Seed Viability of Heracleum mantegazzianum (Apiaceae) Is Quickly Reduced at Temperatures Prevailing in Biogas Plants


Abstract
:1. Introduction
- Seed survival will increase when the incubation temperature decreases,
- Seed survival will increase when seeds are more mature,
- Viable seeds will germinate after stratification, independently from previous incubation.
2. Materials and Methods
2.1. Seed Sampling
2.2. Seed Survival in Water Baths
2.3. Germination after Stratification
2.4. Statistical Evaluation
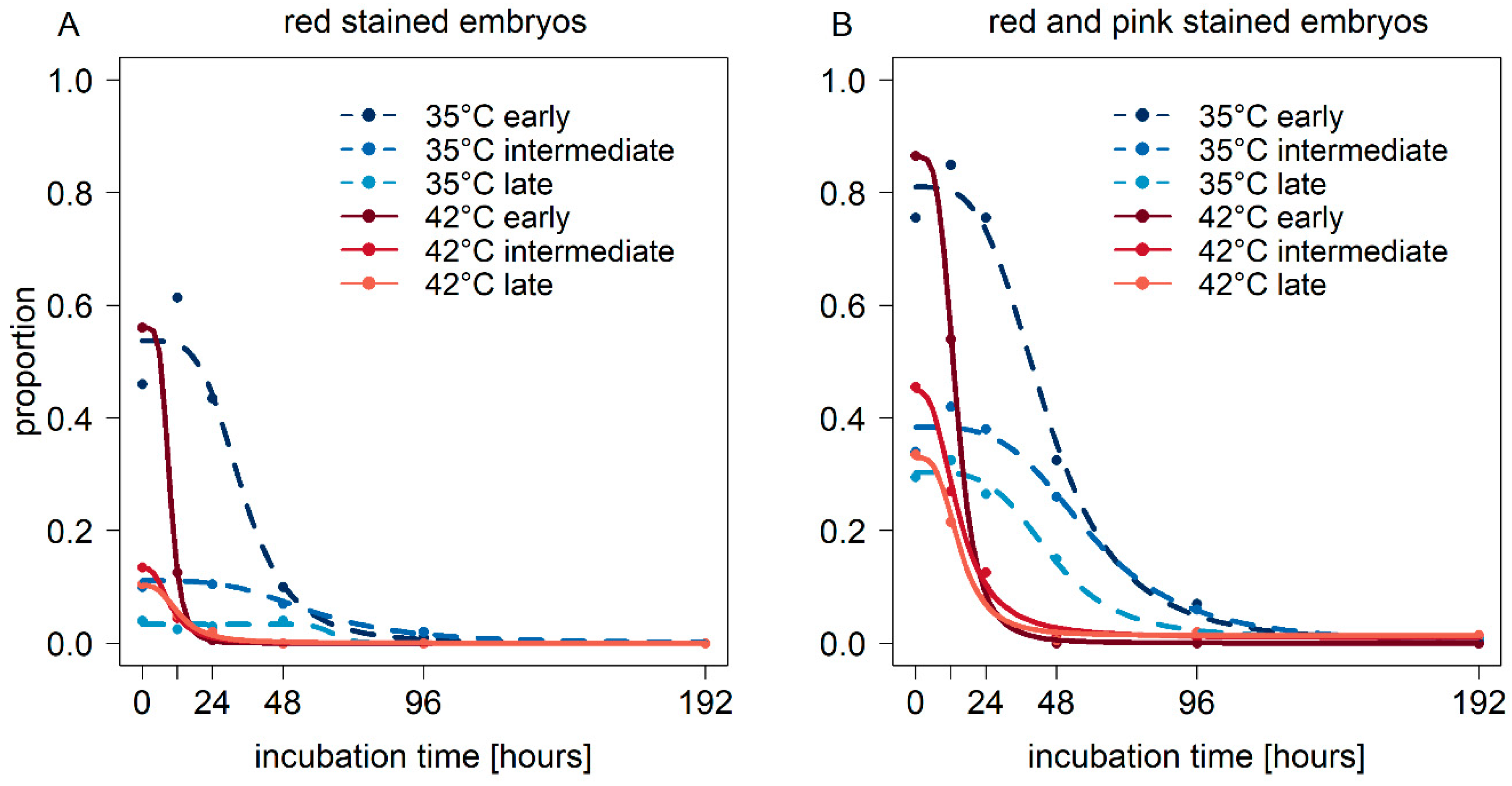
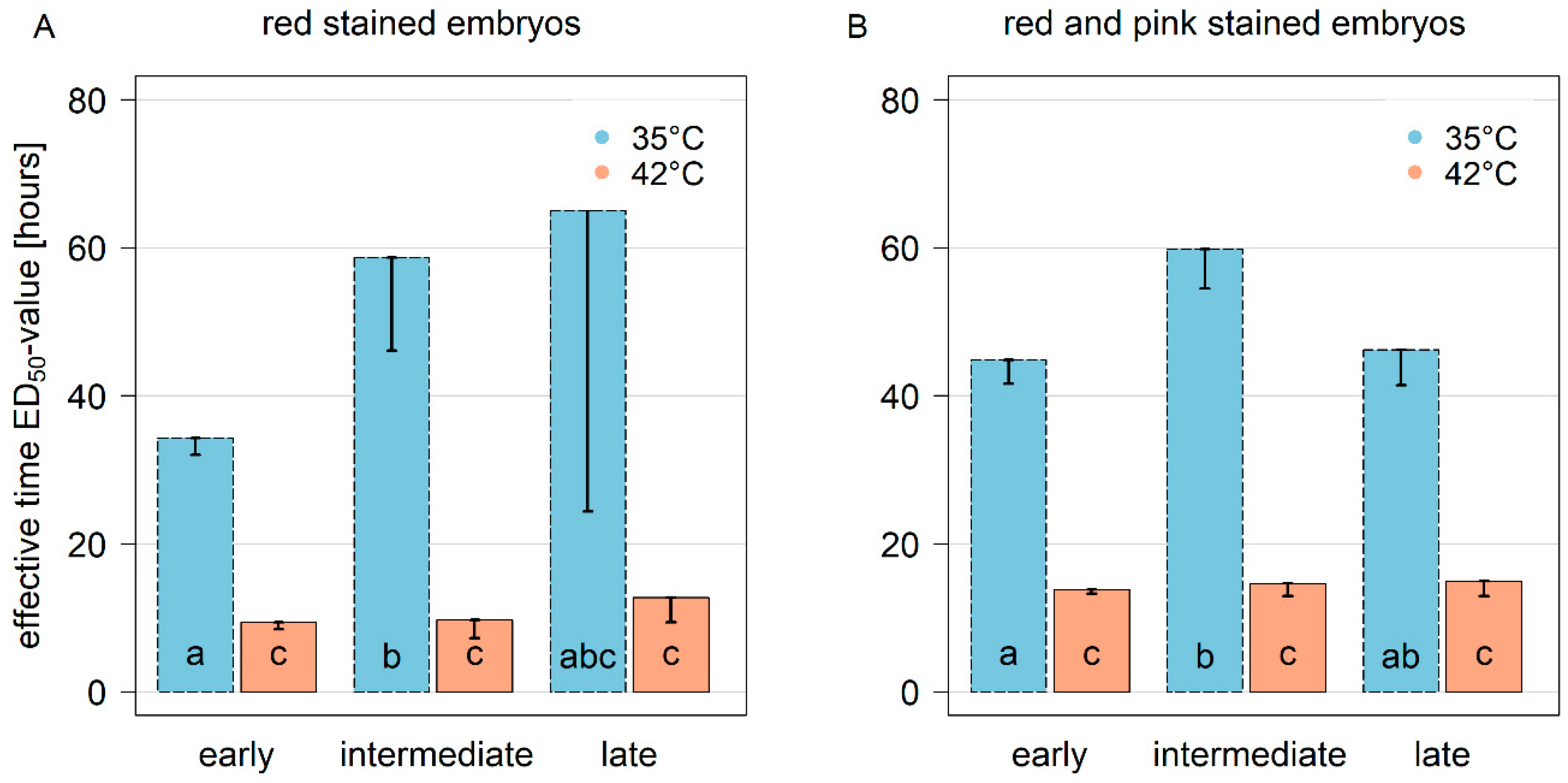
3. Results
3.1. Seed Viability after Incubation
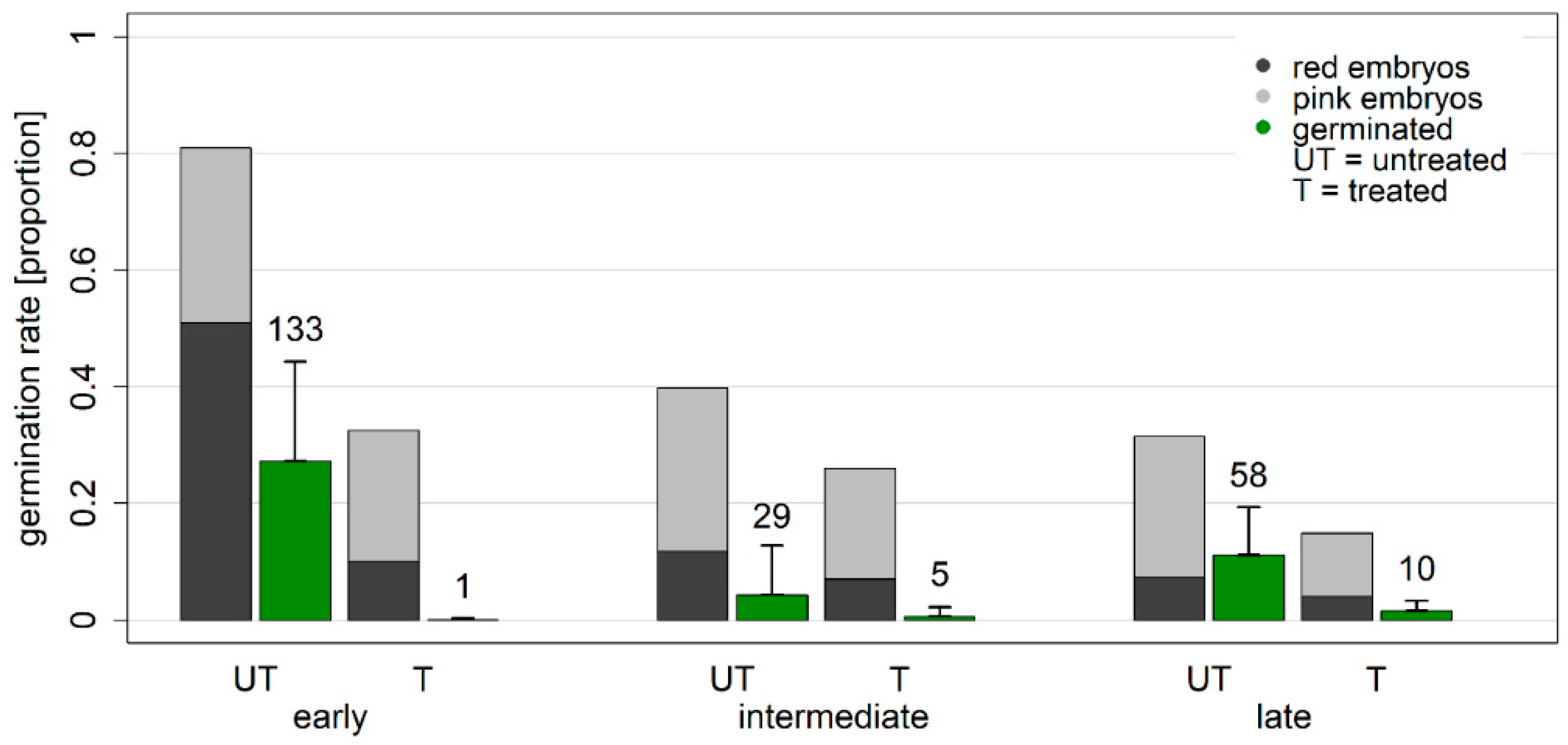
3.2. Germination after Stratification
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

References
- Nielsen, C.; Ravn, H.P.; Nentwig, W.; Wade, M. The Giant Hogweed Best Practice Manual: Guidelines for the Management and Control of An Invasive Weed in Europe; Forest & Landscape Denmark: Hoersholm, Denmark, 2005; 44p, ISBN 87-7903-211-7. [Google Scholar]
- Thiele, J.; Isermann, M.; Kollmann, J.; Otte, A. Impact scores of invasive plants are biased by disregard of environmental co-variation and non-linearity. Neobiota 2011, 10, 65–79. [Google Scholar] [CrossRef]
- Braun, M.; Schindler, S.; Essl, F. Distribution and management of invasive alien plant species in protected areas in Central Europe. J. Nat. Conserv. 2016, 33, 48–57. [Google Scholar] [CrossRef]
- Hejda, M.; Pyšek, P.; Jarošík, V. Impact of invasive plants on the species richness, diversity and composition of invaded communities. J. Ecol. 2009, 97, 393–403. [Google Scholar] [CrossRef]
