
Mesenchymal Stem/Stromal Cells Derived from Human and Animal Perinatal Tissues—Origins, Characteristics, Signaling Pathways, and Clinical Trials

 , , , , ,
, , , , ,  and
and 
Abstract
1. Human and Animal Perinatal Tissues—Anatomical, Histological, and Cellular Characteristics
2. Mesenchymal Stem/Stromal Cells—Origin, Cellular and Molecular Characteristics, and Signaling Pathways Involved in Differentiation
2.1. Different Origins of MSCs—Sources, Cell Characteristics, Possible Applications
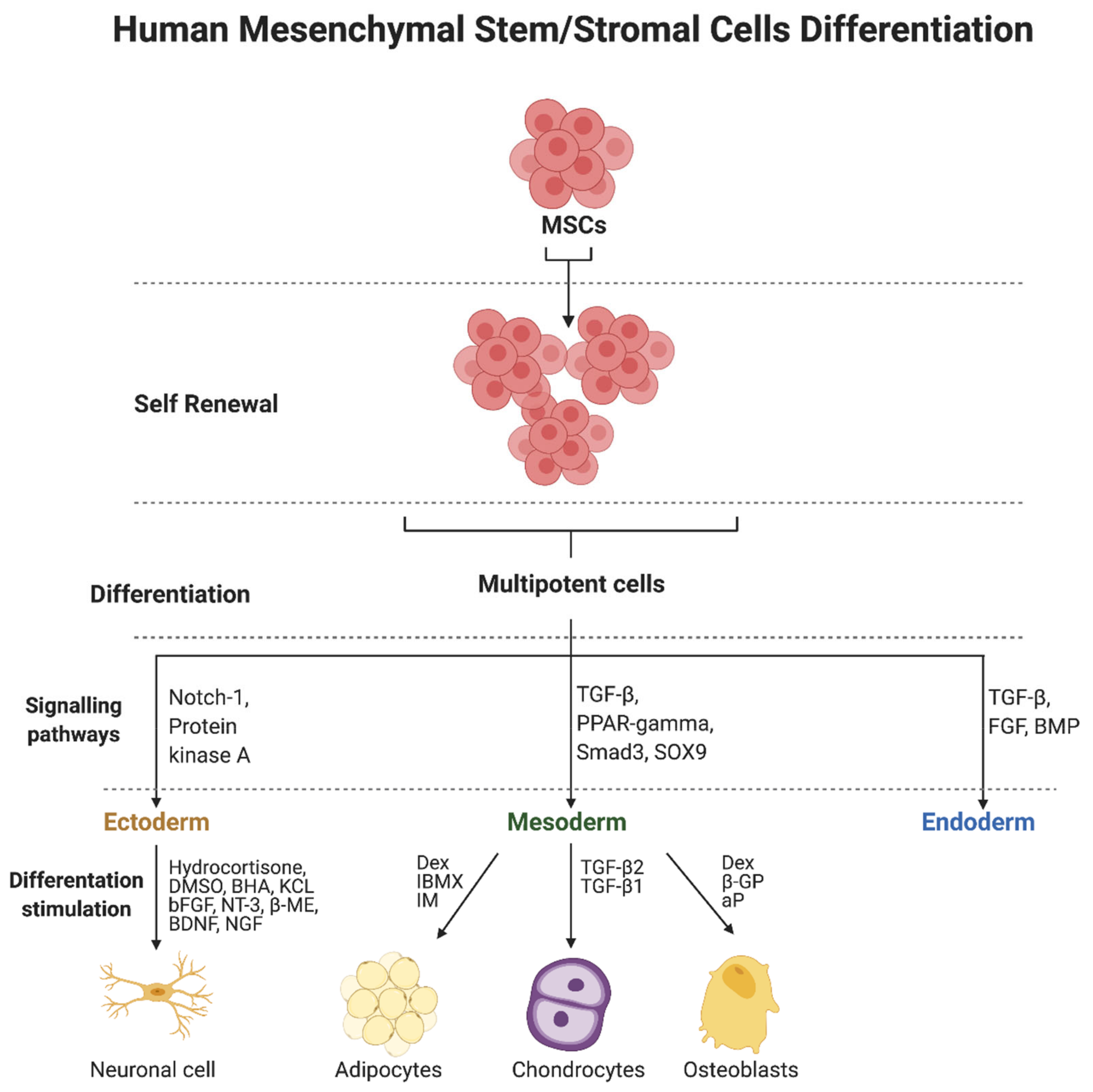
2.2. Differentiation of MSCs—Regulatory Factors and Signaling Pathways
3. Mesenchymal Stem/Stromal Cells Derived from Human and Animal Perinatal Tissues
3.1. Animal Perinatal Mesenchymal Stem/Stromal Cells
3.2. Human Perinatal Mesenchymal Stem/Stromal Cells
3.2.1. Placenta-Derived Mesenchymal Stem/Stromal Cells
3.2.2. Umbilical Cord-Derived Mesenchymal Stem/Stromal Cells
3.2.3. Amniotic Fluid-Derived Mesenchymal Stem/Stromal Cells
4. Signaling Pathways Involved in Fetal-Derived MSC Development and Differentiation
5. Animal Models and Clinical Applications
5.1. Animal Models
5.1.1. Bone and Cartilage Diseases
5.1.2. Cardiac Diseases
5.1.3. Neurological Disorders
5.1.4. Organ Disorders
5.1.5. Ischemia and Wound Healing
5.2. The Application of Perinatal MSC in Human Clinical Trials
5.2.1. Placenta-Derived MSCs
5.2.2. Umbilical Cord and Amniotic Fluid-Derived MSCs

{kind=link}
{kind=link}
{kind=link}
| Condition/ Procedures | Type of Study | Number of Participants | Material | First Author; Year; Reference |
|---|---|---|---|---|
| Knee osteoarthritis | Randomized, double-blind, placebo-controlled clinical trial | 20 | Allogenic placenta-derived MSCs | Soltani; 2019; [301] |
| Hip arthroplasty | Randomized, double blind, placebo-controlled, phase I/IIa clinical trial | 20 | Allogenic placenta-derived MSCs | Winkler; 2018; [302] |
| Erectile dysfunction | Prospective, non-randomized, single-arm clinical trial | 8 | Allogenic placenta-derived MSCs | Levy; 2016; [304] |
| Chronic obstructive pulmonary disease | Prospective, non-randomized, single-arm clinical trial | 30 | Allogenic placenta-derived MSCs-derived product: Exo-d-MAPPS drug | Harrell; 2020; [306] |
| COVID-19-induced ARDS | Prospective, non-randomized, single-arm clinical trial | 11 | Allogenic placenta-derived MSCs (5 cases) Umbilical cord-derived MSCs (6 cases) | Hashemian; 2021; [307] |
| Acute graft-versus-host disease | Prospective, non-randomized, single-arm clinical trial | 38 | Allogenic decidua-derived MSCs | Ringden; 2018; [308] |
| Knee osteoarthritis | Single-arm, open-label clinical trial | 29 | Allogenic umbilical cord-derived MSCs | Dilogo; 2020; [309] |
| Knee osteoarthritis | Randomized, placebo-controlled, phase I/II clinical trial | 26 | Allogenic umbilical cord-derived MSCs | Matas; 2019; [310] |
| Knee osteoarthritis | Open-label, single-arm, single-center, phase I/II clinical trial with 7-year extended follow-up | 7 | Allogeneic human umbilical cord blood-derived MSCs | Park; 2017; [311] |
| Knee osteoarthritis | Randomized, placebo-controlled clinical trial | 36 | Allogenic umbilical cord-derived MSCs | Wang; 2016; [312] |
| Chronic ischemic heart disease | Randomized, double-blind clinical trial | 115 | Collagen scaffolds covered with umbilical cord-derived MSCs | He; 2020; [313] |
| Heart failure | Randomized, controlled, phase I/II clinical trial | 30 | Allogenic umbilical cord-derived MSCs | Bartolucci; 2017; [314] |
| Chronic diabetic skin ulcers | Randomized, clinical trial | 5 | Allogenic umbilical cord-derived MSCs seeded on biological scaffold | Hashemi; 2019; [315] |
| Atopic dermatitis | Prospective, non-randomized, single-arm clinical trial | 28 | Topical drugs with allogenic umbilical cord-derived MSCs conditioned media | Kim; 2020; [316] |
| Ablative CO2 fractional laser treatment | Randomized, double-blinded, controlled clinical trial | 23 | Umbilical cord blood-derived MSCs conditioned media containing serum | Kim; 2020; [317] |
| Rheumatoid arthritis | Randomized, controlled clinical trial | 119 | Allogenic umbilical cord-derived MSCs | Qi; 2020; [319] |
| Rheumatoid arthritis | Randomized, controlled, phase I/II clinical trial | 63 | Allogenic umbilical cord-derived MSCs | He; 2020; [318] |
| Rheumatoid arthritis | Prospective, phase I/II clinical trial | 64 | Allogenic umbilical cord-derived MSCs | Wang; 2019; [320] |
| Rheumatoid arthritis | Open-label, single-arm, single-center, phase Ia clinical trial | 9 | Allogenic umbilical cord blood-derived MSCs | Park; 2018; [321] |
| Lupus nephritis | Randomized, double-blind, placebo-controlled clinical trial | 18 | Allogenic umbilical cord blood-derived MSCs | Deng; 2017; [322] |
| Multiple sclerosis | Prospective, non-randomized, single-arm clinical trial | 20 | Allogenic umbilical cord-derived MSCs | Riordan; 2018; [323] |
| Cerebral palsy | Randomized, controlled clinical trial | 39 | Allogenic umbilical cord-derived MSCs | Gu; 2020; [324] |
| Autism spectrum disorder | Single-arm, phase I/II clinical trial | 20 | Allogenic umbilical cord-derived MSCs | Riordan; 2019; [325] |
| Crohn’s disease | Randomized, controlled clinical trial | 82 | Allogenic umbilical cord-derived MSCs | Zhang; 2018; [326] |
| Kidney transplantation | Multicenter, randomized, controlled clinical trial | 42 | Allogenic umbilical cord-derived MSCs | Sun; 2018; [327] |
| Acute liver allograft rejection | Randomized, controlled, clinical trial | 27 | Allogenic umbilical cord-derived MSCs | Shi; 2017; [328] |
| COVID-19 | Randomized, double-blind, placebo-controlled, phase II clinical trial | 101 | Allogenic umbilical cord-derived MSCs | Shi; 2021; [329] |
| COVID-19 | Parallel assigned controlled, non-randomized, phase I clinical trial | 18 | Allogenic umbilical cord-derived MSCs | Meng; 2020; [330] |
| COVID-19 | Single-center open-label, individually randomized, standard treatment-controlled clinical trial | 41 | Allogenic umbilical cord-derived MSCs | Shu; 2020; [331] |
| Sepsis | Single-center, open-label, dose-escalation phase 1 clinical trial | 15 | Allogenic umbilical cord-derived MSCs | He; 2018; [332] |
6. Conclusions
Funding
Acknowledgments
Conflicts of Interest
References
- Cross, J.; Werb, Z.; Fisher, S. Implantation and the placenta: Key pieces of the development puzzle. Science 1994, 266, 1508–1518. [Google Scholar] [CrossRef] [PubMed]
- Siiteri, P.K.; Stites, D.P. Immunologic and Endocrine Interrelationships in Pregnancy. Biol. Reprod. 1982, 26, 1–14. [Google Scholar] [CrossRef]
- Bartel, H. Embriologia, Textbook for Medical Students, 2nd ed.; Medical Publishing House PZWL: Warsaw, Poland, 1999; ISBN 9788320035933. [Google Scholar]
- Zhang, S.; Mesalam, A.; Joo, M.-D.; Lee, K.-L.; Hwang, J.-Y.; Xu, L.; Song, S.-H.; Koh, P.-O.; Yuan, Y.-G.; Lv, W.; et al. Matrix metalloproteinases improves trophoblast invasion and pregnancy potential in mice. Theriogenology 2020, 151, 144–150. [Google Scholar] [CrossRef] [PubMed]
- Pijnenborg, R.; Dixon, G.; Robertson, W.B.; Brosens, I. Trophoblastic invasion of human decidua from 8 to 18 weeks of pregnancy. Placenta 1980, 1, 3–19. [Google Scholar] [CrossRef]
- Palis, J.; McGrath, K.E.; Kingsley, P.D. Initiation of hematopoiesis and vasculogenesis in murine yolk sac explants. Blood 1995, 86, 156–163. [Google Scholar] [CrossRef] [PubMed]
- Ross, C.; Boroviak, T.E. Origin and function of the yolk sac in primate embryogenesis. Nat. Commun. 2020, 11, 3760. [Google Scholar] [CrossRef] [PubMed]
- Hoyes, A.D. Structure and function of the amnion. Obstet. Gynecol. Annu. 1975, 4, 1–38. [Google Scholar] [PubMed]
- Knezevic, V. Differentiation potential of rat amnion. J. Anat. 1996, 189, 1–7. [Google Scholar] [PubMed]
- Nizard, J. Amniocentesis: Technique and education. Curr. Opin. Obstet. Gynecol. 2010, 22, 152–154. [Google Scholar] [CrossRef] [PubMed]
- Bielańska-Osuchowska, Z. Embriologia, Textbook for Medical Students, 4th ed.; National Agricultural and Forestry Publishing House: Warsaw, Poland, 2001; ISBN 8309017480. [Google Scholar]
- Modena, A.B.; Fieni, S. Amniotic fluid dynamics. Acta Bio-Med. Atenei Parm. 2004, 75, 11–13. [Google Scholar]
- King, B.F. Developmental changes in the fine structure of the chorion laeve (Smooth chorion) of the rhesus monkey placenta. Anat. Rec. 1981, 200, 163–175. [Google Scholar] [CrossRef]
- Watson, E.D.; Cross, J.C. Development of Structures and Transport Functions in the Mouse Placenta. Physiology 2005, 20, 180–193. [Google Scholar] [CrossRef] [PubMed]
- Minh, H.-N.; Douvin, D.; Smadja, A.; Orcel, L. Fetal membrane morphology and circulation of the liquor amnii. Eur. J. Obstet. Gynecol. Reprod. Biol. 1980, 10, 213–223. [Google Scholar] [CrossRef]
- Downs, K.M.; Temkin, R.; Gifford, S.; McHugh, J. Study of the Murine Allantois by Allantoic Explants. Dev. Biol. 2001, 233, 347–364. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Spurway, J.; Logan, P.; Pak, S. The development, structure and blood flow within the umbilical cord with particular reference to the venous system. Australas. J. Ultrasound Med. 2012, 15, 97–102. [Google Scholar] [CrossRef]
- Downs, K.M.; Harmann, C. Developmental potency of the murine allantois. Development 1997, 124, 2769–2780. [Google Scholar] [CrossRef] [PubMed]
- Di Naro, E.; Ghezzi, F.; Raio, L.; Franchi, M.; D’Addario, V. Umbilical cord morphology and pregnancy outcome. Eur. J. Obstet. Gynecol. Reprod. Biol. 2001, 96, 150–157. [Google Scholar] [CrossRef]
- Raio, L.; Ghezzi, F.; di Naro, E.; Franchi, M.; Bruhwiler, H. Prenatal assessment of the Hyrtl anastomosis and evaluation of its function: Case report. Hum. Reprod. 1999, 14, 1890–1893. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Nanaev, A.; Kohnen, G.; Milovanov, A.P.; Domogatsky, S.P.; Kaufmann, P. Stromal differentiation and architecture of the human umbilical cord. Placenta 1997, 18, 53–64. [Google Scholar] [CrossRef]
- Davies, J.E.; Walker, J.T.; Keating, A. Concise Review: Wharton’s Jelly: The Rich, but Enigmatic, Source of Mesenchymal Stromal Cells. Stem Cells Transl. Med. 2017, 6, 1620–1630. [Google Scholar] [CrossRef]
- Turco, M.Y.; Moffett, A. Development of the human placenta. Development 2019, 146, dev163428. [Google Scholar] [CrossRef]
- Furukawa, S.; Kuroda, Y.; Sugiyama, A. A Comparison of the Histological Structure of the Placenta in Experimental Animals. J. Toxicol. Pathol. 2014, 27, 11–18. [Google Scholar] [CrossRef] [PubMed]
- Furukawa, S.; Tsuji, N.; Sugiyama, A. Morphology and physiology of rat placenta for toxicological evaluation. J. Toxicol. Pathol. 2019, 32, 1–17. [Google Scholar] [CrossRef]
- Leiser, R.; Dantzer, V. Initial vascularisation in the pig placenta: II. Demonstration of gland and areola-gland subunits by histology and corrosion casts. Anat. Rec. Adv. Integr. Anat. Evol. Biol. 1994, 238, 326–334. [Google Scholar] [CrossRef] [PubMed]
