Abstract
Dairy and livestock industry drives the economy and food security through sustainable supply of dairy products and meat across the globe. Dairy farm operations produce a large quantity of manure, which is a cheap and abundant plant nutrient source. However, insufficient forage production with low quality matrix are the current challenges of dairy industry in boreal climate due to extreme weather conditions. To address these challenges, a field experiment was conducted for three years to determine the effects of organic (dairy manure-based phosphorus (DMP)) and inorganic phosphorus (P) amendments on forage yield and quality indices of silage-corn cultivated in boreal climate. Experimental treatments were: (i) DMP with high P concentration (DMP1); (ii) DMP with low P concentration (DMP2) and (iii) inorganic P, also designated as control; and five silage-corn genotypes (Fusion-RR, Yukon-R, A4177G3-RIB, DKC23-17RIB, DKC26-28RIB). Results revealed that DMP1 amendment produced significantly higher forage yield compared to inorganic P, whereas non-significant effects were shown on quality indices except P mineral, available and crude protein. Yukon-R and DKC26-28RIB showed superior agronomic performance and produced significantly higher forage yield, whereas A4177G3-RIB produced lowest forage yield but exhibited superior nutritional quality; higher minerals, protein, total digestible nutrients, net energy for gain, net energy for maintenance and calculated milk production compared to other genotypes. Yukon-R not only produced higher forage, but also displayed good forage quality indices which were very close to A4177G3-RIB genotype. Therefore, we conclude that Yukon-R cultivation following DMP as organic amendment could be a sustainable production practice to attain high forage yield with optimum nutritional quality to meet the forage needs of growing dairy industry in boreal climate.
1. Introduction
Sustainable food production and food security is the prime objective of agricultural production systems [1]. The global population is predicted to grow to 9.2 billion through 2050, necessitating enormous increases in food production, as well as reductions in waste [2]. Annual cereal and meat production will need to rise to about 900 and 200 million tons respectively, to feed the burgeoning world population [3]. To achieve this, animal forage with improved quality indices will be required to increase milk and meat production [4].
Corn (Zea mays L.) is one of the most important food and forage crops contributing to the global food security for human and animals [5]. Silage-corn is a high-energy crop with multifaceted benefits compared to other forage species [6]. For example, higher biomass production potential, high energy content, more uniform quality with one cut forage harvest [7,8]. Additionally, silage-corn is highly palatable, digestible and easy to ensile due to high soluble sugar contents [9]. Owing to high energy contents, milk production increased significantly with increased silage-corn acreage compared to other grasses as a sole source of energy [10]. Consequently, dairy farmers need a sustainable supply of high-quality forage which can only be achieved through adaptations of sustainable agricultural practices that make use of local resources to enhance forage production and quality and diversify the forage production systems. Enhancing milk production and quality in response to ever-increasing demands for dairy products requires more sustainable, economically viable, and environmentally beneficial forage cropping systems [11].
Dairy manure (DM) is known as a good soil amendment and a rich source of plant nutrients [12,13]. Therefore, dairy manure-based phosphorus (P) and other nutrient amendments could be a promising strategy to enhance on-farm P and other nutrients recycling, crop growth and forage production [14,15,16]. Phosphorus is one of the essential mineral nutrients required for plant growth and development [17]; however, at the same time, it is immobile in soils. Therefore, P deficiency in crop plants may lead to stunted growth and reduced crop yields [18,19].
The root morphology and architecture plays an important role in exploring the rhizosphere area, which is associated with soil rhizosphere microorganisms (actinomycetes, protozoans, bacteria, and fungi), release organic compounds (phosphatases and organic acids) and inorganic P in the rhizosphere [20,21]. Generally, inorganic fertilization cause P fixation in soils with organic matter or cations making P unavailable for plant uptake [22,23]. Due to such higher reactivity, approximately 70 percent of global croplands have become P deficient making inorganic P fertilization an inevitable input in intensive agricultural production systems. Furthermore, depleting rock phosphate resources is also a growing concern for the agriculture industry in next 50–100 years [24]. Hence, using DMP as a sustainable plant nutrient source could be a promising strategy to attain sustainable supply of forage production, particularly in territories of expanding dairy industry [25]. This can further enhance the efficient utilization of abundantly available DMP as a mineral and carbon source, while simultaneously reducing the dependency on synthetic fertilizers, particularly non-renewable inorganic P [23,24]. Moreover, DMP application may enhance mineral uptake and forage yield [26], acid detergent fiber (ADF), neutral detergent fiber (NDF), and total digestible nutrients (TDN) compared to inorganic nutrient sources [27].
Earlier researchers reported variations in P use efficiency in different crop species and genotypes, most probably due to their genetic makeup [28,29]. Similarly, different silage-corn hybrids expressed significant variations in forage proteins, minerals, ADF, NDF, TDN, forage energies and milk production [30,31,32]. P amendments with different application rates exhibited significant production potential and nutritional quality of forage species [33,34,35]. However, studies reporting the effects of organic and inorganic P amendments on forage production potential and forage qualitative traits of silage-corn genotypes are very limited and inconclusive, particularly relating to podzolic soils common in boreal climate.
Podzolic soils are generally formed in coarse to medium-textured, acidic parent materials, less fertile under forest or heath vegetation in cool climates. These soils may occur in wet sandy sites in sub-humid climates and can also be formed from calcareous parent materials. Most podzolic soils characterized as low fertility, acidic, accumulation of organic matter, Fe, and Al in the illuvial B horizon and low water holding capacity [36]. Therefore, we hypothesized that (i) organic and inorganic P amendments will enhance forage yield potential and forage quality of silage-corn in podzolic soils under boreal climate, and (ii) silage-corn genotypes will exhibit significant variation in forage production potential and quality indices in podzolic soils under boreal climate. Hence, a field research trial was conducted for three years to investigate the effects of organic and inorganic P amendments on forage yield potential and forage quality of silage-corn genotypes in podzol under boreal climate.
2. Materials and Methods
2.1. Experimental Location and Treatments
A field research trial was carried out at Pynn’s Brook Research Station, Pasadena (49° 04ʹ 20” N, 57° 33ʹ 35” W), Canada for three years (2015–2017). The soil, reddish brown to brown, has developed on gravelly sandy fluvial deposit of mixed lithology, with >100 cm depth to bedrock, and soil samples for the study site revealed a gravelly loamy sand soil (sand = 82.0% (± 3.4); silt = 11.6% (± 2.4); clay = 6.4% (± 1.2)), which is classified as orthic humo-ferric podzol [37]. Before start of experiment each year, soil samples were analyzed and detailed soil analyses averaged over three growing seasons are presented in Table 1. The experimental field was rainfed and average rainfall received during the three crop growing seasons was 487.3 ± 103.6 mm, which was 6% higher compared to last thirty years (Table 2). The experimental treatments were comprised of five silage corn genotypes and three P amendments and laid out in randomized complete block design with factorial arrangement with three replications. The five Roundup Ready® silage-corn genotypes were selected based on the required crop heating units (CHU) to assess their adaptation and resilience in cool climate production systems (Table 3). The three P amendments were (i) DMP with high P concentration (DMP1); (ii) DMP with low P concentration (DMP2) and (iii) inorganic P (110 Kg ha−1. We used inorganic P treatment as control to compare the effects of organic P sources (DMP1, DMP2) on forage yield and quality indices in silage-corn genotypes. DMP application is seen as a common practice in dairy and livestock farming sector for cultivating forage crops. Therefore, we aimed to investigate the response of DMP application with two different mineral composition and inorganic P amendment on forage production and forage quality indices in podzolic soils under boreal climates. Well-agitated DMP samples were collected from ten dairy farms of western NL and were sent to the Soil, Plant and Feed Laboratory, Department of Fisheries and Land Resources, St. John’s, NL. DM samples with high and low P concentration were chosen (dairy farms) and designated as DMP1 and DMP2, respectively. Detailed manure analyses reports are given in Table 4. It is pertinent to mention that DMP with high P concentration also contained high N and K as well (Table 4). During the start of each growing season, well-agitated DMP samples were collected from the same two dairy farms and were sent to the laboratory for complete nutrient analyses prior to field application. DMP was applied to the respective treatment plots @ 30,000 L ha−1 according to the local dairy farmers practice and was thoroughly mixed in the top 15–20 cm soil layer before crop seeding. The remaining required nutrients in DMP1 and DMP2 treatments were supplemented through inorganic fertilizers following regional recommendations of silage-corn. Experimental plots were georeferenced after harvest to keep the treatments at the same location over three years’ field trial.

