
Retinal Protection from LED-Backlit Screen Lights by Short Wavelength Absorption Filters

 , , , ,
, , , ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Rearing Conditions
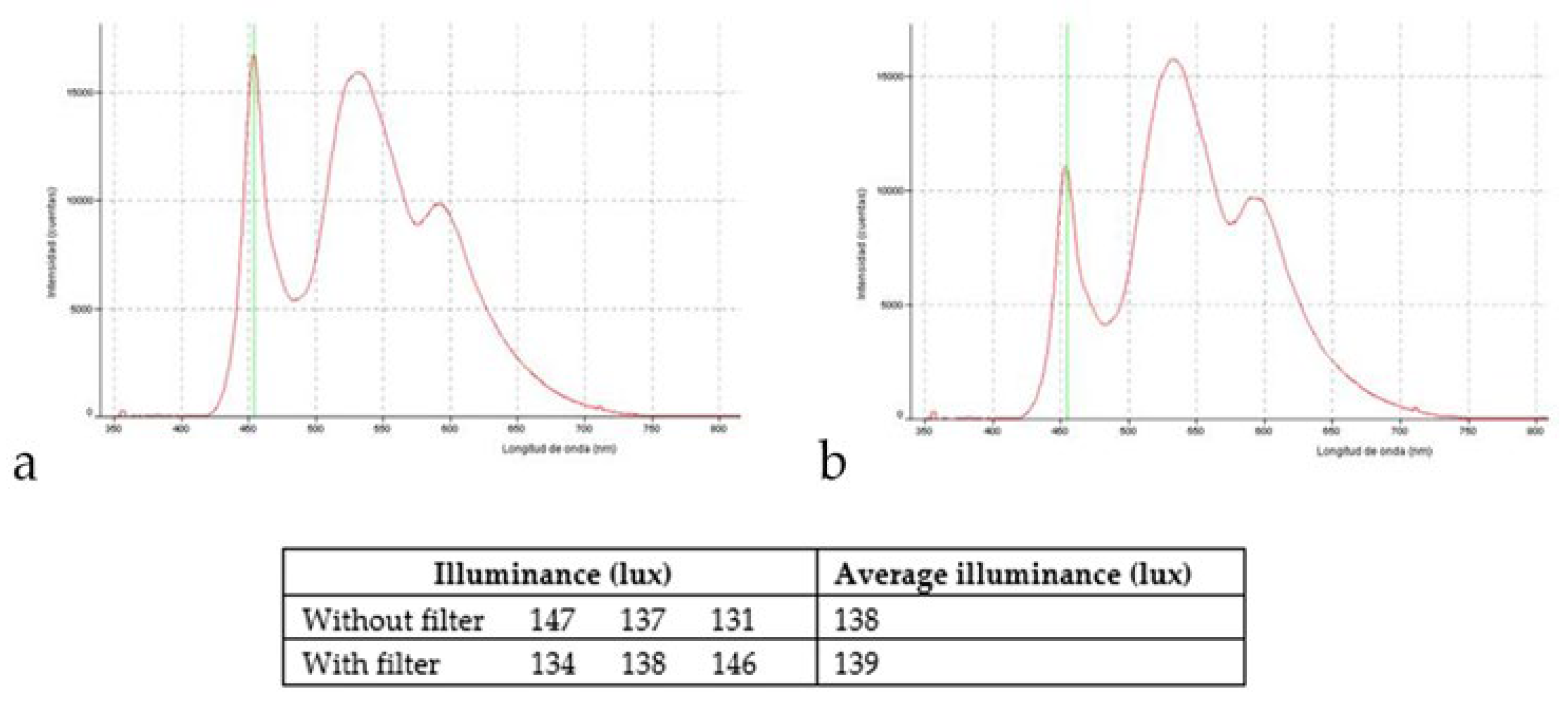
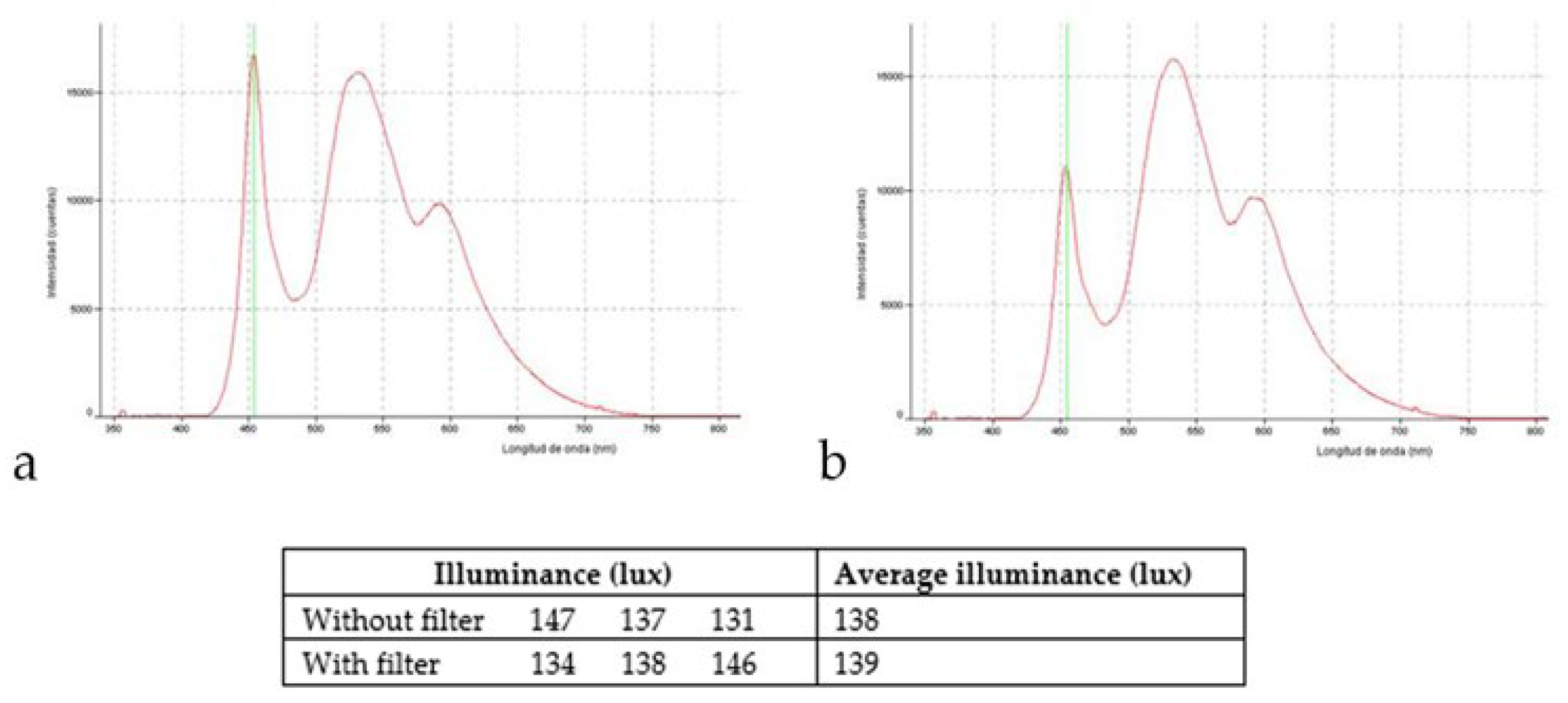
2.2. Light Source and Exposure
2.3. Tissue Collection, RNA Extraction, and qPCR
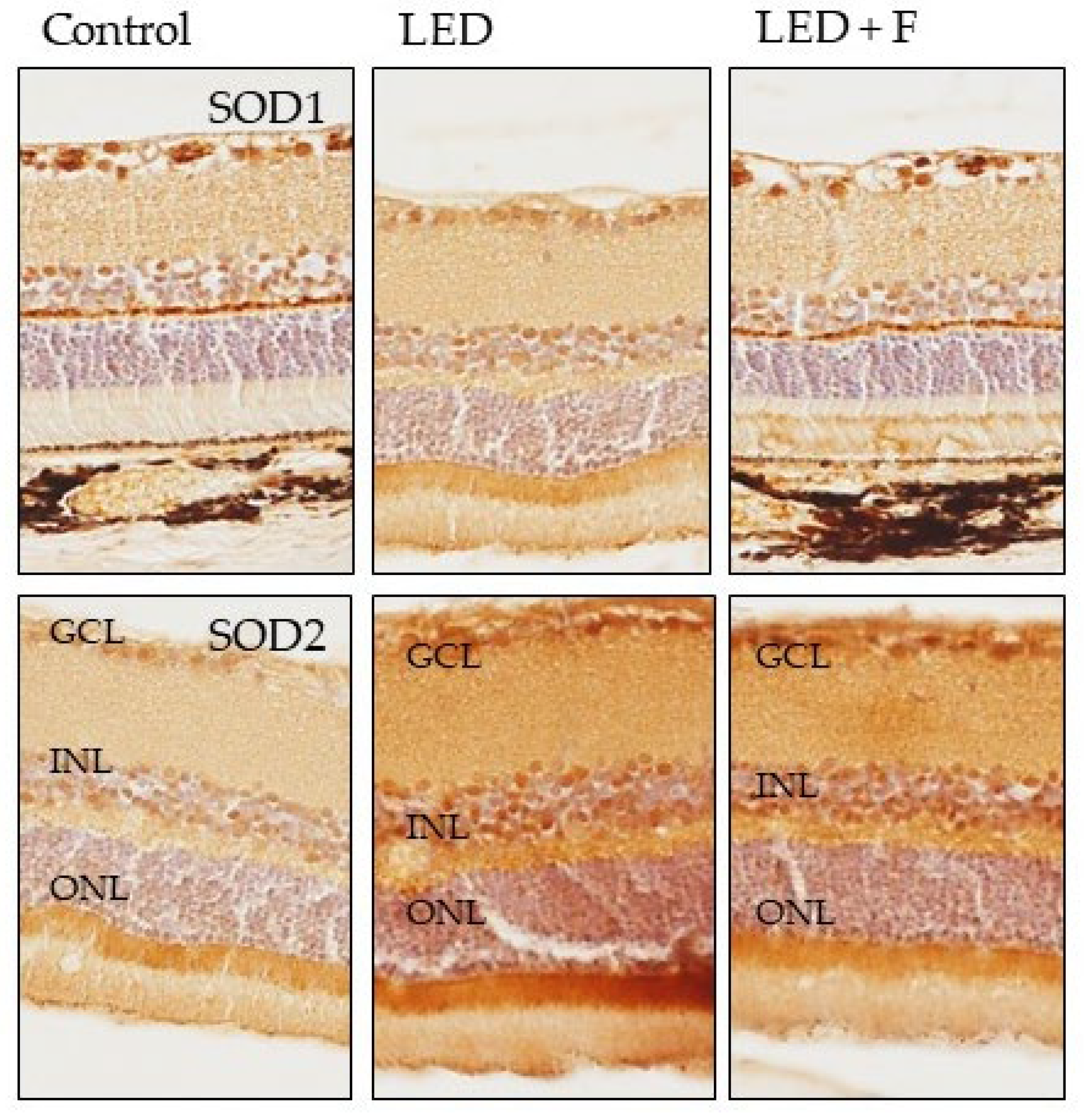
2.4. Structural and Immunohistochemical Studies
2.5. Statistical Analysis
3. Results
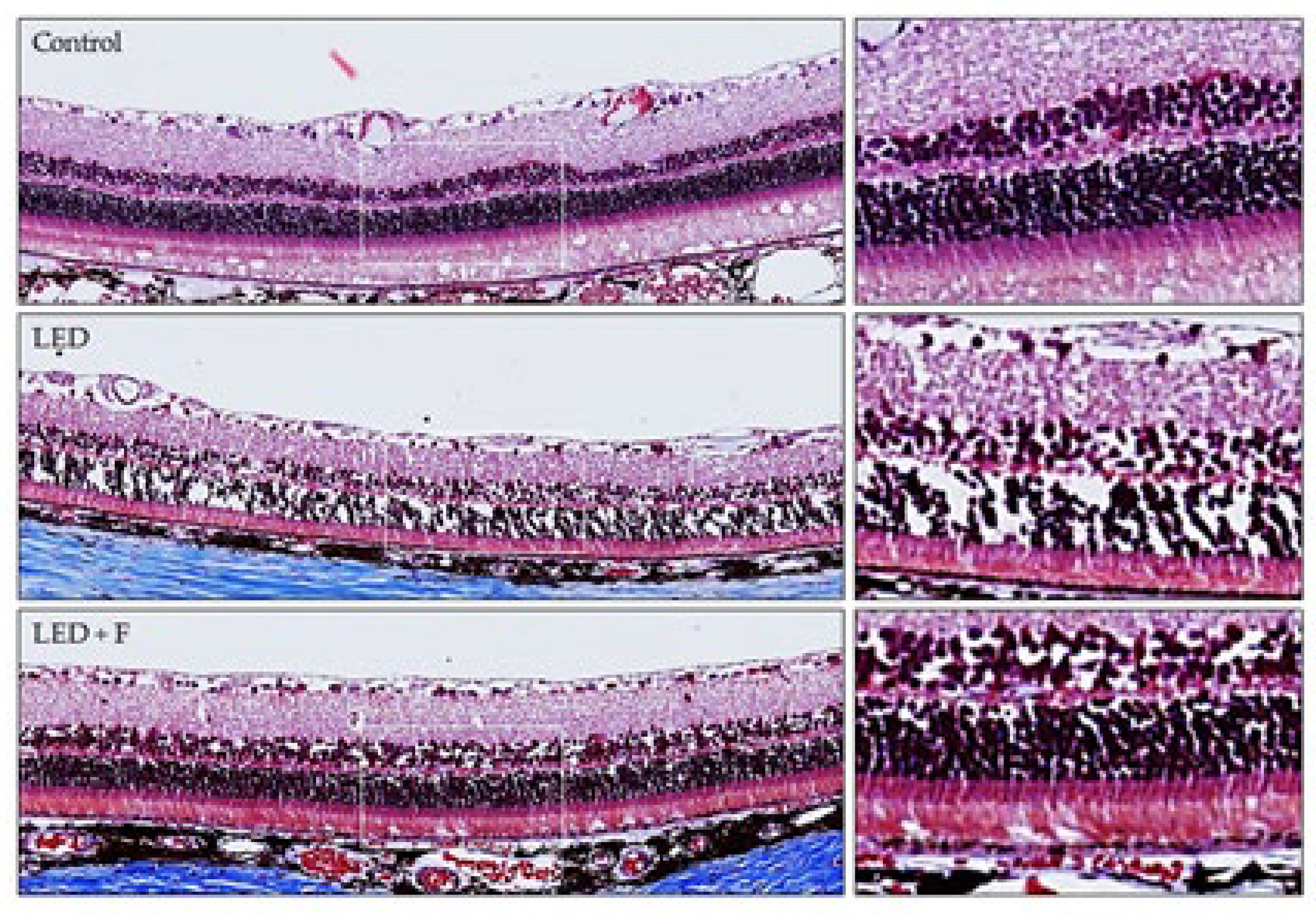
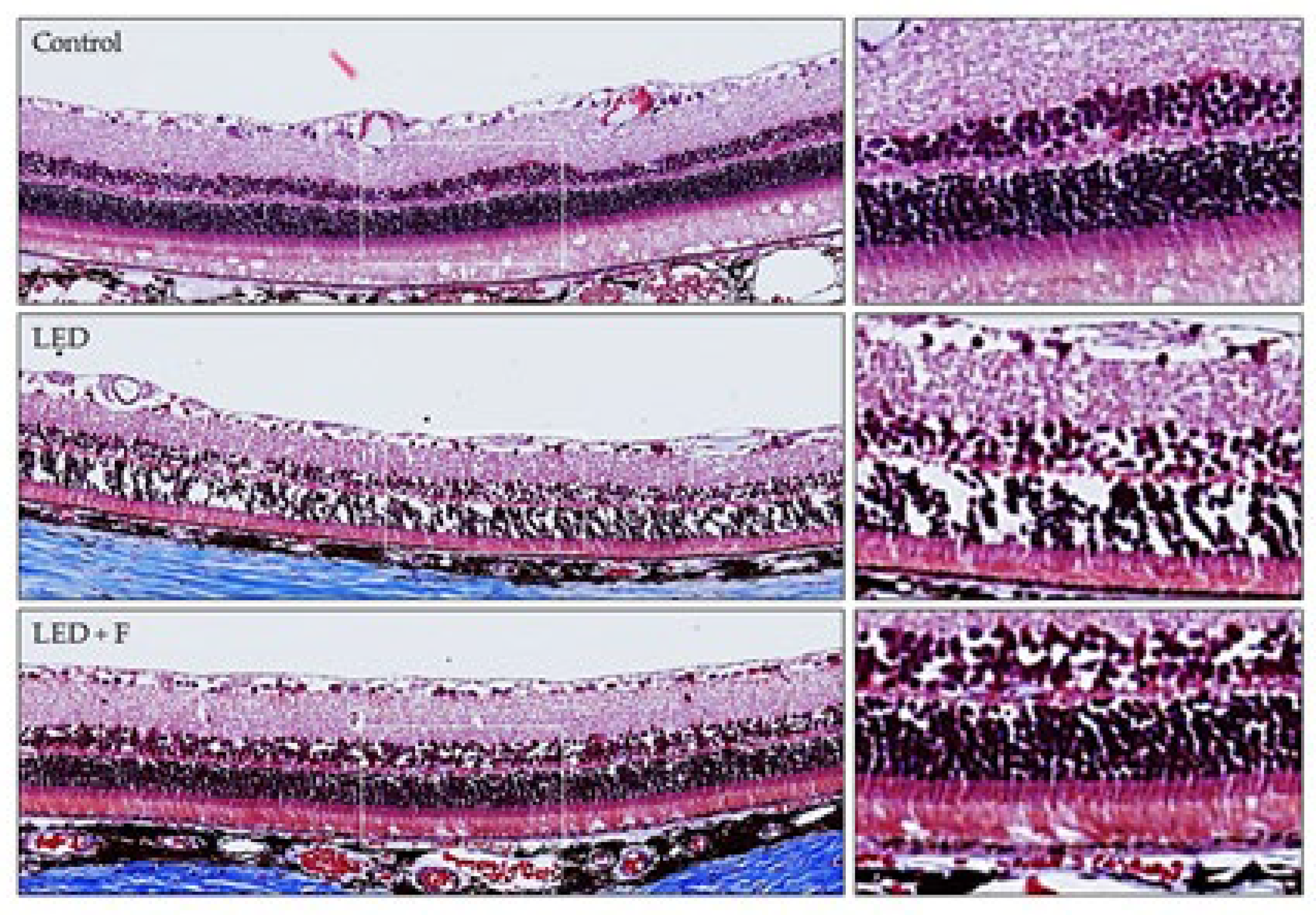
3.1. Structural Study
3.2. Gene Expression Study
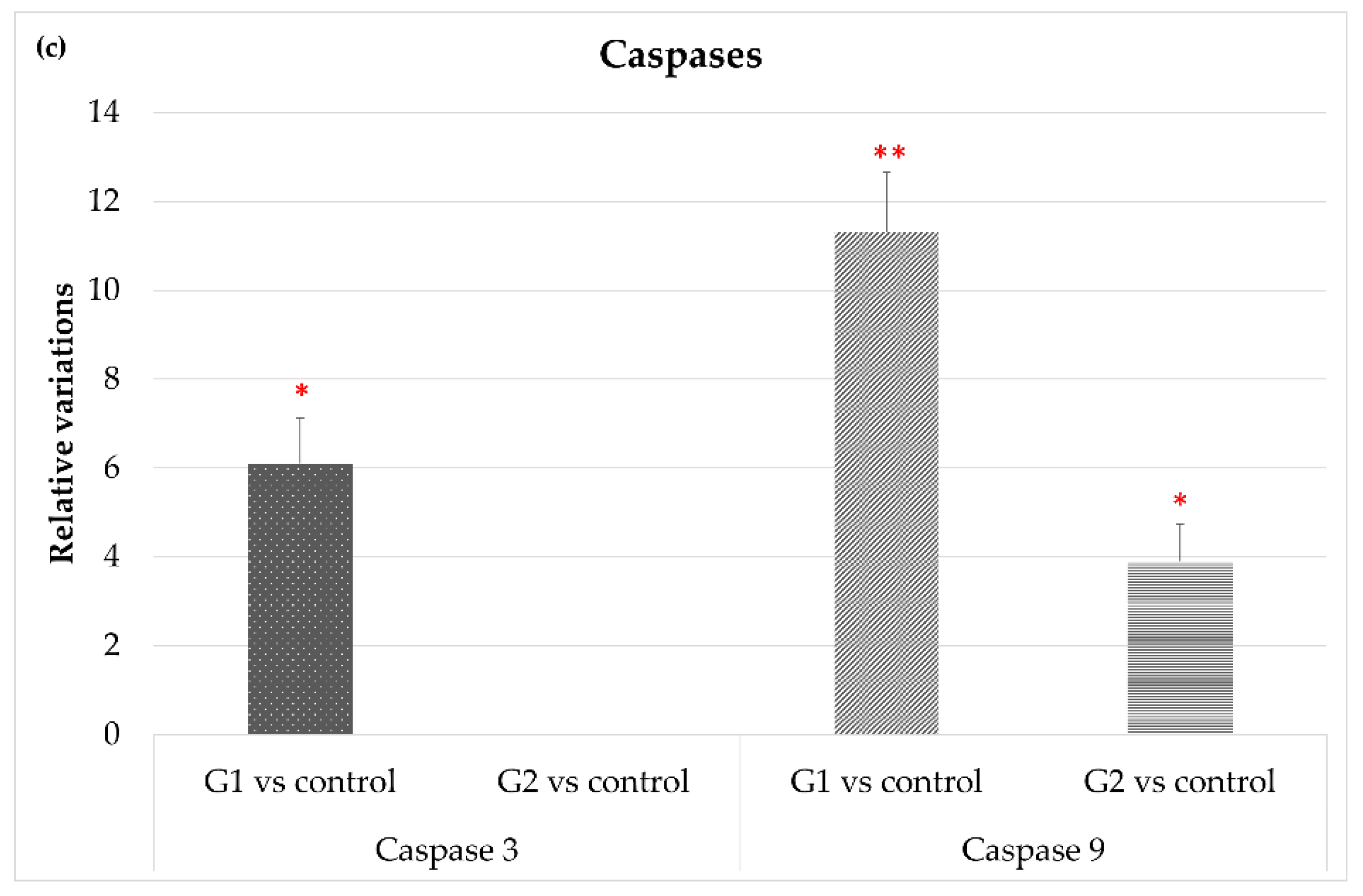
3.2.1. Life/Death Cell-Related Genes
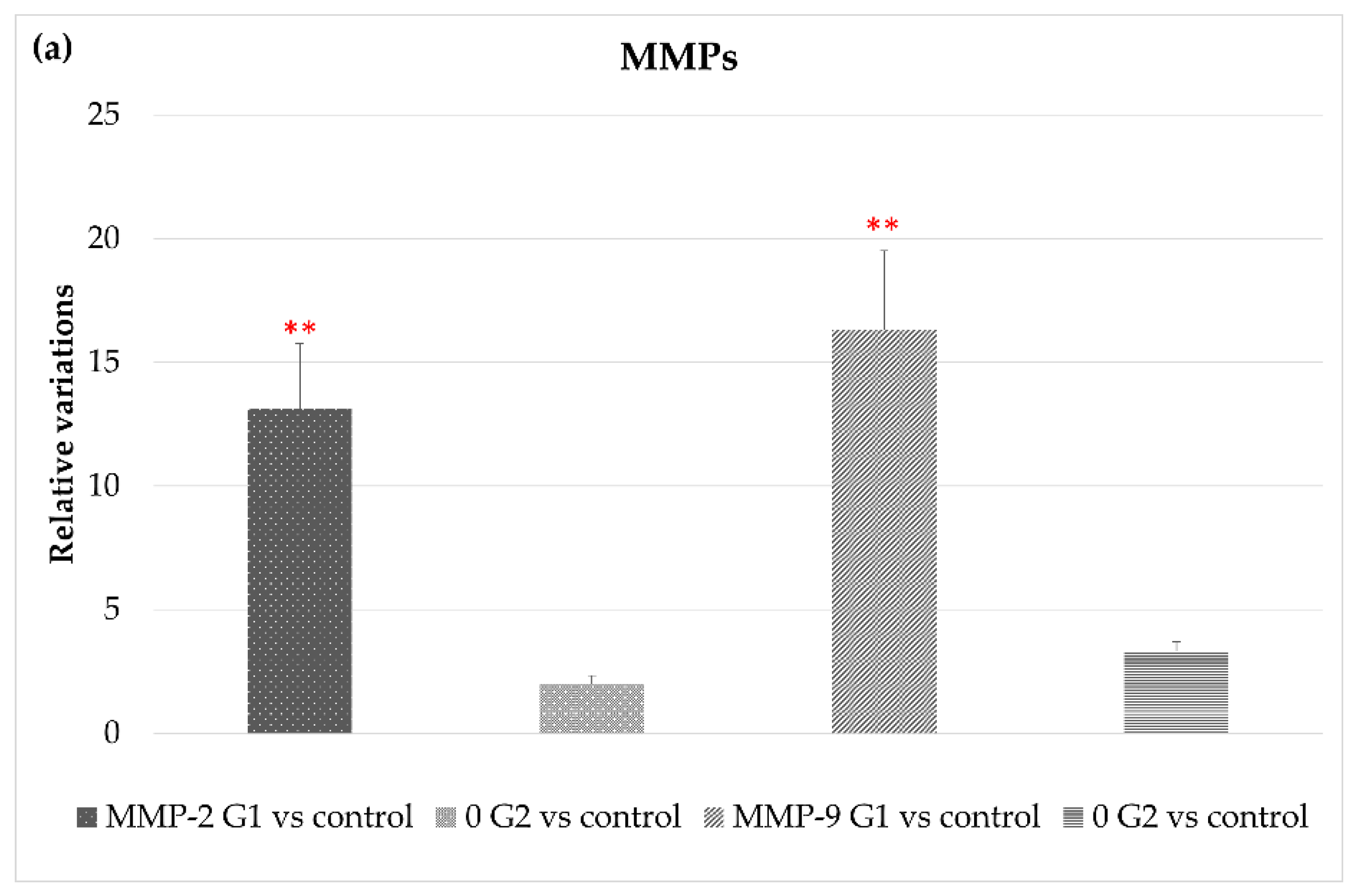
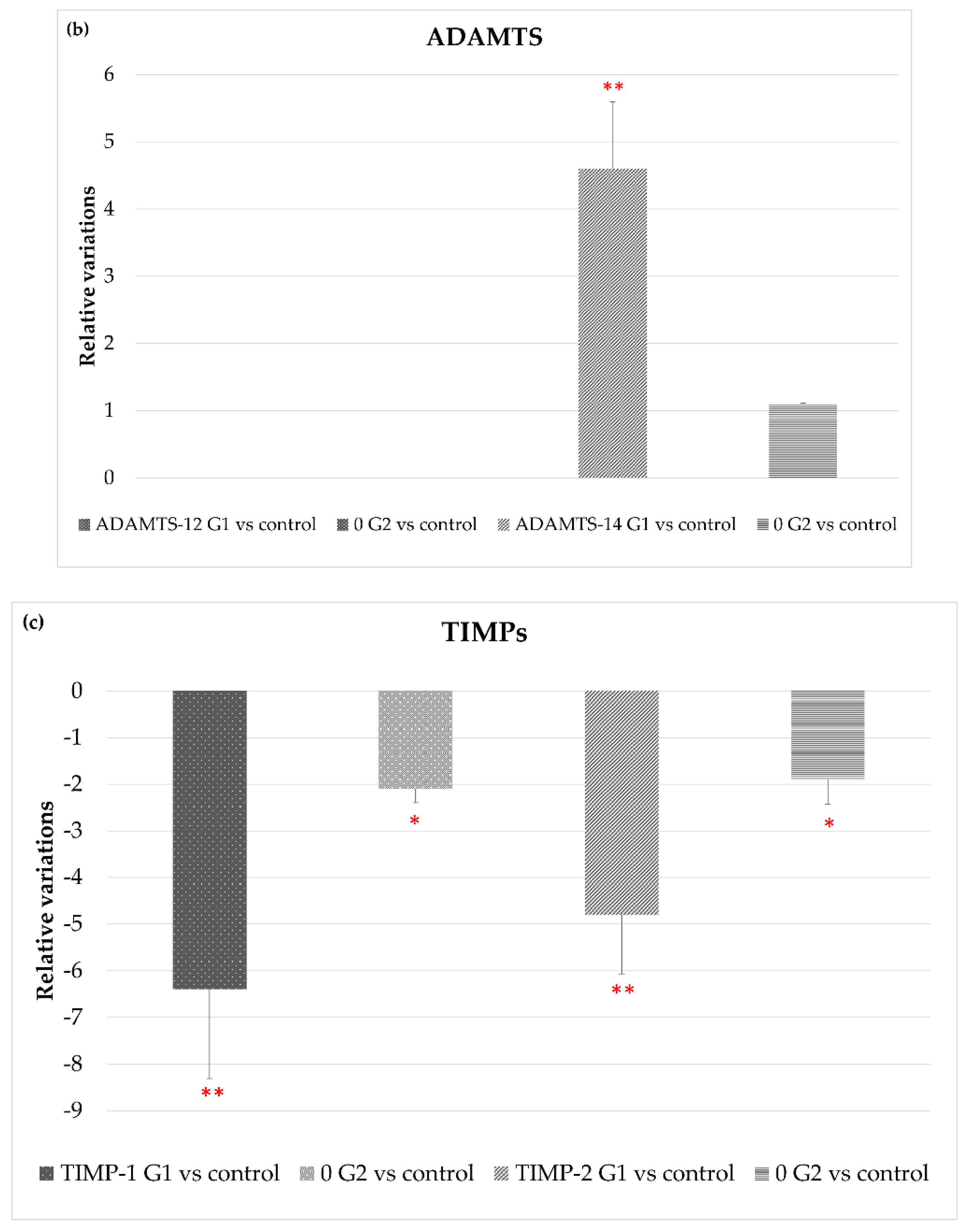
3.2.2. Extracellular Matrix Degradation: MMP/TIMPs System and ADAMTS Genes
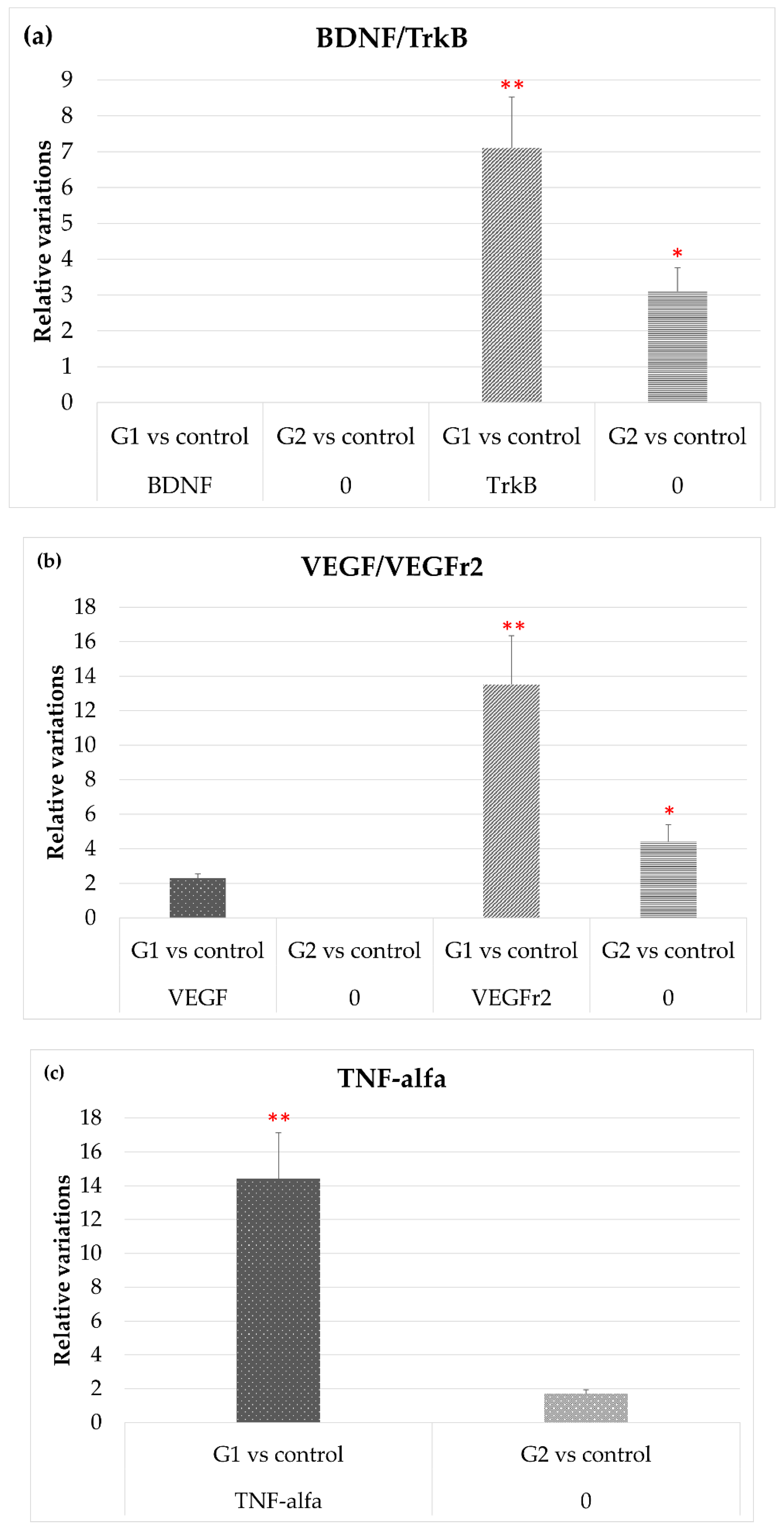
3.2.3. BDNF/TrkB and VEGF/VEGFR-2 System, and TNF-α
3.2.4. Oxidative Stress
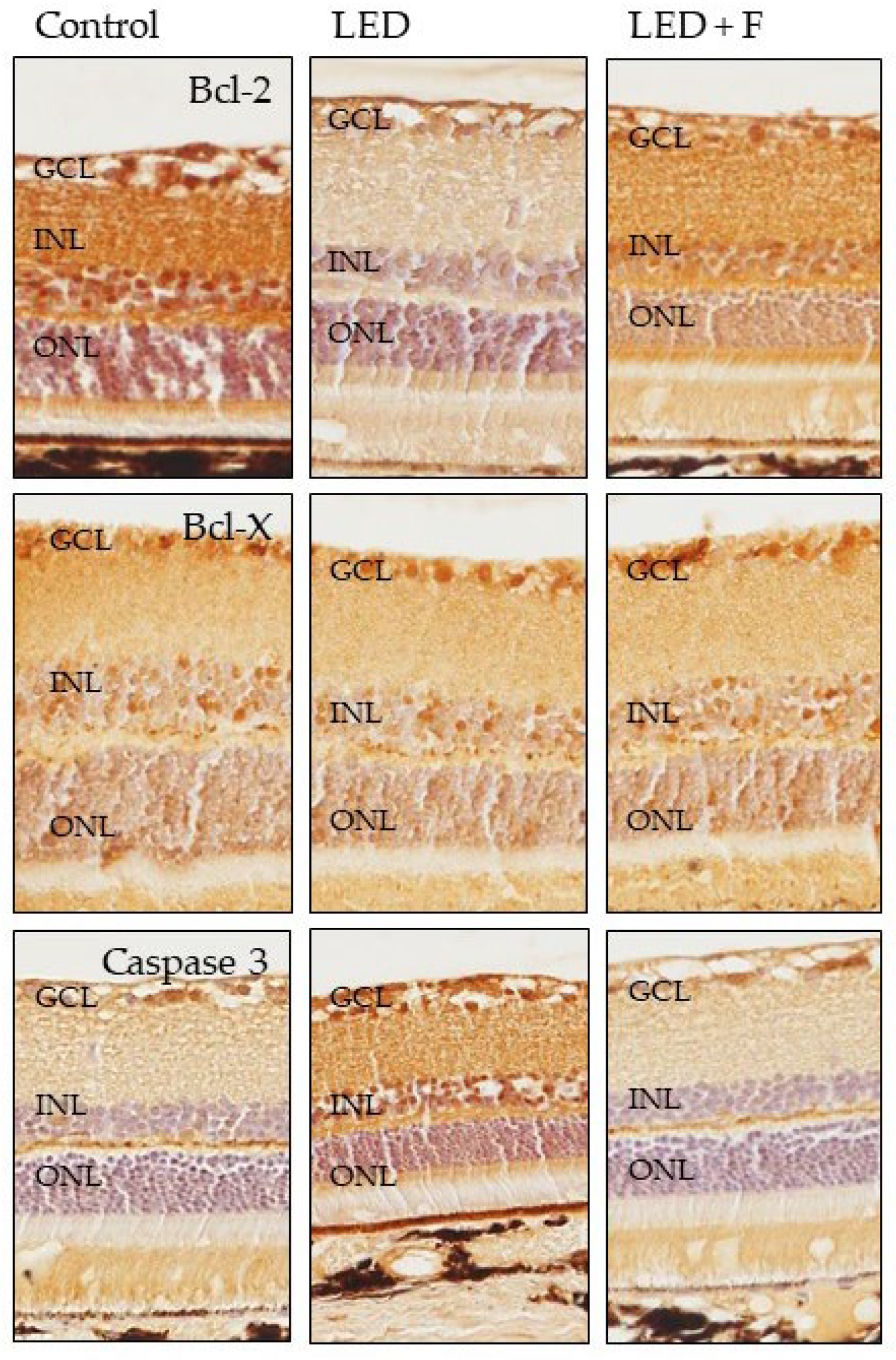
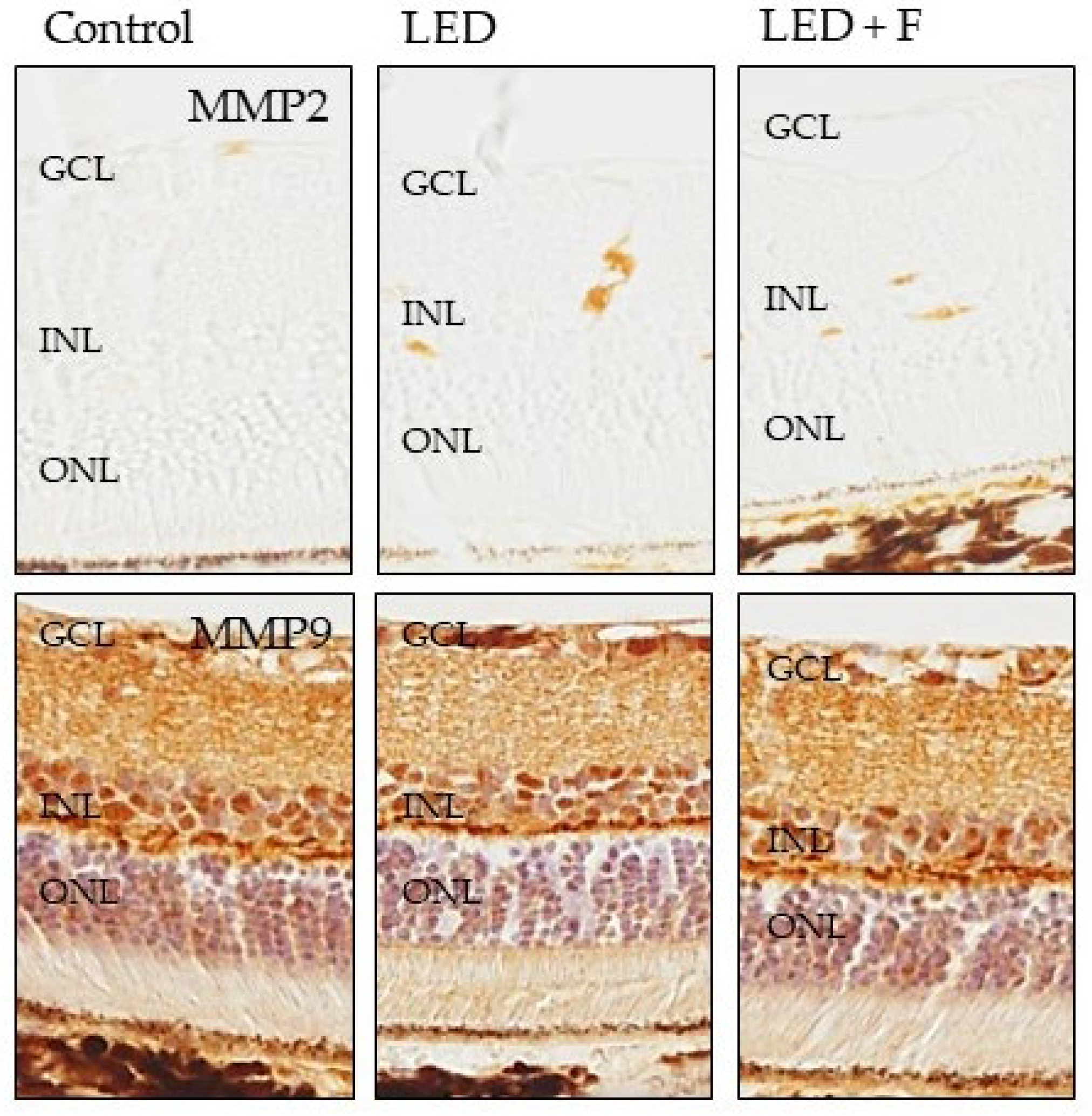
3.3. Immunohistochemistry
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Noell, W.K.; Walker, V.S.; Kang, B.S.; Berman, S. Retinal damage by light in rats. Investig. Ophthalmol. 1966, 5, 450–473. [Google Scholar]
- Jaadane, I.; Boulenguez, P.; Chahory, S.; Carré, S.; Savoldelli, M.; Jonet, L.; Behar-Cohen, F.; Martinsons, C.; Torriglia, A. Retinal damage induced by commercial light emitting diodes (LEDs). Free Radic. Biol. Med. 2015, 84, 373–384. [Google Scholar] [CrossRef] [Green Version]
- Chamorro, E.; Bonnin-Arias, C.; Pérez-Carrasco, M.J.; Muñoz de Luna, J.; Vázquez, D.; Sánchez-Ramos, C. Effects of light-emitting diode radiations on human retinal pigment epithelial cells in vitro. Photochem. Photobiol. 2013, 89, 468–473. [Google Scholar] [CrossRef]
- Roberts, J.E. Ultraviolet radiation as a risk factor for cataract and macular degeneration. Eye Contact Lens 2011, 37, 246–249. [Google Scholar] [CrossRef] [PubMed]
- Wielgus, A.R.; Roberts, J.E. Retinal photodamage by endogenous and xenobiotic agents. Photochem. Photobiol. 2012, 88, 1320–1345. [Google Scholar] [CrossRef]
- Mohan, K.; Kecova, H.; Hernandez-Merino, E.; Kardon, R.H.; Harper, M.M. Retinal ganglion cell damage in an experimental rodent model of blast-mediated traumatic brain injury. Investig. Ophthalmol. Vis. Sci. 2013, 54, 3440–3450. [Google Scholar] [CrossRef] [Green Version]