- Abd-Elnaeim, M.M.; Pfarrer, C.; Saber, A.S.; Abou-Elmagd, A.; Jones, C.J.; Leiser, R. Fetomaternal attachment and anchorage in the early diffuse epitheliochorial placenta of the camel (Camelus dromedarius). Light, transmission, and scanning electron microscopic study. Cells Tissues Organs 1999, 164, 141–154. [Google Scholar] [CrossRef] [PubMed]
- Carter, A.M.; Enders, A.C. Comparative aspects of trophoblast development and placentation. Reprod. Biol. Endocrinol. 2004, 2, 46. [Google Scholar] [CrossRef]
- Carter, A.M. Evolution of Placental Function in Mammals: The Molecular Basis of Gas and Nutrient Transfer, Hormone Secretion, and Immune Responses. Physiol. Rev. 2012, 92, 1543–1576. [Google Scholar] [CrossRef] [PubMed]
- Jiang, W.; Xu, J. Immune modulation by mesenchymal stem cells. Cell Prolif. 2019, 53, e12712. [Google Scholar] [CrossRef]
- Popis, M.; Konwerska, A.; Partyka, M.; Wieczorkiewicz, M.; Ciesiółka, S.; Stefańska, K.; Spaczyńska, J.; Golkar-Narenji, A.; Jeseta, M.; Bukowska, D.; et al. Mesenchymal stem cells and their secretome -candidates for safe and effective therapy for systemic lupus erythematosus. Med. J. Cell Biol. 2021, 9, 110–122. [Google Scholar] [CrossRef]
- Ceccariglia, S.; Cargnoni, A.; Silini, A.R.; Parolini, O. Autophagy: A potential key contributor to the therapeutic action of mesenchymal stem cells. Autophagy 2019, 16, 28–37. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Feng, Z.; Wang, X.; Xiong, Y.; Wang, L.; Ye, L.; Zhang, H. hUC-MSCs Exert a Neuroprotective Effect via Anti-apoptotic Mechanisms in a Neonatal HIE Rat Model. Cell Transplant. 2019, 28, 1552–1559. [Google Scholar] [CrossRef] [PubMed]
- Stavely, R.; Nurgali, K. The emerging antioxidant paradigm of mesenchymal stem cell therapy. STEM CELLS Transl. Med. 2020, 9, 985–1006. [Google Scholar] [CrossRef] [PubMed]
- van Buul, G.M.; Siebelt, M.; Leijs, M.J.C.; Bos, P.K.; Waarsing, J.H.; Kops, N.; Weinans, H.; Verhaar, J.A.N.; Bernsen, M.R.; van Osch, G.J.V.M. Mesenchymal stem cells reduce pain but not degenerative changes in a mono-iodoacetate rat model of osteoarthritis. J. Orthop. Res. 2014, 32, 1167–1174. [Google Scholar] [CrossRef] [PubMed]
- Dominici, M.; Le Blanc, K.; Mueller, I.; Slaper-Cortenbach, I.; Marini, F.C.; Krause, D.S.; Deans, R.J.; Keating, A.; Prockop, D.J.; Horwitz, E.M. Minimal criteria for defining multipotent mesenchymal stromal cells. The International Society for Cellular Therapy position statement. Cytotherapy 2006, 8, 315–317. [Google Scholar] [CrossRef] [PubMed]
- Tondreau, T.; Lagneaux, L.; Dejeneffe, M.; Massy, M.; Mortier, C.; Delforge, A.; Bron, D. Bone marrow–derived mesenchymal stem cells already express specific neural proteins before any differentiation. Differentiation 2004, 72, 319–326. [Google Scholar] [CrossRef] [PubMed]
- Cho, S.-W.; Kim, I.-K.; Lim, S.H.; Kim, D.-I.; Kang, S.-W.; Kim, S.H.; Kim, Y.H.; Lee, E.Y.; Choi, C.Y.; Kim, B.-S. Smooth muscle-like tissues engineered with bone marrow stromal cells. Biomaterials 2004, 25, 2979–2986. [Google Scholar] [CrossRef]
- Hung, S.-C.; Chen, N.-J.; Hsieh, S.-L.; Li, H.; Ma, H.-L.; Lo, W.-H. Isolation and Characterization of Size-Sieved Stem Cells from Human Bone Marrow. STEM CELLS 2002, 20, 249–258. [Google Scholar] [CrossRef] [PubMed]
- Hung, S.-C.; Chang, C.-F.; Ma, H.-L.; Chen, T.-H.; Ho, L.L.-T. Gene expression profiles of early adipogenesis in human mesenchymal stem cells. Gene 2004, 340, 141–150. [Google Scholar] [CrossRef] [PubMed]
- George, S.; Hamblin, M.R.; Abrahamse, H. Differentiation of Mesenchymal Stem Cells to Neuroglia: In the Context of Cell Signalling. Stem Cell Rev. Rep. 2019, 15, 814–826. [Google Scholar] [CrossRef] [PubMed]
- Martin, G.R. Isolation of a pluripotent cell line from early mouse embryos cultured in medium conditioned by teratocarcinoma stem cells. Proc. Natl. Acad. Sci. USA 1981, 78, 7634–7638. [Google Scholar] [CrossRef] [PubMed]
- Carpenter, M.K.; Rößler, E.; Rao, M.S. Characterization and Differentiation of Human Embryonic Stem Cells. Cloning Stem Cells 2003, 5, 79–88. [Google Scholar] [CrossRef] [PubMed]
- Hoffman, L.M.; Carpenter, M.K. Characterization and culture of human embryonic stem cells. Nat. Biotechnol. 2005, 23, 699–708. [Google Scholar] [CrossRef] [PubMed]
- Goldring, C.E.; Duffy, P.A.; Benvenisty, N.; Andrews, P.W.; Ben-David, U.; Eakins, R.; French, N.; Hanley, N.A.; Kelly, L.; Kitteringham, N.R.; et al. Assessing the Safety of Stem Cell Therapeutics. Cell Stem Cell 2011, 8, 618–628. [Google Scholar] [CrossRef] [PubMed]
- Ren, G.; Chen, X.; Dong, F.; Li, W.; Ren, X.; Zhang, Y.; Shi, Y. Concise Review: Mesenchymal Stem Cells and Translational Medicine: Emerging Issues. STEM CELLS Transl. Med. 2012, 1, 51–58. [Google Scholar] [CrossRef] [PubMed]
- Prockop, D.J.; Brenner, M.; Fibbe, W.E.; Horwitz, E.; Le Blanc, K.; Phinney, D.G.; Simmons, P.J.; Sensebe, L.; Keating, A. Defining the risks of mesenchymal stromal cell therapy. Cytotherapy 2010, 12, 576–578. [Google Scholar] [CrossRef] [PubMed]
- Handgretinger, R.; Schilbach, K. The potential role of γδ T cells after allogeneic HCT for leukemia. Blood 2018, 131, 1063–1072. [Google Scholar] [CrossRef] [PubMed]
- Hemmingsen, M.; Vedel, S.; Skafte-Pedersen, P.; Sabourin, D.; Collas, P.; Bruus, H.; Dufva, M. The Role of Paracrine and Autocrine Signaling in the Early Phase of Adipogenic Differentiation of Adipose-derived Stem Cells. PLoS ONE 2013, 8, e63638. [Google Scholar] [CrossRef] [PubMed]
- Moghadam, F.H.; Tayebi, T.; Dehghan, M.; Eslami, G.; Nadri, H.; Moradi, A.; Vahedian-Ardakani, H.; Barzegar, K. Differentiation of Bone Marrow Mesenchymal Stem Cells into Chondrocytes after Short Term Culture in Alkaline Medium. Int. J. Hematol. Stem Cell Res. 2014, 8, 12–19. [Google Scholar]
- Wang, J.-J.; Ye, F.; Cheng, L.-J.; Shi, Y.-J.; Bao, J.; Sun, H.-Q.; Wang, W.; Zhang, P.; Bu, H. Osteogenic differentiation of mesenchymal stem cells promoted by overexpression of connective tissue growth factor. J. Zhejiang Univ. Sci. B 2009, 10, 355–367. [Google Scholar] [CrossRef]
- Tropel, P.; Platet, N.; Platel, J.-C.; Noël, D.; Albrieux, M.; Benabid, A.-L.; Berger, F. Functional Neuronal Differentiation of Bone Marrow-Derived Mesenchymal Stem Cells. STEM CELLS 2006, 24, 2868–2876. [Google Scholar] [CrossRef] [PubMed]
- Hass, R.; Kasper, C.; Böhm, S.; Jacobs, R. Different populations and sources of human mesenchymal stem cells (MSC): A comparison of adult and neonatal tissue-derived MSC. Cell Commun. Signal. 2011, 9, 12. [Google Scholar] [CrossRef]
- Camernik, K.; Mihelič, A.; Mihalič, R.; Haring, G.; Herman, S.; Presen, D.M.; Janež, A.; Trebše, R.; Marc, J.; Zupan, J. Comprehensive analysis of skeletal muscle- and bone-derived mesenchymal stem/stromal cells in patients with osteoarthritis and femoral neck fracture. Stem Cell Res. Ther. 2020, 11, 146. [Google Scholar] [CrossRef] [PubMed]
- Čamernik, K.; Mihelič, A.; Mihalič, R.; Presen, D.M.; Janež, A.; Trebše, R.; Marc, J.; Zupan, J. Increased Exhaustion of the Subchondral Bone-Derived Mesenchymal Stem/Stromal Cells in Primary Versus Dysplastic Osteoarthritis. Stem Cell Rev. Rep. 2020, 16, 742–754. [Google Scholar] [CrossRef]
- Zupan, J.; Strazar, K.; Kocijan, R.; Nau, T.; Grillari, J.; Presen, D.M. Age-related alterations and senescence of mesenchymal stromal cells: Implications for regenerative treatments of bones and joints. Mech. Ageing Dev. 2021, 198, 111539. [Google Scholar] [CrossRef]
- Pevsner-Fischer, M.; Levin, S.; Zipori, D. The Origins of Mesenchymal Stromal Cell Heterogeneity. Stem Cell Rev. Rep. 2011, 7, 560–568. [Google Scholar] [CrossRef] [PubMed]
- Russell, A.L.; Lefavor, R.; Durand, N.; Glover, L.; Zubair, A.C. Modifiers of mesenchymal stem cell quantity and quality. Transfusion 2018, 58, 1434–1440. [Google Scholar] [CrossRef] [PubMed]
- Pizzute, T.; Lynch, K.; Pei, M. Impact of Tissue-Specific Stem Cells on Lineage-Specific Differentiation: A Focus on the Musculoskeletal System. Stem Cell Rev. Rep. 2015, 11, 119–132. [Google Scholar] [CrossRef]
- Mattiucci, D.; Maurizi, G.; Leoni, P.; Poloni, A. Aging- and Senescence-associated Changes of Mesenchymal Stromal Cells in Myelodysplastic Syndromes. Cell Transplant. 2018, 27, 754–764. [Google Scholar] [CrossRef]
- Malgieri, A.; Kantzari, E.; Patrizi, M.P.; Gambardella, S. Bone marrow and umbilical cord blood human mesenchymal stem cells: State of the art. Int. J. Clin. Exp. Med. 2010, 3, 248–269. [Google Scholar] [PubMed]
- D’Ippolito, G.; Schiller, P.C.; Ricordi, C.; Roos, B.A.; Howard, G.A. Age-Related Osteogenic Potential of Mesenchymal Stromal Stem Cells from Human Vertebral Bone Marrow. J. Bone Miner. Res. 1999, 14, 1115–1122. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, A.; Sheng, M.H.; Wasnik, S.; Baylink, D.J.; Lau, K.-H.W. Effect of aging on stem cells. World J. Exp. Med. 2017, 7, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Fukada, S.-I.; Ma, Y.; Uezumi, A. Adult stem cell and mesenchymal progenitor theories of aging. Front. Cell Dev. Biol. 2014, 2, 10. [Google Scholar] [CrossRef] [PubMed]
- Babenko, V.A.; Silachev, D.N.; Danilina, T.I.; Goryunov, K.V.; Pevzner, I.B.; Zorova, L.D.; Popkov, V.A.; Chernikov, V.P.; Plotnikov, E.Y.; Sukhikh, G.T.; et al. Age-Related Changes in Bone-Marrow Mesenchymal Stem Cells. Cells 2021, 10, 1273. [Google Scholar] [CrossRef] [PubMed]
- Fehrer, C.; Lepperdinger, G. Mesenchymal stem cell aging. Exp. Gerontol. 2005, 40, 926–930. [Google Scholar] [CrossRef] [PubMed]
- Miao, Z.; Jin, J.; Chen, L.; Zhu, J.; Huang, W.; Zhao, J.; Qian, H.; Zhang, X. Isolation of mesenchymal stem cells from human placenta: Comparison with human bone marrow mesenchymal stem cells. Cell Biol. Int. 2006, 30, 681–687. [Google Scholar] [CrossRef] [PubMed]
- Beeravolu, N.; Khan, I.; McKee, C.; Dinda, S.; Thibodeau, B.; Wilson, G.; Perez-Cruet, M.; Bahado-Singh, R.; Chaudhry, G.R. Isolation and comparative analysis of potential stem/progenitor cells from different regions of human umbilical cord. Stem Cell Res. 2016, 16, 696–711. [Google Scholar] [CrossRef] [PubMed]
- Beeravolu, N.; McKee, C.; Alamri, A.; Mikhael, S.; Brown, C.; Perez-Cruet, M.; Chaudhry, G.R. Isolation and Characterization of Mesenchymal Stromal Cells from Human Umbilical Cord and Fetal Placenta. J. Vis. Exp. 2017, 122, e55224. [Google Scholar] [CrossRef]
- Raileanu, V.N.; Whiteley, J.; Chow, T.; Kollara, A.; Mohamed, A.; Keating, A.; Rogers, I.M. Banking Mesenchymal Stromal Cells from Umbilical Cord Tissue: Large Sample Size Analysis Reveals Consistency Between Donors. STEM CELLS Transl. Med. 2019, 8, 1041–1054. [Google Scholar] [CrossRef]
- Maurice, S.; Srouji, S.; Livne, E. Isolation of progenitor cells from cord blood using adhesion matrices. Cytotechnology 2007, 52, 125–137. [Google Scholar] [CrossRef]
- Gucciardo, L.; Lories, R.; Ochsenbein-Kölble, N.; Done, E.; Zwijsen, A.; Deprest, J.; Ochsenbein-Kölble, N. Fetal mesenchymal stem cells: Isolation, properties and potential use in perinatology and regenerative medicine. BJOG Int. J. Obstet. Gynaecol. 2008, 116, 166–172. [Google Scholar] [CrossRef] [PubMed]
