Role of Macroautophagy in Nutrient Homeostasis During Fungal Development and Pathogenesis

Abstract
:1. Introduction
2. Induction and Requirement of Autophagy in Development and/or Pathogenesis of Filamentous Fungi

{kind=link}
| Fungus | Host | Mutants analyzed | Phenotypic defects | Deduced Autophagy Function | References |
|---|---|---|---|---|---|
| Magnaporthe oryzae | Rice,Barley | atg1Δ, atg2Δ, atg4Δ, atg5Δ, atg8Δ, atg9Δ, atg18Δ | Reduced conidiation; non-pathogenic | Glycogen breakdown; nuclear degradation; Turgor; lipid droplet degradation; autophagy cell death | [43,54,55,56,61] |
| Aspergillus oryzae | N.A. | atg1Δ | Reduced conidiation | Nuclear degradation | [44] |
| Ustilago maydis | Corn | atg8Δ | Reduced teliospores production and pathogenicity | Possibly glycogen metabolism | [57] |
| Cryptococcus neoformans | Human | vps34Δ,ATG8-RNAi | Reduced virulence | Likely nutrient homeostasis | [45,46] |
| Fusarium graminearum | Rice, Barley Wheat | atg15Δ,atg8Δ | Reduced conidiation; non-pathogenic | Lipid droplet turnover; likely glycogen breakdown | [48,49] |
| Trichoderma reesei | N.A. | atg5Δ | Reduced conidiation | Not clear | [50] |
| Aspergillus fumigatus | Human Murine | atg1Δ | Reduced conidiation | Nitrogen metabolism; metal metabolism | [51,52] |
| Colletotrichum lindemuthianum | Beans | clk1Δ (atg1Δ) | Reduced pathogenicity | Not clear | [53] |
| Colletotrichum orbiculare | Cucumber | atg8Δ | No appressorium formation (non-pathogenic) | Not clear | [58,59] |
| Sordaria macrospora | N.A. | atg7 RNAi | Aberrant fruit-body formation | Not clear | [47] |
| Candida glabrata | Human | atg17Δ | Reduced survival in host | Likely ROS and / or starvation resistance | [60] |
| Candida albicans | Human | atg9Δ | No defects | Not clear | [62,63] |
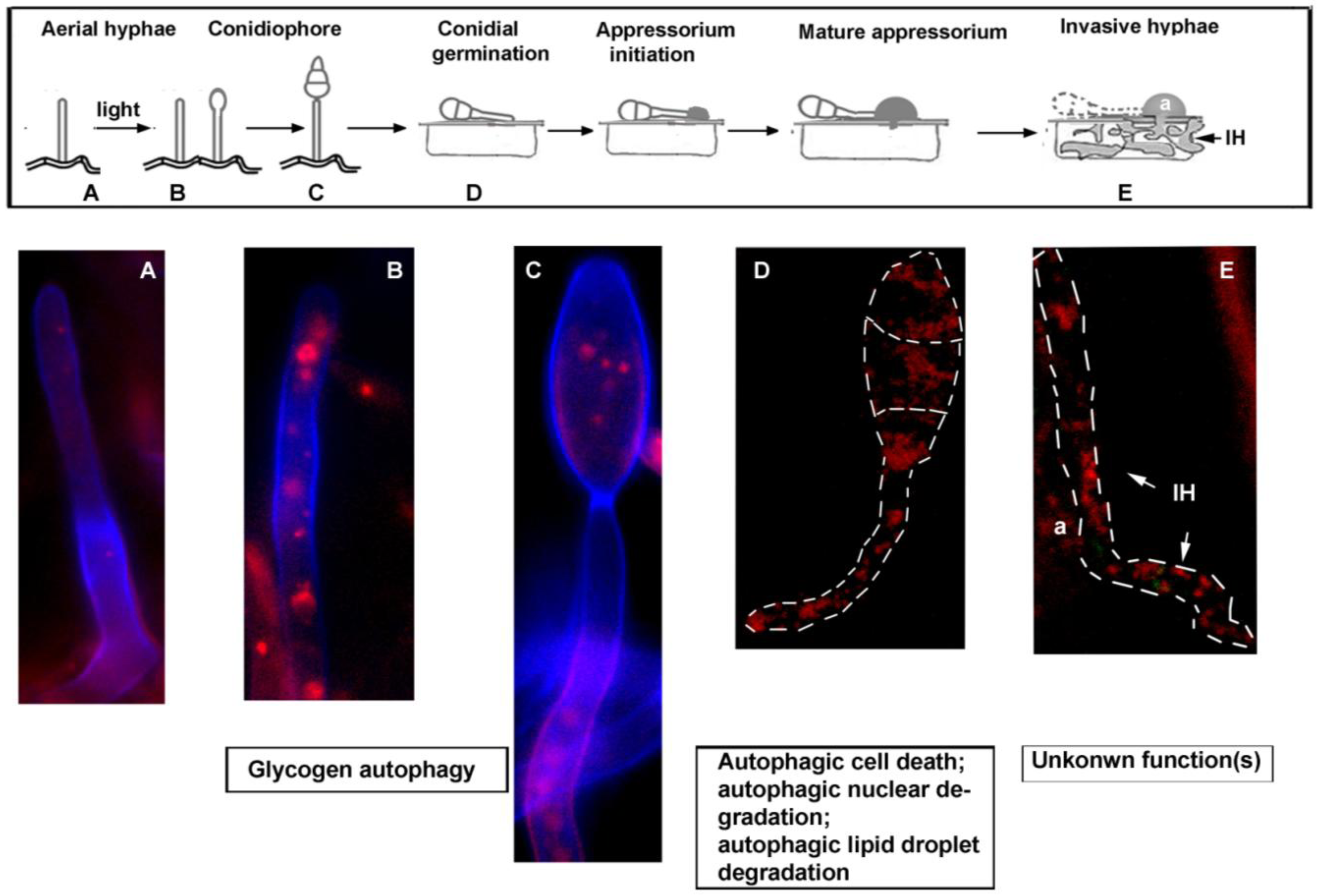
3. Autophagy-Dependent Nutrient Homeostasis in Fungal Sporulation and Pathogenesis
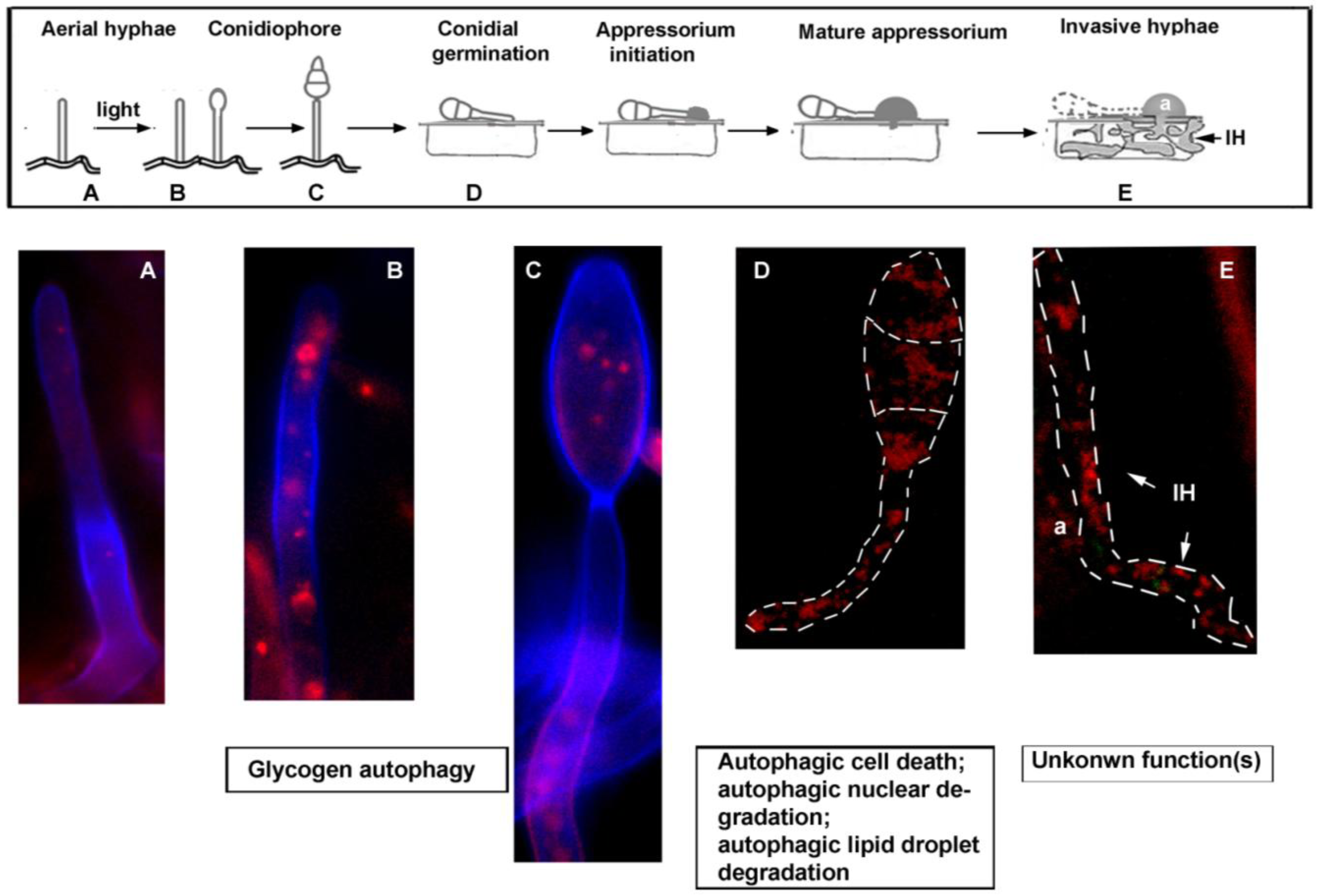
4. Autophagy in Opportunistic Human Fungal Pathogens
5. Host Autophagy as a Defense Mechanism or Facilitator of Infection
6. Conclusions
Acknowledgments
Conflict of Interest
References and Notes
- Nazarko, V.Y.; Nazarko, T.Y.; Farre, J.C.; Stasyk, O.V.; Warnecke, D.; Ulaszewski, S.; Cregg, J.M.; Sibirny, A.A.; Subramani, S. Atg35, a micropexophagy-specific protein that regulates micropexophagic apparatus formation in Pichia pastoris. Autophagy 2011, 7, 375–385. [Google Scholar] [CrossRef]
- Klionsky, D.J.; Cregg, J.M.; Dunn, W.A., Jr.; Emr, S.D.; Sakai, Y.; Sandoval, I.V.; Sibirny, A.; Subramani, S.; Thumm, M.; Veenhuis, M.; et al. A unified nomenclature for yeast autophagy-related genes. Dev. Cell 2003, 5, 539–545. [Google Scholar] [CrossRef]
- Epple, U.D.; Suriapranata, I.; Eskelinen, E.L.; Thumm, M. Aut5/Cvt17p, a putative lipase essential for disintegration of autophagic bodies inside the vacuole. J. Bacteriol. 2001, 183, 5942–5955. [Google Scholar] [CrossRef]
