Isolation and Characterization of the PISTILLATA Ortholog Gene from Cymbidium faberi Rolfe

Abstract
1. Introduction
2. Results
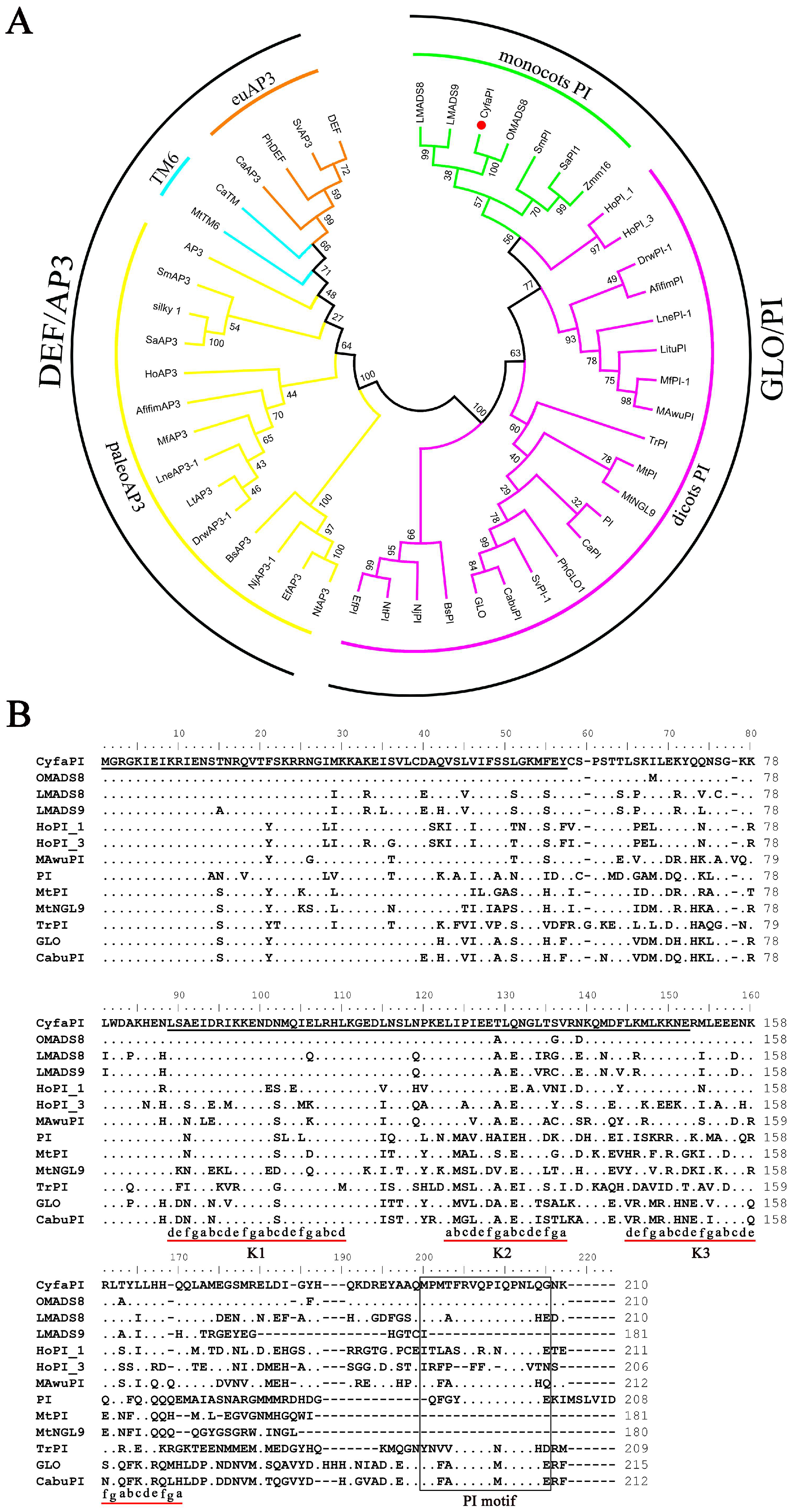
2.1. Cloning and Sequence Alignments of CyfaPI
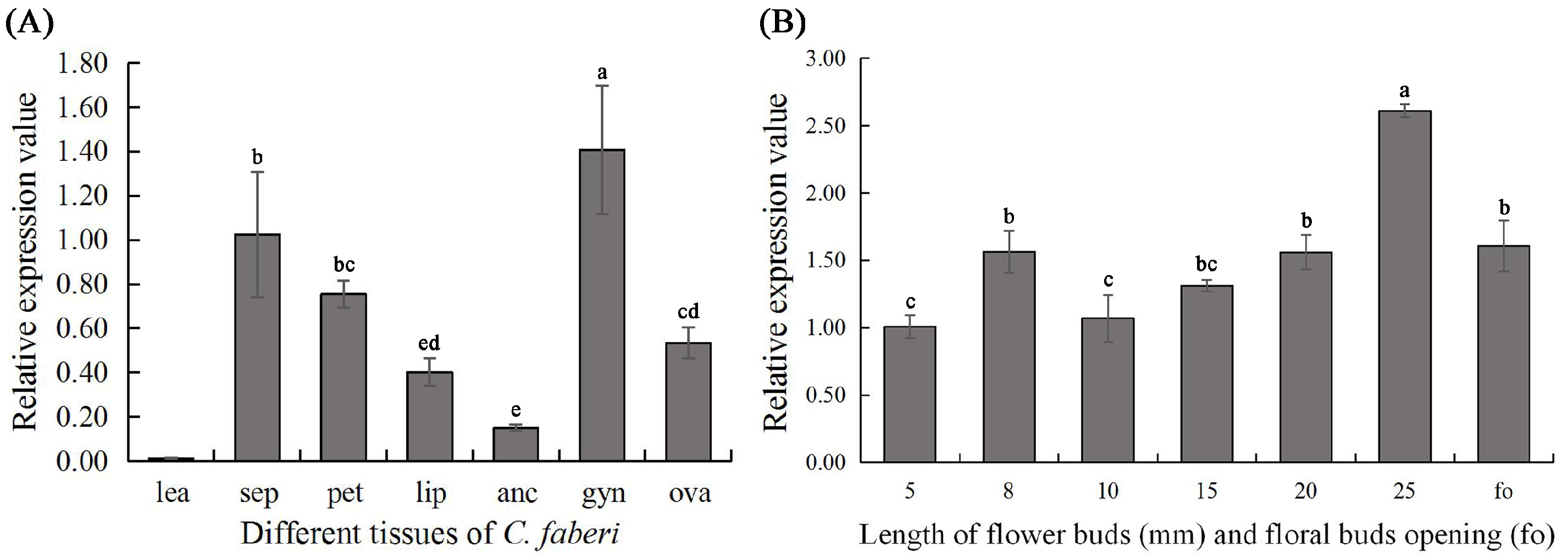
2.2. Expression Analyses of CyfaPI in C. faberi
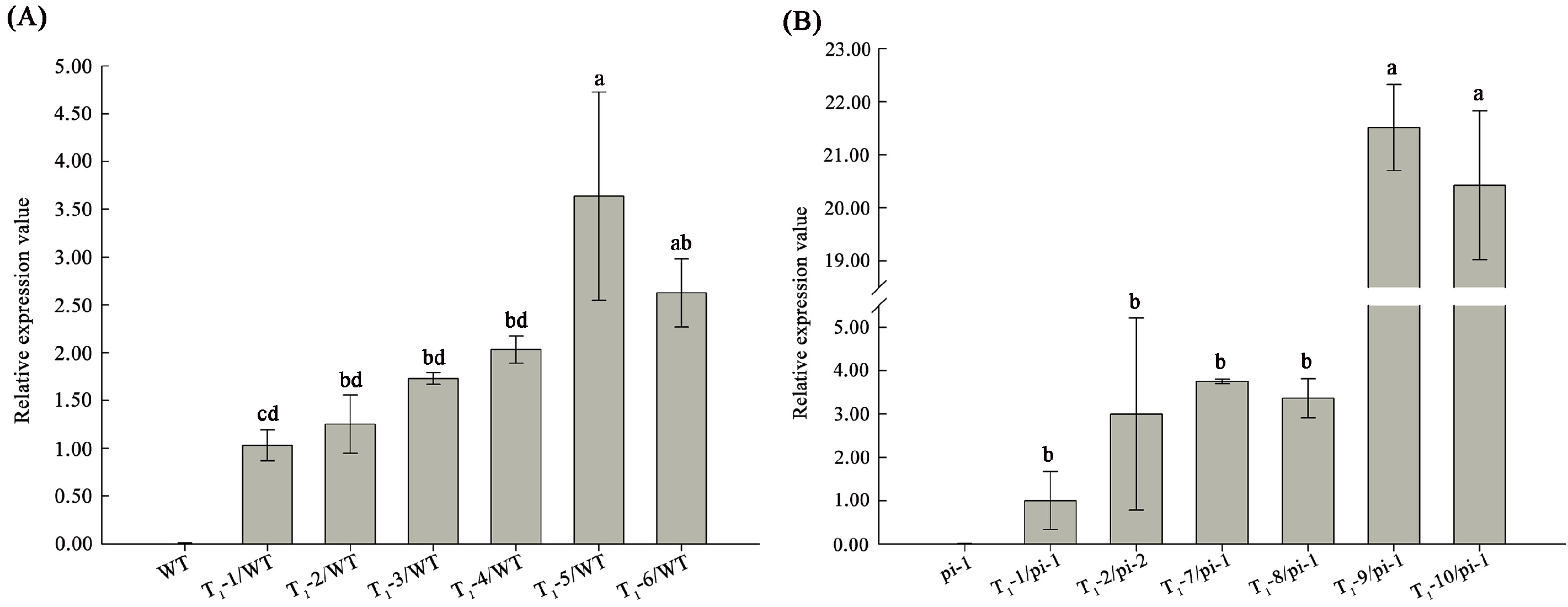
2.3. Ectopic Expression of CyfaPI in Arabidopsis pi-1 Mutant
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. Isolation and Characterization of PI Ortholog from C. faberi
4.3. Expression Analysis of CyfaPI
4.4. Ectopic Expression of CyfaPI in Arabidopsis pi-1 Mutant and Function Analysis
4.5. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| BLAST | Basic local alignment search tool |
| PCR | Polymerase chain reaction |
| RT-qPCR | Quantitative reverse transcription PCR |
| LSD | Least significant difference |
References
- Heijmans, K.; Morel, P.; Vandenbussche, M. MADS-box Genes and Floral Development: The Dark Side. J. Exp. Bot. 2012, 63, 5397–5404. [Google Scholar] [CrossRef] [PubMed]
- Monniaux, M.; Vandenbussche, M. How to Evolve a Perianth: A Review of Cadastral Mechanisms for Perianth Identity. Front. Plant Sci. 2018, 9, 1573. [Google Scholar] [CrossRef] [PubMed]
- Hecht, V. Duplicate MADS genes with split roles. J. Exp. Bot. 2016, 67, 1609–1611. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Lü, S.; Fan, Y.; Liu, L.; Liu, S.; Zhang, W.; Meng, Z. Ectopic expression of TrPI, a Taihangia rupestris (Rosaceae) PI ortholog, causes modifications of vegetative architecture in Arabidopsis. J. Plant Physiol. 2010, 167, 1613–1621. [Google Scholar] [CrossRef] [PubMed]