- Wouters, G.; Grossi, S.; Mesoraca, A.; Bizzoco, D.; Mobili, L.; Cignini, P.; Giorlandino, C. Isolation of amniotic fluid-derived mesenchymal stem cells. J. Prenat. Med. 2007, 1, 39–40. [Google Scholar]
- Kwon, A.; Kim, Y.; Kim, M.; Kim, J.; Choi, H.; Jekarl, D.W.; Lee, S.; Kim, J.M.; Shin, J.-C.; Park, I.Y. Tissue-specific Differentiation Potency of Mesenchymal Stromal Cells from Perinatal Tissues. Sci. Rep. 2016, 6, 23544. [Google Scholar] [CrossRef] [PubMed]
- Caplan, A.I.; Correa, D. The MSC: An Injury Drugstore. Cell Stem Cell 2011, 9, 11–15. [Google Scholar] [CrossRef]
- da Silva Meirelles, L.; Caplan, A.I.; Nardi, N.B. In Search of the In Vivo Identity of Mesenchymal Stem Cells. STEM CELLS 2008, 26, 2287–2299. [Google Scholar] [CrossRef]
- da Silva Meirelles, L.; Fontes, A.M.; Covas, D.T.; Caplan, A.I. Mechanisms involved in the therapeutic properties of mesenchymal stem cells. Cytokine Growth Factor Rev. 2009, 20, 419–427. [Google Scholar] [CrossRef] [PubMed]
- Caplan, A.I.; Dennis, J.E. Mesenchymal stem cells as trophic mediators. J. Cell. Biochem. 2006, 98, 1076–1084. [Google Scholar] [CrossRef] [PubMed]
- Le Blanc, K.; Mougiakakos, D. Multipotent mesenchymal stromal cells and the innate immune system. Nat. Rev. Immunol. 2012, 12, 383–396. [Google Scholar] [CrossRef] [PubMed]
- Caplan, A.I. Adult Mesenchymal Stem Cells: When, Where, and How. Stem Cells Int. 2015, 2015, 628767. [Google Scholar] [CrossRef] [PubMed]
- Caplan, A.I. Mesenchymal Stem Cells: Time to Change the Name! Stem Cells Transl. Med. 2017, 6, 1445–1451. [Google Scholar] [CrossRef] [PubMed]
- Horwitz, E.M.; Le Blanc, K.; Dominici, M.; Mueller, I.; Slaper-Cortenbach, I.; Marini, F.C.; Deans, R.J.; Krause, D.S.; Keating, A. International Society for Cellular Therapy Clarification of the nomenclature for MSC: The International Society for Cellular Therapy position statement. Cytotherapy 2005, 7, 393–395. [Google Scholar] [CrossRef] [PubMed]
- Viswanathan, S.; Shi, Y.; Galipeau, J.; Krampera, M.; Leblanc, K.; Martin, I.; Nolta, J.; Phinney, D.G.; Sensebe, L. Mesenchymal stem versus stromal cells: International Society for Cell & Gene Therapy (ISCT®) Mesenchymal Stromal Cell committee position statement on nomenclature. Cytotherapy 2019, 21, 1019–1024. [Google Scholar] [CrossRef] [PubMed]
- Mushahary, D.; Spittler, A.; Kasper, C.; Weber, V.; Charwat, V. Isolation, cultivation, and characterization of human mesenchymal stem cells. Cytom. Part A 2018, 93, 19–31. [Google Scholar] [CrossRef] [PubMed]
- Salehinejad, P.; Alitheen, N.B.; Ali, A.M.; Omar, A.R.; Mohit, M.; Janzamin, E.; Samani, F.S.; Torshizi, Z.; Nematollahi-Mahani, S.N. Comparison of different methods for the isolation of mesenchymal stem cells from human umbilical cord Wharton’s jelly. Vitr. Cell. Dev. Biol.-Anim. 2012, 48, 75–83. [Google Scholar] [CrossRef]
- Yoon, J.H.; Roh, E.Y.; Shin, S.; Jung, N.H.; Song, E.Y.; Chang, J.Y.; Kim, B.J.; Jeon, H.W. Comparison of Explant-Derived and Enzymatic Digestion-Derived MSCs and the Growth Factors from Wharton’s Jelly. BioMed Res. Int. 2013, 2013, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Hendijani, F. Explant culture: An advantageous method for isolation of mesenchymal stem cells from human tissues. Cell Prolif. 2017, 50, e12334. [Google Scholar] [CrossRef] [PubMed]
- Tkach, M.; Théry, C. Communication by Extracellular Vesicles: Where We Are and Where We Need to Go. Cell 2016, 164, 1226–1232. [Google Scholar] [CrossRef]
- Keerthikumar, S.; Chisanga, D.; Ariyaratne, D.; Al Saffar, H.; Anand, S.; Zhao, K.; Samuel, M.; Pathan, M.; Jois, M.; Chilamkurti, N.; et al. ExoCarta: A Web-Based Compendium of Exosomal Cargo. J. Mol. Biol. 2016, 428, 688–692. [Google Scholar] [CrossRef] [PubMed]
- EL Andaloussi, S.; Mäger, I.; Breakefield, X.O.; Wood, M.J.A. Extracellular vesicles: Biology and emerging therapeutic opportunities. Nat. Rev. Drug Discov. 2013, 12, 347–357. [Google Scholar] [CrossRef] [PubMed]
- Cosenza, S.; Toupet, K.; Maumus, M.; Luz-Crawford, P.; Blanc-Brude, O.; Jorgensen, C.; Noël, D. Mesenchymal stem cells-derived exosomes are more immunosuppressive than microparticles in inflammatory arthritis. Theranostics 2018, 8, 1399–1410. [Google Scholar] [CrossRef]
- Bang, O.Y.; Kim, E.H. Mesenchymal Stem Cell-Derived Extracellular Vesicle Therapy for Stroke: Challenges and Progress. Front. Neurol. 2019, 10, 211. [Google Scholar] [CrossRef] [PubMed]
- Wiklander, O.P.B.; Nordin, J.Z.; O’Loughlin, A.; Gustafsson, Y.; Corso, G.; Mäger, I.; Vader, P.; Lee, Y.; Sork, H.; Seow, Y.; et al. Extracellular vesicle in vivo biodistribution is determined by cell source, route of administration and targeting. J. Extracell. Vesicles 2015, 4, 26316. [Google Scholar] [CrossRef] [PubMed]
- Busato, A.; Bonafede, R.; Bontempi, P.; Scambi, I.; Schiaffino, L.; Benati, D.; Malatesta, M.; Sbarbati, A.; Marzola, P.; Mariotti, R. Labeling and Magnetic Resonance Imaging of Exosomes Isolated from Adipose Stem Cells. Curr. Protoc. Cell Biol. 2017, 75, 3–44. [Google Scholar] [CrossRef] [PubMed]
- da Silva Meirelles, L.; Malta, T.M.; de Deus Wagatsuma, V.M.; Palma, P.V.B.; Araújo, A.G.; Ribeiro Malmegrim, K.C.; Morato De Oliveira, F.; Panepucci, R.A.; Silva, W.A.; Kashima Haddad, S.; et al. Cultured Human Adipose Tissue Pericytes and Mesenchymal Stromal Cells Display a Very Similar Gene Expression Profile. Stem Cells Dev. 2015, 24, 2822–2840. [Google Scholar] [CrossRef] [PubMed]
- da Silva Meirelles, L.; de Deus Wagatsuma, V.M.; Malta, T.; Palma, P.V.B.; Araújo, A.G.; Panepucci, R.A.; Silva, W.A.; Kashima, S.; Covas, D. The gene expression profile of non-cultured, highly purified human adipose tissue pericytes: Transcriptomic evidence that pericytes are stem cells in human adipose tissue. Exp. Cell Res. 2016, 349, 239–254. [Google Scholar] [CrossRef]
- Tang, Y.; Wu, X.; Lei, W.; Pang, L.; Wan, C.; Shi, Z.; Zhao, L.; Nagy, T.R.; Peng, X.; Hu, J.; et al. TGF-beta1-induced migration of bone mesenchymal stem cells couples bone resorption with formation. Nat. Med. 2009, 15, 757–765. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z. Bone regeneration by stem cell and tissue engineering in oral and maxillofacial region. Front. Med. 2011, 5, 401–413. [Google Scholar] [CrossRef] [PubMed]
- Gómez, S.; Vlad, M.D.; López, J.; Fernández, E. Design and properties of 3D scaffolds for bone tissue engineering. Acta Biomater. 2016, 42, 341–350. [Google Scholar] [CrossRef] [PubMed]
- Paspaliaris, V.; Kolios, G. Stem cells in Osteoporosis: From Biology to New Therapeutic Approaches. Stem Cells Int. 2019, 2019, 1–16. [Google Scholar] [CrossRef]
- De Witte, T.-M.; Fratila-Apachitei, L.E.; Zadpoor, A.A.; Peppas, N.A. Bone tissue engineering via growth factor delivery: From scaffolds to complex matrices. Regen. Biomater. 2018, 5, 197–211. [Google Scholar] [CrossRef] [PubMed]
- Vadalà, G.; Ambrosio, L.; Russo, F.; Papalia, R.; Denaro, V. Interaction between Mesenchymal Stem Cells and Intervertebral Disc Microenvironment: From Cell Therapy to Tissue Engineering. Stem Cells Int. 2019, 2019, 1–15. [Google Scholar] [CrossRef]
- Estes, B.T.; Diekman, B.O.; Gimble, J.M.; Guilak, F. Isolation of adipose-derived stem cells and their induction to a chondrogenic phenotype. Nat. Protoc. 2010, 5, 1294–1311. [Google Scholar] [CrossRef] [PubMed]
- Orbay, H.; Devi, K.; Williams, P.A.; Dehghani, T.; Silva, E.A.; Sahar, D.E. Comparison of Endothelial Differentiation Capacities of Human and Rat Adipose-Derived Stem Cells. Plast. Reconstr. Surg. 2016, 138, 1231–1241. [Google Scholar] [CrossRef] [PubMed]
- Xu, T.; Yu, X.; Yang, Q.; Liu, X.; Fang, J.; Dai, X. Autologous Micro-Fragmented Adipose Tissue as Stem Cell-Based Natural Scaffold for Cartilage Defect Repair. Cell Transplant. 2019, 28, 1709–1720. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.-L.; Liu, W.; Wu, Y.-M.; Sun, W.-L.; Dörfer, C.E.; El-Sayed, K.M.F. Oral Mesenchymal Stem/Progenitor Cells: The Immunomodulatory Masters. Stem Cells Int. 2020, 2020, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Gan, L.; Liu, Y.; Cui, D.; Pan, Y.; Zheng, L.; Wan, M. Dental Tissue-Derived Human Mesenchymal Stem Cells and Their Potential in Therapeutic Application. Stem Cells Int. 2020, 2020, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Yang, R.; Wang, J.; Chen, X.; Shi, Y.; Xie, J. Epidermal Stem Cells in Wound Healing and Regeneration. Stem Cells Int. 2020, 2020, 1–11. [Google Scholar] [CrossRef]
- Lee, D.E.; Ayoub, N.; Agrawal, D.K. Mesenchymal stem cells and cutaneous wound healing: Novel methods to increase cell delivery and therapeutic efficacy. Stem Cell Res. Ther. 2016, 7, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Jeon, Y.K.; Jang, Y.H.; Yoo, D.R.; Na Kim, S.; Lee, S.K.; Nam, M.J. Mesenchymal stem cells’ interaction with skin: Wound-healing effect on fibroblast cells and skin tissue. Wound Repair Regen. 2010, 18, 655–661. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.H.; Jin, S.Y.; Song, J.S.; Seo, K.K.; Cho, K.H. Paracrine effects of adipose-derived stem cells on keratinocytes and dermal fibroblasts. Ann. Dermatol. 2012, 24, 136–143. [Google Scholar] [CrossRef]
- Hsieh, J.-Y.; Wang, H.-W.; Chang, S.-J.; Liao, K.-H.; Lee, I.-H.; Lin, W.-S.; Wu, C.-H.; Lin, W.-Y.; Cheng, S.-M. Mesenchymal Stem Cells from Human Umbilical Cord Express Preferentially Secreted Factors Related to Neuroprotection, Neurogenesis, and Angiogenesis. PLoS ONE 2013, 8, e72604. [Google Scholar] [CrossRef]
- I Arno, A.; Amini-Nik, S.; Blit, P.H.; Al-Shehab, M.; Belo, C.; Herer, E.; Tien, C.H.; Jeschke, M.G. Human Wharton’s jelly mesenchymal stem cells promote skin wound healing through paracrine signaling. Stem Cell Res. Ther. 2014, 5, 28. [Google Scholar] [CrossRef]
- Zhao, Y.-X.; Chen, S.-R.; Su, P.-P.; Huang, F.-H.; Shi, Y.-C.; Shi, Q.-Y.; Lin, S. Using Mesenchymal Stem Cells to Treat Female Infertility: An Update on Female Reproductive Diseases. Stem Cells Int. 2019, 2019, 1–10. [Google Scholar] [CrossRef]
- Kossowska-Tomaszczuk, K.; Geyter, C. De Cells with Stem Cell Characteristics in Somatic Compartments of the Ovary. Biomed Res. Int. 2013, 2013, 310859. [Google Scholar] [CrossRef]
- Hoang, S.N.; Ho, Q.C.; Nguyen, T.T.P.; Doan, C.C.; Tran, D.H.; Le, L.T. Evaluation of stemness marker expression in bovine ovarian granulosa cells. Anim. Reprod. 2019, 16, 277–281. [Google Scholar] [CrossRef] [PubMed]
- Varras, M.; Griva, T.; Kalles, V.; Akrivis, C.; Paparisteidis, N. Markers of stem cells in human ovarian granulosa cells: Is there a clinical significance in ART? J. Ovarian Res. 2012, 5, 36. [Google Scholar] [CrossRef] [PubMed]
- Kossowska-Tomaszczuk, K.; Pelczar, P.; Güven, S.; Kowalski, J.; Volpi, E.; De Geyter, C.; Scherberich, A. A Novel Three-Dimensional Culture System Allows Prolonged Culture of Functional Human Granulosa Cells and Mimics the Ovarian Environment. Tissue Eng. Part A 2010, 16, 2063–2073. [Google Scholar] [CrossRef] [PubMed]