- Pyšek, P.; Jarošík, V.; Müllerová, J.; Pergl, J.; Wild, J. Comparing the rate of invasion by Heracleum mantegazzianum at continental, regional, and local scales. Divers. Distrib. 2008, 14, 355–363. [Google Scholar] [CrossRef]
- Dalke, I.V.; Chadin, I.F.; Zakhozhiy, I.G.; Malyshev, R.V.; Maslova, S.P.; Tabalenkova, G.N.; Golovko, T.K. Traits of Heracleum sosnowskyi plants in monostand on invaded area. PLoS ONE 2015, 10, e0142833. [Google Scholar] [CrossRef] [PubMed]
- Thiele, J.; Otte, A. Invasion patterns of Heracleum mantegazzianum in Germany on the regional and landscape scales. J. Nat. Conserv. 2008, 16, 61–71. [Google Scholar] [CrossRef]
- Convention on Biological Diversity; United Nations: New York, NY, USA, 1992; 28 p.
- Camm, E.; Buck, H.W.L.; Mitchell, J.C. Phytophotodermatitis from Heracleum mantegazzianum. Contact Dermat. 1976, 2, 68–72. [Google Scholar] [CrossRef]
- Mack, R.N.; Simberloff, D.; Lonsdale, W.M.; Evans, H.; Clout, M.; Bazzaz, F.A. Biotic invasions: Causes, epidemiology, global consequences, and control. Ecol. Appl. 2000, 10, 689. [Google Scholar] [CrossRef]
- Nielsen, C.; Vanaga, I.; Treikale, O.; Priekule, I. Mechanical and Chemical Control of Heracleum mantegazzianum and Heracleum sosnowskyi. In Ecology and Management of Giant Hogweed (Heracleum Mantegazzianum); Pyšek, P., Cock, M.J.W., Nentwig, W., Ravn, H.P., Eds.; CABI: Wallingford, UK; Cambridge, MA, USA, 2007; pp. 226–239. ISBN 9781845932060. [Google Scholar]
