRef]
- Mouatt-Prigent, A.; Agid, Y.; Hirsch, E.C. Does the calcium binding protein calretinin protect dopaminergic neurons against degeneration in Parkinson’s disease? Brain Res. 1994, 668, 62–70. [Google Scholar] [CrossRef]
- Rodríguez, M.; Barroso-Chinea, P.; Abdala, P.; Obeso, J.; González-Hernández, T. Dopamine cell degeneration induced by intraventricular administration of 6-hydroxydopamine in the rat: Similarities with cell loss in parkinson’s disease. Exp. Neurol. 2001, 169, 163–181. [Google Scholar] [CrossRef]
- González-Hernández, T.; Barroso-Chinea, P.; Rodríguez, M. Response of the GABAergic and dopaminergic mesostriatal projections to the lesion of the contralateral dopaminergic mesostriatal pathway in the rat. Mov. Disord. 2004, 19, 1029–1042. [Google Scholar] [CrossRef] [PubMed]
- Anglade, P.; Vyas, S.; Hirsch, E.C.; Agid, Y. Apoptosis in dopaminergic neurons of the human substantia nigra during normal aging. Histol. Histopathol. 1997, 12, 603–610. [Google Scholar]
- Hawkes, C.H. The prodromal phase of sporadic Parkinson’s disease: Does it exist and if so how long is it? Mov. Disord. 2008, 23, 1799–1807. [Google Scholar] [CrossRef]
- Reeve, A.; Simcox, E.; Turnbull, D. Ageing and Parkinson’s disease: Why is advancing age the biggest risk factor? Ageing Res. Rev. 2014, 14, 19–30. [Google Scholar] [CrossRef]
- Damier, P.; Hirsch, E.C.; Agid, Y.; Graybiel, A.M. The substantia nigra of the human brain. II. Patterns of loss of dopamine-containing neurons in Parkinson’s disease. Brain. 1999, 122, 1437–1448. [Google Scholar] [CrossRef]
- Schwab, M.E. Nogo and axon regeneration. Curr. Opin. Neurobiol. 2004, 14, 118–124. [Google Scholar] [CrossRef]
- Wälchli, T.; Pernet, V.; Weinmann, O.; Shiu, J.Y.; Guzik-Kornacka, A.; Decrey, G.; Yüksel, D.; Schneider, H.; Vogel, J.; Ingber, D.E.; et al. Nogo-A is a negative regulator of CNS angiogenesis. Proc. Natl. Acad. Sci. USA 2013, 110, E1943–E1952. [Google Scholar] [CrossRef] [Green Version]
- Schwab, M.E. Functions of Nogo proteins and their receptors in the nervous system. Nat. Rev. Neurosci. 2010, 11, 799–811. [Google Scholar] [CrossRef]
- Willi, R.; Aloy, E.M.; Yee, B.K.; Feldon, J.; Schwab, M.E. Behavioral characterization of mice lacking the neurite outgrowth inhibitor Nogo-A. Genes Brain Behav. 2009, 8, 181–192. [Google Scholar] [CrossRef] [PubMed]
- Seiler, S.; Di Santo, S.; Widmer, H.R. Nogo-A Neutralization Improves Graft Function in a Rat Model of Parkinson’s Disease. Front. Cell. Neurosci. 2016, 10, 87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seiler, S.; Pollini, D.; Di Santo, S.; Widmer, H.R. Antagonizing Nogo-receptor 1 promotes the number of cultured dopaminergic neurons and elongates their neurites. Neuroreport 2013, 24, 1047–1052. [Google Scholar] [CrossRef] [PubMed]
- Schawkat, K.; Di Santo, S.; Seiler, S.; Ducray, A.D.; Widmer, H.R. Loss of Nogo-A-expressing neurons in a rat model of Parkinson’s disease. Neuroscience. 2015, 288, 59–72. [Google Scholar] [CrossRef]
- Kurowska, Z.; Brundin, P.; Schwab, M.E.; Li, J.Y. Intracellular Nogo-A facilitates initiation of neurite formation in mouse midbrain neurons in vitro. Neuroscience 2014, 256, 456–466. [Google Scholar] [CrossRef]