- Kossowska-Tomaszczuk, K.; De Geyter, C.; De Geyter, M.; Martin, I.; Holzgreve, W.; Scherberich, A.; Zhang, H. The Multipotency of Luteinizing Granulosa Cells Collected from Mature Ovarian Follicles. Stem Cells 2009, 27, 210–219. [Google Scholar] [CrossRef] [PubMed]
- Brevini, T.A.L.; Pennarossa, G.; Rahman, M.M.; Paffoni, A.; Antonini, S.; Ragni, G.; Deeguileor, M.; Tettamanti, G.; Gandolfi, F. Morphological and Molecular Changes of Human Granulosa Cells Exposed to 5-Azacytidine and Addressed Toward Muscular Differentiation. Stem Cell Rev. Rep. 2014, 10, 633–642. [Google Scholar] [CrossRef] [PubMed]
- Brązert, M.; Kranc, W.; Celichowski, P.; Jankowski, M.; Piotrowska-Kempisty, H.; Pawelczyk, L.; Bruska, M.; Zabel, M.; Nowicki, M.; Kempisty, B. Expression of genes involved in neurogenesis, and neuronal precursor cell proliferation and development: Novel pathways of human ovarian granulosa cell differentiation and transdifferentiation capability in vitro. Mol. Med. Rep. 2020, 21, 1749–1760. [Google Scholar] [CrossRef] [PubMed]
- Virant-Klun, I.; Omejec, S.; Stimpfel, M.; Skerl, P.; Novakovic, S.; Jancar, N.; Vrtacnik-Bokal, E. Female Age Affects the Mesenchymal Stem Cell Characteristics of Aspirated Follicular Cells in the In Vitro Fertilization Programme. Stem Cell Rev. Rep. 2019, 15, 543–557. [Google Scholar] [CrossRef] [PubMed]
- Guillot, P.V.; Gotherstrom, C.; Chan, J.; Kurata, H.; Fisk, N.M. Human First-Trimester Fetal MSC Express Pluripotency Markers and Grow Faster and Have Longer Telomeres Than Adult MSC. Stem Cells 2007, 25, 646–654. [Google Scholar] [CrossRef] [PubMed]
- Blum, B.; Benvenisty, N. The Tumorigenicity of Human Embryonic Stem Cells. Adv. Cancer Res. 2008, 100, 133–158. [Google Scholar] [CrossRef] [PubMed]
- Fong, C.-Y.; Gauthaman, K.; Bongso, A. Teratomas from pluripotent stem cells: A clinical hurdle. J. Cell. Biochem. 2010, 111, 769–781. [Google Scholar] [CrossRef] [PubMed]
- Götherström, C.; Ringdén, O.; Tammik, C.; Zetterberg, E.; Westgren, M.; Le Blanc, K. Immunologic properties of human fetal mesenchymal stem cells. Am. J. Obstet. Gynecol. 2004, 190, 239–245. [Google Scholar] [CrossRef]
- Götherström, C.; West, A.; Liden, J.; Uzunel, M.; Lahesmaa, R.; Le Blanc, K. Difference in gene expression between human fetal liver and adult bone marrow mesenchymal stem cells. Haematologica 2005, 90, 1017–1026. [Google Scholar] [PubMed]
- Mirmalek-Sani, S.-H.; Tare, R.S.; Morgan, S.M.; Roach, H.I.; Wilson, D.I.; Hanley, N.A.; Oreffo, R.O. Characterization and Multipotentiality of Human Fetal Femur-Derived Cells: Implications for Skeletal Tissue Regeneration. Stem Cells 2006, 24, 1042–1053. [Google Scholar] [CrossRef]
- Guillot, P.V.; De Bari, C.; Dell’Accio, F.; Kurata, H.; Polak, J.; Fisk, N.M. Comparative osteogenic transcription profiling of various fetal and adult mesenchymal stem cell sources. Differ. 2008, 76, 946–957. [Google Scholar] [CrossRef] [PubMed]
- O’Donoghue, K.; Fisk, N.M. Fetal stem cells. Best Pract. Res. Clin. Obstet. Gynaecol. 2004, 18, 853–875. [Google Scholar]
- Brumbaugh, J.; Rose, C.; Phanstiel, D.H.; Thomson, J.A.; Coon, J.J. Proteomics and pluripotency. Crit. Rev. Biochem. Mol. Biol. 2011, 46, 493–506. [Google Scholar] [CrossRef]
- Radzisheuskaya, A.; Le Bin Chia, G.; dos Santos, R.L.; Theunissen, T.; Castro, L.F.; Nichols, J.; Silva, J.C.R. A defined Oct4 level governs cell state transitions of pluripotency entry and differentiation into all embryonic lineages. Nat. Cell Biol. 2013, 15, 579–590. [Google Scholar] [CrossRef]
- Pan, G.J.; Chang, Z.Y.; Schöler, H.R.; Pei, D. Stem cell pluripotency and transcription factor Oct4. Cell Res. 2002, 12, 321–329. [Google Scholar] [CrossRef] [PubMed]
- Seruya, M.; Shah, A.; Pedrotty, D.; Du Laney, T.; Melgiri, R.; McKee, J.A.; Young, H.E.; Niklason, L.E. Clonal Population of Adult Stem Cells: Life Span and Differentiation Potential. Cell Transplant. 2004, 13, 93–101. [Google Scholar] [CrossRef]
- Xue, G.; Han, X.; Ma, X.; Wu, H.; Qin, Y.; Liu, J.; Hu, Y.; Hong, Y.; Hou, Y. Effect of Microenvironment on Differentiation of Human Umbilical Cord Mesenchymal Stem Cells into Hepatocytes In Vitro and In Vivo. Biomed Res. Int. 2016, 2016, 1–13. [Google Scholar]
- Kusuma, G.D.; Carthew, J.R.; Lim, R.; Frith, J.E. Effect of the Microenvironment on Mesenchymal Stem Cell Paracrine Signaling: Opportunities to Engineer the Therapeutic Effect. Stem Cells Dev. 2017, 26, 617–631. [Google Scholar] [CrossRef] [PubMed]
- Robert, A.W.; Marcon, B.H.; Dallagiovanna, B.; Shigunov, P. Adipogenesis, Osteogenesis, and Chondrogenesis of Human Mesenchymal Stem/Stromal Cells: A Comparative Transcriptome Approach. Front. Cell Dev. Biol. 2020, 8, 561. [Google Scholar] [CrossRef] [PubMed]
- Munir, H.; Ward, L.; Sheriff, L.; Kemble, S.; Nayar, S.; Barone, F.; Nash, G.B.; McGettrick, H.M. Adipogenic Differentiation of Mesenchymal Stem Cells Alters Their Immunomodulatory Properties in a Tissue-Specific Manner. Stem Cells 2017, 35, 1636–1646. [Google Scholar] [CrossRef]
- José, B.; Andrades, J.A.; Ertl, D.C.; Sorgente, N.; Nimni, M.E. Demineralized bone matrix mediates differentiation of bone marrow stromal cells in vitro: Effect of age of cell donor. J. Bone Miner. Res. 2009, 11, 1703–1714. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.; Li, P.; Xu, H.; Pan, Q.; Zeng, R.; Ma, X.; Li, Z.; Lin, H. Dexamethasone-Activated MSCs Release MVs for Stimulating Osteogenic Response. Stem Cells Int. 2018, 2018, 1–12. [Google Scholar] [CrossRef]
- Huang, C.-Y.C.; Hagar, K.L.; Frost, L.E.; Sun, Y.; Cheung, H.S. Effects of Cyclic Compressive Loading on Chondrogenesis of Rabbit Bone-Marrow Derived Mesenchymal Stem Cells. Stem Cells 2004, 22, 313–323. [Google Scholar] [CrossRef] [PubMed]
- Tontonoz, P.; Hu, E.; Spiegelman, B.M. Stimulation of adipogenesis in fibroblasts by PPARγ2, a lipid-activated transcription factor. Cell 1994, 79, 1147–1156. [Google Scholar] [CrossRef]
- Rangwala, S.M.; Lazar, M.A. Transcriptional Control of Adipogenesis. Annu. Rev. Nutr. 2000, 20, 535–559. [Google Scholar] [CrossRef]
- Furumatsu, T.; Tsuda, M.; Taniguchi, N.; Tajima, Y.; Asahara, H. Smad3 Induces Chondrogenesis through the Activation of SOX9 via CREB-binding Protein/p300 Recruitment. J. Biol. Chem. 2005, 280, 8343–8350. [Google Scholar] [CrossRef]
- Chu, M.-S.; Chang, C.-F.; Yang, C.-C.; Bau, Y.-C.; Ho, L.L.-T.; Hung, S.-C. Signalling pathway in the induction of neurite outgrowth in human mesenchymal stem cells. Cell. Signal. 2006, 18, 519–530. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Tio, M.; Lee, W.; Beerheide, W.; Udolph, G. Neural differentiation of mesenchymal-like stem cells from cord blood is mediated by PKA. Biochem. Biophys. Res. Commun. 2007, 357, 1021–1027. [Google Scholar] [CrossRef]
- Yanjie, J.; Jiping, S.; Yan, Z.; Xiaofeng, Z.; Boai, Z.; Yajun, L. Effects of Notch-1 signalling pathway on differentiation of marrow mesenchymal stem cells into neurons in vitro. Neuro Report 2007, 18, 1443–1447. [Google Scholar] [CrossRef]
- Qian, L.; Saltzman, W. Improving the expansion and neuronal differentiation of mesenchymal stem cells through culture surface modification. Biomater. 2004, 25, 1331–1337. [Google Scholar] [CrossRef] [PubMed]
- D’Ippolito, G.; Diabira, S.; Howard, G.A.; Menei, P.; Roos, B.A.; Schiller, P.C. Marrow-isolated adult multilineage inducible (MIAMI) cells, a unique population of postnatal young and old human cells with extensive expansion and differentiation potential. J. Cell Sci. 2004, 117, 2971–2981. [Google Scholar] [CrossRef] [PubMed]
- Ding, D.-C.; Shyu, W.-C.; Chiang, M.-F.; Lin, S.-Z.; Chang, Y.-C.; Wang, H.-J.; Su, C.-Y.; Li, H. Enhancement of neuroplasticity through upregulation of β1-integrin in human umbilical cord-derived stromal cell implanted stroke model. Neurobiol. Dis. 2007, 27, 339–353. [Google Scholar] [CrossRef] [PubMed]
- Inada, M.; Follenzi, A.; Cheng, K.; Surana, M.; Joseph, B.; Benten, D.; Bandi, S.; Qian, H.; Gupta, S. Phenotype reversion in fetal human liver epithelial cells identifies the role of an intermediate meso-endodermal stage before hepatic maturation. J. Cell Sci. 2008, 121, 1002–1013. [Google Scholar] [CrossRef]
- Yoshida, C.A.; Komori, H.; Maruyama, Z.; Miyazaki, T.; Kawasaki, K.; Furuichi, T.; Fukuyama, R.; Mori, M.; Yamana, K.; Nakamura, K.; et al. SP7 Inhibits Osteoblast Differentiation at a Late Stage in Mice. PLoS ONE 2012, 7, e32364. [Google Scholar] [CrossRef]
- Hagh, M.F.; Noruzinia, M.; Mortazavi, Y.; Soleimani, M.; Kaviani, S.; Abroun, S.; Fard, A.D.; Maymand, M.M. Different Methylation Patterns of RUNX2, OSX, DLX5 and BSP in Osteoblastic Differentiation of Mesenchymal Stem Cells. Cell J 2017, 17, 71–82. [Google Scholar]
- Leucht, P.; Lee, S.; Yim, N. Wnt signaling and bone regeneration: Can’t have one without the other. Biomaterials 2019, 196, 46–50. [Google Scholar] [CrossRef]
- Celil, A.B.; Campbell, P.G. BMP-2 and Insulin-like Growth Factor-I Mediate Osterix (Osx) Expression in Human Mesenchymal Stem Cells via the MAPK and Protein Kinase D Signaling Pathways. J. Biol. Chem. 2005, 280, 31353–31359. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Shen, X.; Wan, C.; Zhao, Q.; Zhang, L.; Zhou, Q.; Deng, L. Effects of insulin and insulin-like growth factor 1 on osteoblast proliferation and differentiation: Differential signalling via Akt and ERK. Cell Biochem. Funct. 2012, 30, 297–302. [Google Scholar] [CrossRef] [PubMed]
- Liang, H.; Xu, X.; Feng, X.; Ma, L.; Deng, X.; Wu, S.; Liu, X.; Yang, C. Gold nanoparticles-loaded hydroxyapatite composites guide osteogenic differentiation of human mesenchymal stem cells through Wnt/β-catenin signaling pathway. Int. J. Nanomed. 2019, 2019, 6151–6163. [Google Scholar] [CrossRef] [PubMed]
- Brady, K.; Dickinson, S.C.; Guillot, P.V.; Polak, J.; Blom, A.W.; Kafienah, W.; Hollander, A.P. Human Fetal and Adult Bone Marrow-Derived Mesenchymal Stem Cells Use Different Signaling Pathways for the Initiation of Chondrogenesis. Stem Cells Dev. 2014, 23, 541–554. [Google Scholar] [CrossRef] [PubMed]
- Sibiak, R.; Jaworski, M.; Dorna, Z.; Pieńkowski, W.; Stefańska, K.; Bryl, R.; Žáková, J.; Crha, I.; Ventruba, P.; Ješeta, M.; et al. Human placenta-derived stem cells-recent findings based on the molecular science. Med J. Cell Biol. 2020, 8, 164–169. [Google Scholar] [CrossRef]
- Zhao, Q.; Han, Z.; Wang, J.; Han, Z. Development and investigational new drug application of mesenchymal stem/stromal cells products in China. Stem Cells Transl. Med. 2021, 10, S18–S30. [Google Scholar] [CrossRef]
- Cellular & Gene Therapy Products. Available online: https://www.fda.gov/vaccines-blood-biologics/cellular-gene-therapy-products (accessed on 11 November 2021).
- Blasimme, A.; Rial-Sebbag, E. Regulation of Cell-Based Therapies in Europe: Current Challenges and Emerging Issues. Stem Cells Dev. 2013, 22, 14–19. [Google Scholar] [CrossRef] [PubMed]
- Regulation of Stem Cell Research in Europe. Available online: https://www.eurostemcell.org/pl/regulation-stem-cell-research-europe (accessed on 11 November 2021).