- Klionsky, D.J. The molecular machinery of autophagy: Unanswered questions. J. Cell Sci. 2005, 118, 7–18. [Google Scholar] [CrossRef]
- Kabeya, Y.; Noda, N.N.; Fujioka, Y.; Suzuki, K.; Inagaki, F.; Ohsumi, Y. Characterization of the Atg17-Atg29-Atg31 complex specifically required for starvation-induced autophagy in Saccharomyces cerevisiae. Biochem. Biophys. Res. Commun. 2009, 389, 612–615. [Google Scholar] [CrossRef]
- Cheong, H.; Nair, U.; Geng, J.; Klionsky, D.J. The Atg1 kinase complex is involved in the regulation of protein recruitment to initiate sequestering vesicle formation for nonspecific autophagy in Saccharomyces cerevisiae. Mol. Biol. Cell 2008, 19, 668–681. [Google Scholar]
- Ohsumi, Y. Molecular dissection of autophagy: Two ubiquitin-like systems. Nat. Rev. Mol. Cell Biol. 2001, 2, 211–216. [Google Scholar] [CrossRef]
- Mizushima, N.; Noda, T.; Yoshimori, T.; Tanaka, Y.; Ishii, T.; George, M.D.; Klionsky, D.J.; Ohsumi, M.; Ohsumi, Y.A. Protein conjugation system essential for autophagy. Nature 1998, 395, 395–398. [Google Scholar]
- Mizushima, N.; Noda, T.; Ohsumi, Y. Apg16p is required for the function of the Apg12p-Apg5p conjugate in the yeast autophagy pathway. EMBO J. 1999, 18, 3888–3896. [Google Scholar] [CrossRef]
- Ichimura, Y.; Kirisako, T.; Takao, T.; Satomi, Y.; Shimonishi, Y.; Ishihara, N.; Mizushima, N.; Tanida, I.; Kominami, E.; Ohsumi, M.; et al. A ubiquitin-like system mediates protein lipidation. Nature 2000, 408, 488–492. [Google Scholar]
- Tanida, I.; Mizushima, N.; Kiyooka, M.; Ohsumi, M.; Ueno, T.; Ohsumi, Y.; Kominami, E. Apg7p/Cvt2p: A novel protein-activating enzyme essential for autophagy. Mol. Biol. Cell 1999, 10, 1367–1379. [Google Scholar]
- Kihara, A.; Noda, T.; Ishihara, N.; Ohsumi, Y. Two distinct Vps34 phosphatidylinositol 3-kinase complexes function in autophagy and carboxypeptidase Y sorting in Saccharomyces cerevisiae. J. Cell Biol. 2001, 152, 519–530. [Google Scholar] [CrossRef]
- Shintani, T.; Suzuki, K.; Kamada, Y.; Noda, T.; Ohsumi, Y. Apg2p functions in autophagosome formation on the perivacuolar structure. J. Biol. Chem. 2001, 276, 30452–30460. [Google Scholar]
- Yang, Z.; Huang, J.; Geng, J.; Nair, U.; Klionsky, D.J. Atg22 recycles amino acids to link the degradative and recycling functions of autophagy. Mol. Biol. Cell 2006, 17, 5094–5104. [Google Scholar] [CrossRef]
- Kim, J.; Kamada, Y.; Stromhaug, P.E.; Guan, J.; Hefner-Gravink, A.; Baba, M.; Scott, S.V.; Ohsumi, Y.; Dunn, W.A., Jr.; Klionsky, D.J. Cvt9/Gsa9 functions in sequestering selective cytosolic cargo destined for the vacuole. J. Cell Biol. 2001, 153, 381–396. [Google Scholar] [CrossRef]
- Scott, S.V.; Guan, J.; Hutchins, M.U.; Kim, J.; Klionsky, D.J. Cvt19 is a receptor for the cytoplasm-to-vacuole targeting pathway. Mol. Cell 2001, 7, 1131–1141. [Google Scholar] [CrossRef]
- Nice, D.C.; Sato, T.K.; Stromhaug, P.E.; Emr, S.D.; Klionsky, D.J. Cooperative binding of the cytoplasm to vacuole targeting pathway proteins, Cvt13 and Cvt20, to phosphatidylinositol 3-phosphate at the pre-autophagosomal structure is required for selective autophagy. J. Biol. Chem. 2002, 277, 30198–30207. [Google Scholar]