- Otte, A.; Eckstein, R.L.; Thiele, J. Heracleum mantegazzianum in its primary distribution range of the Western Greater Caucasus. In Ecology and Management of Giant Hogweed (Heracleum mantegazzianum); Pyšek, P., Cock, M.J.W., Nentwig, W., Ravn, H.P., Eds.; CABI: Wallingford, UK; Cambridge, MA, USA, 2007; pp. 20–41. ISBN 9781845932060. [Google Scholar]
- Jahodová, S.; Fröberg, L.; Pyšek, P.; Geltman, D.; Trybush, S.; Karp, A. Taxonomy, identification, genetic relationships and distribution of large Heracleum species in Europe. In Ecology and Management of Giant Hogweed (Heracleum Mantegazzianum); Pyšek, P., Cock, M.J.W., Nentwig, W., Ravn, H.P., Eds.; CABI: Wallingford, UK; Cambridge, MA, USA, 2007; ISBN 9781845932060. [Google Scholar]
- European and Mediterranean Plant Protection Organization. Heracleum mantegazzianum, Heracleum sosnowskyi and Heracleum persicum. EPPO Bull. 2009, 39, 489–499. [Google Scholar] [CrossRef]
- Ochsmann, J. Heracleum mantegazzianum SOMMIER & LEVIER (Apiaceae) in Germany—Studies on biology, distribution, morphology and taxonomy (in German). Feddes Repert. 1996, 107, 557–595. [Google Scholar] [CrossRef]
- Tiley, G.E.D.; Dodd, F.S.; Wade, P.M. Heracleum mantegazzianum Sommier & Levier. J. Ecol. 1996, 84, 297. [Google Scholar] [CrossRef]
- Wade, M.; Darby, E.J.; Courtney, A.D.; Caffrey, J.M. Heracleum mantegazzianum: A problem for river managers in the Republic of Ireland and the United Kingdom. In Plant Invasions: Studies from North America and Europe; Brock, J.H., Wade, M., Pyšek, P., Green, D., Eds.; Backhuys Publishers: Leiden, The Netherlands, 1997; pp. 139–151. ISBN 9073348234. [Google Scholar]
- Pyšek, P. Heracleum mantegazzianum in the Czech Republic: Dynamics of spreading from the historical perspective. Folia Geobot. Phytotax. 1991, 26, 439–454. [Google Scholar] [CrossRef]