- Silini, A.R.; Masserdotti, A.; Papait, A.; Parolini, O. Shaping the Future of Perinatal Cells: Lessons From the Past and Interpretations of the Present. Front. Bioeng. Biotechnol. 2019, 7, 75. [Google Scholar] [CrossRef] [PubMed]
- Müller, M.; Czarnecka, J.; Brzeziński, M.; Prus, J.; Kulak, B.; Hołubowski, A.; Stasiak, M.; Borowiec, B.; Bryl, R.; Moncrieff, L.; et al. Current stem cells technologies used in medicine. Med J. Cell Biol. 2021, 8, 124–138. [Google Scholar] [CrossRef]
- Erices, A.; Conget, P.; Minguell, J.J. Mesenchymal progenitor cells in human umbilical cord blood. Br. J. Haematol. 2000, 109, 235–242. [Google Scholar] [CrossRef] [PubMed]
- Moschidou, D.; Drews, K.; Eddaoudi, A.; Adjaye, J.; De Coppi, P.; Guillot, P.V. Molecular Signature of Human amniotic Fluid Stem Cells During Fetal Development. Curr. Stem Cell Res. Ther. 2013, 8, 73–81. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Koh, S.-E.; Hur, C.Y.; Lee, W.-D.; Lim, J.; Lee, Y.-J. Comparison of human first and third trimester placental mesenchymal stem cell. Cell Biol. Int. 2013, 37, 242–249. [Google Scholar] [CrossRef]
- Izumi, M.; Pazin, B.J.; Minervini, C.F.; Gerlach, J.; Ross, M.A.; Stolz, D.B.; Turner, M.E.; Thompson, R.L.; Miki, T. Quantitative comparison of stem cell marker-positive cells in fetal and term human amnion. J. Reprod. Immunol. 2009, 81, 39–43. [Google Scholar] [CrossRef]
- Savickiene, J.; Treigyte, G.; Baronaite, S.; Valiuliene, G.; Kaupinis, A.; Valius, M.; Arlauskiene, A.; Navakauskienė, R. Human Amniotic Fluid Mesenchymal Stem Cells from Second- and Third-Trimester Amniocentesis: Differentiation Potential, Molecular Signature, and Proteome Analysis. Stem Cells Int. 2015, 2015, 319238. [Google Scholar] [CrossRef] [PubMed]
- Alessio, N.; Pipino, C.; Mandatori, D.; Di Tomo, P.; Ferone, A.; Marchiso, M.; Melone, M.A.; Peluso, G.; Pandolfi, A.; Galderisi, U. Mesenchymal stromal cells from amniotic fluid are less prone to senescence compared to those obtained from bone marrow: An in vitro study. J. Cell. Physiol. 2018, 233, 8996–9006. [Google Scholar] [CrossRef]
- Galderisi, U.; Peluso, G.; Di Bernardo, G. Clinical Trials Based on Mesenchymal Stromal Cells are Exponentially Increasing: Where are We in Recent Years? Stem Cell Rev. Rep. 2021, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Squillaro, T.; Peluso, G.; Galderisi, U. Clinical Trials with Mesenchymal Stem Cells: An Update. Cell Transplant. 2016, 25, 829–848. [Google Scholar] [CrossRef]
- Silini, A.R.; Di Pietro, R.; Lang-Olip, I.; Alviano, F.; Banerjee, A.; Basile, M.; Borutinskaite, V.; Eissner, G.; Gellhaus, A.; Giebel, B.; et al. Perinatal Derivatives: Where Do We Stand? A Roadmap of the Human Placenta and Consensus for Tissue and Cell Nomenclature. Front. Bioeng. Biotechnol. 2020, 8, 174. [Google Scholar] [CrossRef] [PubMed]
- Bailo, M.; Soncini, M.; Vertua, E.; Signoroni, P.B.; Sanzone, S.; Lombardi, G.; Arienti, D.; Calamani, F.; Zatti, D.; Paul, P.; et al. Engraftment Potential of Human Amnion and Chorion Cells Derived from Term Placenta. Transplantation 2004, 78, 1439–1448. [Google Scholar] [CrossRef] [PubMed]
- Bartholomew, S.; Owens, S.D.; Ferraro, G.L.; Carrade, D.D.; Lara, D.J.; Librach, F.A.; Borjesson, D.L.; Galuppo, L.D. Collection of equine cord blood and placental tissues in 40 thoroughbred mares. Equine Veter-J. 2009, 41, 724–728. [Google Scholar] [CrossRef]
- Carrade, D.D.; Owens, S.D.; Galuppo, L.D.; Vidal, M.A.; Ferraro, G.L.; Librach, F.; Buerchler, S.; Friedman, M.S.; Walker, N.J.; Borjesson, D.L. Clinicopathologic findings following intra-articular injection of autologous and allogeneic placentally derived equine mesenchymal stem cells in horses. Cytotherapy 2011, 13, 419–430. [Google Scholar] [CrossRef] [PubMed]
- Toupadakis, C.A.; Wong, A.; Genetos, D.C.; Cheung, W.K.; Borjesson, D.L.; Ferraro, G.L.; Galuppo, L.D.; Leach, J.K.; Owens, S.D.; Yellowley, C.E. Comparison of the osteogenic potential of equine mesenchymal stem cells from bone marrow, adipose tissue, umbilical cord blood, and umbilical cord tissue. Am. J. Veter-Res. 2010, 71, 1237–1245. [Google Scholar] [CrossRef] [PubMed]
- Schuh, E.M.; Friedman, M.S.; Carrade, D.D.; Li, J.; Heeke, D.; Oyserman, S.M.; Galuppo, L.D.; Lara, D.J.; Walker, N.J.; Ferraro, G.L.; et al. Identification of variables that optimize isolation and culture of multipotent mesenchymal stem cells from equine umbilical-cord blood. Am. J. Veter-Res. 2009, 70, 1526–1535. [Google Scholar] [CrossRef] [PubMed]
- Steven Shaw, S.W.; Bollini, S.; Nader, K.A.; Gastadello, A.; Mehta, V.; Filppi, E.; Cananzi, M.; Gaspar, H.B.; Qasim, W.; de Coppi, P.; et al. Autologous transplantation of amniotic fluid-derived mesenchymal stem cells into sheep fetuses. Cell Transplant. 2011, 20, 1015–1031. [Google Scholar] [CrossRef] [PubMed]
- Colosimo, A.; Curini, V.; Russo, V.; Mauro, A.; Bernabò, N.; Marchisio, M.; Alfonsi, M.; Muttini, A.; Mattioli, M.; Barboni, B. Characterization, GFP Gene Nucleofection, and Allotransplantation in Injured Tendons of Ovine Amniotic Fluid-Derived Stem Cells. Cell Transplant. 2013, 22, 99–117. [Google Scholar] [CrossRef]
- Ribitsch, I.; Chang-Rodriguez, S.; Egerbacher, M.; Gabner, S.; Gueltekin, S.; Huber, J.; Schuster, T.; Jenner, F. Sheep Placenta Cotyledons: A Noninvasive Source of Ovine Mesenchymal Stem Cells. Tissue Eng. Part C Methods 2017, 23, 298–310. [Google Scholar] [CrossRef]
- Zhan, X.-S.; El-Ashram, S.; Luo, D.-Z.; Luo, H.-N.; Wang, B.-Y.; Chen, S.-F.; Bai, Y.-S.; Chen, Z.-S.; Liu, C.-Y.; Ji, H.-Q. A Comparative Study of Biological Characteristics and Transcriptome Profiles of Mesenchymal Stem Cells from Different Canine Tissues. Int. J. Mol. Sci. 2019, 20, 1485. [Google Scholar] [CrossRef]
- Borghesi, J.; Lima, M.F.; Mario, L.C.; Anunciação, A.R.D.A.D.; Rabelo, A.C.S.; da Silva, M.G.K.C.; Fernandes, F.A.; Miglino, M.A.; Carreira, A.C.O.; Favaron, P.O. Canine amniotic membrane mesenchymal stromal/stem cells: Isolation, characterization and differentiation. Tissue Cell 2019, 58, 99–106. [Google Scholar] [CrossRef] [PubMed]
- Saulnier, N.; Loriau, J.; Febre, M.; Robert, C.; Rakic, R.; Bonte, T.; Buff, S.; Maddens, S. Canine placenta: A promising potential source of highly proliferative and immunomodulatory mesenchymal stromal cells? Veter-Immunol. Immunopathol. 2016, 171, 47–55. [Google Scholar] [CrossRef] [PubMed]
- Long, C.; Lankford, L.; Kumar, P.; Grahn, R.; Borjesson, D.L.; Farmer, D.; Wang, A. Isolation and characterization of canine placenta-derived mesenchymal stromal cells for the treatment of neurological disorders in dogs. Cytom. Part A 2018, 93, 82–92. [Google Scholar] [CrossRef] [PubMed]
- Parolini, O.; Alviano, F.; Bagnara, G.P.; Bilic, G.; Bühring, H.-J.; Evangelista, M.; Hennerbichler, S.; Liu, B.; Magatti, M.; Mao, N.; et al. Concise Review: Isolation and Characterization of Cells from Human Term Placenta: Outcome of the First International Workshop on Placenta Derived Stem Cells. Stem Cells 2008, 26, 300–311. [Google Scholar] [CrossRef] [PubMed]
- Roselli, E.A.; Lazzati, S.; Iseppon, F.; Manganini, M.; Marcato, L.; Gariboldi, M.B.; Maggi, F.; Grati, F.R.; Simoni, G. Fetal Mesenchymal Stromal Cells From Cryopreserved Human Chorionicvilli: Cytogenetic and Molecular Analysis of Genome Stability in Long-Term Cultures. Cytotherapy 2013, 15, 1340–1351. [Google Scholar] [CrossRef]
- Abumaree, M.H.; Al Jumah, M.A.; Kalionis, B.; Jawdat, D.; Al Khaldi, A.; Altalabani, A.A.; Knawy, B.A. Phenotypic and Functional Characterization of Mesenchymal Stem Cells from Chorionic Villi of Human Term Placenta. Stem Cell Rev. Rep. 2013, 9, 16–31. [Google Scholar] [CrossRef]
- Igura, K.; Zhang, X.; Takahashi, K.; Mitsuru, A.; Yamaguchi, S.; Takahashi, T.A. Isolation and characterization of mesenchymal progenitor cells from chorionic villi of human placenta. Cytotherapy 2004, 6, 543–553. [Google Scholar] [CrossRef] [PubMed]
- Castrechini, N.M.; Murthi, P.; Gude, N.M.; Erwich, J.; Gronthos, S.; Zannettino, A.; Brennecke, S.; Kalionis, B. Mesenchymal stem cells in human placental chorionic villi reside in a vascular Niche. Placenta 2010, 31, 203–212. [Google Scholar] [CrossRef]
- Macias, M.I.; Grande, J.; Moreno, A.; Domínguez, I.; Bornstein, R.; Flores, A.I. Isolation and characterization of true mesenchymal stem cells derived from human term decidua capable of multilineage differentiation into all 3 embryonic layers. Am. J. Obstet. Gynecol. 2010, 203, 495.e9–495.e23. [Google Scholar] [CrossRef]
- Abomaray, F.M.; Al Jumah, M.A.; Alsaad, K.O.; Jawdat, D.; Al Khaldi, A.; Alaskar, A.S.; Al Harthy, S.; Al Subayyil, A.M.; Khatlani, T.; Alawad, A.O.; et al. Phenotypic and Functional Characterization of Mesenchymal Stem/Multipotent Stromal Cells from Decidua Basalis of Human Term Placenta. Stem Cells Int. 2016, 23(9), 1193–1207. [Google Scholar] [CrossRef]
- Portmann-Lanz, C.B.; Schoeberlein, A.; Huber, A.; Sager, R.; Malek, A.; Holzgreve, W.; Surbek, D.V. Placental mesenchymal stem cells as potential autologous graft for pre- and perinatal neuroregeneration. Am. J. Obstet. Gynecol. 2006, 194(3), 664–673. [Google Scholar] [CrossRef]
- Sudo, K.; Kanno, M.; Miharada, K.; Ogawa, S.; Hiroyama, T.; Saijo, K.; Nakamura, Y. Mesenchymal Progenitors Able to Differentiate into Osteogenic, Chondrogenic, and/or Adipogenic Cells In Vitro Are Present in Most Primary Fibroblast-Like Cell Populations. Stem Cells 2007, 25, 1610–1617. [Google Scholar] [CrossRef] [PubMed]
- Wolbank, S.; Peterbauer, A.; Fahrner, M.; Hennerbichler, S.; van Griensven, M.; Stadler, G.; Redl, H.; Gabriel, C. Dose-Dependent Immunomodulatory Effect of Human Stem Cells from Amniotic Membrane: A Comparison with Human Mesenchymal Stem Cells from Adipose Tissue. Tissue Eng. 2007, 13, 1173–1183. [Google Scholar] [CrossRef]
- Lindenmair, A.; Hatlapatka, T.; Kollwig, G.; Hennerbichler, S.; Gabriel, C.; Wolbank, S.; Redl, H.; Kasper, C. Mesenchymal Stem or Stromal Cells from Amnion and Umbilical Cord Tissue and Their Potential for Clinical Applications. Cells 2012, 1, 1061–1088. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.W.; Koo, H.C.; Hwang, S.Y.; Kang, S.K.; Ra, J.C.; Lee, M.H.; Park, Y.H. Immunomodulatory effects of human amniotic membrane-derived mesenchymal stem cells. J. Veter-Sci. 2012, 13, 23–31. [Google Scholar] [CrossRef] [PubMed]
- Bilic, G.; Zeisberger, S.M.; Mallik, A.S.; Zimmermann, R.; Zisch, A.H. Comparative Characterization of Cultured Human Term Amnion Epithelial and Mesenchymal Stromal Cells for Application in Cell Therapy. Cell Transplant. 2008, 17, 955–968. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Kang, H.M.; Kim, H.; Kim, M.R.; Kwon, H.C.; Gye, M.C.; Kang, S.G.; Yang, H.S.; You, J. Ex Vivo Characteristics of Human Amniotic Membrane-Derived Stem Cells. Cloning Stem Cells 2007, 9, 581–594. [Google Scholar] [CrossRef] [PubMed]
- Alviano, F.; Fossati, V.; Marchionni, C.; Arpinati, M.; Bonsi, L.; Franchina, M.; Lanzoni, G.; Cantoni, S.; Cavallini, C.; Bianchi, F.; et al. Term amniotic membrane is a high throughput source for multipotent mesenchymal stem cells with the ability to differentiate into endothelial cells in vitro. BMC Dev. Biol. 2007, 7, 11. [Google Scholar] [CrossRef] [PubMed]
- Soncini, M.; Vertua, E.; Gibelli, L.; Zorzi, F.; Denegri, M.; Albertini, A.; Wengler, G.S.; Parolini, O. Isolation and characterization of mesenchymal cells from human fetal membranes. J. Tissue Eng. Regen. Med. 2007, 1, 296–305. [Google Scholar] [CrossRef] [PubMed]
- Huang, Q.; Yang, Y.; Luo, C.; Wen, Y.; Liu, R.; Li, S.; Chen, T.; Sun, H.; Tang, L. An efficient protocol to generate placental chorionic plate-derived mesenchymal stem cells with superior proliferative and immunomodulatory properties. Stem Cell Res. Ther. 2019, 10, 301. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Wu, J.; Han, L.; Jiang, X.; Yan, L.; Hao, J.; Wang, H. Comparative analysis of mesenchymal stem cells derived from amniotic membrane, umbilical cord, and chorionic plate under serum-free condition. Stem Cell Res. Ther. 2019, 10, 19. [Google Scholar] [CrossRef]
- Kim, M.J.; Shin, K.S.; Jeon, J.H.; Lee, D.R.; Shim, S.H.; Kim, J.K.; Cha, D.-H.; Yoon, T.K.; Kim, G.J. Human chorionic-plate-derived mesenchymal stem cells and Wharton’s jelly-derived mesenchymal stem cells: A comparative analysis of their potential as placenta-derived stem cells. Cell Tissue Res. 2011, 346, 53–64. [Google Scholar] [CrossRef] [PubMed]
- Majore, I.; Moretti, P.; Hass, R.; Kasper, C. Identification of subpopulations in mesenchymal stem cell-like cultures from human umbilical cord. Cell Commun. Signal. 2009, 7, 6. [Google Scholar] [CrossRef] [PubMed]
- Karahuseyinoglu, S.; Cinar, O.; Kilic, E.; Kara, F.; Akay, G.G.; Demiralp, D.Ö.; Tukun, D.; Uckan, D.; Can, A. Biology of Stem Cells in Human Umbilical Cord Stroma: In Situ and In Vitro Surveys. Stem Cells 2007, 25, 319–331. [Google Scholar] [CrossRef]