- Tucker, K.A.; Reggiori, F.; Dunn, W.A., Jr.; Klionsky, D.J. Atg23 is essential for the cytoplasm to vacuole targeting pathway and efficient autophagy but not pexophagy. J. Biol. Chem. 2003, 278, 48445–48452. [Google Scholar]
- Wurmser, A.E.; Emr, S.D. Novel PtdIns(3)P-binding protein Etf1 functions as an effector of the Vps34 PtdIns 3-kinase in autophagy. J. Cell Biol. 2002, 158, 761–772. [Google Scholar] [CrossRef]
- Strømhaug, P.E.; Reggiori, F.; Guan, J.; Wang, C.W.; Klionsky, D.J. Atg21 is a phosphoinositide binding protein required for efficient lipidation and localization of Atg8 during uptake of aminopeptidase I by selective autophagy. Mol. Biol. Cell 2004, 15, 3553–3566. [Google Scholar] [CrossRef]
- Nazarko, T.Y.; Polupanov, A.S.; Manjithaya, R.R.; Subramani, S.; Sibirny, A.A. The requirement of sterol glucoside for pexophagy in yeast is dependent on the species and nature of peroxisome inducers. Mol. Biol. Cell 2007, 18, 106–118. [Google Scholar]
- Monastyrska, I.; Kiel, J.A.; Krikken, A.M.; Komduur, J.A.; Veenhuis, M.; van der Klei, I.J. The Hansenula polymorpha ATG25 gene encodes a novel coiled-coil protein that is required for macropexophagy. Autophagy 2005, 1, 92–100. [Google Scholar]
- Stasyk, O.V.; Stasyk, O.G.; Mathewson, R.D.; Farré, J.C.; Nazarko, V.Y.; Krasovska, O.S.; Subramani, S.; Cregg, J.M.; Sibirny, A.A. Atg28, a novel coiled-coil protein involved in autophagic degradation of peroxisomes in the methylotrophic yeast Pichia pastoris. Autophagy 2006, 2, 30–38. [Google Scholar]
- Farré, J.C.; Manjithaya, R.; Mathewson, R.D.; Subramani, S. PpAtg30 tags peroxisomes for turnover by selective autophagy. Dev. Cell 2008, 14, 365–376. [Google Scholar] [CrossRef]
- Kanki, T.; Wang, K.; Cao, Y.; Baba, M.; Klionsky, D.J. Atg32 is a mitochondrial protein that confers selectivity during mitophagy. Dev. Cell 2009, 17, 98–109. [Google Scholar] [CrossRef]
- Kanki, T.; Wang, K.; Baba, M.; Bartholomew, C.R.; Lynch-Day, M.A.; Du, Z.; Geng, J.; Mao, K.; Yang, Z.; Yen, W.L.; et al. A genomic screen for yeast mutants defective in selective mitochondria autophagy. Mol. Biol. Cell 2009, 20, 4730–4738. [Google Scholar] [CrossRef]
- Liu, Y.; Schiff, M.; Czymmek, K.; Tallóczy, Z.; Levine, B.; Dinesh-Kumar, S.P. Autophagy regulates programmed cell death during the plant innate immune response. Cell 2005, 121, 567–577. [Google Scholar] [CrossRef]
- Liu, Y.; Xiong, Y.; Bassham, D.C. Autophagy is required for tolerance of drought and salt stress in plants. Autophagy 2009, 5, 954–963. [Google Scholar] [CrossRef]
- Ma, J.; Jin, R.; Jia, X.; Dobry, C.J.; Wang, L.; Reggiori, F.; Zhu, J.; Kumar, A. An interrelationship between autophagy and filamentous growth in budding yeast. Genetics 2007, 177, 205–214. [Google Scholar] [CrossRef]
- Mizushima, N. The pleiotropic role of autophagy: from protein metabolism to bactericide. Cell Death Differ. 2005, 12, 1535–1541. [Google Scholar] [CrossRef]