- Jing, D.; Xia, Y.; Chen, F.; Wang, Z.; Zhang, S.; Wang, J. Ectopic expression of a Catalpa bungei (Bignoniaceae) PISTILLATA homologue rescues the petal and stamen identities in Arabidopsis pi-1 mutant. Plant Sci. 2015, 231, 40–51. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Sun, Y.; Du, X.; Xu, Q.; Wu, F.; Meng, Z. Analysis of the APETALA3- and PISTILLATA-like genes in Hedyosmum orientale (Chloranthaceae) provides insight into the evolution of the floral homeotic B-function in angiosperms. Ann. Bot. 2013, 112, 1239–1251. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Shen, X.; Liang, H.; Wang, Y.; He, Z.; Zhang, D.; Chen, F. Isolation and Functional Analysis of PISTILLATA Homolog from Magnolia wufengensis. Front. Plant Sci. 2018, 9, 1743. [Google Scholar] [CrossRef]
- Aceto, S.; Gaudio, L. The MADS and the Beauty: Genes Involved in the Development of Orchid Flowers. Curr. Genom. 2011, 12, 342–356. [Google Scholar] [CrossRef]
- Hsu, H.F.; Hsu, W.H.; Lee, Y.I.; Mao, W.T.; Yang, J.Y.; Li, J.Y.; Yang, C.H. Model for perianth formation in orchids. Nat. Plants 2015, 1, 15046. [Google Scholar] [CrossRef]
- Riechmann, J.L.; Krizek, B.A.; Meyerowitz, E.M. Dimerization specificity of Arabidopsis MADS domain homeotic proteins APETALA1, APETALA3, PISTILLATA, and AGAMOUS. Proc. Natl. Acad. Sci. USA 1996, 93, 4793–4798. [Google Scholar] [CrossRef]
- Yang, Y.; Jack, T. Defining subdomains of the K domain important for protein-protein interactions of plant MADS proteins. Plant Mol. Biol. 2004, 55, 45–59. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Yoo, M.J.; Albert, V.A.; Farris, J.S.; Soltis, P.S.; Soltis, D.E. Phylogeny and diversification of B-function MADS-box genes in angiosperms: Evolutionary and functional implications of a 260-million-year-old duplication. Am. J. Bot. 2004, 91, 2102–2118. [Google Scholar] [CrossRef] [PubMed]