- Schnell, L.; Schwab, M.E. Axonal regeneration in the rat spinal cord produced by an antibody against myelin-associated neurite growth inhibitors. Nature 1990, 343, 269–272. [Google Scholar] [CrossRef]
- Dupuis, L.; Gonzalez de Aguilar, J.L.; di Scala, F.; Rene, F.; de Tapia, M.; Pradat, P.F.; Lacomblez, L.; Seihlan, D.; Prinjha, R.; Walsh, F.S.; et al. Nogo provides a molecular marker for diagnosis of amyotrophic lateral sclerosis. Neurobiol. Dis. 2002, 10, 358–365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gil, V.; Nicolas, O.; Mingorance, A.; Ureña, J.M.; Tang, B.L.; Hirata, T.; Sáez-Valero, J.; Ferrer, I.; Soriano, E.; del Río, J.A. Nogo-A expression in the human hippocampus in normal aging and in Alzheimer disease. J. Neuropathol. Exp. Neurol. 2006, 65, 433–444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.-K.; Kim, J.-E.; Sivula, M.; Strittmatter, S.M. Nogo receptor antagonism promotes stroke recovery by enhancing axonal plasticity. J. Neurosci. 2004, 24, 6209–6217. [Google Scholar] [CrossRef] [PubMed]
- Willi, R.; Schwab, M.E. Nogo and Nogo receptor: Relevance to schizophrenia? Neurobiol. Dis. 2013, 54, 150–157. [Google Scholar] [CrossRef]
- Mohammed, R.; Opara, K.; Lall, R.; Ojha, U.; Xiang, J. Evaluating the effectiveness of anti-Nogo treatment in spinal cord injuries. Neural. Dev. 2020, 15, 1. [Google Scholar] [CrossRef] [PubMed]
- Zlobec, I.; Suter, G.; Perren, A.; Lugli, A. A next-generation tissue microarray (ngTMA) protocol for biomarker studies. J. Vis. Exp. 2014, 51893. [Google Scholar] [CrossRef] [Green Version]
- Zlobec, I.; Koelzer, V.H.; Dawson, H.; Perren, A.; Lugli, A. Next-generation tissue microarray (ngTMA) increases the quality of biomarker studies: An example using CD3, CD8, and CD45RO in the tumor microenvironment of six different solid tumor types. J. Transl Med. 2013, 11, 104. [Google Scholar] [CrossRef] [Green Version]
- Eriksen, N.; Stark, A.K.; Pakkenberg, B. Age and Parkinson’s disease-related neuronal death in the substantia nigra pars compacta. J. Neural. Transm. Suppl. 2009, 203–213. [Google Scholar] [CrossRef]
- Naskar, A.; Mahadevan, A.; Philip, M.; Alladi, P.A. Aging mildly affects dendritic arborisation and synaptic protein expression in human substantia nigra pars compacta. J. Chem. Neuroanat. 2019, 97, 57–65. [Google Scholar] [CrossRef]
- Smedfors, G.; Olson, L.; Karlsson, T.E. A Nogo-Like Signaling Perspective from Birth to Adulthood and in Old Age: Brain Expression Patterns of Ligands, Receptors and Modulators. Front. Mol. Neurosci. 2018, 11. [Google Scholar] [CrossRef] [Green Version]
- Buss, A.; Sellhaus, B.; Wolmsley, A.; Noth, J.; Schwab, M.E.; Brook, G.A. Expression pattern of NOGO-A protein in the human nervous system. Acta Neuropathol. 2005, 110, 113–119. [Google Scholar] [CrossRef] [PubMed]
- Carvey, P.M.; Punati, A.; Newman, M.B. Progressive dopamine neuron loss in Parkinson’s disease: The multiple hit hypothesis. Cell. Transplant. 2006, 15, 239–250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chan, C.S.; Gertler, T.S.; Surmeier, D.J. A molecular basis for the increased vulnerability of substantia nigra dopamine neurons in aging and Parkinson’s disease. Mov. Disord. 2010, 25, S63–S70. [Google Scholar] [CrossRef]
- Kumari, A.; Thakur, M.K. Age-Dependent Decline of Nogo-A Protein in the Mouse Cerebrum. Cell. Mol. Neurobiol. 2014, 34, 1131–1141. [Google Scholar] [CrossRef] [PubMed]