Table 1.
Basic soil properties of experimental site during three growing seasons. Values are means ± standard error for three years.
Table 2.
Mean average air temperature, mean maximum and minimum temperature and rainfall at experimental site during three growing seasons. Crop growing season average represents means of three growing seasons ± standard error (n = 3), whereas the last thirty years data represent means from 1985–2014 for crop growth period.
Table 3.
Genotypic traits of the five silage-corn varieties cultivated under field conditions in a cool climate production system.
Table 4.
Dairy manure analyses report of both sources used in this experiment. Values are means ± standard error for three years (2015–2017).
2.2. Crop Management and Forage Sampling
Silage-corn was seeded with SAMCO system (SAMCO 2200 Agricultural Manufacturing, Limerick, Ireland). The partially perforated plastic sheets cover one-meter-wide corn rows to supplement additional extra heating units to enhance germination and seedling establishment during early crop growth stage. Seeding was done on June 4, May 25 and May 23 and harvested on October 13, 18, and 13 during 2015, 2016 and 2017, respectively. Plant population was maintained by seeding 90,900 plants per hectare. Glyphosate (WeatherMax) herbicide was applied to control weeds according to the instructions on the label (Monsanto Canada Inc.). One square meter area from the centre of each experimental plot was hand-cut at ground level at the black layer stage (R6) which is considered optimum to attain high forage nutritional quality [38]. The fresh forage biomass obtained from one square meter was weighed and recorded; thereafter a subsample was taken, chopped with knife into small pieces and dried in a forced air oven (Shell Labs, Cornelius, OR, USA) at 65 ○C until constant dry weight. The dry matter content (g kg−1) was then calculated based on the total fresh and dry weights of the sample as described by Kwabiah [39]. Total forage biomass yield was converted from one square meter to one hectare on a dry matter basis (equations below) and was reported as Mg ha−1.
2.3. Forage Quality Analysis
Three plants were randomly selected from each experimental plot to carry out forage quality analyses. Whole plants were chopped, pooled and dried at 65 °C in a forced air oven (Shell Labs, Cornelius, OR, USA) until constant weight. The plant samples were ground with a grinder mill (Wily Mill Standard Model-3, Arthur H. Thomas Co. Philadelphia, PA, USA), and sieved using 500 µM screen. Thereafter, sieved samples (50 g) obtained from each experimental plot were packed in small paper bags and sent to Actlabs or Activation Laboratories Ltd. (Ancaster ON, Canada), a member laboratory of Dairy One Feed and Forage Analyses Laboratory (Ithaca, New York, USA) for quality analyses. Forage quality parameters used in this study were divided into measured (minerals, proteins, fibers), and calculated values (non-fibrous carbohydrates, TDN, NFC, NEL, NEM, NEG, calculated milk production) following analysis by near-infrared reflectance spectroscopy (NIRS, FOSS XDS System, Model 6500 Win ISI II v1.5) [27,40,41,42]. For measured parameters, briefly minerals in the forage samples (500 mg) were pre-digested for 10 min with 8 mL HNO3 and 2 mL HCL followed by an additional 10 min with 1 mL (30%) H2O2 using a microwave digestor (CEM System, MARS-6, Matthews, NC, USA). After pre-digestion, samples were digested in two stages: 10 min ramp to 135 °C and held for 3 min at 1500 W (stage one); and for 12 min at 200 °C and held for 15 min at 1600 W. The digested samples were filtered and then analyzed using inductivity coupled plasma spectrometer (iCAP-6300 Series, Thermo Scientific, Cambridge, UK). Acid detergent fiber (ADF) and neutral detergent fiber (NDF) were assessed by weighing 500 mg forage sample into filter bags and digested for 75 min with H2SO4 in a ANKOM A2000 digestion system by adopting ANKOM technology method-12 and method-13, respectively (Ankom Technology Crop, Macedon, NY, USA). Samples were then rinsed three times with boiling water for 5 min in filter bags followed by a 3 min acetone soak and dried at 105 °C for 2 h [43]. Total nitrogen in the forage samples was measured by combusting 1 mm sieved, ground and dried forage sample in a CN analyzer (CN-628, Leco, St. Joseph, MI, USA) and N contents were multiplied by 6.25 to get the crude protein in silage-corn forage samples [15,44]. Estimated parameters such as non-fibrous carbohydrates (NFC) were quantified using summative equation by [43,45]. The total digestible nutrients (TDN) were assessed using a summative equation based on forage quality components [46]. Net energy for lactation (NEL) was predicted by applying discount approach as suggested by Van Soest [45]. Net energy for maintenance (NEM) and net energy for gain (NEG) was predicted using National Research Council standard equations [47] and variable discount approach [45]. Calculated milk production was done according to the standard method [48].
2.4. Statistical Analyses
Data were pooled for three years and analyses were carried out to evaluate observed variables using ANOVA technique by employing the Statistix-10 software package (Analytical Software, Tallahassee, FL, USA). Where treatment effects were significant, the means were compared with Fisher’s least significant difference (LSD) test at p ≤ 0.05. Principal component analysis (PCA) was then performed by (XLStat Premium, Perpetual version 2018.1.1, NY, USA) to determine the similarities or groupings of the genotypes, and P amendments based on forage yield and nutritional quality parameters. Figures were prepared using XLStat (XLStat Premium, Perpetual version 2018.1.1, NY, USA) and Sigma Plot 13.0 (Systat Software Inc., San Jose, CA, USA) software packages.
3. Results
3.1. Forage Yield
Inorganic and organic P amendments had significant effects on forage production and quality indices of silage-corn genotypes (Table 5). DMP1 produced significantly higher forage yield compared to inorganic P which produced lower forage yield; however, it was statistically on par with DMP2 treatment (Figure 1a). Among silage-corn genotypes, Yukon-R and DKC26-28RIB showed superior agronomic performance and produced 29% and 21% higher forage yield compared to A4177G3-RIB which produced lowest forage yield (Figure 1b).
Table 5.
Analyses of variance (ANOVA) regarding forage yield and quality indices of silage-corn genotypes under organic and inorganic amendments.
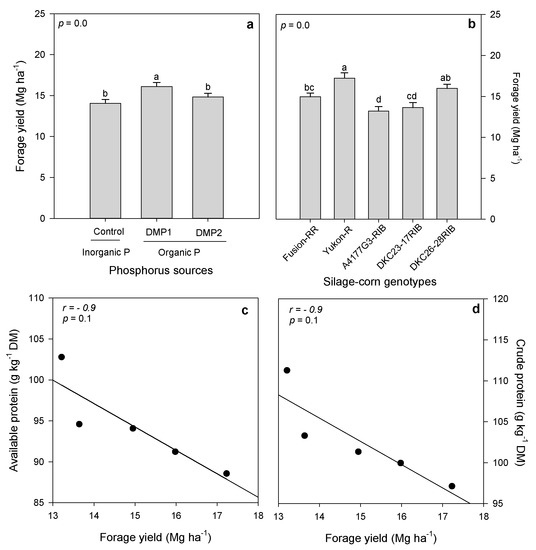
Figure 1.