- Adolphi, K. Neophytic Cultivated Plants As Crop Refugees of the Rhineland; Berlin, Techn. Univ., Diss.; Galunder: Wiehl, Germany, 1995; ISBN 3931251012. (In German) [Google Scholar]
- Page, N.A.; Wall, R.E.; Darbyshire, S.J.; Mulligan, G.A. The Biology of Invasive Alien Plants in Canada. 4. Heracleum mantegazzianum Sommier & Levier. Can. J. Plant Sci. 2006, 86, 569–589. [Google Scholar] [CrossRef]
- Moravcová, L.; Pysek, P.; Krinke, L.; Pergl, J.; Perglová, I.; Thompson, K. Seed germination, dispersal and seed bank in Heracleum mantegazzianum. In Ecology and Management of Giant Hogweed (Heracleum mantegazzianum); Pyšek, P., Cock, M.J.W., Nentwig, W., Ravn, H.P., Eds.; CABI: Wallingford, UK; Cambridge, MA, USA, 2007; pp. 74–91. ISBN 9781845932060. [Google Scholar]
- Moravcová, L.; Gudzinskas, Z.; Pysek, P.; Pergl, J.; Perglová, I. Seed ecology of Heracleum mantegazzianum and H. sosnowskyi, two invasive species with different distributions in Europe. In Ecology and Management of Giant Hogweed (Heracleum mantegazzianum); Pyšek, P., Cock, M.J.W., Nentwig, W., Ravn, H.P., Eds.; CABI: Wallingford, UK; Cambridge, MA, USA, 2007; pp. 157–169. ISBN 9781845932060. [Google Scholar]
- Perglová, I.; Pergl, J.; Pyšek, P. Flowering phenology and reproductive effort of the invasive alien plant Heracleum mantegazzianum. Preslia 2006, 78, 265–285. [Google Scholar]
- Perglová, I.; Pergl, J.; Pysek, P. Reproductive ecology of Heracleum mantegazzianum. In Ecology and Management of Giant Hogweed (Heracleum mantegazzianum); Pyšek, P., Cock, M.J.W., Nentwig, W., Ravn, H.P., Eds.; CABI: Wallingford, UK; Cambridge, MA, USA, 2007; pp. 55–73. ISBN 9781845932060. [Google Scholar]