- Nakamura, M.; Yako, T.; Kuse, Y.; Inoue, Y.; Nishinaka, A.; Nakamura, S.; Shimazawa, M.; Hara, H. Exposure to excessive blue LED light damages retinal pigment epithelium and photoreceptors of pigmented mice. Exp. Eye Res. 2018, 177, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Shang, Y.-M.; Wang, G.-S.; Sliney, D.H.; Yang, C.-H.; Lee, L.-L. Light-emitting-diode induced retinal damage and its wavelength dependency in vivo. Int. J. Ophthalmol. 2017, 10, 191–202. [Google Scholar] [CrossRef]
- Shang, Y.-M.; Wang, G.-S.; Sliney, D.; Yang, C.-H.; Lee, L.-L. White light-emitting diodes (LEDs) at domestic lighting levels and retinal injury in a rat model. Environ. Health Perspect. 2014, 122, 269–276. [Google Scholar] [CrossRef] [Green Version]
- Zhang, P.; Huang, C.; Wang, W.; Wang, M. Early changes in staurosporine-induced differentiated RGC-5 cells indicate cellular injury response to nonlethal blue light exposure. Photochem. Photobiol. Sci. Off. J. Eur. Photochem. Assoc. Eur. Soc. Photobiol. 2015, 14, 1093–1099. [Google Scholar] [CrossRef]
- Kuse, Y.; Ogawa, K.; Tsuruma, K.; Shimazawa, M.; Hara, H. Damage of photoreceptor-derived cells in culture induced by light emitting diode-derived blue light. Sci. Rep. 2014, 4, 5223. [Google Scholar] [CrossRef] [Green Version]
- Behar-Cohen, F.; Martinsons, C.; Viénot, F.; Zissis, G.; Barlier-Salsi, A.; Cesarini, J.P.; Enouf, O.; Garcia, M.; Picaud, S.; Attia, D. Light-emitting diodes (LED) for domestic lighting: Any risks for the eye? Prog. Retin. Eye Res. 2011, 30, 239–257. [Google Scholar] [CrossRef] [PubMed]
- Martinsons, C. Potential Health Issues of Solid-State Lighting; IEA 4E Final Report; IEA: Paris, France, 2014. [Google Scholar]
- Contín, M.A.; Arietti, M.M.; Benedetto, M.M.; Bussi, C.; Guido, M.E. Photoreceptor damage induced by low-intensity light: Model of retinal degeneration in mammals. Mol. Vis. 2013, 19, 1614–1625. [Google Scholar]