- Ma, H.; Depamphilis, C. The ABCs of Floral Evolution. Cell 2000, 101, 5–8. [Google Scholar] [CrossRef]
- Goto, K.; Meyerowitz, E.M. Function and regulation of the Arabidopsis floral homeotic gene PISTILLATA. Genes Dev. 1994, 8, 1548–1560. [Google Scholar] [CrossRef] [PubMed]
- Roque, E.; Fares, M.A.; Yenush, L.; Rochina, M.C.; Wen, J.; Mysore, K.S.; Gomez-Mena, C.; Beltrán, J.P.; Cañas, L.A. Evolution by gene duplication of Medicago truncatula PISTILLATA-like transcription factors. J. Exp. Bot. 2016, 67, 1805–1817. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Chen, M.K.; Hsieh, W.P.; Yang, C.H. Functional analysis reveals the possible role of the C-terminal sequences and PI motif in the function of lily (Lilium longiflorum) PISTILLATA (PI) orthologues. J. Exp. Bot. 2012, 63, 941–961. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.Y.; Kao, N.H.; Li, J.Y.; Hsu, W.H.; Liang, Y.L.; Wu, J.W.; Yang, C.H. Characterization of the possible roles for B class MADS box genes in regulation of perianth formation in orchid. Plant Physiol. 2010, 152, 837–853. [Google Scholar] [CrossRef]
- Mao, W.T.; Hsu, H.F.; Hsu, W.H.; Li, J.Y.; Lee, Y.I.; Yang, C.H. The C-Terminal Sequence and PI motif of the Orchid (Oncidium Gower Ramsey) PISTILLATA (PI) Ortholog Determine its Ability to Bind AP3 Orthologs and Enter the Nucleus to Regulate Downstream Genes Controlling Petal and Stamen Formation. Plant Cell Physiol. 2015, 56, pcv129. [Google Scholar]
- Tsai, W.C.; Lee, P.F.; Chen, H.I.; Hsiao, Y.Y.; Wei, W.J.; Pan, Z.J.; Chuang, M.H.; Kuoh, C.S.; Chen, W.H.; Chen, H.H. PeMADS6, a GLOBOSA/PISTILLATA-like Gene in Phalaenopsis equestris Involved in Petaloid Formation, and Correlated with Flower Longevity and Ovary Development. Plant Cell Physiol. 2005, 46, 1125–1139. [Google Scholar] [CrossRef]