- Shen, W.; Ganetzky, B. Autophagy promotes synapse development in Drosophila. J. Cell Biol. 2009, 187, 71–79. [Google Scholar] [CrossRef]
- Wada, S.; Ishida, H.; Izumi, M.; Yoshimoto, K.; Ohsumi, Y.; Mae, T.; Makino, A. Autophagy plays a role in chloroplast degradation during senescence in individually darkened leaves. Plant Physiol. 2009, 149, 885–893. [Google Scholar]
- Xiong, Y.; Contento, A.L.; Bassham, D.C. Disruption of autophagy results in constitutive oxidative stress in Arabidopsis. Autophagy 2007, 3, 257–258. [Google Scholar]
- Kimmelman, A.C. The dynamic nature of autophagy in cancer. Genes Dev. 2011, 25, 1999–2010. [Google Scholar] [CrossRef]
- Bove, J.; Martinez-Vicente, M.; Vila, M. Fighting neurodegeneration with rapamycin: mechanistic insights. Nat. Rev. Neurosci. 2011, 12, 437–452. [Google Scholar]
- Harris, H.; Rubinsztein, D.C. Control of autophagy as a therapy for neurodegenerative disease. Nat. Rev. Neurol. 2012, 8, 108–117. [Google Scholar]
- Anglade, P.; Vyas, S.; Javoy-Agid, F.; Herrero, M.T.; Michel, P.P.; Marquez, J.; Mouatt-Prigent, A.; Ruberg, M.; Hirsch, E.C.; Agid, Y. Apoptosis and autophagy in nigral neurons of patients with Parkinson's disease. Histol. Histopathol. 1997, 12, 25–31. [Google Scholar]
- Pollack, J.K.; Harris, S.D.; Marten, M.R. Autophagy in filamentous fungi. Fungal. Genet. Biol. 2009, 46, 1–8. [Google Scholar] [CrossRef]
- Bartoszewska, M.; Kiel, J.A. The role of macroautophagy in development of filamentous fungi. Antioxid. Redox Signal. 2011, 14, 2271–2287. [Google Scholar] [CrossRef]
- Deng, Y.Z.; Ramos-Pamplona, M.; Naqvi, N.I. Methods for functional analysis of macroautophagy in filamentous fungi. Methods Enzymol. 2008, 451, 295–310. [Google Scholar]
- Klionsky, D.J.; Abeliovich, H.; Agostinis, P.; Agrawal, D.K.; Aliev, G.; Askew, D.S.; Baba, M.; Baehrecke, E.H.; Bahr, B.A.; Ballabio, A.; et al. Guidelines for the use and interpretation of assays for monitoring autophagy in higher eukaryotes. Autophagy 2008, 4, 151–175. [Google Scholar]
- Mizushima, N. Methods for monitoring macroautophagy. Tanpakushitsu Kakusan Koso. 2006, 51, 1542–1548. [Google Scholar]
- Deng, Y.Z.; Ramos-Pamplona, M.; Naqvi, N.I. Autophagy-assisted glycogen catabolism regulates asexual differentiation in Magnaporthe oryzae. Autophagy 2009, 5, 33–43. [Google Scholar] [CrossRef]
- Kikuma, T.; Ohneda, M.; Arioka, M.; Kitamoto, K. Functional analysis of the ATG8 homologue Aoatg8 and role of autophagy in differentiation and germination in Aspergillus oryzae. Eukaryot. Cell 2006, 5, 1328–1336. [Google Scholar] [CrossRef]
- Hu, G.; Hacham, M.; Waterman, S.R.; Panepinto, J.; Shin, S.; Liu, X.; Gibbons, J.; Valyi-Nagy, T.; Obara, K.; Jaffe, H.A.; et al. PI3K signaling of autophagy is required for starvation tolerance and virulenceof Cryptococcus neoformans. J. Clin. Invest. 2008, 118, 1186–1197. [Google Scholar] [CrossRef]