- Krigel, A.; Berdugo, M.; Picard, E.; Levy-Boukris, R.; Jaadane, I.; Jonet, L.; Dernigoghossian, M.; Andrieu-Soler, C.; Torriglia, A.; Behar-Cohen, F. Light-induced retinal damage using different light sources, protocols and rat strains reveals LED phototoxicity. Neuroscience 2016, 339, 296–307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Landry, R.J.; James, R.H.; Miller, S.A.; Ilev, I.K. Evaluation of Potential Optical Radiation Hazards from LED Flashlights. Health Phys. 2021, 120, 56–61. [Google Scholar] [CrossRef]
- Tao, J.-X.; Zhou, W.-C.; Zhu, X.-G. Mitochondria as potential targets and initiators of the blue light hazard to the retina. Oxid. Med. Cell. Longev. 2019, 2019, 6435364. [Google Scholar] [CrossRef] [Green Version]
- Touitou, Y.; Point, S. Effects and mechanisms of action of light-emitting diodes on the human retina and internal clock. Environ. Res. 2020, 190, 109942. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.-H.; Wu, M.-R.; Li, C.-H.; Cheng, H.-W.; Huang, S.-H.; Tsai, C.-H.; Lin, F.-L.; Ho, J.-D.; Kang, J.-J.; Hsiao, G.; et al. Editor’s Highlight: Periodic Exposure to Smartphone-Mimic Low-Luminance Blue Light Induces Retina Damage Through Bcl-2/BAX-Dependent Apoptosis. Toxicol. Sci. 2017, 157, 196–210. [Google Scholar] [CrossRef]
- Jaadane, I.; Villalpando Rodriguez, G.E.; Boulenguez, P.; Chahory, S.; Carré, S.; Savoldelli, M.; Jonet, L.; Behar-Cohen, F.; Martinsons, C.; Torriglia, A. Effects of white light-emitting diode (LED) exposure on retinal pigment epithelium in vivo. J. Cell. Mol. Med. 2017, 21, 3453–3466. [Google Scholar] [CrossRef] [Green Version]
- Xie, C.; Zhu, H.; Chen, S.; Wen, Y.; Jin, L.; Zhang, L.; Tong, J.; Shen, Y. Chronic retinal injury induced by white LED light with different correlated color temperatures as determined by microarray analyses of genome-wide expression patterns in mice. J. Photochem. Photobiol. B Biol. 2020, 210, 111977. [Google Scholar] [CrossRef]
- Song, J.; Li, D.; Shan, Z.; Kurskaya, O.; Sharshov, K.; Gao, T.; Bi, H. Photocytotoxicity of white light-emitting diode irradiation on human lens epithelium and retinal pigment epithelium via the JNK and p38 MAPK signaling pathways. J. Photochem. Photobiol. B Biol. 2020, 213, 112058. [Google Scholar] [CrossRef]
- Benedetto, M.M.; Contin, M.A. Oxidative stress in retinal degeneration promoted by constant LED light. Front. Cell. Neurosci. 2019, 13, 139. [Google Scholar] [CrossRef]