- Moretti, P.; Hatlapatka, T.; Marten, D.; Lavrentieva, A.; Majore, I.; Hass, R.; Kasper, C. Mesenchymal Stromal Cells Derived from Human Umbilical Cord Tissues: Primitive Cells with Potential for Clinical and Tissue Engineering Applications. Bioreact. Syst. Tissue Eng. II 2009, 123, 29–54. [Google Scholar] [CrossRef]
- He, H.; Nagamura-Inoue, T.; Tsunoda, H.; Yuzawa, M.; Yamamoto, Y.; Yorozu, P.; Agata, H.; Tojo, A. Stage-Specific Embryonic Antigen 4 in Wharton’s Jelly–Derived Mesenchymal Stem Cells Is Not a Marker for Proliferation and Multipotency. Tissue Eng. Part A 2014, 20, 1314–1324. [Google Scholar] [CrossRef] [PubMed]
- Batsali, A.K.; Kastrinaki, M.-C.; Papadaki, H.A.; Pontikoglou, C. Mesenchymal Stem Cells Derived from Wharton’s Jelly of the Umbilical Cord: Biological Properties and Emerging Clinical Applications. Curr. Stem Cell Res. Ther. 2013, 8, 144–155. [Google Scholar] [CrossRef]
- Stefańska, K.; Bryl, R.; Hutchings, G.; Shibli, J.A.; Dyszkiewicz-Konwińska, M. Human umbilical cord stem cells–the discovery, history and possible application. Med J. Cell Biol. 2020, 8, 78–82. [Google Scholar] [CrossRef]
- Moraghebi, R.; Kirkeby, A.; Chaves, P.; Rönn, R.E.; Sitnicka, E.; Parmar, M.; Larsson, M.; Herbst, A.; Woods, N.-B. Term amniotic fluid: An unexploited reserve of mesenchymal stromal cells for reprogramming and potential cell therapy applications. Stem Cell Res. Ther. 2017, 8, 190. [Google Scholar] [CrossRef]
- Moorefield, E.C.; McKee, E.E.; Solchaga, L.; Orlando, G.; Yoo, J.J.; Walker, S.; Furth, M.E.; Bishop, C.E. Cloned, CD117 Selected Human Amniotic Fluid Stem Cells Are Capable of Modulating the Immune Response. PLoS ONE 2011, 6, e26535. [Google Scholar] [CrossRef]
- Araújo, A.B.; Salton, G.D.; Furlan, J.M.; Schneider, N.; Angeli, M.H.; Laureano, Á.M.; Silla, L.; Passos, E.P.; Paz, A.H. Comparison of human mesenchymal stromal cells from four neonatal tissues: Amniotic membrane, chorionic membrane, placental decidua and umbilical cord. Cytotherapy 2017, 19, 577–585. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, M.S.V.; Bienert, M.; Müller, K.; Rath, B.; Goecke, T.; Opländer, C.; Braunschweig, T.; Mela, P.; Brümmendorf, T.H.; Beier, F.; et al. Comprehensive characterization of chorionic villi-derived mesenchymal stromal cells from human placenta. Stem Cell Res. Ther. 2018, 9, 28. [Google Scholar] [CrossRef] [PubMed]
- Tai, C.; Wang, L.; Xie, Y.; Gao, T.; Huang, F.; Wang, B. Analysis of Key Distinct Biological Characteristics of Human Placenta-Derived Mesenchymal Stromal Cells and Individual Heterogeneity Attributing to Donors. Cells Tissues Organs 2021, 210, 45–57. [Google Scholar] [CrossRef]
- Seok, J.; Jun, S.; Lee, J.O.; Kim, G.J. Mitochondrial Dynamics in Placenta-Derived Mesenchymal Stem Cells Regulate the Invasion Activity of Trophoblast. Int. J. Mol. Sci. 2020, 21, 8599. [Google Scholar] [CrossRef] [PubMed]
- Seok, J.; Jun, S.; Cho, J.; Park, S.; Lee, J.O.; Kim, G.J. Human placenta-derived mesenchymal stem cells induce trophoblast invasion via dynamic effects on mitochondrial function. J. Cell. Physiol. 2021, 236, 6678–6690. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.-E.; Ha, C.-W.; Jung, M.; Jin, H.-J.; Lee, M.; Song, H.; Choi, S.; Oh, W.; Yang, Y.-S. Mesenchymal stem/progenitor cells developed in cultures from UC blood. Cytotherapy 2004, 6, 476–486. [Google Scholar] [CrossRef] [PubMed]
- de la Torre, P.; Flores, A.I. Current status and future prospects of perinatal stem cells. Genes 2021, 12, 6. [Google Scholar] [CrossRef] [PubMed]
- Amati, E.; Sella, S.; Perbellini, O.; Alghisi, A.; Bernardi, M.; Chieregato, K.; Lievore, C.; Peserico, D.; Rigno, M.; Zilio, A.; et al. Generation of mesenchymal stromal cells from cord blood: Evaluation of in vitro quality parameters prior to clinical use. Stem Cell Res. Ther. 2017, 8, 14. [Google Scholar] [CrossRef] [PubMed]
- Yoshioka, S.; Miura, Y.; Iwasa, M.; Fujishiro, A.; Yao, H.; Miura, M.; Fukuoka, M.; Nakagawa, Y.; Yokota, A.; Hirai, H.; et al. Isolation of mesenchymal stromal/stem cells from small-volume umbilical cord blood units that do not qualify for the banking system. Int. J. Hematol. 2015, 102, 218–229. [Google Scholar] [CrossRef] [PubMed]
- Seifabadi, Z.S.; Rezaei-Tazangi, F.; Azarbarz, N.; Nejad, D.B.; Mohammadiasl, J.; Darabi, H.; Pezhmanlarki-Tork, S. Assessment of viability of wharton’s jelly mesenchymal stem cells encapsulated in alginate scaffold by WST-8 assay kit. Med J. Cell Biol. 2021, 9, 42–47. [Google Scholar] [CrossRef]
- Borys-Wójcik, S.; Moncrieff, L.; Kempisty, B.; Dyszkiewicz-Konwińska, M. Current state of umbilical cord stem cells in humans. Med J. Cell Biol. 2019, 7, 86–89. [Google Scholar] [CrossRef]
- Borys-Wójcik, S.; Józkowiak, M.; Stefańska, K.; Knap, S.; Pieńkowski, W.; Gutaj, P.; Bruska, M.; Kempisty, B. Historical background of umbilical stem cell culture. Med J. Cell Biol. 2019, 7, 11–14. [Google Scholar] [CrossRef]
- Lim, I.J.; Phan, T.T. Epithelial and Mesenchymal Stem Cells from the Umbilical Cord Lining Membrane. Cell Transplant. 2014, 23, 497–503. [Google Scholar] [CrossRef]
- Wang, H.; Yan, X.; Jiang, Y.; Wang, Z.; Li, Y.; Shao, Q. The human umbilical cord stem cells improve the viability of OA degenerated chondrocytes. Mol. Med. Rep. 2018, 17, 4474–4482. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.-S.; Shih, Y.-T.; Cheng, Y.-C.; Min, M.-Y. Transformation of human umbilical mesenchymal cells into neurons in vitro. J. Biomed. Sci. 2004, 11, 652–660. [Google Scholar] [CrossRef] [PubMed]
- Campard, D.; Lysy, P.A.; Najimi, M.; Sokal, E. Native Umbilical Cord Matrix Stem Cells Express Hepatic Markers and Differentiate Into Hepatocyte-like Cells. Gastroenterology 2008, 134, 833–848. [Google Scholar] [CrossRef] [PubMed]
- Wu, K.H.; Zhou, B.; Lu, S.H.; Feng, B.; Yang, S.G.; Du, W.T.; Gu, D.S.; Han, Z.C.; Liu, Y.L. In vitro and in vivo differentiation of human umbilical cord derived stem cells into endothelial cells. J. Cell. Biochem. 2007, 100, 608–616. [Google Scholar] [CrossRef] [PubMed]
- Stefańska, K.; Bryl, R.; Moncrieff, L.; Pinto, N.; Shibli, J.A.; Dyszkiewicz-Konwińska, M. Mesenchymal stem cells–a historical overview. Med J. Cell Biol. 2020, 8, 83–87. [Google Scholar] [CrossRef]
- Loukogeorgakis, S.P.; De Coppi, P. Concise Review: Amniotic Fluid Stem Cells: The Known, the Unknown, and Potential Regenerative Medicine Applications. STEM CELLS 2017, 35, 1663–1673. [Google Scholar] [CrossRef] [PubMed]
- Eslaminejad, M.B.; Jahangir, S. Amniotic Fluid Stem Cells and Their Application in Cell-Based Tissue Regeneration. Int J Fertil Steril 2012, 6, 147–156. [Google Scholar]
- Loukogeorgakis, S.P.; De Coppi, P. Stem cells from amniotic fluid–Potential for regenerative medicine. Best Pract. Res. Clin. Obstet. Gynaecol. 2016, 31, 45–57. [Google Scholar] [CrossRef]
- Bajek, A.; Olkowska, J.; Walentowicz-Sadłecka, M.; Walentowicz, P.; Sadłecki, P.; Grabiec, M.; Bodnar, M.; Marszałek, A.; Dębski, R.; Porowińska, D.; et al. High Quality Independent From a Donor: Human Amniotic Fluid Derived Stem Cells-A Practical Analysis Based on 165 Clinical Cases. J. Cell. Biochem. 2016, 118, 116–126. [Google Scholar] [CrossRef]
- Tsai, M.S.; Lee, J.L.; Chang, Y.J.; Hwang, S.M. Isolation of human multipotent mesenchymal stem cells from second-trimester amniotic fluid using a novel two-stage culture protocol. Hum. Reprod. 2004, 19, 1450–1456. [Google Scholar] [CrossRef] [PubMed]
- Laowanitwattana, T.; Aungsuchawan, S.; Narakornsak, S.; Markmee, R.; Tancharoen, W.; Keawdee, J.; Boonma, N.; Tasuya, W.; Peerapapong, L.; Pangjaidee, N.; et al. Osteoblastic differentiation potential of human amniotic fluid-derived mesenchymal stem cells in different culture conditions. Acta Histochem. 2018, 120, 701–712. [Google Scholar] [CrossRef]
- Di Trapani, M.; Bassi, G.; Fontana, E.; Giacomello, L.; Pozzobon, M.; Guillot, P.V.; De Coppi, P.; Krampera, M. Immune Regulatory Properties of CD117pos Amniotic Fluid Stem Cells Vary According to Gestational Age. Stem Cells Dev. 2015, 24, 132–143. [Google Scholar] [CrossRef] [PubMed]
- Wolfrum, K.; Wang, Y.; Prigione, A.; Sperling, K.; Lehrach, H.; Adjaye, J. The LARGE Principle of Cellular Reprogramming: Lost, Acquired and Retained Gene Expression in Foreskin and Amniotic Fluid-Derived Human iPS Cells. PLoS ONE 2010, 5, e13703. [Google Scholar] [CrossRef]
- Balbi, C.; Bollini, S. Fetal and perinatal stem cells in cardiac regeneration: Moving forward to the paracrine era. Placenta 2017, 59, 96–106. [Google Scholar] [CrossRef]
- Sibiak, R.; Jaworski, M.; Barrett, S.; Bryl, R.; Gutaj, P.; Kulus, J.; Bukowska, D.; Petitte, J.; Crha, I.; Ventruba, P.; et al. Trophoblast stem cells-methods of isolation, histological and cellular characteristic, and their possible applications in human and animal models. Med J. Cell Biol. 2020, 8, 95–100. [Google Scholar] [CrossRef]
- Bischof, P.; Irminger-Finger, I. The human cytotrophoblastic cell, a mononuclear chameleon. Int. J. Biochem. Cell Biol. 2005, 37, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Gerami-Naini, B.; Dovzhenko, O.V.; Durning, M.; Wegner, F.H.; Thomson, J.A.; Golos, T.G. Trophoblast Differentiation in Embryoid Bodies Derived from Human Embryonic Stem Cells. Endocrinol. 2004, 145, 1517–1524. [Google Scholar] [CrossRef]
- Xu, R.-H.; Chen, X.; Li, D.S.; Li, R.; Addicks, G.C.; Glennon, C.; Zwaka, T.P.; Thomson, J.A. BMP4 initiates human embryonic stem cell differentiation to trophoblast. Nat. Biotechnol. 2002, 20, 1261–1264. [Google Scholar] [CrossRef]
- Genbacev, O.; Miller, R. Post-implantation Differentiation and Proliferation of Cytotrophoblast Cells: In Vitro Models—A Review. Placenta 2000, 21, S45–S49. [Google Scholar] [CrossRef] [PubMed]
- Caniggia, I.; Mostachfi, H.; Winter, J.; Gassmann, M.; Lye, S.J.; Kuliszewski, M.; Post, M. Hypoxia-inducible factor-1 mediates the biological effects of oxygen on human trophoblast differentiation through TGFβ3. J. Clin. Investig. 2000, 105, 577–587. [Google Scholar] [CrossRef] [PubMed]
- Frendo, J.-L.; Olivier, D.; Cheynet, V.; Blond, J.-L.; Bouton, O.; Vidaud, M.; Rabreau, M.; Evain-Brion, D.; Mallet, F. Direct Involvement of HERV-W Env Glycoprotein in Human Trophoblast Cell Fusion and Differentiation. Mol. Cell. Biol. 2003, 23, 3566–3574. [Google Scholar] [CrossRef] [PubMed]
- Frendo, J.-L.; Cronier, L.; Bertin, G.; Guibourdenche, J.; Vidaud, M.; Evain-Brion, D.; Malassiné, A. Involvement of connexin 43 in human trophoblast cell fusion and differentiation. J. Cell Sci. 2003, 116, 3413–3421. [Google Scholar] [CrossRef] [PubMed]
- Okae, H.; Toh, H.; Sato, T.; Hiura, H.; Takahashi, S.; Shirane, K.; Kabayama, Y.; Suyama, M.; Sasaki, H.; Arima, T. Derivation of Human Trophoblast Stem Cells. Cell Stem Cell 2018, 22, 50–63.e6. [Google Scholar] [CrossRef] [PubMed]
- Latos, P.A.; Hemberger, M. From the stem of the placental tree: Trophoblast stem cells and their progeny. Development 2016, 143, 3650–3660. [Google Scholar] [CrossRef] [PubMed]
- Gauthier-Fisher, A.; Szaraz, P.; Librach, C.L. Pericytes in the Umbilical Cord. In Advances in Experimental Medicine and Biology; Springer: Berlin/Heidelberg, Germany, 2019; Volume 1122, pp. 211–233. [Google Scholar]
- Subramanian, A.; Fong, C.-Y.; Biswas, A.; Bongso, A. Comparative Characterization of Cells from the Various Compartments of the Human Umbilical Cord Shows that the Wharton’s Jelly Compartment Provides the Best Source of Clinically Utilizable Mesenchymal Stem Cells. PLoS ONE 2015, 10, e0127992. [Google Scholar] [CrossRef]
- Takechi, K.; Kuwabara, Y.; Mizuno, M. Ultrastructural and immunohistochemical studies of Wharton’s jelly umbilical cord cells. Placenta 1993, 14, 235–245. [Google Scholar] [CrossRef]
- Schugar, R.C.; Chirieleison, S.M.; Wescoe, K.E.; Schmidt, B.T.; Askew, Y.; Nance, J.J.; Evron, J.M.; Péault, B.; Deasy, B.M. High Harvest Yield, High Expansion, and Phenotype Stability of CD146 Mesenchymal Stromal Cells from Whole Primitive Human Umbilical Cord Tissue. J. Biomed. Biotechnol. 2009, 2009, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Crisan, M.; Yap, S.; Casteilla, L.; Chen, C.-W.; Corselli, M.; Park, T.S.; Andriolo, G.; Sun, B.; Zheng, B.; Zhang, L.; et al. A Perivascular Origin for Mesenchymal Stem Cells in Multiple Human Organs. Cell Stem Cell 2008, 3, 301–313. [Google Scholar] [CrossRef] [PubMed]
- Sarugaser, R.; Lickorish, D.; Baksh, D.; Hosseini, M.M.; Davies, J.E. Human Umbilical Cord Perivascular (HUCPV) Cells: A Source of Mesenchymal Progenitors. Stem Cells 2005, 23, 220–229. [Google Scholar] [CrossRef]
- Stratman, A.N.; Schwindt, A.E.; Malotte, K.M.; Davis, G.E. Endothelial-derived PDGF-BB and HB-EGF coordinately regulate pericyte recruitment during vasculogenic tube assembly and stabilization. Blood 2010, 116, 4720–4730. [Google Scholar] [CrossRef] [PubMed]