Effects of inorganic and organic phosphorus amendments on forage yield (a). Variation in forage production among silage-corn genotypes (b). Pearson’s correlation showing the association between forage yield and available protein (c) and crude protein (d) among five silage corn genotypes. Each bar represents mean of three replications ± SE.
3.2. Forage Quality
3.2.1. Mineral Composition
Organic and inorganic P amendments expressed non-significant effects on forage minerals (K, Ca and Mg) (Figure 2b–d), and significant effects on P mineral (Table 5). Inorganic P amendment exhibited higher forage P mineral compared to DMP1, which showed lower P, although DMP2 and inorganic P amendments were statistically non-significant (Figure 2a). Silage-corn genotypes expressed significant variations in forage P, K and Mg, but non-significant variation in Ca (Figure 3a–d). Significantly, higher P and K minerals were observed in A4177G3-RIB and DKC26-28RIB, compared to lowest were noted in Fusion-RR (Figure 3a,b). A4177G3-RIB also produced higher Mg contents compared to Fusion-RR that showed lower Mg (Figure 3d). However, silage-corn genotypes had non-significant effects on Ca (Figure 3c).
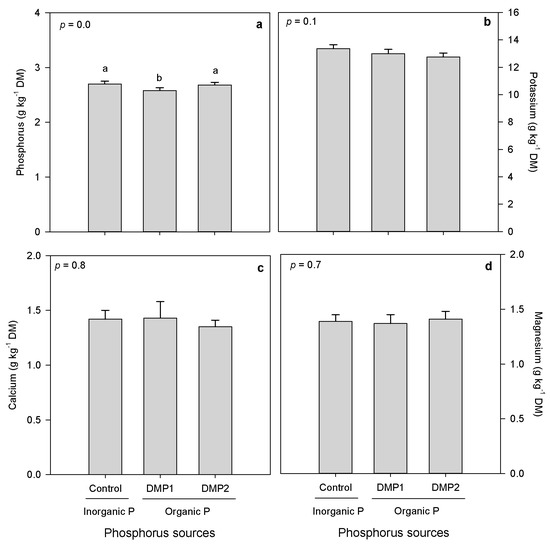
Figure 2.
Effects of inorganic and organic phosphorus amendments on forage minerals of silage-corn (a) phosphorus, (b) potassium, (c) calcium and (d) magnesium grown in boreal climate. Each bar represents mean of three replications ± SE.
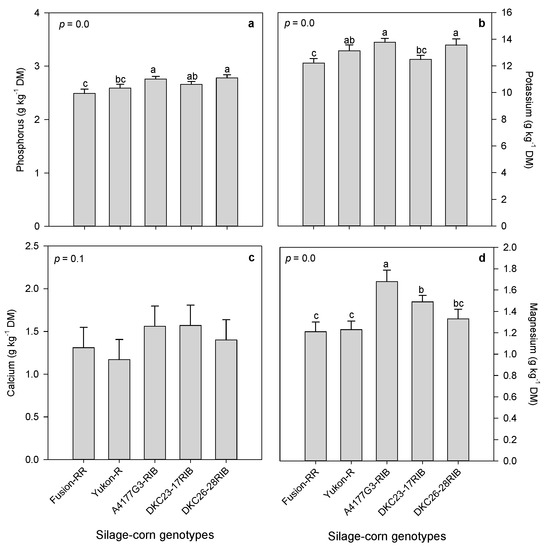
Figure 3.
Variations in forage minerals among silage-corn genotypes: 3(a) phosphorus, 3(b) potassium, 3(c) calcium, and 3(d) magnesium. Each bar represents mean of three replications ± SE.
3.2.2. Forage Proteins
Organic and inorganic P amendments had significant effects on available and crude forage proteins when grown under field conditions in boreal climate (Table 5). Inorganic P amendment exhibited higher available and crude protein, whereas organic P amendments produced lower available and crude protein. However, DMP1 and DMP2 were statistically on par and produced similar available and crude protein (Figure 4a,b). Likewise, silage-corn genotypes expressed significant variation in available and crude protein (Table 5). Higher available and crude protein were observed in A4177G3-RIB followed by DKC23-17RIB and Fusion-RR, whereas minimum was recorded in Yukon-R (Figure 4c,d).
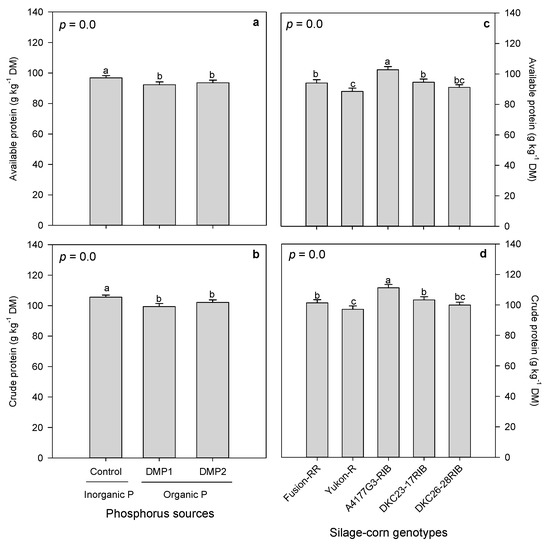
Figure 4.
Effects of inorganic and organic phosphorus amendments on forage proteins: (a) available protein and (b) crude protein. Variation in available protein (c) and crude protein (d) among silage-corn genotypes. Each bar represents mean of three replications ± SE.
3.2.3. Fiber Contents
Phosphorus amendments expressed non-significant effects on ADF, NDF and NFC (Table 5). However, ADF and NDF contents are slightly higher in inorganic P compared to organic P amendment, whereas organic amendment exhibited slightly higher NFC compared to inorganic P amendment (Figure 5a–c). ADF, NDF and NFC varied significantly among five silage-corn genotypes, where Fusion-RR produced higher ADF and NDF contents compared to lowest exhibited by DKC-2628 RIB (Figure 5d,e). However, the trend was quite the opposite for NFC where DKC-2628 RIB produced significantly higher NFC and lowest were recorded in Fusion-RR (Figure 5f).
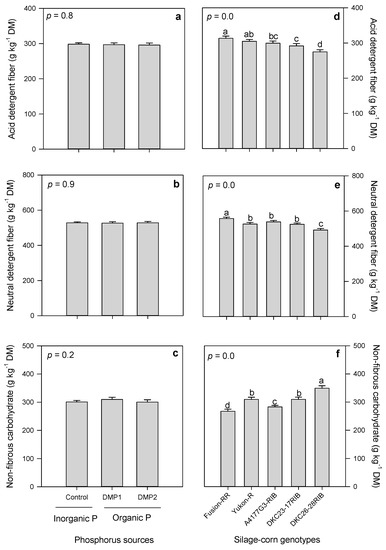
Figure 5.
Effects of inorganic and organic phosphorus amendments on forage fibers: (a) acid detergent fiber (ADF), (b) neutral detergent fiber (NDF), and (c) non-fibrous carbohydrates (NFC). Variation in ADF (d), NDF (e), and NFC (f) among silage-corn genotypes. Each bar represents mean of three replications ± SE.
3.2.4. Forage Energies
Phosphorus amendments expressed non-significant effects on NEM, NEG and NEL. However, DMP1 (organic P amendment) exhibited slightly higher NEM, NEG and NEL compared to inorganic P (Figure 6a–c).