- Otsu, W.; Ishida, K.; Nakamura, S.; Shimazawa, M.; Tsusaki, H.; Hara, H. Blue light-emitting diode irradiation promotes transcription factor EB-mediated lysosome biogenesis and lysosomal cell death in murine photoreceptor-derived cells. Biochem. Biophys. Res. Commun. 2020, 526, 479–484. [Google Scholar] [CrossRef]
- Liu, X.; Zhou, Q.; Lin, H.; Wu, J.; Wu, Z.; Qu, S.; Bi, Y. The Protective Effects of Blue Light-Blocking Films With Different Shielding Rates: A Rat Model Study. Transl. Vis. Sci. Technol. 2019, 8, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, S.-I.; Jang, Y.P. The Protective Effect of Brown-, Gray-, and Blue-Tinted Lenses against Blue LED Light-Induced Cell Death in A2E-Laden Human Retinal Pigment Epithelial Cells. Ophthalmic Res. 2017, 57, 118–124. [Google Scholar] [CrossRef]
- Sanchez-Ramos, C.; Vega, J.A.; del Valle, M.E.; Fernandez-Balbuena, A.; Bonnin-Arias, C.; Benitez-del Castillo, J.M. Role of metalloproteases in retinal degeneration induced by violet and blue light. Adv. Exp. Med. Biol. 2010, 664, 159–164. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Ramos, C.; Bonnin-Arias, C.; Guerrera, M.C.; Calavia, M.G.; Chamorro, E.; Montalbano, G.; López-Velasco, S.; López-Muñiz, A.; Germanà, A.; Vega, J.A. Light regulates the expression of the BDNF/TrkB system in the adult zebrafish retina. Microsc. Res. Tech. 2013, 76, 42–49. [Google Scholar] [CrossRef] [PubMed]
- Dawson, W.; Nakanishi-Ueda, T.; Armstrong, D.; Reitze, D.; Samuelson, D.; Hope, M.; Fukuda, S.; Matsuishi, M.; Ozawa, T.; Ueda, T.; et al. Local fundus response to blue (LED and laser) and infrared (LED and laser) sources. Exp. Eye Res. 2001, 73, 137–147. [Google Scholar] [CrossRef]
- Ueda, T.; Nakanishi-Ueda, T.; Yasuhara, H.; Koide, R.; Dawson, W.W. Eye damage control by reduced blue illumination. Exp. Eye Res. 2009, 89, 863–868. [Google Scholar] [CrossRef]
- Polosa, A.; Bessaklia, H.; Lachapelle, P. Strain Differences in Light-Induced Retinopathy. PLoS ONE 2016, 11, e0158082. [Google Scholar] [CrossRef] [Green Version]
- Sparrow, J.R.; Miller, A.S.; Zhou, J. Blue light-absorbing intraocular lens and retinal pigment epithelium protection in vitro. J. Cataract Refract. Surg. 2004, 30, 873–878. [Google Scholar] [CrossRef] [PubMed]