- Moravcová, L.; Pyšek, P.; Pergl, J.; Perglová, I.; Jarošík, V. Seasonal pattern of germination and seed longevity in the invasive species Heracleum mantegazzianum. Preslia 2006, 78, 287–301. [Google Scholar]
- Moravcová, L.; Pyšek, P.; Krinke, L.; Müllerová, J.; Perglová, I.; Pergl, J. Long-term survival in soil of seed of the invasive herbaceous plant Heracleum mantegazzianum. Preslia 2018, 90, 225–234. [Google Scholar] [CrossRef]
- Pyšek, P. Ecological aspects of invasion by Heracleum mantegazzianum in the Czech Republic. In Ecology and Management of Invasive Riverside Plants; Waal, L.C., Child, L.E., Wade, P.M., Brock, J.H., Eds.; Wiley: Chichester, UK, 1994; pp. 45–54. [Google Scholar]
- Vaverková, M.D.; Winkler, J.; Adamcová, D.; Radziemska, M.; Uldrijan, D.; Zloch, J. Municipal solid waste landfill—Vegetation succession in an area transformed by human impact. Ecol. Eng. 2019, 129, 109–114. [Google Scholar] [CrossRef]
- Andersen, U.V.; Calov, B. Long-term effects of sheep grazing on giant hogweed (Heracleum mantegazzianum). Hydrobiologia 1996, 340, 277–284. [Google Scholar] [CrossRef]
- Lundström, H. Giant hogweed, Heracleum mantegazzianum, a threat to the Swedish countryside. In Proceedings of the Weeds and Weed Control. 25th Swedish Weed Conference, Uppsala, Sweden, February 1984; Volume 1, Reports. pp. 191–200. [Google Scholar]
- Jodaugiene, D.; Marcinkeviciene, A.; Sinkeviciene, A. Control of Heracleum sosnowskyi in Lithuania. Julius Kühn Archiv 2018, 458, 275–280. [Google Scholar] [CrossRef]