- Fan, W.; Kraus, P.R.; Boily, M.J.; Heitman, J. Cryptococcus neoformans gene expression during murine macrophage infection. Eukaryot. Cell 2005, 4, 1420–1433. [Google Scholar] [CrossRef]
- Nolting, N.; Bernhards, Y.; Poggeler, S. SmATG7 is required for viability in the homothallic ascomycete Sordaria macrospora. Fungal. Genet. Biol. 2009, 46, 531–542. [Google Scholar] [CrossRef]
- Nguyen, L.N.; Bormann, J.; Le, G.T.; Stärkel, C.; Olsson, S.; Nosanchuk, J.D.; Giese, H.; Schäfer, W. Autophagy-related lipase FgATG15 of Fusarium graminearum is important for lipid turnover and plant infection. Fungal. Genet. Biol. 2011, 48, 217–224. [Google Scholar] [CrossRef]
- Josefsen, L.; Droce, A.; Sondergaard, T.E.; Sørensen, J.L.; Bormann, J.; Schäfer, W.; Giese, H.; Olsson, S. Autophagy provides nutrients for nonassimilating fungal structures and is necessary for plant colonization but not for infection in the necrotrophic plant pathogen Fusarium graminearum. Autophagy 2012, 8, 326–337. [Google Scholar] [CrossRef]
- Liu, X.H.; Yang, J.; He, R.L.; Lu, J.P.; Zhang, C.L.; Lu, S.L.; Lin, F.C. An autophagy gene, TrATG5, affects conidiospore differentiation in Trichoderma reesei. Res. Microbiol. 2011, 162, 756–763. [Google Scholar] [CrossRef]
- Richie, D.L.; Askew, D.S. Autophagy in the filamentous fungus Aspergillus fumigatus. Methods Enzymol. 2008, 451, 241–250. [Google Scholar]
- Richie, D.L.; Fuller, K.K.; Fortwendel, J.; Miley, M.D.; McCarthy, J.W.; Feldmesser, M.; Rhodes, J.C.; Askew, D.S. Unexpected link between metal ion deficiency and autophagy in Aspergillus fumigatus. Eukaryot. Cell 2007, 6, 2437–2447. [Google Scholar] [CrossRef]
- Dufresne, M.; Bailey, J.A.; Dron, M.; Langin, T. clk1, a serine/threonine protein kinase-encoding gene, is involved in pathogenicity of Colletotrichum lindemuthianum on common bean. Mol. Plant Microbe Interact. 1998, 11, 99–108. [Google Scholar] [CrossRef]
- Liu, X.H.; Lu, J.P.; Zhang, L.; Dong, B.; Min, H.; Lin, F.C. Involvement of a Magnaporthe grisea serine/threonine kinase gene, MgATG1, in appressorium turgor and pathogenesis. Eukaryot. Cell 2007, 6, 997–1005. [Google Scholar] [CrossRef]
- Liu, X.H.; Liu, T.B.; Lin, F.C. Monitoring autophagy in Magnaporthe oryzae. Methods Enzymol. 2008, 451, 271–294. [Google Scholar]
- Veneault-Fourrey, C.; Barooah, M.; Egan, M.; Wakley, G.; Talbot, N.J. Autophagic fungal cell death is necessary for infection by the rice blast fungus. Science 2006, 312, 580–583. [Google Scholar]
- Nadal, M.; Gold, S.E. The autophagy genes ATG8 and ATG1 affect morphogenesis and pathogenicity in Ustilago maydis. Mol. Plant Pathol. 2010, 11, 463–478. [Google Scholar] [CrossRef]
- Takano, Y.; Asakura, M.; Sakai, Y. Atg26-mediated pexophagy and fungal phytopathogenicity. Autophagy 2009, 5, 1041–1042. [Google Scholar] [CrossRef]
- Asakura, M.; Ninomiya, S.; Sugimoto, M.; Oku, M.; Yamashita, S.; Okuno, T.; Sakai, Y.; Takano, Y. Atg26-mediated pexophagy is required for host invasion by the plant pathogenic fungus Colletotrichum orbiculare. Plant Cell 2009, 21, 1291–1304. [Google Scholar] [CrossRef]