- Grzywocz, Z.; Pius-Sadowska, E.; Klos, P.; Gryzik, M.; Wasilewska, D.; Aleksandrowicz, B.; Dworczynska, M.; Sabalinska, S.; Hoser, G.; Machalinski, B.; et al. Growth factors and their receptors derived from human amniotic cells in vitro. Folia Histochem. Et Cytobiol. 2014, 52, 163–170. [Google Scholar] [CrossRef]
- Takashima, Y.; Era, T.; Nakao, K.; Kondo, S.; Kasuga, M.; Smith, A.; Nishikawa, S.-I. Neuroepithelial Cells Supply an Initial Transient Wave of MSC Differentiation. Cell 2007, 129, 1377–1388. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, R.; Griparic, L.; Umana, M.; Burgee, K.; Vargas, V.; Nasrallah, R.; Silva, F.; Patel, A. An Efficient Approach to Isolation and Characterization of Pre- and Postnatal Umbilical Cord Lining Stem Cells for Clinical Applications. Cell Transplant. 2010, 19, 1439–1449. [Google Scholar] [CrossRef] [PubMed]
- Dobreva, M.; Pereira, P.N.G.; Deprest, J.; Zwijsen, A. On the origin of amniotic stem cells: Of mice and men. Int. J. Dev. Biol. 2010, 54, 761–777. [Google Scholar] [CrossRef]
- Fauza, D. Amniotic fluid and placental stem cells. Best Pract. Res. Clin. Obstet. Gynaecol. 2004, 18, 877–891. [Google Scholar] [CrossRef]
- Tong, X. Amniotic fluid may act as a transporting pathway for signaling molecules and stem cells during the embryonic development of amniotes. J. Chin. Med Assoc. 2013, 76, 606–610. [Google Scholar] [CrossRef] [PubMed]
- Torricelli, F.; Brizzi, L.; A Bernabei, P.; Gheri, G.; Di Lollo, S.; Nutini, L.; Lisi, E.; Di Tommaso, M.; Cariati, E. Identification of hematopoietic progenitor cells in human amniotic fluid before the 12th week of gestation. Ital. J. Anat. Embryol. Arch. Ital. Di Anat. Ed Embriol. 1993, 98, 264. [Google Scholar]
- In ’t Anker, P.S.; Scherjon, S.A.; Kleijburg-van der Keur, C.; Noort, W.A.; Claas, F.H.J.; Willemze, R.; Fibbe, W.E.; Kanhai, H.H.H. Amniotic fluid as a novel source of mesenchymal stem cells for therapeutic transplantation. Blood 2003, 102, 1548–1549. [Google Scholar] [CrossRef]
- Kolambkar, Y.M.; Peister, A.; Soker, S.; Atala, A.; Guldberg, R.E. Chondrogenic differentiation of amniotic fluid-derived stem cells. J. Mol. Histol. 2007, 38, 405–413. [Google Scholar] [CrossRef]
- Bajek, A.; Olkowska, J.; Walentowicz-Sadłecka, M.; Sadłecki, P.; Grabiec, M.; Porowinska, D.; Drewa, T.; Roszkowski, K. Human Adipose-Derived and Amniotic Fluid-Derived Stem Cells: A Preliminary In Vitro Study Comparing Myogenic Differentiation Capability. Med Sci. Monit. 2018, 24, 1733–1741. [Google Scholar] [CrossRef]
- Rahman, S.; Spitzhorn, L.-S.; Wruck, W.; Hagenbeck, C.; Balan, P.; Graffmann, N.; Bohndorf, M.; Ncube, A.; Guillot, P.V.; Fehm, T.; et al. The presence of human mesenchymal stem cells of renal origin in amniotic fluid increases with gestational time. Stem Cell Res. Ther. 2018, 9, 1–17. [Google Scholar] [CrossRef]
- Hengstschläger, M.; Siegel, N.; Valli, A.; Fuchs, C.; Rosner, M. Expression of mTOR pathway proteins in human amniotic fluid stem cells. Int. J. Mol. Med. 2009, 23, 779–784. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Valli, A.; Rosner, M.; Fuchs, C.; Siegel, N.; E Bishop, C.; Dolznig, H.; Mädel, U.; Feichtinger, W.; Atala, A.; Hengstschläger, M. Embryoid body formation of human amniotic fluid stem cells depends on mTOR. Oncogene 2010, 29, 966–977. [Google Scholar] [CrossRef] [PubMed]
- Barboni, B.; Russo, V.; Berardinelli, P.; Mauro, A.; Valbonetti, L.; Sanyal, H.; Canciello, A.; Greco, L.; Muttini, A.; Gatta, V.; et al. Placental Stem Cells from Domestic Animals: Translational Potential and Clinical Relevance. Cell Transplant. 2018, 27, 93–116. [Google Scholar] [CrossRef]
- Kusuma, G.D.; Menicanin, D.; Gronthos, S.; Manuelpillai, U.; Abumaree, M.H.; Pertile, M.D.; Brennecke, S.P.; Kalionis, B. Ectopic Bone Formation by Mesenchymal Stem Cells Derived from Human Term Placenta and the Decidua. PLoS ONE 2015, 10, e0141246. [Google Scholar] [CrossRef] [PubMed]
- Reddy, S.; Wasnik, S.; Guha, A.; Kumar, J.M.; Sinha, A.; Singh, S. Evaluation of nano-biphasic calcium phosphate ceramics for bone tissue engineering applications: In vitro and preliminary in vivo studies. J. Biomater. Appl. 2013, 27, 565–575. [Google Scholar] [CrossRef]
- Wei, J.P.; Nawata, M.; Wakitani, S.; Kametani, K.; Ota, M.; Toda, A.; Konishi, I.; Ebara, S.; Nikaido, T. Human Amniotic Mesenchymal Cells Differentiate into Chondrocytes. Cloning Stem Cells 2009, 11, 19–26. [Google Scholar] [CrossRef]
- Li, F.; Chen, Y.-Z.; Miao, Z.-N.; Zheng, S.-Y.; Jin, J. Human Placenta-Derived Mesenchymal Stem Cells with Silk Fibroin Biomaterial in the Repair of Articular Cartilage Defects. Cell. Reprogramming 2012, 14, 334–341. [Google Scholar] [CrossRef] [PubMed]
- Nogami, M.; Tsuno, H.; Koike, C.; Okabe, M.; Yoshida, T.; Seki, S.; Matsui, Y.; Kimura, T.; Nikaido, T. Isolation and Characterization of Human Amniotic Mesenchymal Stem Cells and Their Chondrogenic Differentiation. Transplantation 2012, 93, 1221–1228. [Google Scholar] [CrossRef] [PubMed]
- Zhao, P.; Ise, H.; Hongo, M.; Ota, M.; Konishi, I.; Nikaido, T. Human Amniotic Mesenchymal Cells Have Some Characteristics of Cardiomyocytes. Transplant. 2005, 79, 528–535. [Google Scholar] [CrossRef]
- Zhang, W.; Liu, X.-C.; Yang, L.; Zhu, D.-L.; Zhang, Y.-D.; Chen, Y.; Zhang, H.-Y. Wharton’s jelly-derived mesenchymal stem cells promote myocardial regeneration and cardiac repair after miniswine acute myocardial infarction. Coron. Artery Dis. 2013, 24, 549–558. [Google Scholar] [CrossRef] [PubMed]
- Nascimento, D.S.; Mosqueira, D.; Sousa, L.M.; Teixeira, M.; Filipe, M.; Resende, T.P.; Araújo, A.F.; Valente, M.; Almeida, J.; Martins, J.P.; et al. Human umbilical cord tissue-derived mesenchymal stromal cells attenuate remodeling after myocardial infarction by proangiogenic, antiapoptotic, and endogenous cell-activation mechanisms. Stem Cell Res. Ther. 2014, 5, 5. [Google Scholar] [CrossRef]
- Simioniuc, A.; Campan, M.; Lionetti, V.; Marinelli, M.; Aquaro, G.D.; Cavallini, C.; Valente, S.; Di Silvestre, D.; Cantoni, S.; Bernini, F.; et al. Placental stem cells pre-treated with a hyaluronan mixed ester of butyric and retinoic acid to cure infarcted pig hearts: A multimodal study. Cardiovasc. Res. 2011, 90, 546–556. [Google Scholar] [CrossRef]
- Weiss, M.L.; Medicetty, S.; Bledsoe, A.R.; Rachakatla, R.S.; Choi, M.; Merchav, S.; Luo, Y.; Rao, M.S.; Velagaleti, G.; Troyer, D. Human Umbilical Cord Matrix Stem Cells: Preliminary Characterization and Effect of Transplantation in a Rodent Model of Parkinson’s Disease. Stem Cells 2006, 24, 781–792. [Google Scholar] [CrossRef]
- Fu, Y.-S.; Cheng, Y.-C.; Lin, M.-Y.A.; Cheng, H.; Chu, P.-M.; Chou, S.-C.; Shih, Y.-H.; Ko, M.-H.; Sung, M.-S. Conversion of Human Umbilical Cord Mesenchymal Stem Cells in Wharton’s Jelly to Dopaminergic Neurons In Vitro: Potential Therapeutic Application for Parkinsonism. Stem Cells 2006, 24, 115–124. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Kim, E.; Koh, S.-E.; Maeng, S.; Lee, W.-D.; Lim, J.; Shim, I.; Lee, Y.-J. Dopaminergic differentiation of neural progenitors derived from placental mesenchymal stem cells in the brains of Parkinson’s disease model rats and alleviation of asymmetric rotational behavior. Brain Res. 2012, 1466, 158–166. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.-S.; Kim, H.S.; Park, J.-M.; Kim, H.W.; Park, M.-K.; Lee, H.-S.; Lim, D.S.; Lee, T.H.; Chopp, M.; Moon, J. Long-term immunomodulatory effect of amniotic stem cells in an Alzheimer’s disease model. Neurobiol. Aging 2013, 34, 2408–2420. [Google Scholar] [CrossRef]
- Mohamadi, Y.; Moghahi, S.M.H.N.; Mousavi, M.; Borhani-Haghighi, M.; Abolhassani, F.; Kashani, I.R.; Hassanzadeh, G. Intrathecal transplantation of Wharton’s jelly mesenchymal stem cells suppresses the NLRP1 inflammasome in the rat model of spinal cord injury. J. Chem. Neuroanat. 2019, 97, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Badner, A.; Hacker, J.; Hong, J.; Mikhail, M.; Vawda, R.; Fehlings, M.G. Splenic involvement in umbilical cord matrix-derived mesenchymal stromal cell-mediated effects following traumatic spinal cord injury. J. Neuroinflammation 2018, 15, 219. [Google Scholar] [CrossRef]
- Chudickova, M.; Vackova, I.; Urdzikova, L.M.; Jancova, P.; Kekulova, K.; Rehorova, M.; Turnovcova, K.; Jendelova, P.; Kubinova, S. The Effect of Wharton Jelly-Derived Mesenchymal Stromal Cells and Their Conditioned Media in the Treatment of a Rat Spinal Cord Injury. Int. J. Mol. Sci. 2019, 20, 4516. [Google Scholar] [CrossRef]
- Krupa, P.; Vackova, I.; Ruzicka, J.; Zaviskova, K.; Dubisova, J.; Koci, Z.; Turnovcova, K.; Urdzikova, L.M.; Kubinova, S.; Rehak, S.; et al. The Effect of Human Mesenchymal Stem Cells Derived from Wharton’s Jelly in Spinal Cord Injury Treatment Is Dose-Dependent and Can Be Facilitated by Repeated Application. Int. J. Mol. Sci. 2018, 19, 1503. [Google Scholar] [CrossRef] [PubMed]
- Clark, K.; Zhang, S.; Barthe, S.; Kumar, P.; Pivetti, C.; Kreutzberg, N.; Reed, C.; Wang, Y.; Paxton, Z.; Farmer, D.; et al. Placental Mesenchymal Stem Cell-Derived Extracellular Vesicles Promote Myelin Regeneration in an Animal Model of Multiple Sclerosis. Cells 2019, 8, 1497. [Google Scholar] [CrossRef]
- Fisher-Shoval, Y.; Barhum, Y.; Sadan, O.; Yust-Katz, S.; Ben-Zur, T.; Lev, N.; Benkler, C.; Hod, M.; Melamed, E.; Offen, D. Transplantation of Placenta-Derived Mesenchymal Stem Cells in the EAE Mouse Model of MS. J. Mol. Neurosci. 2012, 48, 176–184. [Google Scholar] [CrossRef] [PubMed]
- Tsai, P.-C.; Fu, T.-W.; Chen, Y.-M.A.; Ko, T.-L.; Chen, T.-H.; Shih, Y.-H.; Hung, S.-C.; Fu, Y.-S. The therapeutic potential of human umbilical mesenchymal stem cells from Wharton’s jelly in the treatment of rat liver fibrosis. Liver Transplant. 2009, 15, 484–495. [Google Scholar] [CrossRef] [PubMed]
- Moodley, Y.; Atienza, D.; Manuelpillai, U.; Samuel, C.S.; Tchongue, J.; Ilancheran, S.; Boyd, R.; Trounson, A. Human Umbilical Cord Mesenchymal Stem Cells Reduce Fibrosis of Bleomycin-Induced Lung Injury. Am. J. Pathol. 2009, 175, 303–313. [Google Scholar] [CrossRef]
- Yu, J.; Hao, G.; Wang, D.; Liu, J.; Dong, X.; Sun, Y.; Pan, Q.; Li, Y.; Shi, X.; Li, L.; et al. Therapeutic Effect and Location of GFP-Labeled Placental Mesenchymal Stem Cells on Hepatic Fibrosis in Rats. Stem Cells Int. 2017, 2017, 1–11. [Google Scholar] [CrossRef]
- Jung, J.; Choi, J.H.; Lee, E.; Park, J.-W.; Oh, I.-H.; Hwang, S.-G.; Kim, K.-S.; Kim, G.J. Human Placenta-Derived Mesenchymal Stem Cells Promote Hepatic Regeneration in CCl4-Injured Rat Liver Model via Increased Autophagic Mechanism. Stem Cells 2013, 31, 1584–1596. [Google Scholar] [CrossRef]
- Zhang, D.; Jiang, M.; Miao, D. Transplanted Human Amniotic Membrane-Derived Mesenchymal Stem Cells Ameliorate Carbon Tetrachloride-Induced Liver Cirrhosis in Mouse. PLoS ONE 2011, 6, e16789. [Google Scholar] [CrossRef]
- Cao, H.; Yang, J.; Yu, J.; Pan, Q.; Li, J.; Zhou, P.; Li, Y.; Pan, X.; Li, J.; Wang, Y.; et al. Therapeutic potential of transplanted placental mesenchymal stem cells in treating Chinese miniature pigs with acute liver failure. BMC Med. 2012, 10, 56. [Google Scholar] [CrossRef]
- Kim, H.; Shin, T.; Lee, B.; Yu, K.; Seo, Y.; Lee, S.; Seo, M.; Hong, I.; Choi, S.W.; Seo, K.; et al. Human Umbilical Cord Blood Mesenchymal Stem Cells Reduce Colitis in Mice by Activating NOD2 Signaling to COX2. Gastroenterol. 2013, 145, 1392–1403.e8. [Google Scholar] [CrossRef]
- Song, J.-Y.; Kang, H.J.; Hong, J.S.; Kim, C.J.; Shim, J.-Y.; Lee, C.W.; Choi, J. Umbilical cord-derived mesenchymal stem cell extracts reduce colitis in mice by re-polarizing intestinal macrophages. Sci. Rep. 2017, 7, 9412. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.S.; Chun, S.Y.; Kil Lee, J.; Lim, H.J.; Bae, J.-S.; Chung, H.-Y.; Atala, A.; Soker, S.; Yoo, J.J.; Kwon, T.G. Human amniotic fluid stem cell injection therapy for urethral sphincter regeneration in an animal model. BMC Med. 2012, 10, 94. [Google Scholar] [CrossRef] [PubMed]
- Sabapathy, V.; Sundaram, B.; VM, S.; Mankuzhy, P.; Kumar, S. Human Wharton’s Jelly Mesenchymal Stem Cells plasticity augments scar-free skin wound healing with hair growth. PLoS ONE 2014, 9, e93726. [Google Scholar] [CrossRef]
- Khalifeh Soltani, S.; Forogh, B.; Ahmadbeigi, N.; Hadizadeh Kharazi, H.; Fallahzadeh, K.; Kashani, L.; Karami, M.; Kheyrollah, Y.; Vasei, M. Safety and efficacy of allogenic placental mesenchymal stem cells for treating knee osteoarthritis: A pilot study. Cytotherapy 2019, 21. [Google Scholar] [CrossRef]
- Winkler, T.; Perka, C.; von Roth, P.; Agres, A.N.; Plage, H.; Preininger, B.; Pumberger, M.; Geissler, S.; Hagai, E.L.; Ofir, R.; et al. Immunomodulatory placental-expanded, mesenchymal stromal cells improve muscle function following hip arthroplasty. J. Cachexia Sarcopenia Muscle 2018, 9, 880–897. [Google Scholar] [CrossRef] [PubMed]