- van Meerbeek, K.; Appels, L.; Dewil, R.; Calmeyn, A.; Lemmens, P.; Muys, B.; Hermy, M. Biomass of invasive plant species as a potential feedstock for bioenergy production. Biofuels Bioprod. Bioref. 2015, 9, 273–282. [Google Scholar] [CrossRef]
- Westerman, P.R.; Gerowitt, B. Weed seed survival during anaerobic digestion in biogas plants. Bot. Rev. 2013, 79, 281–316. [Google Scholar] [CrossRef]
- Dahlquist, R.M.; Prather, T.S.; Stapleton, J.J. Time and temperature requirements for weed seed thermal death. Weed Sci. 2007, 55, 619–625. [Google Scholar] [CrossRef]
- Hahn, J.; Parzych, D.; Schulz, J.; Westerman, P.R.; Gerowitt, B. Wildflower seeds in the biogas reactor: Screening the risk of survival of different species. Julius Kühn Archiv 2018, 458, 41–46. [Google Scholar] [CrossRef]
- Baier, U.; Warthmann, R.; Schleiss, K. Fermentation & Composting Plants As Hygiene Barriers (in German), Report, 2010, Zürich, 42p. Available online: https://www.biomassesuisse.ch/files/biomasse_temp/data/Das_bieten_wir/FE_Bericht_Hygienebarrieren_end3.pdf (accessed on 13 May 2019).
- Hahn, J.; Parzych, D.; Westerman, P.R.; Heiermann, M.; Gerowitt, B. The importance of temperature in the inactivation of seeds in biogas reactors (in German). Julius Kühn Archiv 2016, 452, 123–129. [Google Scholar] [CrossRef]
- Moore, R.P. ISTA Handbuch für die Tetrazoliumuntersuchung; 2. Aufl; Internationale Vereinigung für Saatgutprüfung: Zürich, Switzerland, 1985; ISBN 3906549259. [Google Scholar]
- Ritz, C.; Streibig, J.C. drc: Analysis of Dose-Response Curves. R Package Version 3.0-1. 2016. Available online: https://CRAN.R-project.org/package=drc (accessed on 18 May 2019).