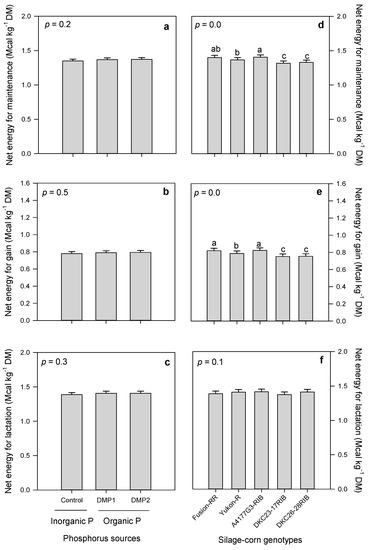
Figure 6.
Effects of inorganic and organic phosphorus amendments on forage energies: NEM (a), NEG (b), and NEL (c). Variations in NEM (d), NEG (e), NEL (f) among silage-corn genotypes grown in boreal climate. Each bar represents mean of three replications ± SE.
Contrarily, silage-corn genotypes displayed significant variations in NEM, NEG and non-significant effects on NEL. Significantly higher NEM and NEG were recorded in A4177G3 RIB compared to lowest was observed in DKC26-28RIB (Figure 6d,e). Likewise, A4177G3-RIB produced slightly higher NEL, although non-significant with other genotypes (Figure 6f).
3.2.5. Total Digestible Nutrients and Calculated Milk Production
Organic and inorganic P amendments expressed non-significant effects on TDN and calculated milk production as depicted in Figure 7a,b. However, silage-corn genotypes had significantly influenced TDN and calculated milk production (Figure 7c,d). A4177G3-RIB showed superior performance in producing higher TDN and calculated milk production, while DKC23-17RIB produced lowest TDN and milk production (Figure 7c,d).
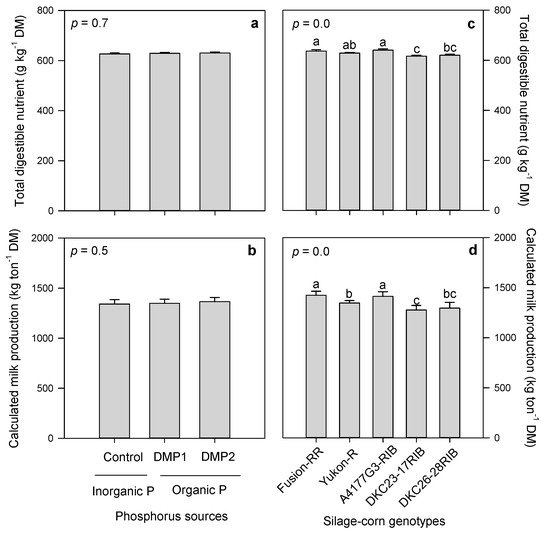
Figure 7.
Effects of inorganic and organic phosphorus amendments on forage total digestible nutrients (a) and calculated milk production (b). Variations in total digestible nutrients (c) and calculated milk production (d) among silage-corn genotypes grown in boreal climate. Each bar represents mean of three replications ± SE.
4. Discussion
4.1. Forage Yield
The pooled weather data over the three growing seasons showed slightly higher (6%) rainfall compared to the last thirty years depicting the changing climatic pattern (Table 2). Increased rainfall during crop growth season in the rainfed areas is predicted to enhance the agricultural production [49]. Results of the present study demonstrated higher forage yield in DMP1 (Figure 1a), probably due to higher N, P, K and micronutrients and organic matter, that improved aggregation and soil structure for better aeration, soil quality and maintained or increased soil pH in acidic soils [50]. P deficiency restricts leaf growth, photosynthesis and eventually forage yield [18]. Recent work reported that P deficiency reduces the orthophosphate concentration in the chloroplast stroma which may lead to inhibit the process of ATP synthesis [51]. Previous research also demonstrated DMP amendment significantly improved photosynthesis and forage yield compared to mineral fertilizers and was attributed to optimum and slow release of nutrients [52]. Moreover, DMP release organic acids which play an important role in the supply of NH4+, Mg2+ and, Fe2+ which are necessary for enzyme activation, chloroplast and chlorophyll formation [53]. Furthermore, DMP application improved optimum supply of soil organic C, N and Mg2+ to crop plants which may lead to increased chlorophyll contents, photosynthesis and forage yield of silage-corn because both compounds are central components of chlorophyll molecules. In the present study, we observed that DMP1 amendment produced significantly higher forage yield compared to inorganic P, largely through increased leaf area, chlorophyll contents and photosynthesis (data not shown), which consequently boosted forage yield.
Yukon-R and DKC26-28RIB showed superior agronomic performance and produced 29% and 21% higher forage yield compared to A4177G3-RIB, which produced lowest forage yield (Figure 1b) but showed superior forage qualitative traits (Figure 8b). Higher forage yield in Yukon-R and DKC26-28RIB might be due their plant heights (Table 3), genetic potential, resilience, and better adaptation in boreal climate [54], as both these genotypes acquired similar heating units (2150) to reach physiological maturity compared to other genotypes (2075 and 2200). A field study conducted in similar climate and soil conditions also observed variation in forage yield in three silage-corn genotypes [39]. However, forage yield of Yukon-R and DKC26-28RIB tested in present study was significantly higher, suggesting better resilience, adaptation, improved genetic material and management practices (Figure 1a,b and Table 2 and Table 3). Therefore, Yukon-R and DKC26-28RIB were segregated in same quadrant (Figure 8a) and could be good choices for dairy and livestock farmers to attain high forage production under boreal climate.
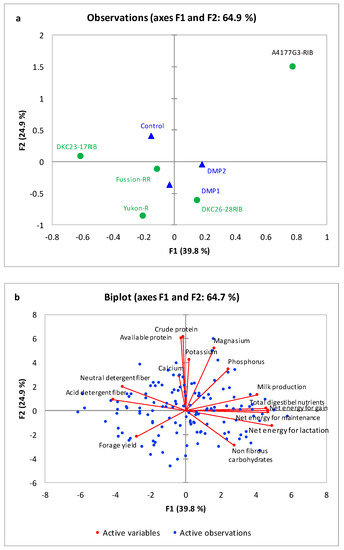
Figure 8.
Principal component analysis (PCA) observation plot (a) and biplot (b) of the first two components performed on the forage yield and forage quality of five silage-corn genotypes affected by organic (DMP1 and DMP2) and inorganic P amendments. Centroids showing segregation of silage corn genotypes and P amendments on axis F1 and F2 based on forage production and forage quality.
4.2. Forage Quality
Forages contain minerals which play an important role in milk production, reproduction and energy provision in animals [55], skeleton formation, protein and lipids synthesis [56]. The performance of dairy animals is dependent on adequate supply of forage minerals which may vary due to soil fertility, fertilization strategies and crop maturity [57]. In the present study, we observed significantly (p = 0.0) higher P uptake with inorganic P application compared to DMP1 treatment while non-significant differences in Ca, K and Mg uptake under inorganic and organic P amendments, although slightly higher uptake with inorganic P application (Figure 2a–d). Manure contains iron that might have reacted with P and formed iron phosphates thereby reducing P solubility. Reduction of Ca uptake in manure could be due to cation competition with K [58]. However, a greenhouse study conducted by Leytem et al. [58] reported increased P, Ca, Mg and K uptake in silage-corn with inorganic P application, while manure and compost treatments showed non-significant difference. Silage-corn genotypes showed significant variation in P, K and Mg and non-significant variation in Ca minerals uptake (Figure 3a–d). Mg and P concentration in forages vary between 1.8–3.6 g kg−1 and 1.8–3.3 g kg−1 of dry matter respectively, and are significantly affected by forage type, growth conditions, and harvesting stage [55,59]. Roth and Heinrichs [60] observed 0.2% P and 0.2% Mg in silage-corn, whereas in the present study we observed up to 0.3% P and 0.1–0.2% Mg in silage-corn genotypes (Figure 3a,d), and was either on par or slightly higher than reported in literature [30].