- Nagai, H.; Hirano, Y.; Yasukawa, T.; Morita, H.; Nozaki, M.; Wolf-Schnurrbusch, U.; Wolf, S.; Ogura, Y. Prevention of increased abnormal fundus autofluorescence with blue light-filtering intraocular lenses. J. Cataract Refract. Surg. 2015, 41, 1855–1859. [Google Scholar] [CrossRef]
- Singh, M.; Tyagi, S.C. Metalloproteinases as mediators of inflammation and the eyes: Molecular genetic underpinnings governing ocular pathophysiology. Int. J. Ophthalmol. 2017, 10, 1308–1318. [Google Scholar] [CrossRef]
- Plantner, J.J.; Smine, A.; Quinn, T.A. Matrix metalloproteinases and metalloproteinase inhibitors in human interphotoreceptor matrix and vitreous. Curr. Eye Res. 1998, 17, 132–140. [Google Scholar] [CrossRef] [PubMed]
- Papp, A.M.; Nyilas, R.; Szepesi, Z.; Lorincz, M.L.; Takács, E.; Abrahám, I.; Szilágyi, N.; Tóth, J.; Medveczky, P.; Szilágyi, L.; et al. Visible light induces matrix metalloproteinase-9 expression in rat eye. J. Neurochem. 2007, 103, 2224–2233. [Google Scholar] [CrossRef] [PubMed]
- Elliot, S.; Catanuto, P.; Stetler-Stevenson, W.; Cousins, S.W. Retinal pigment epithelium protection from oxidant-mediated loss of MMP-2 activation requires both MMP-14 and TIMP-2. Investig. Ophthalmol. Vis. Sci. 2006, 47, 1696–1702. [Google Scholar] [CrossRef] [PubMed]
- Wen, R.; Cheng, T.; Song, Y.; Matthes, M.T.; Yasumura, D.; LaVail, M.M.; Steinberg, R.H. Continuous exposure to bright light upregulates bFGF and CNTF expression in the rat retina. Curr. Eye Res. 1998, 17, 494–500. [Google Scholar] [CrossRef]
- Asai, N.; Abe, T.; Saito, T.; Sato, H.; Ishiguro, S.-I.; Nishida, K. Temporal and spatial differences in expression of TrkB isoforms in rat retina during constant light exposure. Exp. Eye Res. 2007, 85, 346–355. [Google Scholar] [CrossRef]
- Nikoletopoulou, V.; Lickert, H.; Frade, J.M.; Rencurel, C.; Giallonardo, P.; Zhang, L.; Bibel, M.; Barde, Y.-A. Neurotrophin receptors TrkA and TrkC cause neuronal death whereas TrkB does not. Nature 2010, 467, 59–63. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.; Seregard, S.; Algvere, P. V Photochemical damage of the retina. Surv. Ophthalmol. 2006, 51, 461–481. [Google Scholar] [CrossRef]
- Cachafeiro, M.; Bemelmans, A.P.; Samardzija, M.; Afanasieva, T.; Pournaras, J.A.; Grimm, C.; Kostic, C.; Philippe, S.; Wenzel, A.; Arsenijevic, Y. Hyperactivation of retina by light in mice leads to photoreceptor cell death mediated by VEGF and retinal pigment epithelium permeability. Cell Death Dis. 2013, 4, e781. [Google Scholar] [CrossRef]