- Roetzer, A.; Gratz, N.; Kovarik, P.; Schüller, C. Autophagy supports Candida glabrata survival during phagocytosis. Cell Microbiol. 2010, 12, 199–216. [Google Scholar] [CrossRef]
- Kershaw, M.J.; Talbot, N.J. Genome-wide functional analysis reveals that infection-associated fungal autophagy is necessary for rice blast disease. Proc. Natl. Acad. Sci. USA 2009, 106, 15967–15972. [Google Scholar] [CrossRef]
- Palmer, G.E. Autophagy in the invading pathogen. Autophagy 2007, 3, 251–253. [Google Scholar]
- Palmer, G.E. Autophagy in Candida albicans. Methods Enzymol. 2008, 451, 311–322. [Google Scholar]
- Dice, J.F.; Walker, C.D.; Byrne, B.; Cardiel, A. General characteristics of protein degradation in diabetes and starvation. Proc. Natl. Acad. Sci. USA 1978, 75, 2093–2097. [Google Scholar] [CrossRef]
- Adams, T.H.; Timberlake, W.E. Developmental repression of growth and gene expression in Aspergillus. Proc. Natl. Acad. Sci. USA 1990, 87, 5405–5409. [Google Scholar]
- Ou, S.H. Rice Diseases; The Commonwealth Mycological Institute: Surrey, UK, 1985. [Google Scholar]
- Cole, G.T. Models of cell differentiation in conidial fungi. Microbiol. Rev. 1986, 50, 95–132. [Google Scholar]
- Teng, P.S.; Klein-Gebbinck, H.W.; Pinnschmidt, H. An analysis of the blast pathosystem to guide modeling and forecasting. In Rice Blast Modeling and Forecasting; International Rice Research Institute: Manila, Philippines, 1991; pp. 1–30. [Google Scholar]
- Lee, Y.H.; Dean, R.A. cAMP Regulates Infection Structure Formation in the Plant Pathogenic Fungus Magnaporthe grisea. Plant Cell 1993, 5, 693–700. [Google Scholar]
- Gilbert, R.D.; Johnson, A.M.; Dean, R.A. Chemical signals responsible for appressorium formation in the rice blast fungus Magnaporthe grisea. Physiol. Mol. Plant Pathol. 1996, 48, 335–346. [Google Scholar] [CrossRef]
- Wang, Z.Y.; Jenkinson, J.M.; Holcombe, L.J.; Soanes, D.M.; Veneault-Fourrey, C.; Bhambra, G.K.; Talbot, N.J. The molecular biology of appressorium turgor generation by the rice blast fungus Magnaporthe grisea. Biochem. Soc. Trans. 2005, 33, 384–388. [Google Scholar]
- Talbot, N.J. Having a blast: Exploring the pathogenicity of Magnaporthe grisea. Trends Microbiol. 1995, 3, 9–16. [Google Scholar] [CrossRef]
- Deng, Y.; Naqvi, N.I. A vacuolar glucoamylase, Sga1, participates in glycogen autophagy for proper asexual differentiation in Magnaporthe oryzae. Autophagy 2010, 6, 455–461. [Google Scholar] [CrossRef]
- Devos, P.; Baudhuin, P.; Van Hoof, F.; Hers, H.G. The alpha particulate liver glycogen. A morphometric approach to the kinetics of its synthesis and degradation. Biochem. J. 1983, 209, 159–165. [Google Scholar]
- Willms, K.; Robert, L.; Caro, J.A. Ultrastructure of smooth muscle, gap junctions and glycogen distribution in Taenia solium tapeworms from experimentally infected hamsters. Parasitol. Res. 2003, 89, 308–316. [Google Scholar]