Silage-corn is generally low in Ca (1.4–3.0 g kg−1), whereas K was up to 1.2% on dry weight basis [59]. In the present study, we found Ca ranges between 1.2–1.6 g kg−1 and K between 12–14 g kg−1 among five silage-corn genotypes (Figure 3b). It is interesting to note that under our three-year field experiment, all five-tested silage-corn genotypes expressed slightly higher or similar range of P, Ca and K and were consistent with reports in the literature [30,60,61]. A4177G3-RIB genotype exhibited superior forage quality indices and produced significantly higher P, K and Mg; however, it was statistically on par with DKC26-28-RIB and ranked as high forage quality genotype according to the literature.
Proteins are organic compounds, composed of amino acid and are important animal feed component. Phosphorus is a part of nucleic acid, which is known to regulate protein synthesis. Organic based fertilizers contain N, P and micro-nutrients but generally they are slowly available to plants, whereas macro and micro-nutrients are readily available to the plants through inorganic fertilizer application which may affect the composition of forage quality. In the present study, we observed significantly higher available protein (AP) and crude protein (CP) with inorganic P compared to organic P amendment (Figure 4a,b). Conversely, Moreno-Resendez et al. [27] reported increased protein in forage treated with organic source compared to inorganic fertilizer. According to literature, protein contents in silage-corn vary between 5.2–7.5% [30], whereas, in good quality forage, CP should be more than 7% [62]. AP and CP contents in the present study were >10% in A4177G3-RIB (Figure 4a,b) and were 9% in Yukon-R among five genotypes, which were 36% higher than reported in the literature [27,61,62], and therefore, can be considered as high quality forage genotypes [62]. However, it appears a trade-off between AP, CP and forage production in silage corn genotypes [15,63]; therefore, a strong negative correlation between forage production and AP (r = −0.9), and CP (r = −0.9) was noted (Figure 1c,d), which might be due to N dilution effects [64,65].
ADF is comprised of cellulose and lignin which determines the forage quality [27]. This is the least digestible part and increased ADF result in low forage digestibility and energy contents [66,67]. The N-based organic amendment gave higher ADF contents, whereas P based organic treatments produced similar ADF contents to inorganic treatment. This could be attributed to differences in N contents among these treatments because they strongly influence fiber content as well [68]. In the present study, we observed non-significant in ADF in response to organic and inorganic P amendments (Figure 5a). Consistent with our findings, Nazli et al. [69] observed non-significant effects of P based cattle manure, poultry litter and inorganic fertilizer on ADF contents in silage-corn. NDF is of prime importance in ration formulation, as it comprised of ADF plus hemicellulose and they reflect the animal forage intake [27,70]. Organic amendment produced higher NDF contents (53.5%), whereas inorganic fertilizer produced lower (42.6%) NDF in silage-corn forage [27]. In the present study, we observed non-significant difference in NDF with inorganic and organic P based amendments (Figure 5b). NFC contents are more digestible than fiber, and therefore, a significant source of instant energy for rumen microbes [70,71]. For example, volatile fatty acids are produced during the NFC fermentation process and are assimilated by the rumens and utilized as a source of energy for maintenance, synthesis of milk and body tissues [72]. Moreno-Reséndez et al. [27] observed higher NFC with inorganic compared to lower with organic amendment in forage maize. However, in the present study, we observed non-significant effects of organic and inorganic P amendments on NFC (Figure 5c). Good quality forage crops contain ADF and NDF less than 28%, and 50%, and NFC > 30% [27,66,67]. Observed ADF, NDF and NFC in our experiment were 28%, 49.6% and >30% in DKC26-28-RIB (Figure 5d–f), which meets the criteria of good quality forage as reported in literature [27,66,67]. ADF, NDF and NFC contents produced by Yukon-R were very close to DKC26-28-RIB (Figure 5d–f). Manitoba corn committee observed 28.7% and 30.5% ADF in Yukon-R and Fusion RR, whereas in the present study we observed 30.5% and 31.5% ADF in the same genotypes. Lower ADF, NDF and higher NFC in DKC26-28-RIB explained the superior forage quality cultivated in boreal climate. Interestingly, Yukon-R showed ADF slightly higher above 30% and DKC26-28RIB produced 28% and were below 30%; they were therefore, ranked as high-quality forage genotypes, as these both produced higher forage production (Figure 1b and Figure 5d).
NEM and NEG represent the quality of forage species [73]. Forages NEM keep the animals at a stable weight, whereas NEG provides energy for body maintenance [71]. Inorganic and organic P amendments had non-significant effects on NEM, NEG and NEL, whereas silage-corn genotypes showed significant variations in NEM and NEG and non-significant in NEL (Table 2 and Figure 6). Literature does not provide any information about the effects of organic and inorganic P amendments on NEM, NEG and NEL. Observed NEM (1.3–1.4 Mcal kg−1) and NEG (up to 0.8 Mcal kg−1) among silage-corn genotypes used in the present study were slightly lower than those observed by Ballard et al. [61], which were 1.5–1.7 Mcal kg−1 NEM and 0.9–1.1 Mcal kg−1 NEG of dry matter. Manitoba corn committee observed 1.0 and 0.9 Mcal kg−1 NEG in Yukon-R and Fusion-RR. However, observed NEG in Yukon-R and Fusion-RR in the present study were 0.8 and 0.8 Mcal kg−1 respectively, which were slightly lower than Manitoba research trials. This slight decrease in NEM and NEG might be due to the crop harvesting stage where forage energy can be reduced due to increased forage fiber contents and slightly lower TDN [32,74]. Significantly higher NEM and NEG were observed in A4177G3-RIB (1.4 and 0.8 Mcal kg−1) and lower were noted in DKC26-28-RIB (1.3 and 0.8 Mcal kg−1) which were very similar to that reported in literature [61]. Furthermore, the observed differences in NEG and NEM among five silage-corn genotypes could be due to differences in proteins, fibers and TDN (Figure 4c,d, Figure 5d–f, Figure 7c). NEL is an important energy forage quality parameter estimated using TDN, CP, and NDF values [46]. NEL is instrumental in animal’s body maintenance as well as milk production during lactation period [70] and vary between 1.3–1.5 Mcal kg−1 of dry matter in silage-corn [32,75,76], whereas we observed 1.4–1.4 Mcal kg−1 of dry matter among five silage-corn genotypes, although statistically non-significant (Figure 6f). In Manitoba corn research trials, Yukon-R and Fusion-RR produced 1.6 and 1.5 Mcal kg−1 NEL respectively, while, in the present study, observed NEL values were 1.4 and 1.4 Mcal kg−1 in Yukon-R and Fusion-RR (Figure 6f).
TDN is a measure of forage energy as well as the forage digestibility in the animals [46,77,78]. Inorganic and organic P amendments had non-significant effects on TDN and calculated milk production, whereas silage corn genotypes showed significant variations in TDN and calculated milk production (Figure 7a–d). Silage-corn dry matter digestibility is an indication of energy contents or TDN; higher digestibility depends on hybrid, planting date, environmental conditions, and maturity at harvest [40]. Good quality forage must contain TDN equal or greater than 65% [79], however, observed TDN in present study among five silage-corn genotypes ranged approximately between 62–64% (Figure 7c), although significantly higher in A4177G3RIB (64%) which falls within the range of good quality forage as reported above. Kim et al. [78] found higher TDN when harvested between 106‒111 DAS (days after sowing) compared to lower TDN harvested between 119‒125 DAS, possibly due to increased forage fiber contents. Manitoba Corn research trials exhibited 67.7, 66.4 and 67.1% TDN in A4177G3-RIB, Yukon-R and Fusion-RR respectively, which were slightly higher than observed in the present experiment in the same genotypes (63.9, 62.9 and 63.6%). Lower TDN contents in present study might be due to late harvest (130 DAS), which enhanced fibers contents resulted in reduced TDN [78]. Higher calculated milk production in A4177G3-RIB could be due to higher TDN (Figure 7c), available protein (Figure 4c), NEM (Figure 6d), NEG (Figure 6e), minerals (Figure 3a–d), which eventually enhanced calculated milk production. Superior forage quality of genotypes tested in boreal climate might have led to higher calculated milk production and associated quality parameters.