- Hsieh, M.H.; Lu, H.C.; Pan, Z.J.; Yeh, H.H.; Wang, S.S.; Chen, W.H.; Chen, H.H. Optimizing virus-induced gene silencing efficiency with Cymbidium mosaic virus in Phalaenopsis flower. Plant Sci. 2013, 201, 25–41. [Google Scholar] [CrossRef]
- Tsai, W.C.; Pan, Z.J.; Hsiao, Y.Y.; Jeng, M.F.; Wu, T.F.; Chen, W.H.; Chen, H.H. Interactions of B-class complex proteins involved in tepal development in Phalaenopsis orchid. Plant Cell Physiol. 2008, 49, 814–824. [Google Scholar] [CrossRef] [PubMed]
- Pan, Z.J.; Cheng, C.C.; Tsai, W.C.; Chung, M.C.; Chen, W.H.; Hu, J.M.; Chen, H.H. The Duplicated B-class MADS-Box Genes Display Dualistic Characters in Orchid Floral Organ Identity and Growth. Plant Cell Physiol. 2011, 52, 1515–1531. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Salemme, M.; Sica, M.; Gaudio, L.; Aceto, S. Expression pattern of two paralogs of the PI/GLO-like locus during Orchis italica (Orchidaceae, Orchidinae) flower development. Dev. Genes Evol. 2011, 221, 241–246. [Google Scholar] [CrossRef] [PubMed]
- Cantone, C.; Gaudio, L.; Aceto, S. The PI/GLO-like locus in orchids: Duplication and purifying selection at synonymous sites within Orchidinae (Orchidaceae). Gene 2011, 481, 48–55. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Zhang, D.; Liu, D.; Li, F.; Lu, H. Exon skipping of AGAMOUS homolog PrseAG in developing double flowers of Prunus lannesiana (Rosaceae). Plant Cell Rep. 2013, 32, 227–237. [Google Scholar] [CrossRef]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular Evolutionary Genetics Analysis Using Maximum Likelihood, Evolutionary Distance, and Maximum Parsimony Methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Clough, S.J.; Bent, A.F. Floral dip: A simplified method forAgrobacterium-mediated transformation ofArabidopsis thaliana. Plant J. 1998, 16, 735–743. [Google Scholar] [CrossRef]