- Trifunovski, A.; Josephson, A.; Bickford, P.C.; Olson, L.; Brené, S. Selective decline of Nogo mRNA in the aging brain. Neuroreport 2006, 17, 913–916. [Google Scholar] [CrossRef] [PubMed]
- Seiler, S.; Di Santo, S.; Widmer, H.R. Non-canonical actions of Nogo-A and its receptors. Biochem. Pharm. 2016, 100, 28–39. [Google Scholar] [CrossRef] [PubMed]
- Pradhan, L.K.; Das, S.K. The Regulatory Role of Reticulons in Neurodegeneration: Insights Underpinning Therapeutic Potential for Neurodegenerative Diseases. Cell. Mol. Neurobiol. 2021, 41, 1157–1174. [Google Scholar] [CrossRef]
- Ineichen, B.V.; Kapitza, S.; Bleul, C.; Good, N.; Plattner, P.S.; Seyedsadr, M.S.; Kaiser, J.; Schneider, M.P.; Zörner, B.; Martin, R.; et al. Nogo-A antibodies enhance axonal repair and remyelination in neuro-inflammatory and demyelinating pathology. Acta. Neuropathol. 2017, 134, 423–440. [Google Scholar] [CrossRef] [Green Version]
- Kucher, K.; Johns, D.; Maier, D.; Abel, R.; Badke, A.; Baron, H.; Thietje, R.; Casha, S.; Meindl, R.; Gomez-Mancilla, B.; et al. First-in-Man Intrathecal Application of Neurite Growth-Promoting Anti-Nogo-A Antibodies in Acute Spinal Cord Injury. Neurorehabil. Neural. Repair 2018, 32, 578–589. [Google Scholar] [CrossRef] [Green Version]
- Xu, W.; Xiao, P.; Fan, S.; Chen, Y.; Huang, W.; Chen, X.; Liu, G.; Dang, C.; Zeng, J.; Xing, S. Blockade of Nogo-A/Nogo-66 receptor 1 (NgR1) Inhibits Autophagic Activation and Prevents Secondary Neuronal Damage in the Thalamus after Focal Cerebral Infarction in Hypertensive Rats. Neuroscience 2020, 431, 103–114. [Google Scholar] [CrossRef]
- Singh-Bains, M.K.; Mehrabi, N.F.; Tan, A.Y.S.; Faull, R.L.M.; Dragunow, M. Preparation, construction and high-throughput automated analysis of human brain tissue microarrays for neurodegenerative disease drug development. Nat. Protoc. 2021, 16, 2308–2343. [Google Scholar] [CrossRef]
- Twelves, D.; Perkins, K.S.; Counsell, C. Systematic review of incidence studies of Parkinson’s disease. Mov. Disord. 2003, 18, 19–31. [Google Scholar] [CrossRef]
- Tysnes, O.-B.; Storstein, A. Epidemiology of Parkinson’s disease. J. Neural. Transm. 2017, 124, 901–905. [Google Scholar] [CrossRef]
- Haaxma, C.A.; Bloem, B.R.; Borm, G.F.; Oyen, W.J.; Leenders, K.L.; Eshuis, S.; Booij, J.; Dluzen, D.E.; Horstink, M.W. Gender differences in Parkinson’s disease. J. Neurol. Neurosurg. Psychiatry 2007, 78, 819–824. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martinez-Martin, P.; Pecurariu, C.F.; Odin, P.; Van Hilten, J.J.; Antonini, A.; Rojo-Abuin, J.M.; Borges, V.; Trenkwalder, C.; Aarsland, D.; Brooks, D.J. Gender-related differences in the burden of non-motor symptoms in Parkinson’s disease. J. Neurol. 2012, 259, 1639–1647. [Google Scholar] [CrossRef] [PubMed]
- Cholerton, B.; Johnson, C.O.; Fish, B.; Quinn, J.F.; Chung, K.A.; Peterson-Hiller, A.L.; Rosenthal, L.S.; Dawson, T.M.; Albert, M.S.; Hu, S.-C. Sex differences in progression to mild cognitive impairment and dementia in Parkinson’s disease. Parkinsonism Relat. Disord. 2018, 50, 29–36. [Google Scholar] [CrossRef] [PubMed]
- Gillies, G.E.; Murray, H.E.; Dexter, D.; McArthur, S. Sex dimorphisms in the neuroprotective effects of estrogen in an animal model of Parkinson’s disease. Pharmacol. Biochem. Behav. 2004, 78, 513–522. [Google Scholar] [CrossRef]