5. Conclusions
This study demonstrated that DMP1 (high N, P and K) produced higher forage yield compared to inorganic P. Contrarily, inorganic P amendment exhibited higher P mineral and protein contents compared to DMP1 but on par with DMP2 (low N, P and K), whereas non-significant differences in Ca, Mg and K minerals, fibers, NFC, energy parameters, TDN and milk production were shown under inorganic and organic P amendments. Among genotypes, Yukon-R and DKC26-28RIB showed superior agronomic performance over three growing seasons and produced significantly higher forage yield, whereas A4177G3-RIB genotype produced lowest forage yield but exhibited superior nutritional quality indices suggesting some tradeoff between forage yield and quality traits. It is pertinent to mention here that DKC26-28RIB produced P, K, AP, and NFC which were either comparable or close to A4177G3-RIB genotype followed by Yukon-R. In sum, we can conclude that Yukon-R cultivation following DM amendment could be a sustainable and viable agricultural production practice to attain high forage yield with optimum nutritional quality to meet the forage needs of growing dairy industry in boreal climate.
Author Contributions
M.C. conceptualized, funding acquisition and supervised the overall research project; M.N., V.K. and W.A. performed the layout and seeding of field experiment; W.A., M.N., W.A., M.Z. data collection. R.T., M.Z. and W.A. performed analysis. W.A., and M.N. wrote the first draft, while M.N., R.T. and M.C. edited and interpreted the results and discussion of the manuscript.
Funding
This work was supported by research grants from Research & Development Corporation of Newfoundland and Labrador, and Atlantic Corporation Opportunities Agency (ACOA), Canada.
Acknowledgments
Authors are thankful to Adrian Reid and Danny Brock for assisting in field operations and seeding, Sanaz Rajabi for data collection and Tom Halford for proofreading of manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Tilman, D.; Balzer, C.; Hill, J.; Befort, B.L. Global food demand and the sustainable intensification of agriculture. Proc. Natl. Acad. Sci. USA 2011, 108, 20260–20264. [Google Scholar] [CrossRef] [PubMed]
- Fedoroff, N.V. Food in a future of 10 billion. Agric. Food Secur. 2015, 4, 11. [Google Scholar] [CrossRef]
- FAO. How to Feed the World in 2050. Available online: http://www.fao.org/fileadmin/templates/wsfs/docs/expert_paper/How_to_Feed_the_World_in_2050.pdf (accessed on 15 March 2019).
- Ittersum, V. Food Security in 2050 on a Global Scale Achievable but Greatly Challenging. Available online: https://phys.org/news/2011-05-food-global-scale-greatly.html (accessed on 9 May 2019).
- Ranum, P.; Pena-Rosas, J.P.; Garcia-Casal, M.N. Global maize production, utilization, and consumption. Ann. N. Y. Acad. Sci. 2014, 1312, 105–112. [Google Scholar] [CrossRef] [PubMed]
- Khan, N.A.; Yu, P.; Ali, M.; Cone, J.W.; Hendriks, W.H. Nutritive value of maize silage in relation to dairy cow performance and milk quality. J. Sci. Food Agric. 2015, 95, 238–252. [Google Scholar] [CrossRef] [PubMed]
- Rankin, M. Choosing between Alfalfa and Corn Silage; University of Wisconsin-Extension: Madison, WI, USA, 2014. [Google Scholar]
- Kirkland, R.M.; Steen, R.W.J.; Gordon, F.J.; Keady, T.W.J. The influence of grass and maize silage quality on apparent diet digestibility, metabolizable energy concentration and intake of finishing beef cattle. Grass Forage Sci. 2005, 60, 244–253. [Google Scholar] [CrossRef]
- Karsten, H.D.; Roth, G.W.; Muller, L.D. Evaluation of corn hybrids at two stages of development for grazing heifers. Agrono. J. 2003, 95, 870–877. [Google Scholar] [CrossRef]
- Fitzgerald, J.J.; Murphy, J.J. A comparison of low starch maize silage and grass silage and the effect of concentrate supplementation of the forages or inclusion of maize grain with the maize silage on milk production by dairy cows. Livest. Prod. Sci. 1999, 57, 95–111. [Google Scholar] [CrossRef]
- Martin, N.P.; Russelle, M.P.; Powell, J.M.; Sniffen, C.J.; Smith, S.I.; Tricarico, J.M.; Grant, R.J. Invited review: Sustainable forage and grain crop production for the US dairy industry. J. Dairy Sci. 2017, 100, 9479–9494. [Google Scholar] [CrossRef]
- Hart, J.M.; Marx, E.S.; Christensen, N.W.; Moore, J.A. Nutrient management strategies. J. Dairy Sci. 1997, 80, 2659–2666. [Google Scholar] [CrossRef]
- Walsh, J.J.; Jones, D.L.; Edwards-Jones, G.; Williams, A.P. Replacing inorganic fertilizer with anaerobic digestate may maintain agricultural productivity at less environmental cost. J. Plant Nutr. Soil Sci. 2012, 175, 840–845. [Google Scholar] [CrossRef]
- Agriculture, N.L. A Factsheet Series on Environmental Guidelines for Livestock Producers; Agriculture, Newfoundland and Labrador; Publication SLM 045; Department of Forest Resources and Agrifoods: Carbonear, NL, Canada, 2002. [Google Scholar]
- Seleiman, M.F.; Selim, S.; Jaakkola, S.; Mäkelä, P.S.A. Chemical composition and in vitro digestibility of whole-crop maize fertilized with synthetic fertilizer or digestate and harvested at two maturity stages in Boreal growing conditions. Agric. Food Sci. 2017, 26, 47–55. [Google Scholar] [CrossRef]
- Seleiman, M.F.; Santanen, A.; Stoddard, F.L.; Makela, P. Feedstock quality and growth of bioenergy crops fertilized with sewage sludge. Chemosphere 2012, 89, 1211–1217. [Google Scholar] [CrossRef] [PubMed]
- Glass, A.D.M.; Beaton, J.D.; Bomke, A. Role of P in plant nutrition. In Proceedings of the Western Canada Phosphate Symposium, Calgary, AB, Canada, 11–12 March 1980; pp. 357–368. [Google Scholar]
- Plénet, D.; Etchebest, S.; Mollier, A.; Pellerin, S. Growth analysis of maize field crops under phosphorus deficiency-I. Leaf growth. Plant Soil 2000, 223, 117–130. [Google Scholar] [CrossRef]
- Assuero, S.G.; Mollier, A.; Pellerin, S. The decrease in growth of phosphorus-deficient maize leaves is related to a lower cell production. Plant Cell Environ. 2004, 27, 887–895. [Google Scholar] [CrossRef]
- Raghothama, K.G. Phosphate acquisition. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1999, 50, 665–693. [Google Scholar] [CrossRef] [PubMed]
- Kulhánek, M.; Balík, J.; Černý, J.; Nedvěd, V.; Kotková, B. The influence of different intensities of phosphorus fertilizing on available phosphorus contents in soils and uptake by plants. Plant Soil Environ. 2007, 53, 382–387. [Google Scholar] [CrossRef]