- R Core Team. 2019. Available online: http://www.R-project.org/ (accessed on 18 May 2019).
- Ritz, C.; Baty, F.; Streibig, J.C.; Gerhard, D. Dose-response analysis using R. PLoS ONE 2015, 10, e0146021. [Google Scholar] [CrossRef]
- Hothorn, T.; Bretz, F.; Westfall, P. Simultaneus inference in general parametric models. Biom. J. 2008, 50, 346–363. [Google Scholar] [CrossRef]
- Hahn, J.; Westerman, P.R.; Heiermann, M.; Gerowitt, B. Survival of seeds from wild-flowering mixtures in the mesophilic, anaerobic digestion of the biogas process. Asp. Appl. Biol. 2015, 131, 187–192. [Google Scholar]
- Westerman, P.R.; Hildebrandt, F.; Gerowitt, B. Weed seed survival following ensiling and mesophilic anaerobic digestion in batch reactors. Weed Res. 2012, 52, 286–295. [Google Scholar] [CrossRef]
- Westerman, P.R.; Heiermann, M.; Pottberg, U.; Rodemann, B.; Gerowitt, B. Weed seed survival during mesophilic anaerobic digestion in biogas plants. Weed Res. 2012, 52, 307–316. [Google Scholar] [CrossRef]
- Egley, G.H. High-temperature effects on germination and survival of weed seeds in soil. Weed Sci. 1990, 38, 429–435. [Google Scholar] [CrossRef]
- Eckford, R.E.; Newman, J.C.; Li, X.; Watson, P.R. Thermophilic anaerobic digestion of cattle manure reduces seed viability for four weed species. Int. J. Agric. Biol. Eng. 2012, 5, 71–75. [Google Scholar]
- Strauß, G.; Kaplan, T.; Jacobi, T. Germination capability of seeds of different (genetically modified) crop plants depending on process parameters and residence time in a biogas plant (in German). J. Verbr. Lebensm. 2012, 7, 19–25. [Google Scholar] [CrossRef]
- Schrade, S.; Oechsner, H.; Perkun, C.; Claupein, W. Influence of the biogas process on the germination capability of seeds. Landtechnik 2003, 90–91. (In German) [Google Scholar]
- Thompson, A.J.; Jones, N.E.; Blair, A.M. The effect of temperature on viability of imbibed weed seeds. Ann. Appl. Biol. 1997, 130, 123–134. [Google Scholar] [CrossRef]
- Leonhardt, C.; Weinhappel, M.; Gansberger, M.; Brandstetter, A.; Schally, H.; Pfundtner, E. Investigations on the Risk of Propagation of Seedborne Diseases, Weeds and Sproutable Plant Parts with Fermentation End Products from Biogas Plants: Final Report on the Research Project 100296/2 (in German). Available online: https://www.dafne.at/prod/dafne_plus_common/attachment_download/9e9168bbb84a533b517ef792c752ca06/100296_Endbericht_biogas_dafne_letztfassung.pdf (accessed on 18 May 2019).
- Hay, F.; Probert, R.J. Seed maturity and the effects of different drying conditions on desiccation tolerance and seed longevity in foxglove (Digitalis purpurea L.). Ann. Bot. 1995, 76, 639–647. [Google Scholar] [CrossRef]
- Moravcová, L.; Perglová, I.; Pyšek, P.; Jarošík, V.; Pergl, J. Effects of fruit position on fruit mass and seed germination in the alien species Heracleum mantegazzianum (Apiaceae) and the implications for its invasion. Acta Oecol. 2005, 28, 1–10. [Google Scholar] [CrossRef]
- Oechsner, H.; Knödler, P.; Gerhards, R. Conditions for the Inactivation of Weed Seeds in the Biogas Process: Biogas Infotage Ulm. Available online: http://docplayer.org/75306345-Bedingungen-zur-inaktivierung-von-unkrautsamen-im-biogasprozess.html (accessed on 18 May 2019).