- Norgren, L.; Weiss, N.; Nikol, S.; Hinchliffe, R.J.; Lantis, J.C.; Patel, M.R.; Reinecke, H.; Ofir, R.; Rosen, Y.; Peres, D.; et al. PLX-PAD Cell Treatment of Critical Limb Ischaemia: Rationale and Design of the PACE Trial. Eur. J. Vasc. Endovasc. Surg. 2019, 57. [Google Scholar] [CrossRef] [PubMed]
- Levy, J.A.; Marchand, M.; Iorio, L.; Cassini, W.; Zahalsky, M.P. Determining the Feasibility of Managing Erectile Dysfunction in Humans With Placental-Derived Stem Cells. J. Osteopat. Med. 2016, 116, e1–e5. [Google Scholar] [CrossRef] [PubMed]
- Zeng, X.; Tang, Y.; Hu, K.; Jiao, W.; Ying, L.; Zhu, L.; Liu, J.; Xu, J. Three-week topical treatment with placenta-derived mesenchymal stem cells hydrogel in a patient with diabetic foot ulcer: A case report. Medcine 2017, 96(51), e9212. [Google Scholar] [CrossRef]
- Harrell, C.R.; Miloradovic, D.; Sadikot, R.; Fellabaum, C.; Markovic, B.S.; Miloradovic, D.; Acovic, A.; Djonov, V.; Arsenijevic, N.; Volarevic, V. Molecular and Cellular Mechanisms Responsible for Beneficial Effects of Mesenchymal Stem Cell-Derived Product “Exo-d-MAPPS” in Attenuation of Chronic Airway Inflammation. Anal. Cell. Pathol. 2020, 2020, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Hashemian, S.-M.R.; Aliannejad, R.; Zarrabi, M.; Soleimani, M.; Vosough, M.; Hosseini, S.-E.; Hossieni, H.; Keshel, S.H.; Naderpour, Z.; Hajizadeh-Saffar, E.; et al. Mesenchymal stem cells derived from perinatal tissues for treatment of critically ill COVID-19-induced ARDS patients: A case series. Stem Cell Res. Ther. 2021, 12, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Ringden, O.; Baygan, A.; Remberger, M.; Gustafsson, B.; Winiarski, J.; Khoein, B.; Moll, G.; Klingspor, L.; Westgren, M.; Sadeghi, B. Placenta-Derived Decidua Stromal Cells for Treatment of Severe Acute Graft-Versus-Host Disease. Stem Cells Transl. Med. 2018, 7, 325–331. [Google Scholar] [CrossRef] [PubMed]
- Dilogo, I.H.; Canintika, A.F.; Hanitya, A.L.; Pawitan, J.A.; Liem, I.K.; Pandelaki, J. Umbilical cord-derived mesenchymal stem cells for treating osteoarthritis of the knee: A single-arm, open-label study. Eur. J. Orthop. Surg. Traumatol. 2020, 30, 799–807. [Google Scholar] [CrossRef] [PubMed]
- Matas, J.; Orrego, M.; Amenabar, D.; Infante, C.; Tapia-Limonchi, R.; Cadiz, M.I.; Alcayaga-Miranda, F.; González, P.L.; Muse, E.; Khoury, M.; et al. Umbilical Cord-Derived Mesenchymal Stromal Cells (MSCs) for Knee Osteoarthritis: Repeated MSC Dosing Is Superior to a Single MSC Dose and to Hyaluronic Acid in a Controlled Randomized Phase I/II Trial. Stem Cells Transl. Med. 2019, 8, 215–224. [Google Scholar] [CrossRef] [PubMed]
- Park, Y.-B.; Ha, C.-W.; Lee, C.-H.; Yoon, Y.C. Cartilage Regeneration in Osteoarthritic Patients by a Composite of Allogeneic Umbilical Cord Blood-Derived Mesenchymal Stem Cells and Hyaluronate Hydrogel: Results from a Clinical Trial for Safety and Proof-of-Concept with 7 Years of Extended Follow-Up. Stem Cells Transl. Med. 2016, 6, 613–621. [Google Scholar] [CrossRef]
- Wang, Y.; Jin, W.; Liu, H.; Cui, Y.; Mao, Q.; Fei, Z.; Xiang, C. Curative effect of human umbilical cord mesenchymal stem cells by intra-articular injection for degenerative knee osteoarthritis. Zhongguo Xiu Fu Chong Jian Wai Ke Za Zhi = Zhongguo Xiufu Chongjian Waike Zazhi = Chin. J. Reparative Reconstr. Surg. 2016, 30, 1472–1477. [Google Scholar] [CrossRef]
- He, X.; Wang, Q.; Zhao, Y.; Zhang, H.; Wang, B.; Pan, J.; Li, J.; Yu, H.; Wang, L.; Dai, J.; et al. Effect of Intramyocardial Grafting Collagen Scaffold With Mesenchymal Stromal Cells in Patients with Chronic Ischemic Heart Disease: A Randomized Clinical Trial. JAMA Netw. Open 2020, 3, e2016236. [Google Scholar] [CrossRef] [PubMed]
- Bartolucci, J.; Verdugo, F.J.; González, P.L.; Larrea, R.E.; Abarzua, E.; Goset, C.; Rojo, P.; Palma, I.; Lamich, R.; Pedreros, P.A.; et al. Safety and efficacy of the intravenous infusion of umbilical cord mesenchymal stem cells in patients with heart failure: A phase 1/2 randomized controlled trial (RIMECARD trial [Randomized clinical trial of intravenous infusion umbilical cord mesenchymal. Circ. Res. 2017, 121, 1192–1204. [Google Scholar] [CrossRef] [PubMed]
- Hashemi, S.; Mohammadi, A.A.; Kabiri, H.; Hashempoor, M.R.; Mahmoodi, M.; Amini, M.; Mehrabani, D.; Mahmudi, M. The healing effect of Wharton’s jelly stem cells seeded on biological scaffold in chronic skin ulcers: A randomized clinical trial. J. Cosmet. Dermatol. 2019, 18, 1961–1967. [Google Scholar] [CrossRef]
- Kim, Y.J.; Ahn, H.J.; Lee, S.H.; Lee, M.H.; Kang, K.S. Effects of conditioned media from human umbilical cord blood-derived mesenchymal stem cells in the skin immune response. Biomed. Pharmacother. 2020, 131, 110789. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Kim, B.; Kim, S.; Lee, Y.I.; Kim, J.; Lee, J.H. The effect of human umbilical cord blood–derived mesenchymal stem cell media containing serum on recovery after laser treatment: A double-blinded, randomized, split-face controlled study. J. Cosmet. Dermatol. 2020, 19, 651–656. [Google Scholar] [CrossRef]
- He, X.; Yang, Y.; Yao, M.; Yang, L.; Ao, L.; Hu, X.; Li, Z.; Wu, X.; Tan, Y.; Xing, W.; et al. Combination of human umbilical cord mesenchymal stem (stromal) cell transplantation with IFN-γtreatment synergistically improves the clinical outcomes of patients with rheumatoid arthritis. Ann. Rheum. Dis. 2020, 79, 1298–1304. [Google Scholar] [CrossRef] [PubMed]
- Qi, T.; Gao, H.; Dang, Y.; Huang, S.; Peng, M. Cervus and cucumis peptides combined umbilical cord mesenchymal stem cells therapy for rheumatoid arthritis. Medicine 2020, 99, e21222. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Huang, S.; Li, S.; Li, M.; Shi, J.; Bai, W.; Wang, Q.; Zheng, L.; Liu, Y. Efficacy and Safety of Umbilical Cord Mesenchymal Stem Cell Therapy for Rheumatoid Arthritis Patients: A Prospective Phase I/II Study. Drug Des. Dev. Ther. 2019, 13, 4331–4340. [Google Scholar] [CrossRef] [PubMed]
- Park, E.H.; Lim, H.; Lee, S.; Roh, K.; Seo, K.; Kang, K.; Shin, K. Intravenous Infusion of Umbilical Cord Blood-Derived Mesenchymal Stem Cells in Rheumatoid Arthritis: A Phase Ia Clinical Trial. Stem Cells Transl. Med. 2018, 7, 636–642. [Google Scholar] [CrossRef] [PubMed]
- Deng, D.; Zhang, P.; Guo, Y.; Lim, T.O. A randomised double-blind, placebo-controlled trial of allogeneic umbilical cord-derived mesenchymal stem cell for lupus nephritis. Ann. Rheum. Dis. 2017, 76, 1436–1439. [Google Scholar] [CrossRef]
- Riordan, N.H.; Morales, I.; Fernández, G.; Allen, N.; Fearnot, N.E.; Leckrone, M.E.; Markovich, D.J.; Mansfield, D.; Avila, D.; Patel, A.N.; et al. Clinical feasibility of umbilical cord tissue-derived mesenchymal stem cells in the treatment of multiple sclerosis. J. Transl. Med. 2018, 16, 57. [Google Scholar] [CrossRef]
- Gu, J.; Huang, L.; Zhang, C.; Wang, Y.; Zhang, R.; Tu, Z.; Wang, H.; Zhou, X.; Xiao, Z.; Liu, Z.; et al. Therapeutic evidence of umbilical cord-derived mesenchymal stem cell transplantation for cerebral palsy: A randomized, controlled trial. Stem Cell Res. Ther. 2020, 11, 43. [Google Scholar] [CrossRef]
- Riordan, N.H.; Hincapié, M.L.; Morales, I.; Fernández, G.; Allen, N.; Leu, C.; Madrigal, M.; Paz Rodríguez, J.; Novarro, N. Allogeneic Human Umbilical Cord Mesenchymal Stem Cells for the Treatment of Autism Spectrum Disorder in Children: Safety Profile and Effect on Cytokine Levels. Stem Cells Transl. Med. 2019, 8, 1008–1016. [Google Scholar] [CrossRef]
- Zhang, J.; Lv, S.; Liu, X.; Song, B.; Shi, L. Umbilical Cord Mesenchymal Stem Cell Treatment for Crohn’s Disease: A Randomized Controlled Clinical Trial. Gut Liver 2018, 12, 73–78. [Google Scholar] [CrossRef] [PubMed]
- Sun, Q.; Huang, Z.; Han, F.; Zhao, M.; Cao, R.; Zhao, D.; Hong, L.; Na, N.; Li, H.; Miao, B.; et al. Allogeneic mesenchymal stem cells as induction therapy are safe and feasible in renal allografts: Pilot results of a multicenter randomized controlled trial. J. Transl. Med. 2018, 16, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Shi, M.; Liu, Z.; Wang, Y.; Xu, R.; Sun, Y.; Zhang, M.; Yu, X.; Wang, H.; Meng, L.; Su, H.; et al. A Pilot Study of Mesenchymal Stem Cell Therapy for Acute Liver Allograft Rejection. Stem Cells Transl. Med. 2017, 6, 2053–2061. [Google Scholar] [CrossRef]
- Shi, L.; Huang, H.; Lu, X.; Yan, X.; Jiang, X.; Xu, R.; Wang, S.; Zhang, C.; Yuan, X.; Xu, Z.; et al. Effect of human umbilical cord-derived mesenchymal stem cells on lung damage in severe COVID-19 patients: A randomized, double-blind, placebo-controlled phase 2 trial. Signal Transduct. Target. Ther. 2021, 6, 1–9. [Google Scholar] [CrossRef]
- Meng, F.; Xu, R.; Wang, S.; Xu, Z.; Zhang, C.; Li, Y.; Yang, T.; Shi, L.; Fu, J.; Jiang, T.; et al. Human umbilical cord-derived mesenchymal stem cell therapy in patients with COVID-19: A phase 1 clinical trial. Signal Transduct. Target. Ther. 2020, 5, 172. [Google Scholar] [CrossRef] [PubMed]
- Shu, L.; Niu, C.; Li, R.; Huang, T.; Wang, Y.; Huang, M.; Ji, N.; Zheng, Y.; Chen, X.; Shi, L.; et al. Treatment of severe COVID-19 with human umbilical cord mesenchymal stem cells. Stem Cell Res. Ther. 2020, 11, 361. [Google Scholar] [CrossRef] [PubMed]
- He, X.; Ai, S.; Guo, W.; Yang, Y.; Wang, Z.; Jiang, D.; Xu, X. Umbilical cord-derived mesenchymal stem (stromal) cells for treatment of severe sepsis: Aphase 1 clinical trial. Transl. Res. 2018, 199, 52–61. [Google Scholar] [CrossRef] [PubMed]
- Squillaro, T.; Antonucci, I.; Alessio, N.; Esposito, A.; Cipollaro, M.; Melone, M.A.B.; Peluso, G.; Stuppia, L.; Galderisi, U. Impact of lysosomal storage disorders on biology of mesenchymal stem cells: Evidences from in vitro silencing of glucocerebrosidase (GBA) and alpha-galactosidase A (GLA) enzymes. J. Cell. Physiol. 2017, 232, 3454–3467. [Google Scholar] [CrossRef] [PubMed]



| Cells | Positive Expression (+) | Negative Expression (−) | References |
|---|---|---|---|
| Amniotic MSCs | CD29, CD44, CD73, CD90, CD105, CD166, SSEA-3/4, CK18, HCAM-1, HLA ABC, Oct-3/4, GATA-4, Rex-1, BMP-4, SCF, NCAM, nestin, HFN-4alpha, CK18, vimentin | CD14, CD34, CD45, TRA-1-60, VCAM-1, PECAM-1, HLA-DR, BMP2, FGF-5, Pax-6, TERT | [199,200,201] |
| Chorionic membrane MSCs | CD13, CD29, CD44, CD54, CD73, CD105, CD166 | CD3, CD14, CD34, CD45, CD31 | [202] |
| Chorionic villi MSCs | CD44, CD73, CD90, CD105, HLA-ABC, Sox-2 | CD45, CD34, CD19, HLA-DR, CD14, CD40, CD56, CD80, CD83, CD86, CD275 | [36,202] |
| Chorionic plate MSCs | CD44, CD73, CD90, CD105, CD166, Oct-4, NANOG, Sox-2 | CD14, CD19, CD34, CD45, HLA-DR | [203,204,205] |
| Decidua MSCs | CD44, CD90, CD105, CD146, CD166, HLA-ABC | CD40, CD80, CD83, CD86, HLA-DR | [193] |
| Umbilical cord MSCs | CD13, CD29, CD44, CD73, CD90, CD105, C10, CD49b-e, CD146, CD166, HLA-ABC, NANOG, Rex-1, Sox-2 Uncertain expression: Oct-3/4, SSEA-3, SSEA-4, STRO-1, TRA-1-60, TRA-1-81 | CD14, CD31, CD34, CD45, CD51/61, CD64, CD106, HLA-DR | [206,207,208,209,210,211] |
| Amniotic fluid MSCs | CD73, CD90, CD105, Oct-4 | CD45, CD34, CD31 | [212] |
| Amniotic fluid stromal cells | CD29, CD44, CD73, CD90, CD105, SSEA-4, Oct-4, MHC-I, NANOG, SSEA-3, TRA-1-60, TRA-1-81 | MHC-II, CD80, CD86 | [167,213] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kulus, M.; Sibiak, R.; Stefańska, K.; Zdun, M.; Wieczorkiewicz, M.; Piotrowska-Kempisty, H.; Jaśkowski, J.M.; Bukowska, D.; Ratajczak, K.; Zabel, M.; et al. Mesenchymal Stem/Stromal Cells Derived from Human and Animal Perinatal Tissues—Origins, Characteristics, Signaling Pathways, and Clinical Trials. Cells 2021, 10, 3278. https://doi.org/10.3390/cells10123278
Kulus M, Sibiak R, Stefańska K, Zdun M, Wieczorkiewicz M, Piotrowska-Kempisty H, Jaśkowski JM, Bukowska D, Ratajczak K, Zabel M, et al. Mesenchymal Stem/Stromal Cells Derived from Human and Animal Perinatal Tissues—Origins, Characteristics, Signaling Pathways, and Clinical Trials. Cells. 2021; 10(12):3278. https://doi.org/10.3390/cells10123278
Chicago/Turabian StyleKulus, Magdalena, Rafał Sibiak, Katarzyna Stefańska, Maciej Zdun, Maria Wieczorkiewicz, Hanna Piotrowska-Kempisty, Jędrzej M. Jaśkowski, Dorota Bukowska, Kornel Ratajczak, Maciej Zabel, and et al. 2021. "Mesenchymal Stem/Stromal Cells Derived from Human and Animal Perinatal Tissues—Origins, Characteristics, Signaling Pathways, and Clinical Trials" Cells 10, no. 12: 3278. https://doi.org/10.3390/cells10123278
APA StyleKulus, M., Sibiak, R., Stefańska, K., Zdun, M., Wieczorkiewicz, M., Piotrowska-Kempisty, H., Jaśkowski, J. M., Bukowska, D., Ratajczak, K., Zabel, M., Mozdziak, P., & Kempisty, B. (2021). Mesenchymal Stem/Stromal Cells Derived from Human and Animal Perinatal Tissues—Origins, Characteristics, Signaling Pathways, and Clinical Trials. Cells, 10(12), 3278. https://doi.org/10.3390/cells10123278