- Hinsinger, P. Bioavailability of soil inorganic P in the rhizosphere as affected by root-induced chemical changes: A review. Plant Soil 2001, 237, 173–195. [Google Scholar] [CrossRef]
- Vance, C.P.; Uhde-Stone, C.; Allan, D.L. Phosphorus acquisition and use: Critical adaptations by plants for securing a nonrenewable resource. New Phytol. 2003, 157, 423–447. [Google Scholar] [CrossRef]
- Cordell, D.; Drangert, J.O.; White, S. The story of phosphorus: Global food security and food for thought. Glob. Environ. Chang. 2009, 19, 292–305. [Google Scholar] [CrossRef]
- Newton, G.L.; Bernard, J.K.; Hubbard, R.K.; Allison, J.R.; Lowrance, R.R.; Gascho, G.J.; Gates, R.N.; Vellidis, G. Managing manure nutrients through multi-crop forage production. J. Dairy Sci. 2003, 86, 2243–2252. [Google Scholar] [CrossRef]
- Cherney, J.H.; Cherney, D.J.R. Impact of fertilization on forage production and animal performance. In Proceedings of the Colloque sur les plantes fourrageres, Drummondville, QC, Canada, 9 December 2015; pp. 1–7. [Google Scholar]
- Moreno-Reséndez, A.; Cantú Brito, J.E.; Reyes-Carrillo, J.L.; Contreras-Villarreal, V. Forage maize nutritional quality according to organic and inorganic fertilization. Sci. Agropecu. 2017, 8, 127–135. [Google Scholar] [CrossRef][Green Version]
- Corrales, I.; Amenós, M.; Poschenrieder, C.; Barceló, J. Phosphorus efficiency and root exudates in two contrasting tropical maize varieties. J. Plant Nutr. 2007, 30, 887–900. [Google Scholar] [CrossRef]
- Ozturk, L.; Eker, S.; Torun, B.; Cakmak, I. Variation in phosphorus efficiency among 73 bread and durum wheat genotypes grown in a phosphorus-deficient calcareous soil. Plant Soil 2005, 269, 69–80. [Google Scholar] [CrossRef]
- Amodu, J.T.; Akpensuen, T.T.; Dung, D.D.; Tanko, R.J.; Musa, A.; Abubakar, S.A.; Hassan, M.R.; Jegede, J.O.; Sani, I. Evaluation of maize accessions for nutrients composition, forage and silage yields. J. Agric. Sci. 2014, 6, 178–187. [Google Scholar] [CrossRef]
- Faisal, S.; Shah, S.N.M.; Majid, A.; Khan, A. Effect of organic and inorganic fertilizers on protein, yield and related traits of maize varieties. Int. J. Agric. Crop Sci. 2013, 6, 1299–1303. [Google Scholar]
- Peña-Ramos, A.; Núñez-Hernández, G.; González-Castañeda, F. Forage potential of some maize populations and relationships between their agronomic characteristics and nutritional quality. Téc. Pecu. Méx. 2002, 40, 215–228. [Google Scholar]
- Eghball, B.; Power, J.F. Phosphorus- and nitrogen-based manure and compost applications: Corn production and soil phosphorus. Soil Sci. Soc. Am. J. 1999, 63, 895–901. [Google Scholar] [CrossRef]
- Hirzel, J.; Matus, I.; Novoa, F.; Walter, I.; Walter, I. Effect of poultry litter on silage maize (Zea mays L.) production and nutrient uptake. Span. J. Agric. Res. 2007, 5, 8. [Google Scholar] [CrossRef][Green Version]
- Toth, J.D.; Dou, Z.; Ferguson, J.D.; Galligan, D.T.; Ramberg, C.F. Nitrogen- vs. phosphorus-based dairy manure applications to field crops. J. Environ. Qual. 2006, 35, 2302–2312. [Google Scholar] [CrossRef]
- Group, S.C.W. The Canadian System of Soi1 Classification, 3rd ed.; Agriculture and Agrifoods Canada, National Research Council of Canada: Ottawa, ON, Canada, 1998. [Google Scholar]
- Kirby, G.E. Soils of the Pasadena-Deer Lake Area, Newfoundland; Soil and Land Irlanagement Division: St. John’s, NL, Canada, 1988; Available online: http://sis.agr.gc.ca/cansis/publications/surveys/nf/nf17/nf17report.pdf (accessed on 15 March 2019).
- Bal, M.A.; Coors, J.G.; Shaver, R.D. Impact of the maturity of corn for use as silage in the diets of dairy cows on intake, digestion, and milk production. J. Dairy Sci. 1997, 80, 2497–2503. [Google Scholar] [CrossRef]
- Kwabiah, A.B. Growth, maturity, and yield responses of silage maize (Zea mays L.) to hybrid, planting date and plastic mulch. J. New Seeds 2005, 7, 37–59. [Google Scholar] [CrossRef]
- Guyader, J.; Baron, V.S.; Beauchemin, K.A. Corn forage yield and quality for silage in short growing season areas of the Canadian Prairies. Agronomy 2018, 8, 164. [Google Scholar] [CrossRef]
- Min, D.H.; Vough, L.R.; Reeves, J.B. Dairy slurry effects on forage quality of orchardgrass, reed canarygrass and alfalfa-grass mixtures. Anim. Feed Sci. Technol. 2002, 95, 143–157. [Google Scholar] [CrossRef]
- Ferreira, G.; Alfonso, M.; Depino, S.; Alessandri, E. Effect of planting density on nutritional quality of green-chopped corn for silage. J. Dairy Sci. 2014, 97, 5918–5921. [Google Scholar] [CrossRef] [PubMed]
- Gallo, A.; Moschini, M.; Cerioli, C.; Masoero, F. Use of principal component analysis to classify forages and predict their calculated energy content. Anim. Int. J. Anim. Biosci. 2013, 7, 930–939. [Google Scholar] [CrossRef] [PubMed]
- Horwitz, W. Official methods of analysis. In The Association of Officeal Analytical Chemists; Benjamin Frankline Statio: Washington, DC, USA, 1980; p. 1018. [Google Scholar]
- Van Soest, P.J.; Fox, D.G. Discounts for net energy and protein-5th Revision. In Proceedings of the Cornell Nutrition Conference for Feed Manufacturers, Rochester, NY, USA, 13–15 October 1992; p. 40. [Google Scholar]
- Weiss, W.P.; Conrad, H.R.; St. Pierre, N.R. A theoretically-based model for predicting total digestible nutrient values of forages and concentrates. Anim. Feed Sci. Technol. 1992, 39, 95–110. [Google Scholar] [CrossRef]
- National, R.C. Nutrient Requirements of Dairy Cattle, 7th ed.; National Academy of Science: Washington, DC, USA, 2001. [Google Scholar]
- Norell, R.J. The Wisconsin system fo revaluating corn silage. In Proceedings of the 2005 Idaho Alfalfa and Forage Conference, Twin Falls, ID, USA, 7–8 February 2005; pp. 44–51. [Google Scholar]
- King, M.; Altdorff, D.; Li, P.; Galagedara, L.; Holden, J.; Unc, A. Northward shift of the agricultural climate zone under 21(st)-century global climate change. Sci. Rep. 2018, 8, 7904. [Google Scholar] [CrossRef]
- Yan, F.; Schubert, S.; Mengel, K. Soil pH changes during legume growth and application of plant material. Biol. Fertil. Soils 1996, 23, 236–242. [Google Scholar] [CrossRef]
- Carstensen, A.; Herdean, A.; Schmidt, S.B.; Sharma, A.; Spetea, C.; Pribil, M.; Husted, S. The Impacts of phosphorus deficiency on the photosynthetic electron transport chain. Plant Physiol. 2018, 177, 271–284. [Google Scholar] [CrossRef]