- Li, L.Y.; Fang, Z.W.; Liu, Z.X. Isolation and characterization of the C-class MADS-box gene from the distylous pseudo-cereal Fagopyrum esculentum. J. Plant Biol. 2017, 60, 189–198. [Google Scholar] [CrossRef]
- Lamb, R.S.; Irish, V.F. Functional divergence within the APETALA3/PISTILLATA floral homeotic gene lineages. Proc. Natl. Acad. Sci. USA 2003, 100, 6558–6563. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Transgenic Lines | Phenotype of Flower | No. Plants |
|---|---|---|
| 35S:: CyfaPI homozygous pi-1 transformants | strong complementation | 3(27.27%) |
| medium complementation | 2(18.18%) | |
| no complementation | 6(54.55%) | |
| 35S:: CyfaPI wild-type transformants | severe phenotype change | 3(42.86%) |
| weak phenotype change | 3(42.86%) | |
| no phenotype change | 1(14.28%) |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fei, Y.; Liu, Z.-X. Isolation and Characterization of the PISTILLATA Ortholog Gene from Cymbidium faberi Rolfe. Agronomy 2019, 9, 425. https://doi.org/10.3390/agronomy9080425
Fei Y, Liu Z-X. Isolation and Characterization of the PISTILLATA Ortholog Gene from Cymbidium faberi Rolfe. Agronomy. 2019; 9(8):425. https://doi.org/10.3390/agronomy9080425
Chicago/Turabian StyleFei, Yue, and Zhi-Xiong Liu. 2019. "Isolation and Characterization of the PISTILLATA Ortholog Gene from Cymbidium faberi Rolfe" Agronomy 9, no. 8: 425. https://doi.org/10.3390/agronomy9080425
APA StyleFei, Y., & Liu, Z.-X. (2019). Isolation and Characterization of the PISTILLATA Ortholog Gene from Cymbidium faberi Rolfe. Agronomy, 9(8), 425. https://doi.org/10.3390/agronomy9080425