- Buck, S.A.; Steinkellner, T.; Aslanoglou, D.; Villeneuve, M.; Bhatte, S.H.; Childers, V.C.; Rubin, S.A.; De Miranda, B.R.; O’Leary, E.I.; Neureiter, E.G.; et al. Vesicular glutamate transporter modulates sex differences in dopamine neuron vulnerability to age-related neurodegeneration. Aging Cell 2021, 20, e13365. [Google Scholar] [CrossRef]
- Shin, J.W.; Shim, E.S.; Hwang, G.H.; Jung, H.S.; Park, J.H.; Sohn, N.W. Cell size-dependent Nogo-A expression in layer V pyramidal neurons of the rat primary somatosensory cortex. Neurosci. Lett. 2006, 394, 117–120. [Google Scholar] [CrossRef]
- Jiang, W.; Xia, F.; Han, J.; Wang, J. Patterns of Nogo-A, NgR, and RhoA expression in the brain tissues of rats with focal cerebral infarction. Transl. Res. 2009, 154, 40–48. [Google Scholar] [CrossRef]
- Nagamoto-Combs, K.; Morecraft, R.J.; Darling, W.G.; Combs, C.K. Long-term gliosis and molecular changes in the cervical spinal cord of the rhesus monkey after traumatic brain injury. J. Neurotrauma. 2010, 27, 565–585. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Theotokis, P.; Lourbopoulos, A.; Touloumi, O.; Lagoudaki, R.; Kofidou, E.; Nousiopoulou, E.; Poulatsidou, K.N.; Kesidou, E.; Tascos, N.; Spandou, E.; et al. Time course and spatial profile of Nogo-A expression in experimental autoimmune encephalomyelitis in C57BL/6 mice. J. Neuropathol. Exp. Neurol. 2012, 71, 907–920. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liechti, R.; Ducray, A.D.; Jensen, P.; Di Santo, S.; Seiler, S.; Jensen, C.H.; Meyer, M.; Widmer, H.R. Characterization of fetal antigen 1/delta-like 1 homologue expressing cells in the rat nigrostriatal system: Effects of a unilateral 6-hydroxydopamine lesion. PLoS ONE 2015, 10, e0116088. [Google Scholar] [CrossRef] [Green Version]
- Seiler, S.; Di Santo, S.; Andereggen, L.; Widmer, H.R. Antagonization of the Nogo-Receptor 1 Enhances Dopaminergic Fiber Outgrowth of Transplants in a Rat Model of Parkinson’s Disease. Front. Cell. Neurosci. 2017, 11, 151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Eyer, G.-C.; Di Santo, S.; Hewer, E.; Andereggen, L.; Seiler, S.; Widmer, H.R. Co-Expression of Nogo-A in Dopaminergic Neurons of the Human Substantia Nigra Pars Compacta Is Reduced in Parkinson’s Disease. Cells 2021, 10, 3368. https://doi.org/10.3390/cells10123368
Eyer G-C, Di Santo S, Hewer E, Andereggen L, Seiler S, Widmer HR. Co-Expression of Nogo-A in Dopaminergic Neurons of the Human Substantia Nigra Pars Compacta Is Reduced in Parkinson’s Disease. Cells. 2021; 10(12):3368. https://doi.org/10.3390/cells10123368
Chicago/Turabian StyleEyer, Gian-Carlo, Stefano Di Santo, Ekkehard Hewer, Lukas Andereggen, Stefanie Seiler, and Hans Rudolf Widmer. 2021. "Co-Expression of Nogo-A in Dopaminergic Neurons of the Human Substantia Nigra Pars Compacta Is Reduced in Parkinson’s Disease" Cells 10, no. 12: 3368. https://doi.org/10.3390/cells10123368
APA StyleEyer, G.-C., Di Santo, S., Hewer, E., Andereggen, L., Seiler, S., & Widmer, H. R. (2021). Co-Expression of Nogo-A in Dopaminergic Neurons of the Human Substantia Nigra Pars Compacta Is Reduced in Parkinson’s Disease. Cells, 10(12), 3368. https://doi.org/10.3390/cells10123368