- Algvere, P.V.; Kvanta, A.; Seregard, S. Drusen maculopathy: A risk factor for visual deterioration. Acta Ophthalmol. 2016, 94, 427–433. [Google Scholar] [CrossRef] [PubMed]
- Yanagi, Y.; Inoue, Y.; Iriyama, A.; Jang, W.-D. Effects of yellow intraocular lenses on light-induced upregulation of vascular endothelial growth factor. J. Cataract Refract. Surg. 2006, 32, 1540–1544. [Google Scholar] [CrossRef] [PubMed]
- Hui, S.; Yi, L.; Fengling, Q.L. Effects of light exposure and use of intraocular lens on retinal pigment epithelial cells in vitro. Photochem. Photobiol. 2009, 85, 966–969. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Sparrow, J.R. Light filtering in a retinal pigment epithelial cell culture model. Optom. Vis. Sci. Off. Publ. Am. Acad. Optom. 2011, 88, 759–765. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Accession Number | Forward | Reverse | |
|---|---|---|---|
| Life/Death | |||
| Bcl-2 | NM_016993 | 5′-ATCTTCTCCTTCCAGCCTGA3′ | 5′-CTGGACATCTCTGCAAAGTC3′ |
| Bcl-xl | BC094213 | 5′-AAGAAACTGAACCAGAAAGG3′ | 5′-TAGATCACTGAATGCTCTCC3′ |
| Bax | NM_017059 | 5′-CTACAGGGTTTCATCCAGGA3′ | 5′-ATCCACATCAGCAATCATCC3′ |
| Bak | AF259504 | 5′-CTATTTAAGAGCGGCATCAG3′ | 5′-ATTACCACTACACTCAGGAT3′ |
| Bcl-xs | AF279286 | 5′-CGGAGAGCATTCAGTGAT 3′ | 5′-CCAGCAGAACTACACCAG 3′ |
| Caspase 3 | NM_012922 | 5′-GTCATGGAGATGAAGGAGTA3′ | 5′-AGAGTAAGCATACAGGAAGT3′ |
| Caspase 9 | NM_031632 | 5′-GTCTGTACTCCAGGGAAGAT3′ | 5′-TTAGCAGTCAGGTCGTTCTT3′ |
| ECM degradation | |||
| MMP2 | NM_031054 | 5′-GCAATACCTGAACACTTTCT3′ | 5′-ATCTGATTCTTGTCCCACTT3′ |
| MMP9 | NM_031055 | 5′-GACTACGACACAGACAGAAA3′ | 5′-GAGTAGGACAGAAGCCATAC3′ |
| ADAMTS-12 | NM_001106420 | 5′-CTGCCAGAATACCACATAGT3′ | 5′-TATCTCCTCTCCACGACATA3′ |
| ADAMTS-14 | NM_001107636 | 5′-GCTACCTCCTATCCTACAAT3′ | 5′-CTTGGTCTTGCAGAAGTATG3′ |
| TIMP1 | U06179 | 5′-CCACCTTATACCAGCGTTAT3′ | 5′-CTGGGACTTGTGGACATATC3′ |
| TIMP22 | NM_021989 | 5′-AGATGTTCAAAGGACCTGAC3′ | 5′-CTTCTTCTGGGTGATGCTAA3′ |
| Growth factors | |||
| BDNF | NM_001270630 | 5′-GTGACAACAATGTGACTCCA3′ | 5′-CATTCACGCTCTCCAGAGTC3′ |
| TrkB | AY265419 | 5′-CCAGAGAACATCACCGAAAT3′ | 5′-ATCAGGTCAGACAAGTCAAG3′ |