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Untreated | Early | Untreated | Intermediate | Untreated | Late | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Red | Pink | White | Rotten | n | Red | Pink | White | Rotten | n | Red | Pink | White | Rotten | n | ||||||
| 204 | 120 | 13 | 63 | 400 | 47 | 112 | 74 | 167 | 400 | 29 | 97 | 64 | 210 | 400 | ||||||
| 12 h | 35 °C | 122 | 47 | 2 | 28 | 199 | 12 h | 35 °C | 25 | 59 | 29 | 87 | 200 | 12 h | 35 °C | 5 | 60 | 28 | 107 | 200 |
| 42 °C | 25 | 83 | 57 | 35 | 200 | 42 °C | 9 | 45 | 59 | 88 | 201 | 42 °C | 10 | 33 | 54 | 103 | 200 | |||
| 1 d | 35 °C | 87 | 64 | 12 | 37 | 200 | 1 d | 35 °C | 21 | 55 | 29 | 95 | 200 | 1 d | 35 °C | 6 | 47 | 25 | 122 | 200 |
| 42 °C | 1 | 19 | 141 | 40 | 201 | 42 °C | 3 | 22 | 71 | 103 | 199 | 42 °C | 4 | 12 | 72 | 110 | 198 | |||
| 2 d | 35 °C | 20 | 45 | 94 | 41 | 200 | 2 d | 35 °C | 14 | 38 | 38 | 110 | 200 | 2 d | 35 °C | 8 | 22 | 57 | 114 | 201 |
| 42 °C | 0 | 0 | 158 | 42 | 200 | 42 °C | 0 | 3 | 91 | 106 | 200 | 42 °C | 0 | 1 | 98 | 101 | 200 | |||
| 4 d | 35 °C | 2 | 12 | 153 | 33 | 200 | 4 d | 35 °C | 4 | 8 | 84 | 104 | 200 | 4 d | 35 °C | 0 | 4 | 86 | 110 | 200 |
| 42 °C | 0 | 0 | 163 | 37 | 200 | 42 °C | 0 | 2 | 101 | 98 | 201 | 42 °C | 0 | 4 | 73 | 123 | 200 | |||
| 8 d | 35 °C | 0 | 0 | 164 | 36 | 200 | 8 d | 35 °C | 0 | 1 | 90 | 109 | 200 | 8 d | 35 °C | 0 | 2 | 83 | 114 | 199 |
| 42 °C | 0 | 0 | 161 | 39 | 200 | 42 °C | 0 | 3 | 87 | 110 | 200 | 42 °C | 0 | 3 | 74 | 123 | 200 | |||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tanke, A.; Müller, J.; de Mol, F. Seed Viability of Heracleum mantegazzianum (Apiaceae) Is Quickly Reduced at Temperatures Prevailing in Biogas Plants. Agronomy 2019, 9, 332. https://doi.org/10.3390/agronomy9060332
Tanke A, Müller J, de Mol F. Seed Viability of Heracleum mantegazzianum (Apiaceae) Is Quickly Reduced at Temperatures Prevailing in Biogas Plants. Agronomy. 2019; 9(6):332. https://doi.org/10.3390/agronomy9060332
Chicago/Turabian StyleTanke, Anja, Jürgen Müller, and Friederike de Mol. 2019. "Seed Viability of Heracleum mantegazzianum (Apiaceae) Is Quickly Reduced at Temperatures Prevailing in Biogas Plants" Agronomy 9, no. 6: 332. https://doi.org/10.3390/agronomy9060332
APA StyleTanke, A., Müller, J., & de Mol, F. (2019). Seed Viability of Heracleum mantegazzianum (Apiaceae) Is Quickly Reduced at Temperatures Prevailing in Biogas Plants. Agronomy, 9(6), 332. https://doi.org/10.3390/agronomy9060332