- Marchand, I.; Chorneyko, K.; Tarnopolsky, M.; Hamilton, S.; Shearer, J.; Potvin, J.; Graham, T.E. Quantification of subcellular glycogen in resting human muscle: granule size, number, and location. J. Appl. Physiol. 2002, 93, 1598–1607. [Google Scholar]
- He, M.; Kershaw, M.J.; Soanes, D.M.; Xia, Y.; Talbot, N.J. Infection-associated nuclear degeneration in the rice blast fungus Magnaporthe oryzae requires non-selective macro-autophagy. PLoS One 2012, 7, e33270. [Google Scholar]
- Yi, M.; Park, J.H.; Ahn, J.H.; Lee, Y.H. MoSNF1 regulates sporulation and pathogenicity in the rice blast fungus Magnaporthe oryzae. Fungal. Genet. Biol. 2008, 45, 1172–1181. [Google Scholar] [CrossRef]
- Wilson, R.A.; Jenkinson, J.M.; Gibson, R.P.; Littlechild, J.A.; Wang, Z.Y.; Talbot, N.J. Tps1 regulates the pentose phosphate pathway, nitrogen metabolism and fungal virulence. EMBO J. 2007, 26, 3673–3685. [Google Scholar]
- Gancedo, J.M. Yeast carbon catabolite repression. Microbiol. Mol. Biol. Rev. 1998, 62, 334–361. [Google Scholar]
- Wang, Z.; Wilson, W.A.; Fujino, M.A.; Roach, P.J. Antagonistic controls of autophagy and glycogen accumulation by Snf1p, the yeast homolog of AMP-activated protein kinase, and the cyclin-dependent kinase Pho85p. Mol. Cell Biol. 2001, 21, 5742–5752. [Google Scholar] [CrossRef]
- Shoji, J.Y.; Kikuma, T.; Arioka, M.; Kitamoto, K. Macroautophagy-mediated degradation of whole nuclei in the filamentous fungus Aspergillus oryzae. PLoS One 2010, 5, e15650. [Google Scholar]
- Denisov, Y.; Freeman, S.; Yarden, O. Inactivation of Snt2, a BAH/PHD-containing transcription factor, impairs pathogenicity and increases autophagosome abundance in Fusarium oxysporum. Mol. Plant Pathol. 2011, 12, 449–461. [Google Scholar] [CrossRef]
- Pungartnik, C.; Melo, S.C.; Basso, T.S.; Macena, W.G.; Cascardo, J.C.; Brendel, M. Reactive oxygen species and autophagy play a role in survival and differentiation of the phytopathogen Moniliophthora perniciosa. Fungal. Genet. Biol. 2009, 46, 461–472. [Google Scholar] [CrossRef]
- Emri, T.; Molnár, Z.; Szilágyi, M.; Pócsi, I. Regulation of autolysis in Aspergillus nidulans. Appl. Biochem. Biotechnol. 2008, 151, 211–220. [Google Scholar] [CrossRef]
- Cebollero, E.; Gonzalez, R. Induction of autophagy by second-fermentation yeasts during elaboration of sparkling wines. Appl. Environ. Microbiol. 2006, 72, 4121–4127. [Google Scholar] [CrossRef]
- Lorenz, M.C.; Bender, J.A.; Fink, G.R. Transcriptional response of Candida albicans upon internalization by macrophages. Eukaryot. Cell 2004, 3, 1076–1087. [Google Scholar] [CrossRef]
- Nakagawa, Y.; Kanbe, T.; Mizuguchi, I. Disruption of the human pathogenic yeast Candida albicans catalase gene decreases survival in mouse-model infection and elevates susceptibility to higher temperature and to detergents. Microbiol. Immunol. 2003, 47, 395–403. [Google Scholar]
- Palmer, G.E.; Askew, D.S.; Williamson, P.R. The diverse roles of autophagy in medically important fungi. Autophagy 2008, 4, 982–988. [Google Scholar]