| VEG | NM_001110333 | 5′-GTATATCTTCAAGCCGTCCT3′ | 5′-CATTCACATCTGCTATGCTG3′ |
| VEGFR2 | NM_013062 | 5′-GGCAAATACAACCCTTCAGA3′ | 5′-CCGATAGAAGCACTTGTAGG3′ |
| TNFα | NM_012675 | 5′-GCTCTTCTGTCTACTGAACT3′ | 5′-CTTTGAGATCCATGCCATTG3′ |
| Oxidative stress | |||
| SOD1 | NM_017050 | 5′-CTTTGAGATCCATGCCATT3′ | 5′-ACACGATCTTCAATGGACAC3′ |
| SOD2 | NM_017051 | 5′-GAGAACCCAAAGGAGAGTTG3′ | 5′-CTGAAGATAGTAAGCGTGCT3′ |
| β-actin | NM_031144 | 5′-ATCGTGCGTGACATTAAAGA3′ | 5′-GATGCCACAGGATTCCATAC3′ |
| Antigen (Clone) | Origin | Dilution | Supplier |
|---|---|---|---|
| Life/Death | |||
| Bcl-2 Cat # PA5-11379 | Rabbit | 1:200 | ThermoFisher 1 |
| Bcl-xl Cat # PA5-17805 | Rabbit | 1:100 | ThermoFisher 1 |
| Caspase 3 Cat # PA5-77887 | Rabbit | 1:400 | ThermoFisher 1 |
| Caspase 9 ab25758 | Rabbit | 1:200 | Abcam 2 |
| ECM degradation | |||
| MMP2 (clone MMP2/2C1) LS-C2814-100 | Mouse | 1:100 | LifeSpan Biosciences 3 |
| MMP9 ab38898 | Rabbit | 1:200 | Abcam 4 |
| Growth factors/Cytokines | |||
| TrkB catalog # sc-12 | Rabbit | 1:100 | Santa Cruz Biotechnol 5 |
| VEGFR2 PA5-16487 | Rabbit | 1:100 | Invitrogen 6 |
| Oxidative stress | |||
| SOD1 Ab13498 | Rabbit | 1:200 | Abcam 4 |
| SOD2 ab13533 | Rabbit | 1:100 | Abcam 4 |
| Gene Expression | Group 1 Average vs. Control | Group 2 Average vs. Control |
|---|---|---|
| Life/Death | ||
| Bcl-2 | −12.6 ± 2.71 | −2.7 ± 0.3 |
| Bcl-xl | −3.4 ± 0.4 | 0.4 ± 0.01 |
| Bax | 5.2 ± 0.86 | 2.2 ± 0.2 |
| Bak | 3.6 ± 0.63 | 0.1 ± 0.0 |
| Bcl-xs | 1623 ± 2.22 | 2.3 ± 0.61 |
| Caspase 3 | 6.1 ± 1.03 | 0.0 ± 0.0 |
| Caspase 9 | 11.3 ± 1.37 | 3.9 ± 0.83 |
| ECM degradation | ||
| MMP2 | 13.1 ± 2.65 | 2 ± 0.33 |
| MMP9 | 16.3 ± 3.21 | 3.3. ± 0.39 |
| ADAMTS-12 | 0.0 ± 0.0 | 0.0 ± 0.0 |
| ADAMTS-14 | 4.6 ± 0.99 | 1.1 ± 0.0 |
| TIMP1 | −6.4 ± 1.91 | −2.1 ± 0.3 |
| TIMP22 | −4.8 ± 1.27 | −1.9 ± 0.52 |
| Growth factors | ||
| BDNF | 0.0 ± 0.0 | 0.0 ± 0.0 |
| TrkB | 7.1 ± 1.42 | 3.1 ± 0.66 |
| VEG | 2.3 ± 0.25 | 0.0 ± 0.0 |
| VEGFR2 | 11.5 ± 2.84 | 4.4 ± 0.98 |
| TNFα | 14.4 ± 2.72 | 1.7 ± 0.23 |
| Oxidative stress | ||
| SOD1 | 24.6 ± 6.33 | 5.3 ± 1.75 |
| SOD2 | 19.3 ± 3.77 | 6.1 ± 1.42 |
| Retinal Layer | Protein | Control | LED | LED + F |
|---|---|---|---|---|
| Photoreceptors | Blc2 | +/++ | −/+ | + |
| BclX | − | − | − | |