- Efthimiadou, A.; Bilalis, D.; Karkanis, A.; Froud-Williams, B. Combined organic/inorganic fertilization enhance soil quality and increased yield, photosynthesis and sustainability of sweet maize crop. Aust. J. Crop Sci. 2009, 4, 722–729. [Google Scholar]
- Elhindi, K. Evaluation of composted green waste fertigation through surface and subsurface drip irrigation systems on pot marigold plants (Calendula officinalis L.) grown on sandy soil. Aust. J. Crop Sci. 2012, 6, 1249–1259. [Google Scholar]
- Nadeem, M.; Pham, T.H.; Thomas, R.; Galagedara, L.; Kavanagh, V.; Zhu, X.; Ali, W.; Cheema, M. Potential role of root membrane phosphatidic acid in superior agronomic performance of silage-corn cultivated in cool climate cropping systems. Physiol. Plant. 2019. [Google Scholar] [CrossRef] [PubMed]
- Kronqvist, C. Minerals to Dairy Cows with Focus on Calcium and Magnesium Balance. Ph.D. Thesis, Swedish University of Agricultural Sciences, Uppsala, Sweden, 2011. [Google Scholar]
- Plaizier, J.C.; Garner, T.; Droppo, T.; Whiting, T. Nutritional practices on Manitoba dairy farms. Can. J. Anim. Sci. 2004, 84, 501–509. [Google Scholar] [CrossRef]
- Holtenius, K.; Kronqvist, C.; Briland, E.; Sporndly, R. Magnesium absorption by lactating dairy cows on a grass silage-based diet supplied with different potassium and magnesium levels. J. Dairy Sci. 2008, 91, 743–748. [Google Scholar] [CrossRef] [PubMed]
- Leytem, A.B.; Dungun, R.S.; Moore, A. Nutrient availability to corn from dairy manures and fertilizer in a calcareous soil. Soil Sci. 2011, 176, 1–9. [Google Scholar] [CrossRef]
- Suttle, N. Mineral Nutrition of Livestock, 4th ed.; CABI: Wallingford, UK, 2010. [Google Scholar]
- Roth, G.W.; Heinrichs, A.J. Maize Silage Production and Management; Agronomy fact-18; Information and Communication Technology, The College of Agricultural Sciences, Pennsylvania State University: State College, PA, USA, 2001. [Google Scholar]
- Ballard, C.S.; Thomas, E.D.; Tsang, D.S.; Mandebvu, P.; Sniffen, C.J.; Endres, M.I.; Carter, M.P. Effect of corn silage hybrid on dry matter yield, nutrient composition, in vitro digestion, intake by dairy heifers, and milk production by dairy cows. J. Dairy Sci. 2001, 84, 442–452. [Google Scholar] [CrossRef]
- Garibay, S.V.; Stamp, P.; Ammon, H.U.; Feil, B. Yield and quality components of silage maize in killed and live cover crops sods. Eur. J. Agron. 1997, 6, 179–190. [Google Scholar] [CrossRef]
- Seleiman, M.F.; Santanen, A.; Kleemola, J.; Stoddard, F.L.; Mäkelä, P.S.A. Improved sustainability of feedstock production with sludge and interacting mycorrhiza. Chemosphere 2013, 91, 1236–1242. [Google Scholar] [CrossRef] [PubMed]
- Fletcher, A.L.; Chakwizira, E. Developing a critical nitrogen dilution curve for forage brassicas. Grass Forage Sci. 2012, 67, 13–23. [Google Scholar] [CrossRef]
- Bélanger, G.; Gastal, F. Nitrogen utilization by forage grasses. Can. J. Plant Sci. 2000, 80, 11–20. [Google Scholar] [CrossRef]
- Castillo-Jimenez, M.; Roja-Bourrillon, A.; WingChing-Jones, R. Nutritional value of silage made with a mixture of corn and mung bean (Vigna radiata). Agron. Costarric. 2009, 33, 133–146. [Google Scholar]
- Gallegos-Ponce, A.; Martínez-Ríos, A.; Fernando-Sánchez, M.; Figueroa-Viramontes, R.; Berumen-Padilla, S.; Venegas-Soto, J.; Quevedo-Guillen, J.D.D.; Escobedo-López, D.; Silos-Calzada, M.C. Nutritional quality of forage maize (Zea mays L.) under limited water logging conditions. Agrofaz 2012, 12, 59–66. [Google Scholar]
- Johnson, C.R.; Reiling, B.A.; Mislevy, P.; Hall, M.B. Effects of nitrogen fertilization and harvest date on yield, digestibility, fiber, and protein fractions of tropical grasses. J. Anim. Sci. 2001, 79, 2439–2448. [Google Scholar] [CrossRef] [PubMed]
- Nazli, R.I.; Inal, I.; Kusvuran, A.; Demirbas, A.; Tansi, V. Effects of different organic materials on forage yield and nutrient uptake of silage maize (Zea mays L.). J. Plant Nutr. 2016, 39, 912–921. [Google Scholar] [CrossRef]
- Schwab, E.C.; Shaver, R.D.; Lauer, J.G.; Coors, J.G. Estimating silage energy value and milk yield to rank corn hybrids. Anim. Feed Sci. Technol. 2003, 109, 1–18. [Google Scholar] [CrossRef]
- DePeters, E.J.; Fadel, J.G.; Arana, M.J.; Ohanesian, N.; Etchebarne, M.A.; Hamilton, C.A.; Hinders, R.G.; Maloney, M.D.; Old, C.A.; Riordan, T.J.; et al. Variability in the chemical composition of seventeen selected by-product feedstuffs used by the California dairy industry. Prof. Anim. Sci. 2000, 16, 69–99. [Google Scholar] [CrossRef]
- Harris, B., Jr. CIR1122 Nonstructrual and Structural Carbohydrates in Dairy Cattle Rations; University of Florida IFAS Extension: Gainesville, FL, USA, 1993; pp. 1–5. [Google Scholar]
- Tine, M.A.; McLeod, K.R.; Erdman, R.A.; Baldwin, R.L.T. Effects of brown midrib corn silage on the energy balance of dairy cattle. J. Dairy Sci. 2001, 84, 885–895. [Google Scholar] [CrossRef]
- Di Marco, O.N.; Aello, M.S.; Nomdedeu, M.; Van Houtte, S. Effect of maize crop maturity on silage chemical composition and digestibility (in vivo, in situ and in vitro). Anim. Feed Sci. Technol. 2002, 99, 37–43. [Google Scholar] [CrossRef]
- de la Cruz-Lázaro, E.; Rodríguez-Herrera, S.A.; Palomo-Gil, A.; López Benítez, A.; Robledo-Torres, V.; Gómez-Vázquez, A.; Osorio-Osorio, R. Combining ability of protein high quality maíz inbred lines for forage characteristics. Ecosistemas Recur. Agropecu. 2007, 23, 57–67. [Google Scholar]
- Núñez-Hernández, G.; Contreras, G.F.; Faz-Contreras, R. Yield, chemical composition and in vitro digestibility of tropical and temperate corn hybrids in the arid region of Mexico. Av. Investig. Agropecu. 2004, 8, 1–19. [Google Scholar]
- Posada, O.S.; Rosero, N.R.; Rodríguez, N.; Costa, C.A. Comparison of methods to determine the energy value of feeds for ruminants. Rev. MVZ Córdoba 2012, 17, 3184–3192. [Google Scholar]
- Kim, J.D.; Kwon, C.H.; Kim, D.A. Yield and quality of silage corn as affected by hybrid maturity, Pplanting date and harvest stage. Asian Australas. J. Anim. Sci. 2001, 14, 1705–1711. [Google Scholar] [CrossRef]
- Olague-Ramirez, J.; Montemayor-Trejo, J.A.; Bravo-Sánchez, S.R.; Fortis-Hernández, M.; Aldaco-Nuncio, R.A.; Ruiz-Cerda, E. Agronomic characteristic and forage quality corn with subsurface drip irrigation. Tec. Pecu. Mex. 2006, 44, 351–357. [Google Scholar]
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).