- Karkowska-Kuleta, J.; Rapala-Kozik, M.; Kozik, A. Fungi pathogenic to humans: molecular bases of virulence of Candida albicans, Cryptococcus neoformans and Aspergillus fumigatus. Acta Biochim. Pol. 2009, 56, 211–224. [Google Scholar]
- Yano, T.; Kurata, S. Intracellular recognition of pathogens and autophagy as an innate immune host defence. J. Biochem. 2011, 150, 143–149. [Google Scholar] [CrossRef]
- Yoshimoto, K.; Takano, Y.; Sakai, Y. Autophagy in plants and phytopathogens. FEBS Lett. 2010, 584, 1350–1358. [Google Scholar] [CrossRef]
- Yang, C.S.; Lee, J.S.; Rodgers, M.; Min, C.K.; Lee, J.Y.; Kim, H.J.; Lee, K.H.; Kim, C.J.; Oh, B.; Zandi, E.; et al. Autophagy protein Rubicon mediates phagocytic NADPH oxidase activation in response to microbial infection or TLR stimulation. Cell Host Microbe 2012, 11, 264–276. [Google Scholar] [CrossRef]
- Talbot, N.J.; Kershaw, M.J. The emerging role of autophagy in plant pathogen attack and host defence. Curr. Opin. Plant Biol. 2009, 12, 444–450. [Google Scholar] [CrossRef]
- Lenz, H.D.; Vierstra, R.D.; Nürnberger, T.; Gust, A.A. ATG7 contributes to plant basal immunity towards fungal infection. Plant Signal. Behav. 2011, 6, 1040–1042. [Google Scholar] [CrossRef]
- Lenz, H.D.; Haller, E.; Melzer, E.; Kober, K.; Wurster, K.; Stahl, M.; Bassham, D.C.; Vierstra, R.D.; Parker, J.E.; Bautor, J.; et al. Autophagy differentially controls plant basal immunity to biotrophic and necrotrophic pathogens. Plant J. 2011, 66, 818–830. [Google Scholar]
- Lai, Z.; Wang, F.; Zheng, Z.; Fan, B.; Chen, Z. A critical role of autophagy in plant resistance to necrotrophic fungal pathogens. Plant J. 2011, 66, 953–968. [Google Scholar] [CrossRef]
- Deng, Y.; Qu, Z.; He, Y.; Naqvi, N.I. Sorting nexin Snx41 is essential for conidiation and mediates glutathione-based antioxidant defense during invasive growth in Magnaporthe oryzae. Autophagy 2012, in press. [Google Scholar]
- Qin, Q.M.; Luo, J.; Lin, X.; Pei, J.; Li, L.; Ficht, T.A.; de Figueiredo, P. Functional analysis of host factors that mediate the intracellular lifestyle of Cryptococcus neoformans. PLoS Pathog. 2011, 7, e1002078. [Google Scholar] [CrossRef]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Deng, Y.; Qu, Z.; Naqvi, N.I. Role of Macroautophagy in Nutrient Homeostasis During Fungal Development and Pathogenesis. Cells 2012, 1, 449-463. https://doi.org/10.3390/cells1030449
Deng Y, Qu Z, Naqvi NI. Role of Macroautophagy in Nutrient Homeostasis During Fungal Development and Pathogenesis. Cells. 2012; 1(3):449-463. https://doi.org/10.3390/cells1030449
Chicago/Turabian StyleDeng, Yizhen, Ziwei Qu, and Naweed I. Naqvi. 2012. "Role of Macroautophagy in Nutrient Homeostasis During Fungal Development and Pathogenesis" Cells 1, no. 3: 449-463. https://doi.org/10.3390/cells1030449
APA StyleDeng, Y., Qu, Z., & Naqvi, N. I. (2012). Role of Macroautophagy in Nutrient Homeostasis During Fungal Development and Pathogenesis. Cells, 1(3), 449-463. https://doi.org/10.3390/cells1030449