| Caspase 3 | + | +/++ | + | |
| MMP9 | − | −/+ | − | |
| TrkB | +/++ | +++ | ++ | |
| VEGFR2 | − | − | − | |
| SOD1 | −/+ | ++ | + | |
| SOD2 | + | +++ | +/++ | |
| ONL | Blc2 | ++ | − | −/+ |
| BclX | +/++ | +/++ | +/++ | |
| Caspase 3 | −/+ | +/++ | −/+ | |
| MMP9 | +/++ | +/++ | +/++ | |
| TrkB | +/++ | +/++ | +/++ | |
| VEGFR2 | + | ++ | ++ | |
| SOD1 | + | + | + | |
| SOD2 | + | ++/+++ | ++ | |
| INL | Blc2 | ++ | − | − |
| BclX | +/++ | +/++ | +/++ | |
| Caspase 3 | − | +/++ | − | |
| MMP9 | ++ | ++/+++ | ++/+++ | |
| TrkB | +/++ | +++ | ++ | |
| VEGFR2 | + | ++ | +/++ | |
| SOD1 | −/+ | −/+ | −/+ | |
| SOD2 | ||||
| Ganglionic cells | Blc2 | +++ | + | ++ |
| BclX | ++ | ++ | ++ | |
| Caspase 3 | +/++ | ++/+++ | −/+ | |
| MMP9 | ++ | +++ | +++ | |
| TrkB | ++/+++ | +++ | +++ | |
| VEGFR2 | ++ | ++ | ++ | |
| SOD1 | ++/+++ | + | ++/+++ | |
| SOD2 | +/+ | +++ | +++ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sanchez-Ramos, C.; Bonnin-Arias, C.; Blázquez-Sánchez, V.; Aguirre-Vilacoro, V.; Cobo, T.; García-Suarez, O.; Perez-Carrasco, M.J.; Alvarez-Peregrina, C.; Vega, J.A. Retinal Protection from LED-Backlit Screen Lights by Short Wavelength Absorption Filters. Cells 2021, 10, 3248. https://doi.org/10.3390/cells10113248
Sanchez-Ramos C, Bonnin-Arias C, Blázquez-Sánchez V, Aguirre-Vilacoro V, Cobo T, García-Suarez O, Perez-Carrasco MJ, Alvarez-Peregrina C, Vega JA. Retinal Protection from LED-Backlit Screen Lights by Short Wavelength Absorption Filters. Cells. 2021; 10(11):3248. https://doi.org/10.3390/cells10113248
Chicago/Turabian StyleSanchez-Ramos, Celia, Cristina Bonnin-Arias, Vanesa Blázquez-Sánchez, Victoria Aguirre-Vilacoro, Teresa Cobo, Olivia García-Suarez, María Jesús Perez-Carrasco, Cristina Alvarez-Peregrina, and José A. Vega. 2021. "Retinal Protection from LED-Backlit Screen Lights by Short Wavelength Absorption Filters" Cells 10, no. 11: 3248. https://doi.org/10.3390/cells10113248
APA StyleSanchez-Ramos, C., Bonnin-Arias, C., Blázquez-Sánchez, V., Aguirre-Vilacoro, V., Cobo, T., García-Suarez, O., Perez-Carrasco, M. J., Alvarez-Peregrina, C., & Vega, J. A. (2021). Retinal Protection from LED-Backlit Screen Lights by Short Wavelength Absorption Filters. Cells, 10(11), 3248. https://doi.org/10.3390/cells10113248